
The Bovhyaluronidase Azoximer (Longidaza®) Disrupts Candida albicans and Candida albicans-Bacterial Mixed Biofilms and Increases the Efficacy of Antifungals
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Strains and Growth Conditions
2.3. Minimum Inhibitory Concentrations (MICs)
2.4. Biofilms Staining and Quantification
2.5. The Anti-Biofilm Activity
2.6. Scanning Electron Microscopy
2.7. Statistical Analysis
3. Results
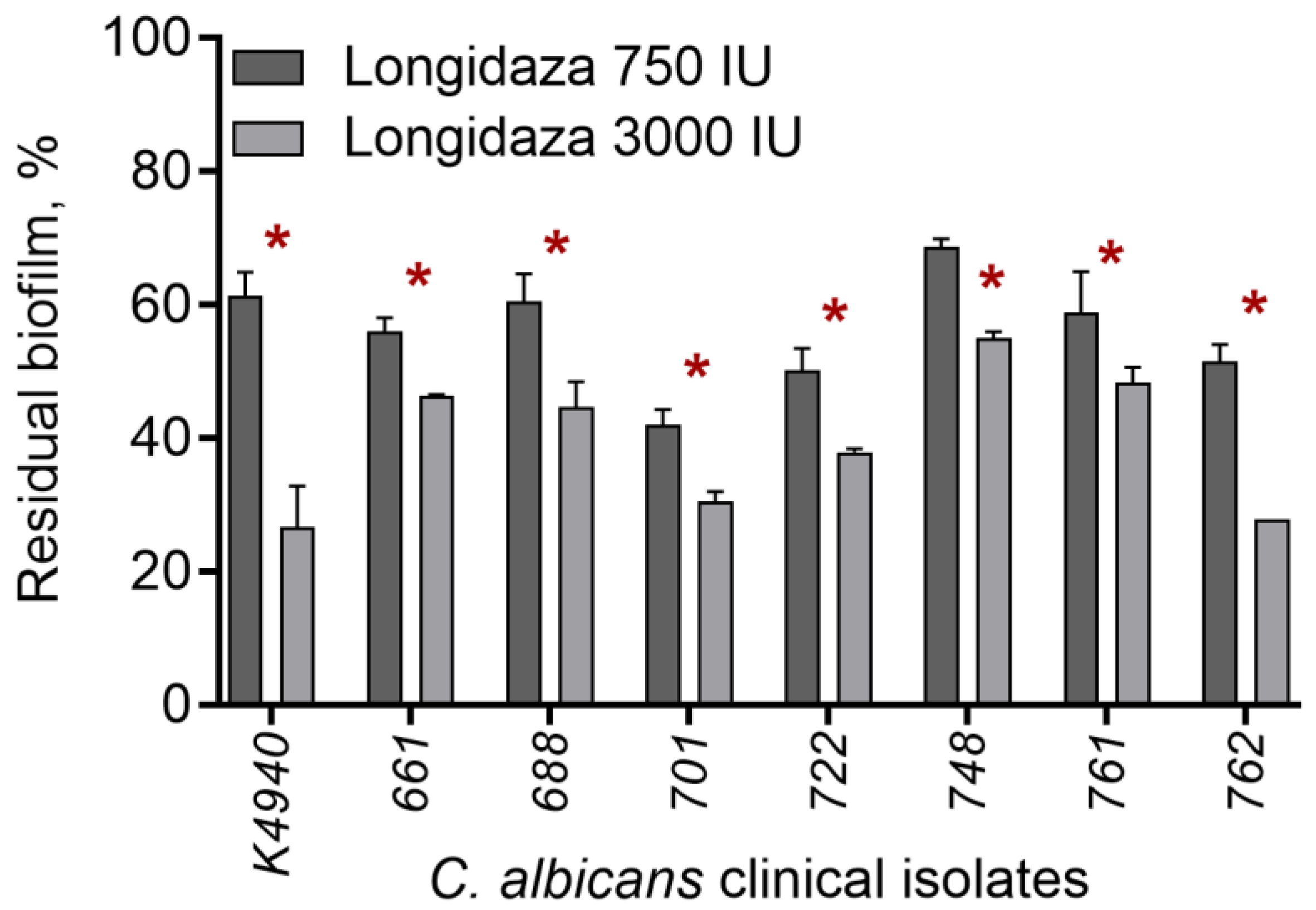
3.1. The Effect of Bovhyaluronidaze Azoximer (Longidaza®) on C. albicans Biofilms
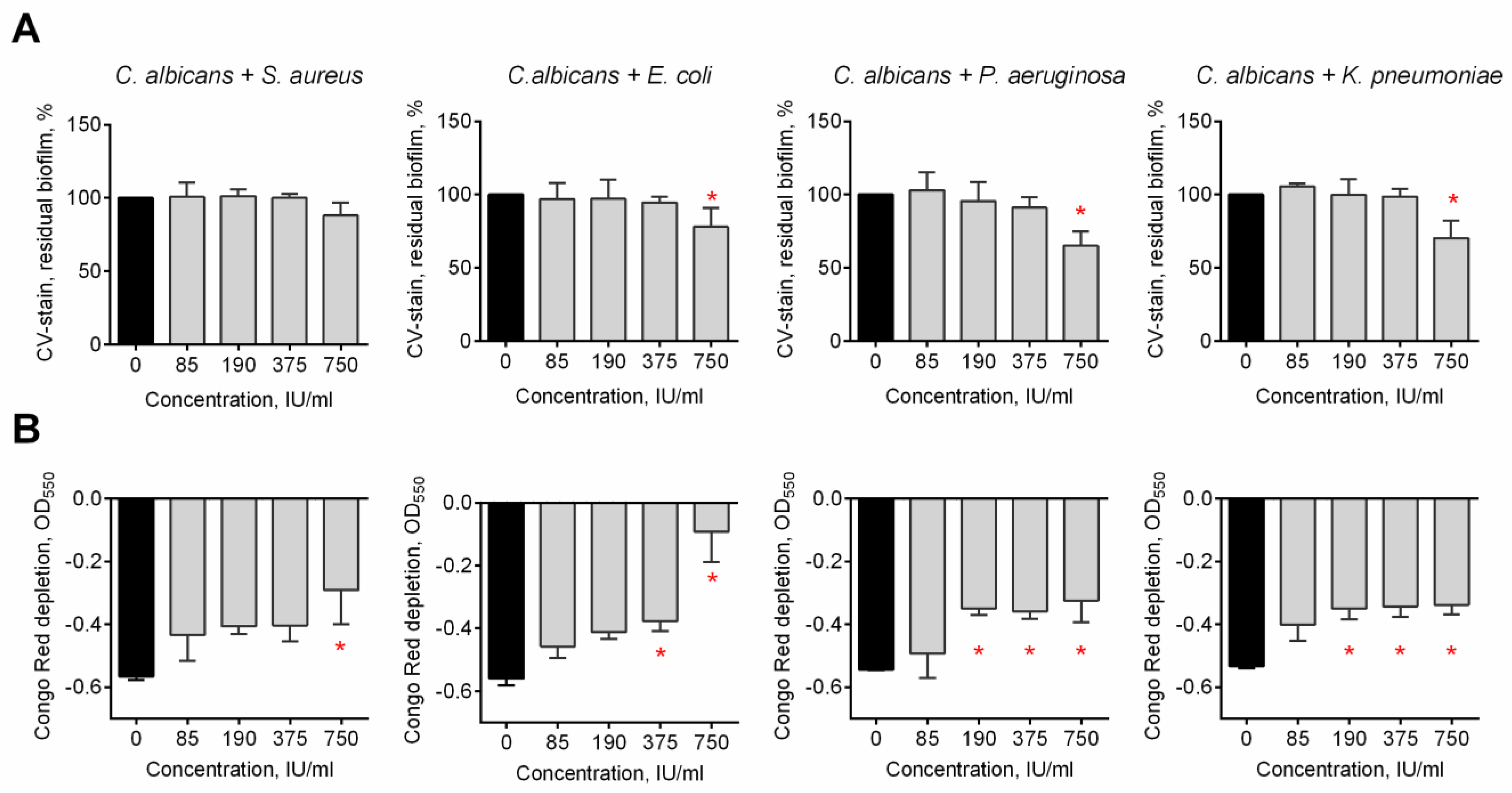
3.2. The Effect of Bovhyaluronidaze Azoximer (Longidaza®) on C. albicans–Bacterial Mixed Biofilms
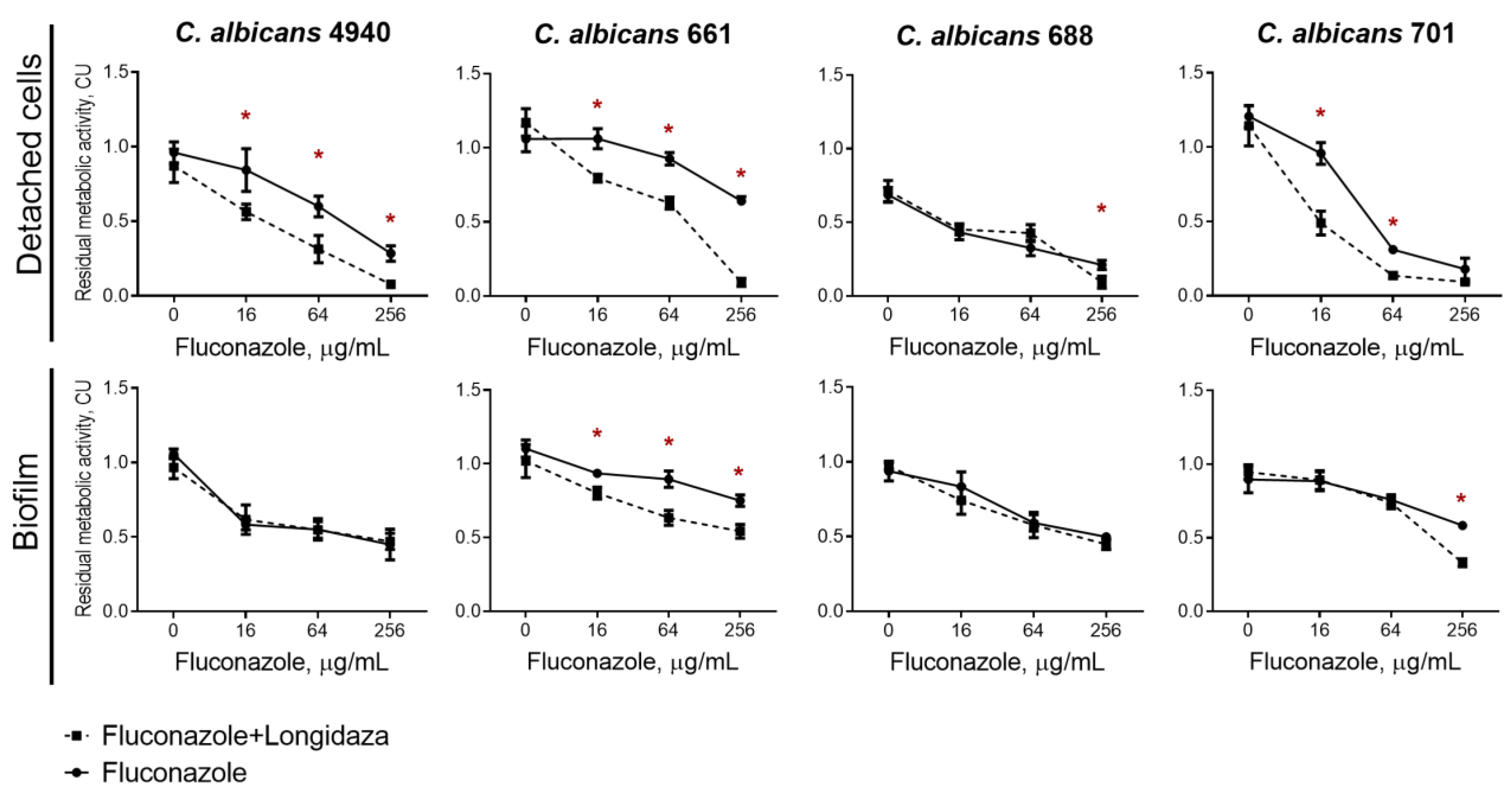
3.3. The effect of Longidaza® on the Susceptibility of Biofilms-Embedded C. albicans to Antifungals
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single- and multi-species Candida albicans biofilms. Nat. Rev. Microbiol. 2018, 16, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, C.G.; Vila, T.; Romo, J.A.; Montelongo-Jauregui, D.; Wall, G.; Ramasubramanian, A.; Lopez-Ribot, J.L. The Candida albicans Biofilm Matrix: Composition, Structure and Function. J. Fungi 2017, 3, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, K.F.; Zarnowski, R.; Andes, D.R. Fungal Super Glue: The Biofilm Matrix and Its Composition, Assembly, and Functions. PLoS Pathog. 2016, 12, e1005828. [Google Scholar] [CrossRef] [Green Version]
- Corte, L.; Roscini, L.; Colabella, C.; Tascini, C.; Leonildi, A.; Sozio, E.; Menichetti, F.; Merelli, M.; Scarparo, C.; Meyer, W.; et al. Exploring ecological modelling to investigate factors governing the colonization success in nosocomial environment of Candida albicans and other pathogenic yeasts. Sci. Rep. 2016, 6, 26860. [Google Scholar] [CrossRef]
- Cavalheiro, M.; Teixeira, M.C. Candida Biofilms: Threats, Challenges, and Promising Strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Al-Fattani, M.A.; Douglas, L.J. Biofilm matrix of Candida albicans and Candida tropicalis: Chemical composition and role in drug resistance. J. Med. Microbiol. 2006, 55, 999–1008. [Google Scholar] [CrossRef]
- Tobudic, S.; Kratzer, C.; Lassnigg, A.; Presterl, E. Antifungal susceptibility of Candida albicans in biofilms. Mycoses 2012, 55, 199–204. [Google Scholar] [CrossRef]
- Taff, H.T.; Mitchell, K.F.; Edward, J.A.; Andes, D.R. Mechanisms of Candida biofilm drug resistance. Future Microbiol. 2013, 8, 1325–1337. [Google Scholar] [CrossRef] [Green Version]
- Zarnowski, R.; Westler, W.M.; Lacmbouh, G.A.; Marita, J.M.; Bothe, J.R.; Bernhardt, J.; Sahraoui, A.L.H.; Fontaine, J.; Sanchez, H.; Hatfield, R.D.; et al. Novel Entries in a Fungal Biofilm Matrix Encyclopedia. Mbio 2014, 5, e01333-14. [Google Scholar] [CrossRef]
- Wolcott, R.; Costerton, J.W.; Raoult, D.; Cutler, S.J. The polymicrobial nature of biofilm infection. Clin. Microbiol. Infect. 2013, 19, 107–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, B.M.; Jabra-Rizk, M.A.; O’May, G.A.; Costerton, J.W.; Shirtliff, M.E. Polymicrobial Interactions: Impact on Pathogenesis and Human Disease. Clin. Microbiol. Rev. 2012, 25, 193–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peleg, A.Y.; Hogan, D.A.; Mylonakis, E. Medically important bacterial-fungal interactions. Nat. Rev. Microbiol. 2010, 8, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Carolus, H.; Van Dyck, K.; Van Dijck, P. Candida albicans and Staphylococcus species: A threatening twosome. Front. Microbiol. 2019, 10, 2162. [Google Scholar] [CrossRef] [PubMed]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans biofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, A.K.; Hogan, D.A. Candida albicans: Molecular interactions with Pseudomonas aeruginosa and Staphylococcus aureus. Fungal Biol. Rev. 2014, 28, 85–96. [Google Scholar] [CrossRef]
- Fox, E.P.; Cowley, E.S.; Nobile, C.J.; Hartooni, N.; Newman, D.K.; Johnson, A.D. Anaerobic Bacteria Grow within Candida albicans Biofilms and Induce Biofilm Formation in Suspension Cultures. Curr. Biol. 2014, 24, 2411–2416. [Google Scholar] [CrossRef] [Green Version]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting microbial biofilms: Current and prospective therapeutic strategies. Nat. Rev. Microbiol. 2017, 15, 740–755. [Google Scholar] [CrossRef]
- Hacioglu, M.; Oyardi, O.; Bozkurt-Guzel, C.; Savage, P.B. Antibiofilm activities of ceragenins and antimicrobial peptides against fungal-bacterial mono and multispecies biofilms. J. Antibiot. 2020, 73, 455–462. [Google Scholar] [CrossRef]
- Weiland-Bräuer, N.; Malek, I.; Schmitz, R.A. Metagenomic quorum quenching enzymes affect biofilm formation of Candida albicans and Staphylococcus epidermidis. PLoS ONE 2019, 14, e0211366. [Google Scholar] [CrossRef]
- Sharafutdinov, I.S.; Ozhegov, G.D.; Sabirova, A.E.; Novikova, V.V.; Lisovskaya, S.A.; Khabibrakhmanova, A.M.; Kurbangalieva, A.R.; Bogachev, M.I.; Kayumov, A.R. Increasing Susceptibility of Drug-Resistant. Molecules 2020, 25, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thallinger, B.; Prasetyo, E.N.; Nyanhongo, G.S.; Guebitz, G.M. Antimicrobial enzymes: An emerging strategy to fight microbes and microbial biofilms. Biotechnol. J. 2013, 8, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Nahar, S.; Mizan, M.F.R.; Ha, A.J.W.; Ha, S.D. Advances and Future Prospects of Enzyme-Based Biofilm Prevention Approaches in the Food Industry. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1484–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donelli, G.; Francolini, I.; Romoli, D.; Guaglianone, E.; Piozzi, A.; Ragunath, C.; Kaplan, J.B. Synergistic activity of dispersin B and cefamandole nafate in inhibition of staphylococcal biofilm growth on polyurethanes. Antimicrob. Agents Chemother. 2007, 51, 2733–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, J.B. Therapeutic potential of biofilm-dispersing enzymes. Int. J. Artif. Organs 2009, 32, 545–554. [Google Scholar] [CrossRef]
- Elchinger, P.H.; Delattre, C.; Faure, S.; Roy, O.; Badel, S.; Bernardi, T.; Taillefumier, C.; Michaud, P. Immobilization of proteases on chitosan for the development of films with anti-biofilm properties. Int. J. Biol. Macromol. 2015, 72, 1063–1068. [Google Scholar] [CrossRef]
- Baidamshina, D.R.; Trizna, E.Y.; Holyavka, M.G.; Bogachev, M.I.; Artyukhov, V.G.; Akhatova, F.S.; Rozhina, E.V.; Fakhrullin, R.F.; Kayumov, A.R. Targeting microbial biofilms using Ficin, a nonspecific plant protease. Sci. Rep. 2017, 7, 46068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baidamshina, D.R.; Koroleva, V.A.; Trizna, E.Y.; Pankova, S.M.; Agafonova, M.N.; Chirkova, M.N.; Vasileva, O.S.; Akhmetov, N.; Shubina, V.V.; Porfiryev, A.G.; et al. Anti-biofilm and wound-healing activity of chitosan-immobilized Ficin. Int. J. Biol. Macromol. 2020, 164, 4205–4217. [Google Scholar] [CrossRef]
- Baidamshina, D.R.; Koroleva, V.A.; Olshannikova, S.S.; Trizna, E.Y.; Bogachev, M.I.; Artyukhov, V.G.; Holyavka, M.G.; Kayumov, A.R. Biochemical Properties and Anti-Biofilm Activity of Chitosan-Immobilized Papain. Mar. Drugs 2021, 19, 197. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.G.; Nigam, Y.; Sawyer, J.; Mack, D.; Pritchard, D.I. Lucilia sericata Chymotrypsin Disrupts Protein Adhesin-Mediated Staphylococcal Biofilm Formation. Appl. Environ. Microbiol. 2013, 79, 1393–1395. [Google Scholar] [CrossRef]
- Selan, L.; Berlutti, F.; Passariello, C.; Comodiballanti, M.R.; Thaller, M.C. Proteolytic-enzymes—A new treatment strategy for prosthetic infections. Antimicrob. Agents Chemother. 1993, 37, 2618–2621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panariello, B.H.D.; Klein, M.I.; Alves, F.; Pavarina, A.C. DNase increases the efficacy of antimicrobial photodynamic therapy on Candida albicans biofilms. Photodiagn. Photodyn. Ther. 2019, 27, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Sachivkina, N.; Lenchenko, E.; Blumenkrants, D.; Ibragimova, A.; Bazarkina, O. Effects of farnesol and lyticase on the formation of Candida albicans biofilm. Vet. World 2020, 13, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, A.; Natalja, P.; Sergej, M. Stable Cosmetic Composition Based on Immobilized Hyaluronidase and Preparation Method Thereof. Russian Patent RU2604160C1, 12 October 2015. [Google Scholar]
- Klasen, H.J. A review on the nonoperative removal of necrotic tissue from burn wounds. Burns 2000, 26, 207–222. [Google Scholar] [CrossRef]
- McCarty, S.M.; Cochrane, C.A.; Clegg, P.D.; Percival, S.L. The role of endogenous and exogenous enzymes in chronic wounds: A focus on the implications of aberrant levels of both host and bacterial proteases in wound healing. Wound Repair Regen. 2012, 20, 125–136. [Google Scholar] [CrossRef]
- Liese, A.; Hilterhaus, L. Evaluation of immobilized enzymes for industrial applications. Chem. Soc. Rev. 2013, 42, 6236–6249. [Google Scholar] [CrossRef]
- Troshina, N.; Dolgushin, I.; Dolgushina, V.; Bakhareva, L.; Kurnosenko, I.; Nadvikova, T. Longidaza Influence on microbial landscape of the cervical canal and the uterine cavity in the treatment of women with chronic nonspecific endometritis and uterine myoma. Zhurnal Mikrobiol. Epidemiol. Immunobiol. 2015, 4, 71–74. [Google Scholar]
- Abramova, S.N.; Lazareva, G.A. Pharmacotherapy exacerbations of chronic inflammatory conditions of female genital sphere using to Gepon and Longidaza. Res. Results Pharmacol. 2016, 2, 55–67. [Google Scholar] [CrossRef]
- Kulchavenya, E.; Shvetsova, O.; Breusov, A. Rationale of use and effectiveness of Longidaza in patients with chronic prostatitis. Urologiia 2018, 4, 64–71. [Google Scholar] [CrossRef]
- Kotov, S.; Bolotov, A.; Belomyttsev, S.; SN, D.; Vorobyova, A. Multicenter randomized study of bovhyalu-ronidase azoximer (Longidaza) in men after transuretral resection of the prostate. Urologiia 2021, 3, 61–69. [Google Scholar] [CrossRef]
- Kayumov, A.R.; Khakimullina, E.N.; Sharafutdinov, I.S.; Trizna, E.Y.; Latypova, L.Z.; Lien, H.T.; Margulis, A.B.; Bogachev, M.I.; Kurbangalieva, A.R. Inhibition of biofilm formation in Bacillus subtilis by new halogenated furanones. J. Antibiot. 2015, 68, 297–301. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A.; Kolter, R. Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: A genetic analysis. Mol. Microbiol. 1998, 28, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Lafi, R.; Montasser, I.; Hafiane, A. Adsorption of congo red dye from aqueous solutions by prepared activated carbon with oxygen-containing functional groups and its regeneration. Adsorpt. Sci. Technol. 2019, 37, 160–181. [Google Scholar] [CrossRef] [Green Version]
- Grela, E.; Kozlowska, J.; Grabowiecka, A. Current methodology of MTT assay in bacteria—A review. Acta Histochem. 2018, 120, 303–311. [Google Scholar] [CrossRef]
- Loiselle, M.; Anderson, K.W. The use of cellulase in inhibiting biofilm formation from organisms commonly found on medical implants. Biofouling 2003, 19, 77–85. [Google Scholar] [CrossRef]
- Rodrigues, M.E.; Gomes, F.; Rodrigues, C.F. Candida spp./Bacteria Mixed Biofilms. J. Fungi 2019, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Holá, V.; Ruzicka, F.; Horka, M. Microbial diversity in biofilm infections of the urinary tract with the use of sonication techniques. FEMS Immunol. Med. Microbiol. 2010, 59, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Gaston, J.R.; Johnson, A.O.; Bair, K.L.; White, A.N.; Armbruster, C.E. Polymicrobial interactions in the urinary tract: Is the enemy of my enemy my friend? Infect. Immun. 2021, 89, e00652-20. [Google Scholar] [CrossRef]
- Daboor, S.M.; Rohde, J.R.; Cheng, Z. Disruption of the extracellular polymeric network of Pseudomonas aeruginosa biofilms by alginate lyase enhances pathogen eradication by antibiotics. J. Cyst. Fibros. 2021, 20, 264–270. [Google Scholar] [CrossRef]
- Bamford, N.C.; Le Mauff, F.; Subramanian, A.S.; Yip, P.; Millán, C.; Zhang, Y.; Zacharias, C.; Forman, A.; Nitz, M.; Codée, J.D.C.; et al. Ega3 from the fungal pathogen. J. Biol. Chem. 2019, 294, 13833–13849. [Google Scholar] [CrossRef] [Green Version]
- Fleming, D.; Chahin, L.; Rumbaugh, K. Glycoside Hydrolases Degrade Polymicrobial Bacterial Biofilms in Wounds. Antimicrob. Agents Chemother. 2017, 61, e01998-16. [Google Scholar] [CrossRef] [PubMed]
- Matias, R.R.; Sepúlveda, A.M.G.; Batista, B.N.; de Lucena, J.M.V.M.; Albuquerque, P.M. Degradation of Staphylococcus aureus Biofilm Using Hydrolytic Enzymes Produced by Amazonian Endophytic Fungi. Appl. Biochem. Biotechnol. 2021, 193, 2145–2161. [Google Scholar] [CrossRef]
- Lin, Y.; Zhou, X.; Li, Y. Strategies for Streptococcus mutans biofilm dispersal through extracellular polymeric substances disruption. Mol. Oral Microbiol. 2022, 37, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, D.; Nag, M.; Dey, A.; Sarkar, T.; Ray, R.R.; Rebezov, M.; Shariati, M.A.; Thiruvengadam, M.; Simal-Gandara, J. Immobilized enzymes as potent antibiofilm agent. Biotechnol. Prog. 2022, 38, e3281. [Google Scholar] [CrossRef] [PubMed]
- Trizna, E.; Bogachev, M.I.; Kayumov, A. Degrading of the Pseudomonas aeruginosa Biofilm by Extracellular Levanase SacC from Bacillus subtilis. Bionanoscience 2019, 9, 48–52. [Google Scholar] [CrossRef]
- Kim, H.E.; Dhall, A.; Liu, Y.; Bawazir, M.; Koo, H.; Hwang, G. Intervening in Symbiotic Cross-Kingdom Biofilm Interactions: A Binding Mechanism-Based Nonmicrobicidal Approach. mBio 2021, 12, e00651-21. [Google Scholar] [CrossRef] [PubMed]
- Farisa Banu, S.; Thamotharan, S.; Gowrishankar, S.; Karutha Pandian, S.; Nithyanand, P. Marine bacterial DNase curtails virulence and disrupts biofilms of Candida albicans and non-albicans Candida species. Biofouling 2019, 35, 975–985. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Source | MIC, µg/mL |
|---|---|---|
| C. albicans K4940 | Buccal swab | 64 |
| C. albicans 661 | Pharynx, mucosa of the tongue | 1024 |
| C. albicans 688 | Mucosa of the pharynx | 1024 |
| C. albicans 701 | Mucosa of tonsils | 1024 |
| C. albicans 722 | Mucosa of the pharynx | 1024 |
| C. albicans 748 | Mucosa of the pharynx | 64 |
| C. albicans 761 | Vaginal swab, cervical canal | 1024 |
| C. albicans 762 | Mucosa of the urethra | 1024 |
| Isolate | Total Biomass of the Biofilm, OD570 | Residual Biomass of the Biofilm, OD490 | α-Polysacch, Relative Units | β-Polysacch, Relative UNits | Proteins, Relative Units |
|---|---|---|---|---|---|
| C. albicans K4940 | 0.11 | 0.07 | 22.80 | 22.80 | 7.01 |
| C. albicans 661 | 0.12 | 0.07 | 19.13 | 16.64 | 11.65 |
| C. albicans 688 | 0.10 | 0.07 | 37.48 | 40.37 | 11.53 |
| C. albicans 701 | 0.15 | 0.06 | 17.90 | 9.94 | 7.95 |
| C. albicans 722 | 0.13 | 0.06 | 19.36 | 18.58 | 4.65 |
| C. albicans 748 | 0.09 | 0.06 | 38.35 | 41.84 | 27.89 |
| C. albicans 761 | 0.11 | 0.07 | 25.99 | 23.20 | 21.35 |
| C. albicans 762 | 0.13 | 0.07 | 12.38 | 13.15 | 6.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gatina, A.; Trizna, E.; Kolesnikova, A.; Baidamshina, D.; Gorshkova, A.; Drucker, V.; Bogachev, M.; Kayumov, A. The Bovhyaluronidase Azoximer (Longidaza®) Disrupts Candida albicans and Candida albicans-Bacterial Mixed Biofilms and Increases the Efficacy of Antifungals. Medicina 2022, 58, 1710. https://doi.org/10.3390/medicina58121710
Gatina A, Trizna E, Kolesnikova A, Baidamshina D, Gorshkova A, Drucker V, Bogachev M, Kayumov A. The Bovhyaluronidase Azoximer (Longidaza®) Disrupts Candida albicans and Candida albicans-Bacterial Mixed Biofilms and Increases the Efficacy of Antifungals. Medicina. 2022; 58(12):1710. https://doi.org/10.3390/medicina58121710
Chicago/Turabian StyleGatina, Alina, Elena Trizna, Alena Kolesnikova, Diana Baidamshina, Anna Gorshkova, Valentin Drucker, Mikhail Bogachev, and Airat Kayumov. 2022. "The Bovhyaluronidase Azoximer (Longidaza®) Disrupts Candida albicans and Candida albicans-Bacterial Mixed Biofilms and Increases the Efficacy of Antifungals" Medicina 58, no. 12: 1710. https://doi.org/10.3390/medicina58121710