
Staphylococcus epidermidis Pathogenesis: Interplay of icaADBC Operon and MSCRAMMs in Biofilm Formation of Isolates from Pediatric Bacteremia in Peshawar, Pakistan
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Identification
2.2. Phenotypic Analysis
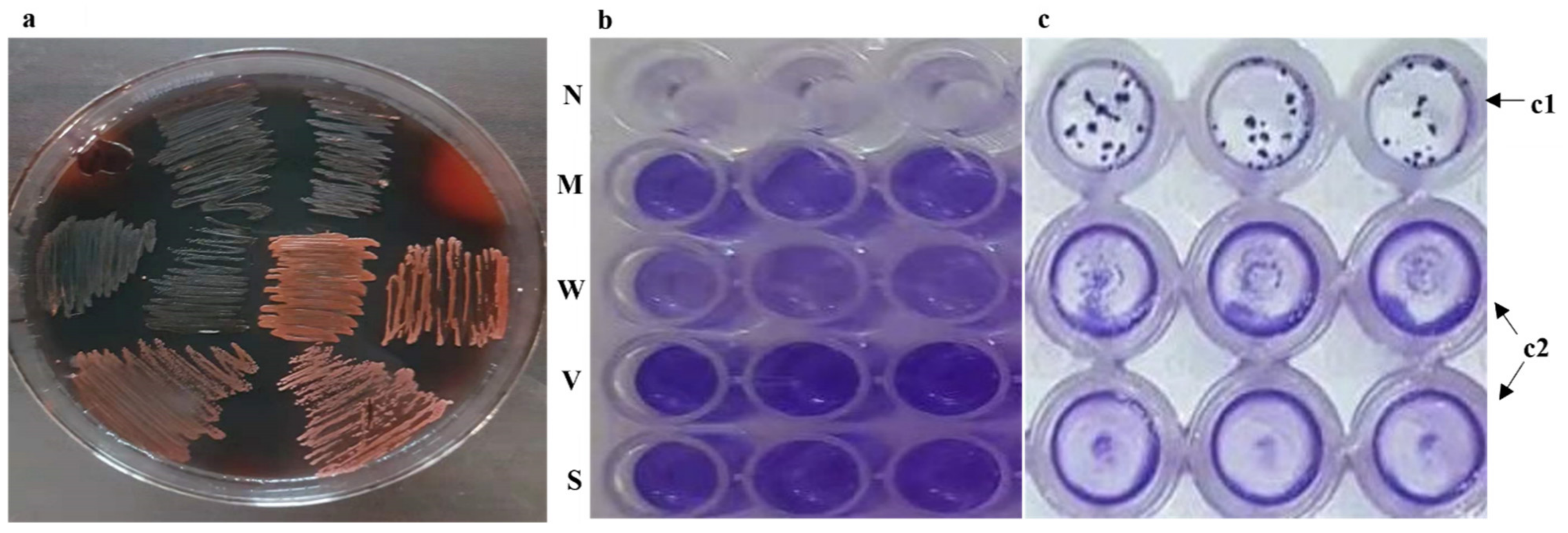
2.2.1. Congo Red Assay for icaADBC Operon Activity
2.2.2. Biofilm-Formation Assay under Plasma Stress
2.3. Detection of icaADBC Operon, MSCRAMMs, and IS256
2.4. Statistical Analysis
3. Results
3.1. Settings and Clinical Isolates
3.2. Screening and Presence of Complete, Missing, and Defective icaADBC Operon
3.3. Activity of Complete, Missing, and Defective icaADBC Operon
3.4. Distribution of Genes Encoding MSCRAMMs
3.5. Biofilm Formation Associated with the icaADBC Operon and MSCRAMMs
3.6. Screening for IS256 and Association with Biofilm Formation and icaADBC Operon
3.7. Association of icaADBC Operon and MSCRAMMs with Biofilm Mass Density
3.8. Appearance of Biofilm Morphotypes (BM) and Association with icaADBC Operon and MSCRAMMs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michels, R.; Last, K.; Becker, S.L.; Papan, C. Update on coagulase-negative Staphylococci—What the clinician should know. Microorganisms 2021, 9, 830. [Google Scholar] [CrossRef] [PubMed]
- Hogan, S.; Stevens, N.; Humphreys, H.; O’Gara, J.; O’Neill, E. Current and Future Approaches to the Prevention and Treatment of Staphylococcal Medical Device-Related Infections. Curr. Pharm. Des. 2014, 21, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Hira, V.; Sluijter, M.; Goessens, W.H.F.; Ott, A.; de Groot, R.; Hermans, P.W.M.; Kornelisse, R.F. Coagulase-Negative Staphylococcal Skin Carriage among Neonatal Intensive Care Unit Personnel: From Population to Infection. J. Clin. Microbiol. 2010, 48, 3876–3881. [Google Scholar] [CrossRef] [Green Version]
- Gerke, C.; Kraft, A.; Süßmuth, R.; Schweitzer, O.; Götz, F. Characterization of the N-Acetylglucosaminyltransferase Activity Involved in the Biosynthesis of the Staphylococcus epidermidis Polysaccharide Intercellular Adhesin. J. Biol. Chem. 1998, 273, 18586–18593. [Google Scholar] [CrossRef] [Green Version]
- Valle, J.; Vergara, M.; Merino, N.; Penades, R.; Lasa, I. B regulates IS256-mdiated Staphylococcus aureus biofilm phenotypic variation. J. Bacteriol. 2007, 189, 2886–2896. [Google Scholar] [CrossRef] [Green Version]
- Ziebuhr, W.; Krimmer, V.; Rachid, S.; Lossner, I.; Gotz, F.; Hacker, J. A novel mechanism of phase variation of virulence in Staphylococcus epidermidis: Evidence for control of the polysaccharide intercellular adhesin synthesis by alternating insertion and excision of the insertion sequence element IS256. Mol. Microbiol. 1999, 32, 345–3566. [Google Scholar] [CrossRef]
- Henning, S.; Wai, S.N.; Zeibuhr, W. Spontaneous switch to PIA-independent biofilm formation in an Ica-positive Staphylococcus epidermidis isolate. Int. J. Med. Microbiol. 2007, 297, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Li, H.; Li, M.; Vuong, C.; Otto, M.; Wen, Y.; Gao, Q. Bacterial insertion sequence IS256 as a potential molecular marker to discriminate invasive strains from commensal strains of Staphylococcus epidermidis. J. Hosp. Infect. 2005, 61, 342–348. [Google Scholar] [CrossRef]
- Conlon, K.M.; Humphreys, H.; O’Gara, J.P. icaR encodes a transcriptional repressor involved in environmental regulation of ica operon expression and biofilm formation in Staphylococcus epidermidis. J. Bacteriol. 2002, 184, 4400–4408. [Google Scholar] [CrossRef] [Green Version]
- Heilmann, C.; Schweitzer, O.; Gerke, C.; Vanittanakom, N.; Mack, D.; Götz, F. Molecular basis of intercellular adhesion in the biofilm-forming Staphylococcus epidermidis. Mol. Microbiol. 1996, 20, 1083–1091. [Google Scholar] [CrossRef]
- Mack, D.; Nedelmann, M.; Krokotsch, A.; Schwarzkopf, A.; Heesemann, J.; Laufs, R. Characterization of transposon mutants of biofilm-producing Staphylococcus epidermidis impaired in the accumulative phase of biofilm production: Genetic identification of a hexosamine-containing polysaccharide intercellular adhesin. Infect. Immun. 1994, 62, 3244–3253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christner, M.; Franke, G.C.; Schommer, N.N.; Wendt, U.; Wegert, K.; Pehle, P.; Kroll, G.; Schulze, C.; Buck, F.; Mack, D. The giant extracellular matrix-binding protein of Staphylococcus epidermidis mediates biofilm accumulation and attachment to fibronectin. Mol. Microbiol. 2010, 75, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Schommer, N.N.; Christner, M.; Hentschke, M.; Ruckdeschel, K.; Aepfelbacher, M.; Rohde, H. Staphylococcus epidermidis Uses Distinct Mechanisms of Biofilm Formation To Interfere with Phagocytosis and Activation of Mouse Macrophage-Like Cells 774A.1. Infect. Immun. 2011, 79, 2267–2276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, M. Staphylococcus epidermidis—The ‘accidental’ pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitale, C.; Ma, T.M.; Sim, J.; Altheim, C.; Martinez-Nieves, E.; Kadiyala, U.; Solomon, M.J.; VanEpps, J.S. Staphylococcus epidermidis Has Growth Phase Dependent Affinity for Fibrinogen and Resulting Fibrin Clot Elasticity. Front. Microbiol. 2021, 12, 1561. [Google Scholar] [CrossRef]
- Skovdal, S.M.; Hansen, L.K.; Ivarsen, D.M.; Zeng, G.; Büttner, H.; Rohde, H.; Jørgensen, N.P.; Meyer, R.L. Host factors abolish the need for polysaccharides and extracellular matrix-binding protein in Staphylococcus epidermidis biofilm formation. J. Med. Microbiol. 2021, 70, 001287. [Google Scholar] [CrossRef]
- Lamy, B.; Dargère, S.; Arendrup, M.C.; Parienti, J.-J.; Tattevin, P. How to optimize the use of blood cultures for the diagnosis of bloodstream infections? A state-of-the art. Front. Microbiol. 2016, 7, 697. [Google Scholar] [CrossRef]
- Cheesbrough, M. District Laboratory Practice in Tropical Countries, Part 2; Cambridge University Press: Cambridge, UK, 2006; pp. 64–67. [Google Scholar]
- Freeman, D.J.; Falkiner, F.R.; Keane, C.T. New method for detecting slime production by coagulase negative staphylococci. J. Clin. Pathol. 1989, 42, 872–874. [Google Scholar] [CrossRef] [Green Version]
- Arciola, C.R.; Campoccia, D.; Gamberini, S.; Cervellati, M.; Donati, E.; Montanaro, L. Detection of slime production by means of an optimised Congo red agar plate test based on a colourimetric scale in Staphylococcus epidermidis clinical isolates genotyped for ica locus. Biomaterials 2002, 23, 4233–4239. [Google Scholar] [CrossRef]
- Kwasny, S.M.; Opperman, T.J. Static Biofilm Cultures of Gram-Positive Pathogens Grown in a Microtiter Format Used for Anti-Biofilm Drug Discovery. Curr. Protoc. Pharmacol. 2010, 50, 13A.8.1–13A.8.23. [Google Scholar] [CrossRef]
- Vuong, C.; Kidder, J.B.; Jacobson, E.R.; Otto, M.; Proctor, R.A.; Somerville, G.A. Staphylococcus epidermidis Polysaccharide Intercellular Adhesin Production Significantly Increases during Tricarboxylic Acid Cycle Stress. J. Bacteriol. 2005, 187, 2967–2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shome, B.; Das Mitra, S.; Bhuvana, M.; Krithiga, N.; Velu, D.; Shome, R.; Isloor, S.; Barbuddhe, S.; Rahman, H. Multiplex PCR assay for species identification of bovine mastitis pathogens. J. Appl. Microbiol. 2011, 111, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Seng, R.; Kitti, T.; Thummeepak, R.; Kongthai, P.; Leungtongkam, U.; Wannalerdsakun, S.; Sitthisak, S. Biofilm formation of methicillin-resistant coagulase negative staphylococci (MR-CoNS) isolated from community and hospital environments. PLoS ONE 2017, 12, e0184172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Haqan, A.; Boswihi, S.S.; Pathan, S.; Udo, E.E. Antimicrobial resistance and virulence determinants in coagulase-negative staphylococci isolated mainly from preterm neonates. PLoS ONE 2020, 15, e0236713. [Google Scholar] [CrossRef]
- Kuzdan, C.; Soysal, A.; Çulha, G.; Altinkanat, G.; Soyletir, G.; Bakir, M. Three-year study of health care-associated infections in a Turkish pediatric ward. J. Infect. Dev. Ctries. 2014, 8, 1415–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlersson, G.; Hellmark, B.; Svartström, O.; Stenmark, B.; Söderquist, B. Phenotypic characterisation of coagulase-negative staphylococci isolated from blood cultures in newborn infants, with a special focus on Staphylococcus capitis. Acta Paediatr. 2017, 106, 1576–1582. [Google Scholar] [CrossRef]
- Mack, D.; Fischer, W.; Krokotsch, A.; Leopold, K.; Hartmann, R.; Egge, H.; Laufs, R. The intercellular adhesin involved in biofilm accumulation of Staphylococcus epidermidis is a linear beta-1, 6-linked glucosaminoglycan: Purification and structural analysis. J. Bacteriol. 1996, 178, 175–183. [Google Scholar] [CrossRef] [Green Version]
- Salina, A.; Guimarães, F.; Pereira, V.; Menozzi, B.; Rall, V.; Langoni, H. Detection of icaA, icaD, and bap genes and biofilm production in Staphylococcus aureus and non-aureus staphylococci isolated from subclinical and clinical bovine mastitis. Arq. Bras. Med. Veterinária Zootec. 2020, 72, 1034–1038. [Google Scholar] [CrossRef]
- Gajewska, J.; Chajęcka-Wierzchowska, W. Biofilm formation ability and presence of adhesion genes among coagulase-negative and coagulase-positive staphylococci isolates from raw cow’s milk. Pathogens 2020, 9, 654. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Nguyen, T.H.; Otto, M. The staphylococcal exopolysaccharide PIA—Biosynthesis and role in biofilm formation, colonization, and infection. Comput. Struct. Biotechnol. J. 2020, 18, 3324–3334. [Google Scholar] [CrossRef]
- Manandhar, S.; Singh, A.; Varma, A.; Pandey, S.; Shrivastava, N. Evaluation of methods to detect in vitro biofilm formation by staphylococcal clinical isolates. BMC Res. Notes 2018, 11, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solati, S.M.; Tajbakhsh, E.; Khamesipour, F.; Gugnani, H.C. Prevalence of virulence genes of biofilm producing strains of Staphylococcus epidermidis isolated from clinical samples in Iran. AMB Express 2015, 5, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darwish, S.F.; Asfour, H.A. Investigation of biofilm forming ability in Staphylococci causing bovine mastitis using phenotypic and genotypic assays. Sci. World J. 2013, 2013, 378492. [Google Scholar] [CrossRef] [Green Version]
- Mathur, T.; Singhal, S.; Khan, S.; Upadhyay, D.; Fatma, T.; Rattan, A. Detection of biofilm formation among the clinical isolates of staphylococci: An evaluation of three different screening methods. Indian J. Med. Microbiol. 2006, 24, 25–29. [Google Scholar] [CrossRef]
- Peerayeh, S.N.; Moghadas, A.J.; Behmanesh, M. Prevalence of virulence-related determinants in clinical isolates of staphylococcus epidermidis. Jundishapur J. Microbiol. 2016, 9, e30593. [Google Scholar]
- Salgueiro, V.C.; Iorio, N.L.P.; Ferreira, M.C.; Chamon, R.C.; Dos Santos, K.R.N. Methicillin resistance and virulence genes in invasive and nasal Staphylococcus epidermidis isolates from neonates. BMC Microbiol. 2017, 17, 15. [Google Scholar] [CrossRef] [Green Version]
- Mekni, M.A.; Bouchami, O.; Achour, W.; Ben Hassen, A. Strong biofilm production but not adhesion virulence factors can discriminate between invasive and commensal Staphylococcus epidermidis strains. Apmis 2012, 120, 605–611. [Google Scholar] [CrossRef]
- Okee, M.S.; Joloba, M.L.; Okello, M.; Najjuka, F.C.; Katabazi, F.A.; Bwanga, F.; Nanteza, A.; Kateete, D.P. Prevalence of virulence determinants in Staphylococcus epidermidis from ICU patients in Kampala, Uganda. J. Infect. Dev. Ctries. 2011, 6, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Sellman, B.R.; Timofeyeva, Y.; Nanra, J.; Scott, A.; Fulginiti, J.P.; Matsuka, Y.V.; Baker, S.M. Expression of Staphylococcus epidermidis SdrG Increases following Exposure to an In Vivo Environment. Infect. Immun. 2008, 76, 2950–2957. [Google Scholar] [CrossRef] [Green Version]
- França, A.; Cerca, N. Plasma is the main regulator of Staphylococcus epidermidis biofilms virulence genes transcription in human blood. FEMS Pathog. Dis. 2016, 74, ftv125. [Google Scholar] [CrossRef] [Green Version]
- Franca, A.; Pier, G.B.; Vilanova, M.; Cerca, N. Transcriptomic analysis of Staphylococcus epidermidis biofilm-released cells upon interaction with human blood circulating immune cells and soluble factors. Front. Microbiol. 2016, 7, 1143. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Abercrombie, J.J.; Jeffrey, N.R.; Leung, K.P. An improved medium for growing Staphylococcus aureus biofilm. J. Microbiol. Methods 2012, 90, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Sato, A.; Yamaguchi, T.; Hamada, M.; Ono, D.; Sonoda, S.; Oshiro, T.; Nagashima, M.; Kato, K.; Okazumi, S.; Katoh, R. Morphological and biological characteristics of Staphylococcus aureus biofilm formed in the presence of plasma. Microb. Drug Resist. 2019, 25, 668–676. [Google Scholar] [PubMed] [Green Version]
- Kiem, S.; Oh, W.S.; Peck, K.R.; Lee, N.Y.; Lee, J.-Y.; Song, J.-H.; Hwang, E.S.; Kim, E.-C.; Cha, C.Y.; Choe, K.-W. Phase variation of biofilm formation in Staphylococcus aureus by IS256 insertion and its impact on the capacity adhering to polyurethane surface. J. Korean Med. Sci. 2004, 19, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Murugesan, S.; Mani, S.; Kuppusamy, I.; Krishnan, P. Role of Insertion Sequence Element IS256 as a Virulence Marker and Its Association with Biofilm Formation among Methicillin-Resistant Staphylococcus epidermidis from Hospital and Community Settings in Chennai, South India. Indian J. Med. Microbiol. 2018, 36, 124–126. [Google Scholar] [CrossRef]
- Arciola, C.R.; Campoccia, D.; Ravaioli, S.; Montanaro, L. Polysaccharide intercellular adhesin in biofilm: Structural and regulatory aspects. Front. Cell. Infect. Microbiol. 2015, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Olson, M.E.; Todd, D.A.; Schaeffer, C.R.; Paharik, A.E.; Van Dyke, M.J.; Büttner, H.; Dunman, P.M.; Rohde, H.; Cech, N.B.; Fey, P.D. Staphylococcus epidermidis agr quorum-sensing system: Signal identification, cross talk, and importance in colonization. J. Bacteriol. 2014, 196, 3482–3493. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Khan, B.A.; Cheung, G.Y.; Bach, T.-H.L.; Jameson-Lee, M.; Kong, K.-F.; Queck, S.Y.; Otto, M. Staphylococcus epidermidis surfactant peptides promote biofilm maturation and dissemination of biofilm-associated infection in mice. J. Clin. Investig. 2011, 121, 238–248. [Google Scholar]
- Dobinsky, S.; Kiel, K.; Rohde, H.; Bartscht, K.; Knobloch, J.K.-M.; Horstkotte, M.A.; Mack, D. Glucose-Related Dissociation between icaADBC Transcription and Biofilm Expression by Staphylococcus epidermidis: Evidence for an Additional Factor Required for Polysaccharide Intercellular Adhesin Synthesis. J. Bacteriol. 2003, 185, 2879–2886. [Google Scholar] [CrossRef]
- Cramton, S.E.; Gerke, C.; Schnell, N.F.; Nichols, W.W.; Götz, F. The Intercellular Adhesion (ica) Locus Is Present in Staphylococcus aureus and Is Required for Biofilm Formation. Infect. Immun. 1999, 67, 5427–5433. [Google Scholar]


{kind=link}
| Name | Abbreviation | Primers (5ʹ 3ʹ) | Ampli-Con Size (bp) |
|---|---|---|---|
| S. epidermidis species specific | Rdr | F: AAGAGCGTGGAGAAAAGTATCAAG R: TCGATACCATCAAAAAGTTGG | 130 |
| * Intercellular adhesin D | icaD | F: ATGGTCAAGCCCAGACAGAG R: CGTGTTTTCAACATTTAATGCAA | 198 |
| Insertion Sequence 256 | IS256 | F: AGTCCTTTTACGGTACAATG R: TGTGCGCATCAGAAATAACG | 762 |
| ** Intercellular adhesin A | icaA | F: TCTCTTGCAGGAGCAATCAA R: TCAGGCACTAACATCCAGCA | 188 |
| Laminin binding Protein/Enolase | Eno | F: ACGTGCAGCAGCTGACT R: CAACAGCATCTTCAGTACCTTC | 301 |
| Fibrinogen binding Protein | fbe/SdrG | F: CTACAAGTTCAGGTCAAGGACAAGG R: GCGTCGGCGTATATCCTTCAG | 273 |
| Accumulation associated protein | Aap | F: AAACGGTGGTATCTTACGTGAA R: CAATGTTGCACCATCTAAATCAGCT | 465 |
| Bap homologue Protein | Bhp | F: ATGGTATTAGCAAGCTCTCAGCTGG R: AGGGTTTCCATCTGGATCCG | 1585 |
| Extracellular matrix binding protein | Embp | F: AGCGGTACAAATGTCAAT R: AGAAGTGCTCTAGCATCATCC | 455 |
| Biofilm associated protein | Bap | F: CCCTATATCGAAGGTGTAGAATTG R: GCTGTTGAAGTTAATACTGTACCTGC | 971 |
| MSCRAMMs/CWA | Biofilm Positive | Biofilm Negative | p-Values | |||||
|---|---|---|---|---|---|---|---|---|
| In Biofilm Positive Isolates n (%) | Total Strong n (%) 55 (84.6%) | Moderate n (%) | Weak n (%) | Negative n (%) | SG * n (%) | |||
| V. Strong n (%) | Strong n (%) | |||||||
| fbe a | 59 (90.7%) | 11 (84.6%) | 42 (100%) | 5 (71.4%) | 1 (33.3%) | 4 (80%) | 2 (40%) | 0.007 |
| embp a | 57 (87. 6%) | 10 (77%) | 42 (100%) | 4 (57.1%) | 1 (33.3%) | 4 (80%) | 2 (40%) | 0.026 |
| eno b | 60 (92.3%) | 11 (84.6%) | 42 (100%) | 5 (71.4%) | 2 (66.6%) | 5 (100%) | 4 (80%) | 0.802 |
| aap c | 33 (50.7%) | 5 (38.4%) | 25 (59.5%) | 2 (28.5%) | 1 (33.3%) | 2 (40%) | 0 (0%) | 0.069 |
| bhp c | 15 (23%) | 4 (30.7%) | 11 (26.1%) | 0 (0%) | 0 (0%) | 2 (40%) | 1 (20%) | 0.633 |
| bap c | 1 (1.5%) | 0 (0%) | 1 (2.3%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | NA |
| Total isolates (n = 75) | 65 (86.6%) | 13 (20%) | 42 (64.6%) | 7 (10.7%) | 3 (4.6%) | 5 (7.6%) | 5 (7.6%) | NA |
| icaADBC Operon | Biofilm Positive (n = 65) | Biofilm Negative (n = 10) | p-Values | |||||
|---|---|---|---|---|---|---|---|---|
| In Biofilm Positive Isolates n (%) | Total Strong n (%) 55 (84.6%) | Moderate n (%) | Weak n (%) | Negative n (%) | SG * n (%) | |||
| V. Strong n (%) | Strong n (%) | |||||||
| Complete a (Actively complete) b | 26 (40%) (21 (80.7% ** & 28% ***)) | 5 (19%) 4 (19%) | 19 (73%) 15 (71.4%) | 2 (7.6%) 2 (9.5%) | 0 (0%) 0 (0%) | 0 (0%) 0 (0%) | 1 (3.8%) 1 (4.7%) | .065 0.149 |
| Defective c | 15 (23%) | 2 (13.3%) | 11 (73.3%) | 0 (0%) | 2 (13.3%) | 2 (13.3%) | 2 (13.3%) | 0.252 |
| Missing d | 24 (37%) | 0 (0%) | 16 (66.6%) | 6 (25%) | 2 (8.3%) | 3 (8.8%) | 2 (5.8%) | 0.429 |
| Total isolates (n = 75) | 65 (86.6%) | 13 (20%) | 42 (64.6%) | 7 (10.7%) | 3(4.6%) | 5 (7.6%) | 5 (7.6%) | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, S.; Rahman, H.; Qasim, M.; Nawab, J.; Alzahrani, K.J.; Alsharif, K.F.; Alzahrani, F.M. Staphylococcus epidermidis Pathogenesis: Interplay of icaADBC Operon and MSCRAMMs in Biofilm Formation of Isolates from Pediatric Bacteremia in Peshawar, Pakistan. Medicina 2022, 58, 1510. https://doi.org/10.3390/medicina58111510
Ahmad S, Rahman H, Qasim M, Nawab J, Alzahrani KJ, Alsharif KF, Alzahrani FM. Staphylococcus epidermidis Pathogenesis: Interplay of icaADBC Operon and MSCRAMMs in Biofilm Formation of Isolates from Pediatric Bacteremia in Peshawar, Pakistan. Medicina. 2022; 58(11):1510. https://doi.org/10.3390/medicina58111510
Chicago/Turabian StyleAhmad, Saghir, Hazir Rahman, Muhammad Qasim, Javed Nawab, Khalid J. Alzahrani, Khalaf F. Alsharif, and Fuad M. Alzahrani. 2022. "Staphylococcus epidermidis Pathogenesis: Interplay of icaADBC Operon and MSCRAMMs in Biofilm Formation of Isolates from Pediatric Bacteremia in Peshawar, Pakistan" Medicina 58, no. 11: 1510. https://doi.org/10.3390/medicina58111510