

Molecular Pathways Related to Sulforaphane as Adjuvant Treatment: A Nanomedicine Perspective in Breast Cancer

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Effects of Glucosinolates Derived from Cruciferous Vegetables
3. Apoptosis, Cell Cycle, Autophagy, and Suppression of the PI3K-AKT-mTOR Pathway Are the Key Biological Processes Influenced by Sulforaphane
3.1. Sulforaphane Promotes Mitochondria-Mediated Apoptosis
3.2. Effects of Sulforaphane Treatment on Cell Cycle
3.3. Sulforaphane Treatment Promotes Autophagy
3.4. Sulforaphane Treatment Has Negative Effects on the PI3K-AKT-mTOR Oncogenic Signalling Pathway Preventing Carcinogenesis and Angiogenesis Process
4. Tumour-Suppressive Effects of Sulforaphane in Breast Cancer
5. Sulforaphane as Potential Chemotherapy Adjuvants in Breast Cancer
6. Anti-Tumour Effects of Sulforaphane Nanoparticles: Promising Chemotherapeutic Adjuvants in Breast Cancer
Potential Effects of Sulforaphane Nanoparticles as Adjuvant
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative Stress, Inflammation, and Cancer: How Are They Linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malumbres, M.; Carnero, A. Cell Cycle Deregulation: A Common Motif in Cancer. Prog. Cell Cycle Res. 2003, 5, 5–18. [Google Scholar] [PubMed]
- Klaunig, J.E. Oxidative Stress and Cancer. Curr. Pharm. Des. 2019, 24, 4771–4778. [Google Scholar] [CrossRef] [PubMed]
- Resnik, D.B. Responsibility for Health: Personal, Social, and Environmental. J. Med. Ethics 2007, 33, 444–445. [Google Scholar] [CrossRef]
- Chen, Y.; Michalak, M.; Agellon, L.B. Importance of Nutrients and Nutrient Metabolism on Human Health. Yale J. Biol. Med. 2018, 91, 95–103. [Google Scholar]
- Wiseman, M.J. Nutrition and Cancer: Prevention and Survival. Br. J. Nutr. 2019, 122, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Baena Ruiz, R.; Salinas Hernández, P. Diet and Cancer: Risk Factors and Epidemiological Evidence. Maturitas 2014, 77, 202–208. [Google Scholar] [CrossRef]
- Kensler, T.W. Chemoprevention by Inducers of Carcinogen Detoxication Enzymes. Environ. Health Perspect. 1997, 105, 965–970. [Google Scholar] [CrossRef]
- Moloney, J.N.; Cotter, T.G. ROS Signalling in the Biology of Cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef]
- Ramsey, A.T.; Baker, T.B.; Pham, G.; Stoneking, F.; Smock, N.; Colditz, G.A.; James, A.S.; Liu, J.; Bierut, L.J.; Chen, L.-S. Low Burden Strategies Are Needed to Reduce Smoking in Rural Healthcare Settings: A Lesson from Cancer Clinics. Int. J. Environ. Res. Public Health 2020, 17, 1728. [Google Scholar] [CrossRef] [Green Version]
- Surman, M.; Janik, M.E. Stress and Its Molecular Consequences in Cancer Progression. Postepy Hig. Med. Dosw. Online 2017, 71, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Fiolet, T.; Srour, B.; Sellem, L.; Kesse-Guyot, E.; Allès, B.; Méjean, C.; Deschasaux, M.; Fassier, P.; Latino-Martel, P.; Beslay, M.; et al. Consumption of Ultra-Processed Foods and Cancer Risk: Results from NutriNet-Santé Prospective Cohort. BMJ 2018, 360, k322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bober, P.; Alexovič, M.; Tomková, Z.; Kilík, R.; Sabo, J. RHOA and MDia1 Promotes Apoptosis of Breast Cancer Cells via a High Dose of Doxorubicin Treatment. Open Life Sci. 2019, 14, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Tantamango-Bartley, Y.; Jaceldo-Siegl, K.; Fan, J.; Fraser, G. Vegetarian Diets and the Incidence of Cancer in a Low-Risk Population. Cancer Epidemiol. Biomark. Prev. 2013, 22, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Samanta, S.K.; Choudhury, P.; Sarma, P.P.; Gogoi, B.; Gogoi, N.; Devi, R. Dietary Phytochemicals/Nutrients as Promising Protector of Breast Cancer Development: A Comprehensive Analysis. Pharmacol. Rep. 2022, 74, 583–601. [Google Scholar] [CrossRef]
- Abdull Razis, A.F.; Noor, N.M. Cruciferous Vegetables: Dietary Phytochemicals for Cancer Prevention. Asian Pac. J. Cancer Prev. 2013, 14, 1565–1570. [Google Scholar] [CrossRef] [Green Version]
- Higdon, J.; Delage, B.; Williams, D.; Dashwood, R. Cruciferous Vegetables and Human Cancer Risk: Epidemiologic Evidence and Mechanistic Basis. Pharmacol. Res. 2007, 55, 224–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, A.R. Taxonomy and Evolution of Broccoli (Brassica oleracea Var. Italica). Econ. Bot. 1982, 36, 397–410. [Google Scholar] [CrossRef]
- Pickering, R.J.; Rosado, C.J.; Sharma, A.; Buksh, S.; Tate, M.; de Haan, J.B. Recent Novel Approaches to Limit Oxidative Stress and Inflammation in Diabetic Complications. Clin. Transl. Immunol. 2018, 7, e1016. [Google Scholar] [CrossRef]
- Fischer, R.; Maier, O. Interrelation of Oxidative Stress and Inflammation in Neurodegenerative Disease: Role of TNF. Oxid. Med. Cell. Longev. 2015, 2015, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Lynch, R.; Diggins, E.L.; Connors, S.L.; Zimmerman, A.W.; Singh, K.; Liu, H.; Talalay, P.; Fahey, J.W. Sulforaphane from Broccoli Reduces Symptoms of Autism: A Follow-up Case Series from a Randomized Double-Blind Study. Glob. Adv. Health Med. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Sivapalan, T.; Melchini, A.; Saha, S.; Needs, P.W.; Traka, M.H.; Tapp, H.; Dainty, J.R.; Mithen, R.F. Bioavailability of Glucoraphanin and Sulforaphane from High-Glucoraphanin Broccoli. Mol. Nutr. Food Res. 2018, 62, 1700911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.-H.; Lin, K.-H.; Chen, S.-C.; Shen, Y.-H.; Lo, H.-F. Proteomic Analysis of Broccoli (Brassica oleracea) under High Temperature and Waterlogging Stresses. Bot. Stud. 2015, 56, 18. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, S.; Thamilarasan, S.; Park, J.-I.; Chung, M.-Y.; Nou, I.-S. Molecular Modeling of Myrosinase from Brassica Oleracea: A Structural Investigation of Sinigrin Interaction. Genes 2015, 6, 1315–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludikhuyze, L.; Rodrigo, L.; Hendrickx, M. The Activity of Myrosinase from Broccoli (Brassica oleracea L. Cv. Italica): Influence of Intrinsic and Extrinsic Factors. J. Food Prot. 2000, 63, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Egner, P.A.; Agyeman, A.S.; Visvanathan, K.; Groopman, J.D.; Chen, J.-G.; Chen, T.-Y.; Fahey, J.W.; Talalay, P. Keap1–Nrf2 Signaling: A Target for Cancer Prevention by Sulforaphane. In Natural Products in Cancer Prevention and Therapy; Pezzuto, J.M., Suh, N., Eds.; Topics in Current Chemistry; Springer: Berlin/Heidelberg, Germany, 2012; Volume 329, pp. 163–177. ISBN 978-3-642-34574-6. [Google Scholar]
- Cottaz, S.; Rollin, P.; Driguez, H. ChemInform Abstract: Synthesis of 2-Deoxy-2-Fluoro-Glucotropaeolin, a Thioglucosidase Inhibitor. ChemInform 2010, 28. [Google Scholar] [CrossRef]
- de Oliveira, M.R. Sulforaphane and Its Modulation of Brain Redox Status: The Mitochondria as a Target. In Oxidative Stress and Dietary Antioxidants in Neurological Diseases; Academic Press: Cambridge, MA, USA, 2020; pp. 377–391. [Google Scholar] [CrossRef]
- Holst, B.; Williamson, G. A Critical Review of the Bioavailability of Glucosinolates and Related Compounds. Nat. Prod. Rep. 2004, 21, 425. [Google Scholar] [CrossRef]
- Faulkner, K. Selective Increase of the Potential Anticarcinogen 4- Methylsulphinylbutyl Glucosinolate in Broccoli. Carcinogenesis 1998, 19, 605–609. [Google Scholar] [CrossRef] [Green Version]
- Preobrazhenskaya, M.N.; Korolev, A.M. Indole Derivatives in Vegetables of the Cruciferae Family. Russ. J. Bioorganic Chem. 2000, 26, 85–97. [Google Scholar] [CrossRef]
- Kong, X.Y.; Kissen, R.; Bones, A.M. Characterization of Recombinant Nitrile-Specifier Proteins (NSPs) of Arabidopsis Thaliana: Dependency on Fe(II) Ions and the Effect of Glucosinolate Substrate and Reaction Conditions. Phytochemistry 2012, 84, 7–17. [Google Scholar] [CrossRef]
- Foo, H.L.; Grønning, L.M.; Goodenough, L.; Bones, A.M.; Danielsen, B.-E.; Whiting, D.A.; Rossiter, J.T. Purification and Characterisation of Epithiospecifier Protein from Brassica Napus: Enzymic Intramolecular Sulphur Addition within Alkenyl Thiohydroximates Derived from Alkenyl Glucosinolate Hydrolysis. FEBS Lett. 2000, 468, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, N.; Kar, S.; Parbin, S.; Sengupta, D.; Deb, M.; Das, L.; Patra, S.K. Epigenetic Dietary Interventions for Prevention of Cancer. In Epigenetics of Cancer Prevention; Elsevier: Amsterdam, The Netherlands, 2019; pp. 23–48. ISBN 978-0-12-812494-9. [Google Scholar]
- Bauman, J.E.; Zang, Y.; Sen, M.; Li, C.; Wang, L.; Egner, P.A.; Fahey, J.W.; Normolle, D.P.; Grandis, J.R.; Kensler, T.W.; et al. Prevention of Carcinogen-Induced Oral Cancer by Sulforaphane. Cancer Prev. Res. 2016, 9, 547–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, R.; Gong Essel, K.; Mangiaracina Benbrook, D.; Garland, J.; Daniel Zhao, Y.; Chandra, V. Preclinical Efficacy and Involvement of AKT, MTOR, and ERK Kinases in the Mechanism of Sulforaphane against Endometrial Cancer. Cancers 2020, 12, 1273. [Google Scholar] [CrossRef] [PubMed]
- Hudecova, S.; Markova, J.; Simko, V.; Csaderova, L.; Stracina, T.; Sirova, M.; Fojtu, M.; Svastova, E.; Gronesova, P.; Pastorek, M.; et al. Sulforaphane-Induced Apoptosis Involves the Type 1 IP3 Receptor. Oncotarget 2016, 7, 61403–61418. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, S.; Horinaka, M.; Sakai, T. Sulforaphane Enhances Apoptosis Induced by Lactobacillus Pentosus Strain S-PT84 via the TNFα Pathway in Human Colon Cancer Cells. Oncol. Lett. 2019, 18, 4253–4261. [Google Scholar] [CrossRef] [Green Version]
- Myzak, M.C.; Hardin, K.; Wang, R.; Dashwood, R.H.; Ho, E. Sulforaphane Inhibits Histone Deacetylase Activity in BPH-1, LnCaP and PC-3 Prostate Epithelial Cells. Carcinogenesis 2006, 27, 811–819. [Google Scholar] [CrossRef]
- Mokhtari, R.B.; Qorri, B.; Baluch, N.; Sparaneo, A.; Fabrizio, F.P.; Muscarella, L.A.; Tyker, A.; Kumar, S.; Cheng, H.-L.M.; Szewczuk, M.R.; et al. Next-Generation Multimodality of Nutrigenomic Cancer Therapy: Sulforaphane in Combination with Acetazolamide Actively Target Bronchial Carcinoid Cancer in Disabling the PI3K/Akt/MTOR Survival Pathway and Inducing Apoptosis. Oncotarget 2021, 12, 1470–1489. [Google Scholar] [CrossRef]
- Hahm, E.-R.; Singh, K.B.; Kim, S.-H.; Powolny, A.A.; Singh, S.V. The Role of Lysosome-Associated Membrane Protein 2 in Prostate Cancer Chemopreventive Mechanisms of Sulforaphane. Cancer Prev. Res. 2020, 13, 661–672. [Google Scholar] [CrossRef]
- Sundaram, M.K.; Khan, M.A.; Alalami, U.; Somvanshi, P.; Bhardwaj, T.; Pramodh, S.; Raina, R.; Shekfeh, Z.; Haque, S.; Hussain, A. Phytochemicals Induce Apoptosis by Modulation of Nitric Oxide Signaling Pathway in Cervical Cancer Cells. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 11827–11844. [Google Scholar] [CrossRef]
- Myzak, M.C.; Dashwood, R.H. Chemoprotection by Sulforaphane: Keep One Eye beyond Keap1. Cancer Lett. 2006, 233, 208–218. [Google Scholar] [CrossRef] [Green Version]
- Hao, Q.; Wang, M.; Sun, N.; Zhu, C.; Lin, Y.; Li, C.; Liu, F.; Zhu, W. Sulforaphane Suppresses Carcinogenesis of Colorectal Cancer through the ERK/Nrf2-UDP Glucuronosyltransferase 1A Metabolic Axis Activation. Oncol. Rep. 2020, 43, 1067–1080. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.V.; Herman-Antosiewicz, A.; Singh, A.V.; Lew, K.L.; Srivastava, S.K.; Kamath, R.; Brown, K.D.; Zhang, L.; Baskaran, R. Sulforaphane-Induced G2/M Phase Cell Cycle Arrest Involves Checkpoint Kinase 2-Mediated Phosphorylation of Cell Division Cycle 25C. J. Biol. Chem. 2004, 279, 25813–25822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, G.; Xu, C.; Chen, C.; Hebbar, V.; Kong, A.-N.T. P53-Independent G1 Cell Cycle Arrest of Human Colon Carcinoma Cells HT-29 by Sulforaphane Is Associated with Induction of P21CIP1 and Inhibition of Expression of Cyclin D1. Cancer Chemother. Pharmacol. 2006, 57, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Traka, M.; Gasper, A.V.; Smith, J.A.; Hawkey, C.J.; Bao, Y.; Mithen, R.F. Transcriptome Analysis of Human Colon Caco-2 Cells Exposed to Sulforaphane. J. Nutr. 2005, 135, 1865–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parnaud, G.; Li, P.; Cassar, G.; Rouimi, P.; Tulliez, J.; Combaret, L.; Gamet-Payrastre, L. Mechanism of Sulforaphane-Induced Cell Cycle Arrest and Apoptosis in Human Colon Cancer Cells. Nutr. Cancer 2004, 48, 198–206. [Google Scholar] [CrossRef]
- Kung, C.-P.; Budina, A.; Balaburski, G.; Bergenstock, M.K.; Murphy, M. Autophagy in Tumor Suppression and Cancer Therapy. Crit. Rev. Eukaryot. Gene Expr. 2011, 21, 71–100. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Ren, Y.; Yang, L.; Jia, A.; Hu, Y.; Zhao, Y.; Zhao, W.; Yu, B.; Zhao, W.; Zhang, J.; et al. Inhibiting Autophagy Enhances Sulforaphane-Induced Apoptosis via Targeting NRF2 in Esophageal Squamous Cell Carcinoma. Acta Pharm. Sin. B 2021, 11, 1246–1260. [Google Scholar] [CrossRef]
- Alzahrani, A.S. PI3K/Akt/MTOR Inhibitors in Cancer: At the Bench and Bedside. Semin. Cancer Biol. 2019, 59, 125–132. [Google Scholar] [CrossRef]
- Liu, P.; Atkinson, S.J.; Akbareian, S.E.; Zhou, Z.; Munsterberg, A.; Robinson, S.D.; Bao, Y. Sulforaphane Exerts Anti-Angiogenesis Effects against Hepatocellular Carcinoma through Inhibition of STAT3/HIF-1α/VEGF Signalling. Sci. Rep. 2017, 7, 12651. [Google Scholar] [CrossRef] [Green Version]
- Davis, R.; Singh, K.P.; Kurzrock, R.; Shankar, S. Sulforaphane Inhibits Angiogenesis through Activation of FOXO Transcription Factors. Oncol. Rep. 2009, 22, 1473–1478. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Wang, H.; Zhang, Z.; Jiang, B.; Luo, J.; Shi, X. Sulforaphane Inhibited Expression of Hypoxia-inducible Factor-1α in Human Tongue Squamous Cancer Cells and Prostate Cancer Cells. Int. J. Cancer 2008, 123, 1255–1261. [Google Scholar] [CrossRef] [PubMed]
- Castro, N.P.; Rangel, M.C.; Merchant, A.S.; MacKinnon, G.; Cuttitta, F.; Salomon, D.S.; Kim, Y.S. Sulforaphane Suppresses the Growth of Triple-Negative Breast Cancer Stem-like Cells In Vitro and In Vivo. Cancer Prev. Res. 2019, 12, 147–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royston, K.J.; Paul, B.; Nozell, S.; Rajbhandari, R.; Tollefsbol, T.O. Withaferin A and Sulforaphane Regulate Breast Cancer Cell Cycle Progression through Epigenetic Mechanisms. Exp. Cell Res. 2018, 368, 67–74. [Google Scholar] [CrossRef]
- Lewinska, A.; Adamczyk-Grochala, J.; Deregowska, A.; Wnuk, M. Sulforaphane-Induced Cell Cycle Arrest and Senescence Are Accompanied by DNA Hypomethylation and Changes in MicroRNA Profile in Breast Cancer Cells. Theranostics 2017, 7, 3461–3477. [Google Scholar] [CrossRef] [PubMed]
- Pore, S.K.; Hahm, E.-R.; Kim, S.-H.; Singh, K.B.; Nyiranshuti, L.; Latoche, J.D.; Anderson, C.J.; Adamik, J.; Galson, D.L.; Weiss, K.R.; et al. A Novel Sulforaphane-Regulated Gene Network in Suppression of Breast Cancer–Induced Osteolytic Bone Resorption. Mol. Cancer Ther. 2020, 19, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lu, Q.; Li, N.; Xu, M.; Miyamoto, T.; Liu, J. Sulforaphane Suppresses Metastasis of Triple-Negative Breast Cancer Cells by Targeting the RAF/MEK/ERK Pathway. NPJ Breast Cancer 2022, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Yasunaga, A.; Ono, M.; Takeshima, M.; Nakano, S. Sulforaphane Suppresses the Growth of EGFR-overexpressing MDA-MB-468 Triple-negative Breast Cancer Cells in Vivo and in Vitro. Int. J. Funct. Nutr. 2022, 3, 3. [Google Scholar] [CrossRef]
- Mielczarek, L.; Krug, P.; Mazur, M.; Milczarek, M.; Chilmonczyk, Z.; Wiktorska, K. In the Triple-Negative Breast Cancer MDA-MB-231 Cell Line, Sulforaphane Enhances the Intracellular Accumulation and Anticancer Action of Doxorubicin Encapsulated in Liposomes. Int. J. Pharm. 2019, 558, 311–318. [Google Scholar] [CrossRef]
- Kamal, M.M.; Nazzal, S. Novel Sulforaphane-Enabled Self-Microemulsifying Delivery Systems (SFN-SMEDDS) of Taxanes: Formulation Development and in Vitro Cytotoxicity against Breast Cancer Cells. Int. J. Pharm. 2018, 536, 187–198. [Google Scholar] [CrossRef]
- Xu, Y.; Han, X.; Li, Y.; Min, H.; Zhao, X.; Zhang, Y.; Qi, Y.; Shi, J.; Qi, S.; Bao, Y.; et al. Sulforaphane Mediates Glutathione Depletion via Polymeric Nanoparticles to Restore Cisplatin Chemosensitivity. ACS Nano 2019, 13, 13445–13455. [Google Scholar] [CrossRef]
- Huang, J.; Tao, C.; Yu, Y.; Yu, F.; Zhang, H.; Gao, J.; Wang, D.; Chen, Y.; Gao, J.; Zhang, G.; et al. Simultaneous Targeting of Differentiated Breast Cancer Cells and Breast Cancer Stem Cells by Combination of Docetaxel- and Sulforaphane-Loaded Self-Assembled Poly(D, L-Lactide-Co-Glycolide)/Hyaluronic Acid Block Copolymer-Based Nanoparticles. J. Biomed. Nanotechnol. 2016, 12, 1463–1477. [Google Scholar] [CrossRef] [PubMed]
- Keshandehghan, A.; Nikkhah, S.; Tahermansouri, H.; Heidari-Keshel, S.; Gardaneh, M. Co-Treatment with Sulforaphane and Nano-Metformin Molecules Accelerates Apoptosis in HER2+ Breast Cancer Cells by Inhibiting Key Molecules. Nutr. Cancer 2020, 72, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Krug, P.; Mielczarek, L.; Wiktorska, K.; Kaczyńska, K.; Wojciechowski, P.; Andrzejewski, K.; Ofiara, K.; Szterk, A.; Mazur, M. Sulforaphane-Conjugated Selenium Nanoparticles: Towards a Synergistic Anticancer Effect. Nanotechnology 2019, 30, 065101. [Google Scholar] [CrossRef] [PubMed]
- Krug, P.; Wiktorska, K.; Kaczyńska, K.; Ofiara, K.; Szterk, A.; Kuśmierz, B.; Mazur, M. Sulforaphane-assisted preparation of tellurium flower-like nanoparticles. Nanotechnology 2019, 31, 055603. [Google Scholar] [CrossRef] [PubMed]
- Milczarek, M.; Wiktorska, K.; Mielczarek, L.; Koronkiewicz, M.; Dąbrowska, A.; Lubelska, K.; Matosiuk, D.; Chilmonczyk, Z. Autophagic Cell Death and Premature Senescence: New Mechanism of 5-Fluorouracil and Sulforaphane Synergistic Anti-cancer Effect in MDA-MB-231 Triple Negative Breast Cancer Cell Line. Food Chem. Toxicol. 2018, 111, 1–8. [Google Scholar] [CrossRef]
- Bose, C.; Awasthi, S.; Sharma, R.; Beneš, H.; Hauer-Jensen, M.; Boerma, M.; Singh, S.P. Sulforaphane Potentiates Anticancer Effects of Doxorubicin and Attenuates Its Cardiotoxicity in a Breast Cancer Model. PLoS ONE 2018, 13, e0193918. [Google Scholar] [CrossRef]
- Rong, Y.; Huang, L.; Yi, K.; Chen, H.; Liu, S.; Zhang, W.; Yuan, C.; Song, X.; Wang, F. Co-Administration of Sulforaphane and Doxorubicin Attenuates Breast Cancer Growth by Preventing the Accumulation of Myeloid-Derived Suppressor Cells. Cancer Lett. 2020, 493, 189–196. [Google Scholar] [CrossRef]
- Burnett, J.P.; Lim, G.; Li, Y.; Shah, R.B.; Lim, R.; Paholak, H.J.; McDermott, S.P.; Sun, L.; Tsume, Y.; Bai, S.; et al. Sulforaphane Enhances the Anticancer Activity of Taxanes against Triple Negative Breast Cancer by Killing Cancer Stem Cells. Cancer Lett. 2017, 394, 52–64. [Google Scholar] [CrossRef]
- Kim, S.-H.; Park, H.-J.; Moon, D.-O. Sulforaphane Sensitizes Human Breast Cancer Cells to Paclitaxel-Induced Apoptosis by Downregulating the NF-ΚB Signaling Pathway. Oncol. Lett. 2017, 13, 4427–4432. [Google Scholar] [CrossRef] [Green Version]
- Danafar, H.; Sharafi, A.; Kheiri, S.; Kheiri Manjili, H. Co -Delivery of Sulforaphane and Curcumin with PEGylated Iron Oxide-Gold Core Shell Nanoparticles for Delivery to Breast Cancer Cell Line. Iran. J. Pharm. Res. IJPR 2018, 17, 480–494. [Google Scholar]
- Danafar, H.; Sharafi, A.; Kheiri Manjili, H.; Andalib, S. Sulforaphane Delivery Using MPEG–PCL Co-Polymer Nanoparticles to Breast Cancer Cells. Pharm. Dev. Technol. 2017, 22, 642–651. [Google Scholar] [CrossRef] [PubMed]
- Kheiri Manjili, H.; Ma’mani, L.; Tavaddod, S.; Mashhadikhan, M.; Shafiee, A.; Naderi-Manesh, H. D, L-Sulforaphane Loaded Fe3O4@ Gold Core Shell Nanoparticles: A Potential Sulforaphane Delivery System. PLoS ONE 2016, 11, e0151344. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Ren, F.; Mao, X.; Du, M. Mineralized and GSH-Responsive Hyaluronic Acid Based Nano-Carriers for Potentiating Repressive Effects of Sulforaphane on Breast Cancer Stem Cells-like Properties. Carbohydr. Polym. 2021, 269, 118294. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
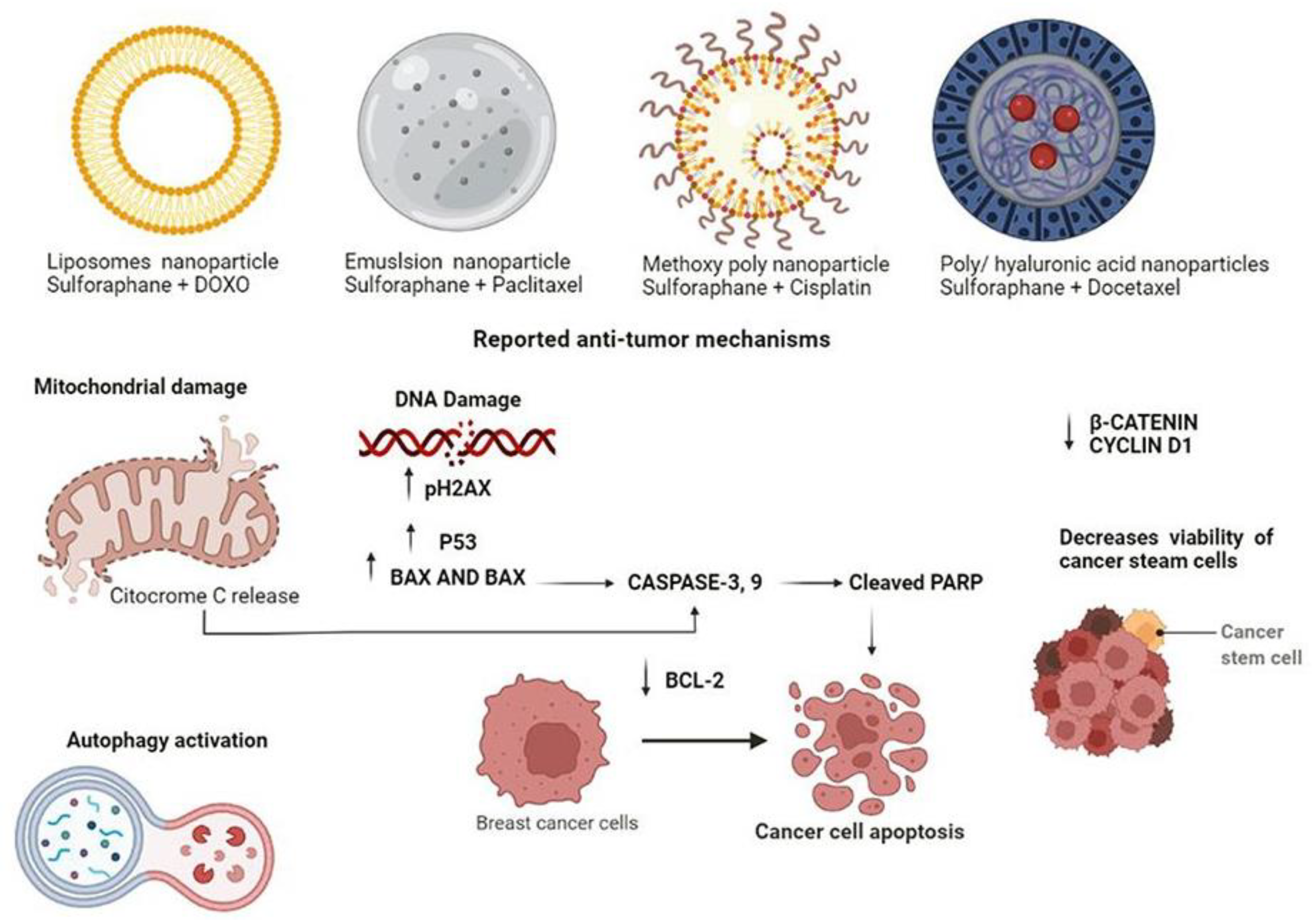{kind=link}
| Precursor | Molecule Products | Conditions or Necessary Enzyme | Reference |
|---|---|---|---|
| Glucoraphanin | Thiohydroxamate-O-sulfonate and glucose | Myrosinase | Kensler, et al. [26] Cottaz, et al. [27] de Oliveira [28] |
| Stable Isothiocyanates | |||
| Thiohydroxamate-O-sulfonate | Sulforaphane | pH 6–7, microbial thioglucosidase | Kensler, et al. [26] Holst [29] Faulkner [30] |
| Erucin | |||
| Iberin | |||
| Unstable isothiocyanates | |||
| β-hydroxylated Isothiocyanate | Goitrin | pH 6–7 and spontaneous cyclisation | Ludikhuyze, et al. [25] |
| Indolylmethyl-isothiocyanate | 3,3-Diindolylmethane | Trimeres, tetramers, etc. of Indole-3-carbinol | Probrazhenskaya [31] |
| Ascorbigen | Ascorbic acid presence | Faulkner [30] Preobrazhenskaya [31] | |
| Nitrile aliphatic and aromatic, indole and β-OH-nitriles | |||
| Thiohydroxamate-O-sulfonate | Crambene | pH 3–7 and the presence of nitrile-forming factor | Kong, et al. [32] |
| Others | |||
| Thiohydroxamate-O-sulfonate | Epithioalkylnitrile | Presence of double bound at Radical, epithiospecifer protein and Fe2+ | Cottaz, et al. [27] Kong, et al. [32] Foo, et al. [33] |
| Thiocyanate | Not clear | Pradhan [34] | |
| Subjects | Sulforaphane Dosage | Anticancer Effect | Genes Targets | Reference | |
|---|---|---|---|---|---|
| MDA-MB 231. | 40 µm | Apoptosis and necrosis | ZEB1, Claudin, and Fibronectin | Mokhtari et al. [40] | |
| BalbC/nude mice | 50 mg/kg | Apoptosis and decrease in tumour volume. | ALDH1A1, NANOG, GDF3 and FOXD3 | Castro et al. [55] | |
| MC7-cells and MDA-MB-231 | 5.0 μM | Cell cycle arrest | CCND1 and CDK4 | Royston et al. [56] | |
| MCF-7 and MDA-MB-231 | 4.05 M, 19.35 M, and 16.64 M | Cell cycle arrest | cell cycle arrest | Lewinska et al. [57] | |
| MDA-MB-231, SK-BR-3, and MCF-7 | 20 ng/mL | Cell cycle, suppression of osteolytic bone resorption | RUNX2 and NF-κB1 | Pore et al. [58] | |
| MDA-MB-231, SK-BR-3, and MCF-7 | 7.5–30 µM | Migration and invasion of breast cancer cells | RAF/MEK/ERK pathway | Zhang et al. [59] | |
| MDA-MB-468 | 1.8 µM | Cell cycle arrest | EGFR, BCL2, BAX and Akt/mTOR pathway | Yasunaga et al. [60] | |
| Nanoparticles in sulforaphane treatment | |||||
| Subjects | Sulforaphane dosage | Anticancer Effect | Genes Targets | Nanoparticles | References |
| MDA-MB-231 and MCF7 | 487.5 mg/g + 9.375 mg/g of DOX | Inhibit tumour cell growth, ROS generation, mitochondrial damage and autophagy | Did not report | Liposome nanoparticles | Mielczarek et al. [61] |
| MDA-MB-231 and MCF-7 | 8.7 µM and 500 nM of paclitaxel (PTX) | Growth suppression | Did not report | Micro-emulsion nanoparticle | Kamal et al. [62] |
| MCF-7 | 48.97 µM and cisplatin at 72.59 µM | Induction of apoptosis through DNA damage. | p-H2AX, p53, PARP and Bcl-2 | Methoxy poly (ethylene glycol)-poly nanoparticle | Xu et al. [63] |
| Breast cancer CD44+ CD24- | 350 µg and docetaxel (DTX) 5 nM | Cell growth inhibition | B-catenin and Cyclin D1 | Poly (D, L-lactideco-glycolide)/hyaluronic acid nanoparticles | Huang et al. [64] |
| MCF-7 and BT-474 | 48 nM and metformin 42 nM | Cell viability decrease and apoptosis | WNT-1, B-catenin, and CD44. | Carbon nanoparticles | Keshandehghan et al. [65] |
| Adult male Wistar rats | 10 mg/kg | Cell viability decrement | Nrf2 pathway | Selenium nanoparticle | Krug et al. [66] |
| Adult male Wistar rats | 10 mg/kg | Induction of ROS production | Did not report | flower-like tellurium nanoparticles | Krug et al. [67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saavedra-Leos, M.Z.; Jordan-Alejandre, E.; Puente-Rivera, J.; Silva-Cázares, M.B. Molecular Pathways Related to Sulforaphane as Adjuvant Treatment: A Nanomedicine Perspective in Breast Cancer. Medicina 2022, 58, 1377. https://doi.org/10.3390/medicina58101377
Saavedra-Leos MZ, Jordan-Alejandre E, Puente-Rivera J, Silva-Cázares MB. Molecular Pathways Related to Sulforaphane as Adjuvant Treatment: A Nanomedicine Perspective in Breast Cancer. Medicina. 2022; 58(10):1377. https://doi.org/10.3390/medicina58101377
Chicago/Turabian StyleSaavedra-Leos, María Zenaida, Euclides Jordan-Alejandre, Jonathan Puente-Rivera, and Macrina Beatriz Silva-Cázares. 2022. "Molecular Pathways Related to Sulforaphane as Adjuvant Treatment: A Nanomedicine Perspective in Breast Cancer" Medicina 58, no. 10: 1377. https://doi.org/10.3390/medicina58101377