
An Evaluation of the Anti-Inflammatory Effects of a Thai Traditional Polyherbal Recipe TPDM6315 in LPS-Induced RAW264.7 Macrophages and TNF-α-Induced 3T3-L1 Adipocytes

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Materials
2.3. Preparation of the Extracts
2.3.1. Ethanolic Extract
2.3.2. Aqueous Extract
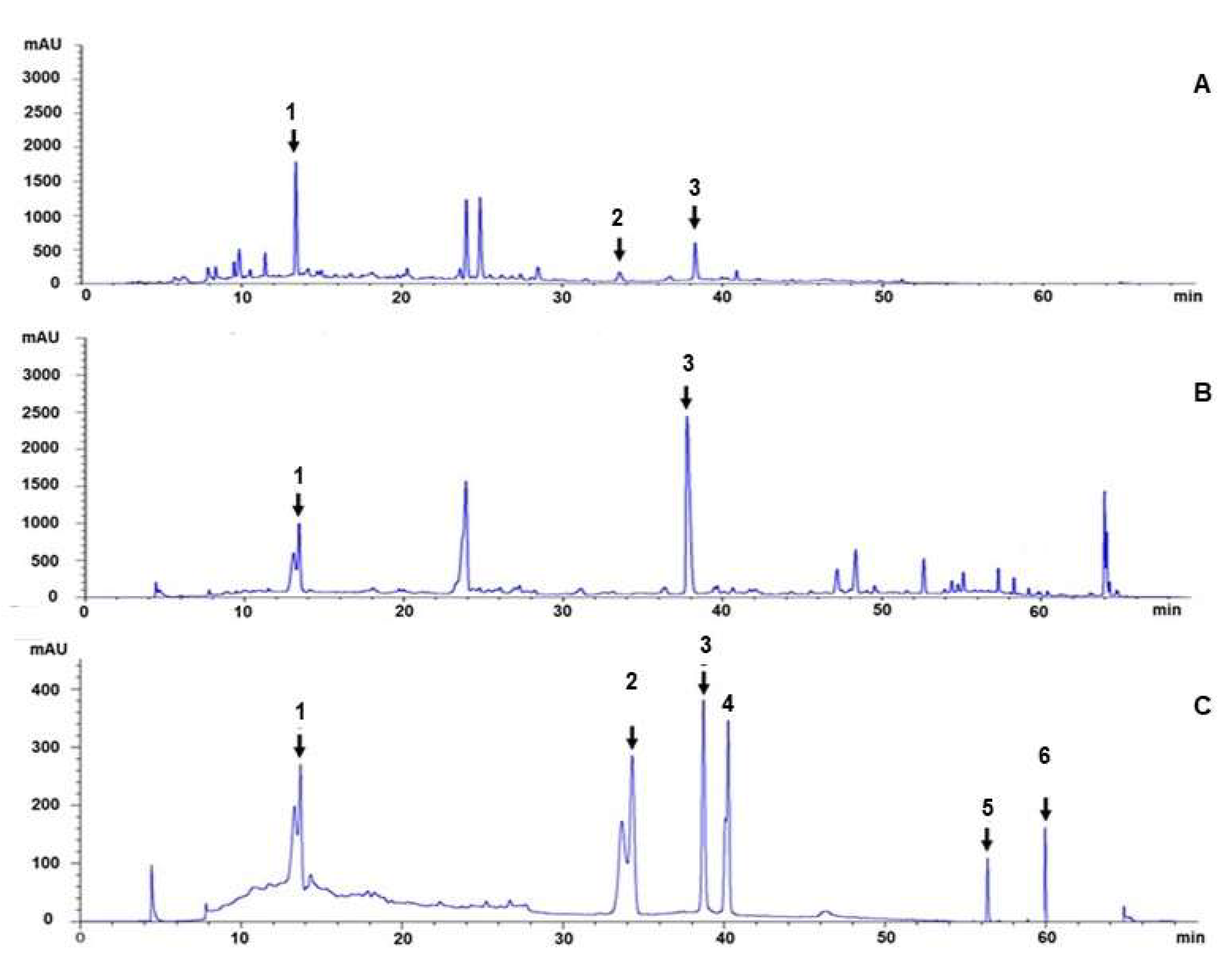
2.4. HPLC Chromatogram
2.5. Evaluation of Anti-Inflammatory Activities of the Extracts in RAW264.7 Macrophages
2.6. The Effects of the Extracts on 3T3-L1 Adipocytes
2.6.1. Cell Culture and Differentiation of 3T3-L1 Adipocytes
2.6.2. Determination of the Effect of Extracts on Lipid Accumulation in 3T3-L1 Adipocytes
2.6.3. Effect of the Extracts on TNF-α-Induced 3T3-L1 Adipocytes
2.7. qPCR Analysis in LPS-Induced RAW264.7 Macrophages and TNF-α-Induced 3T3-L1 Adipocytes
2.8. Statistical Analysis
3. Results
3.1. Characterization of the Recipe Extracts
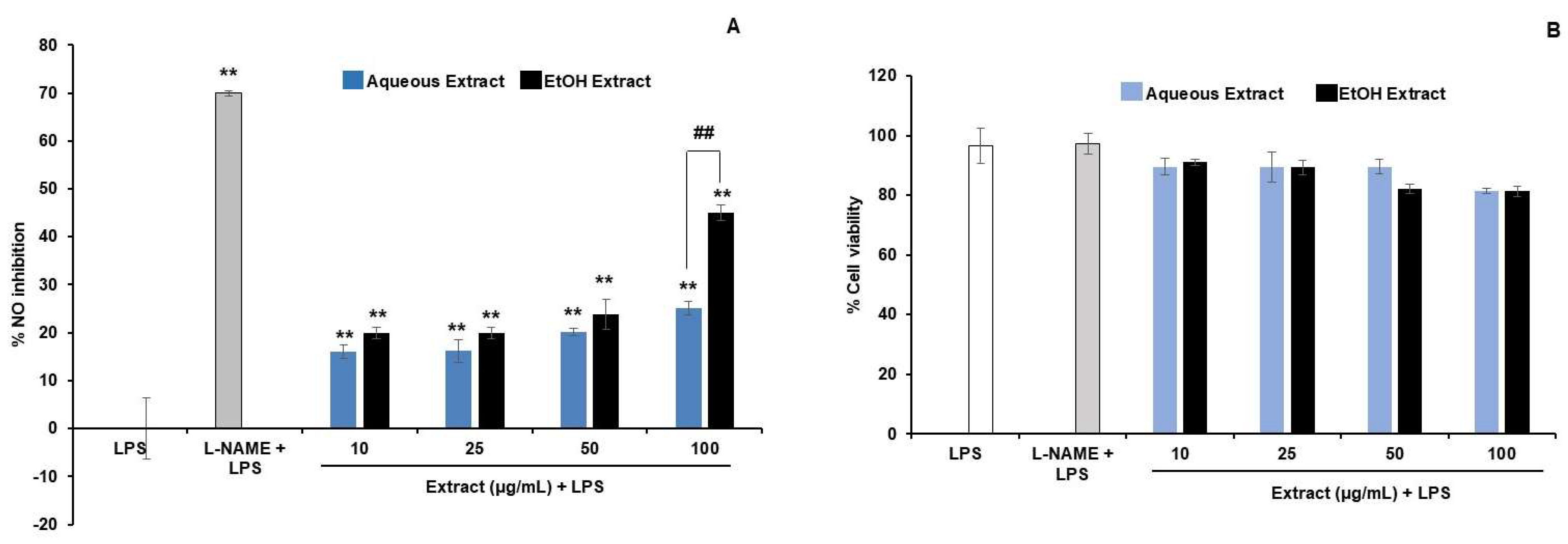
3.2. Anti-Inflammatory Effects on RAW264.7 Macrophages
3.2.1. NO Inhibitory Activity of TPDM6315 Extracts
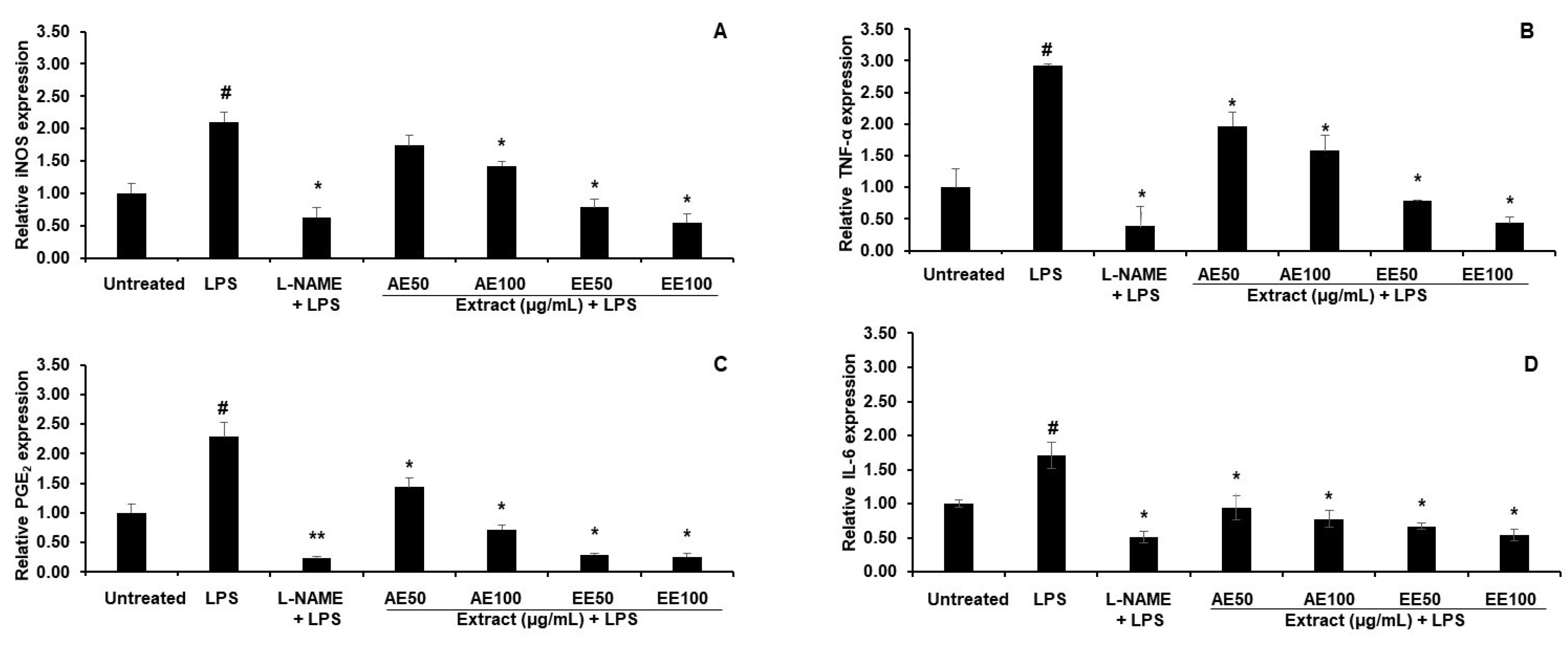
3.2.2. Effect of TPDM6315 Extracts on Gene Expression in LPS-Induced RAW264.7 Macrophages
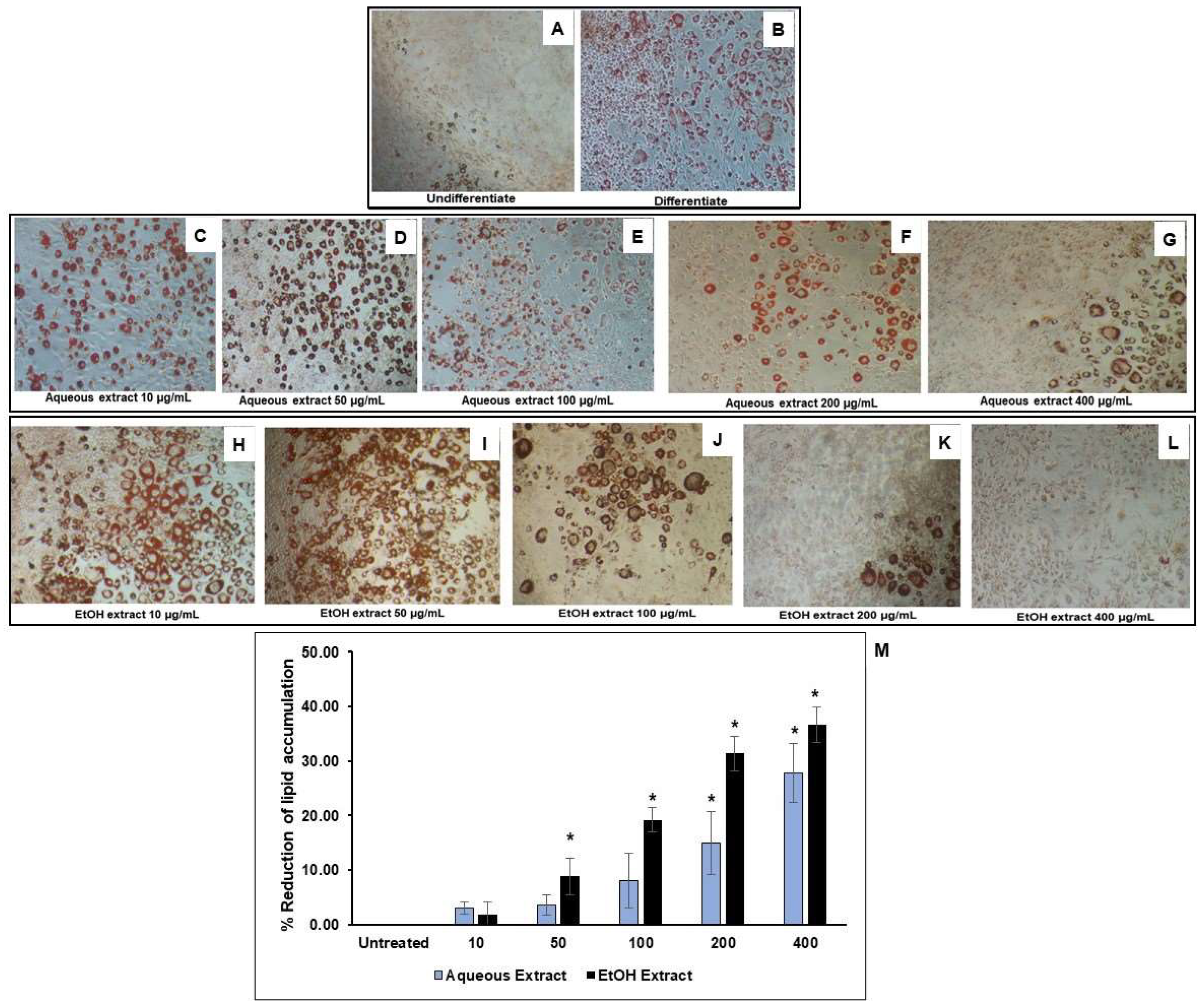
3.3. Reduction of Lipid Accumulation in 3T3-L1-Adipocytes
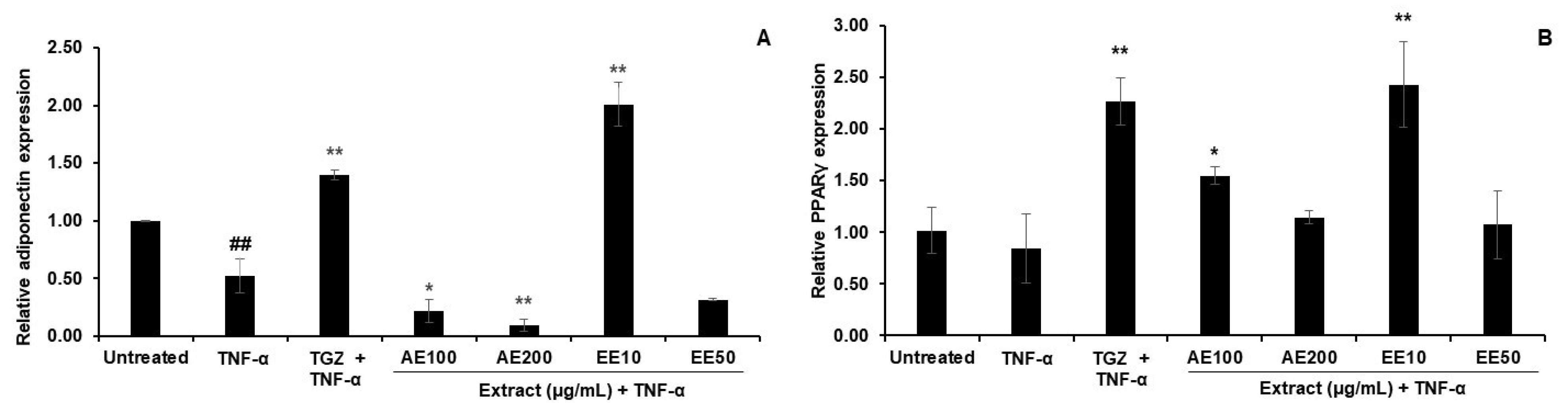
3.4. Effects of TPDM6315 Extracts on TNF-α-Induced 3T3-L1 Adipocytes
3.4.1. Effect on Gene Expression
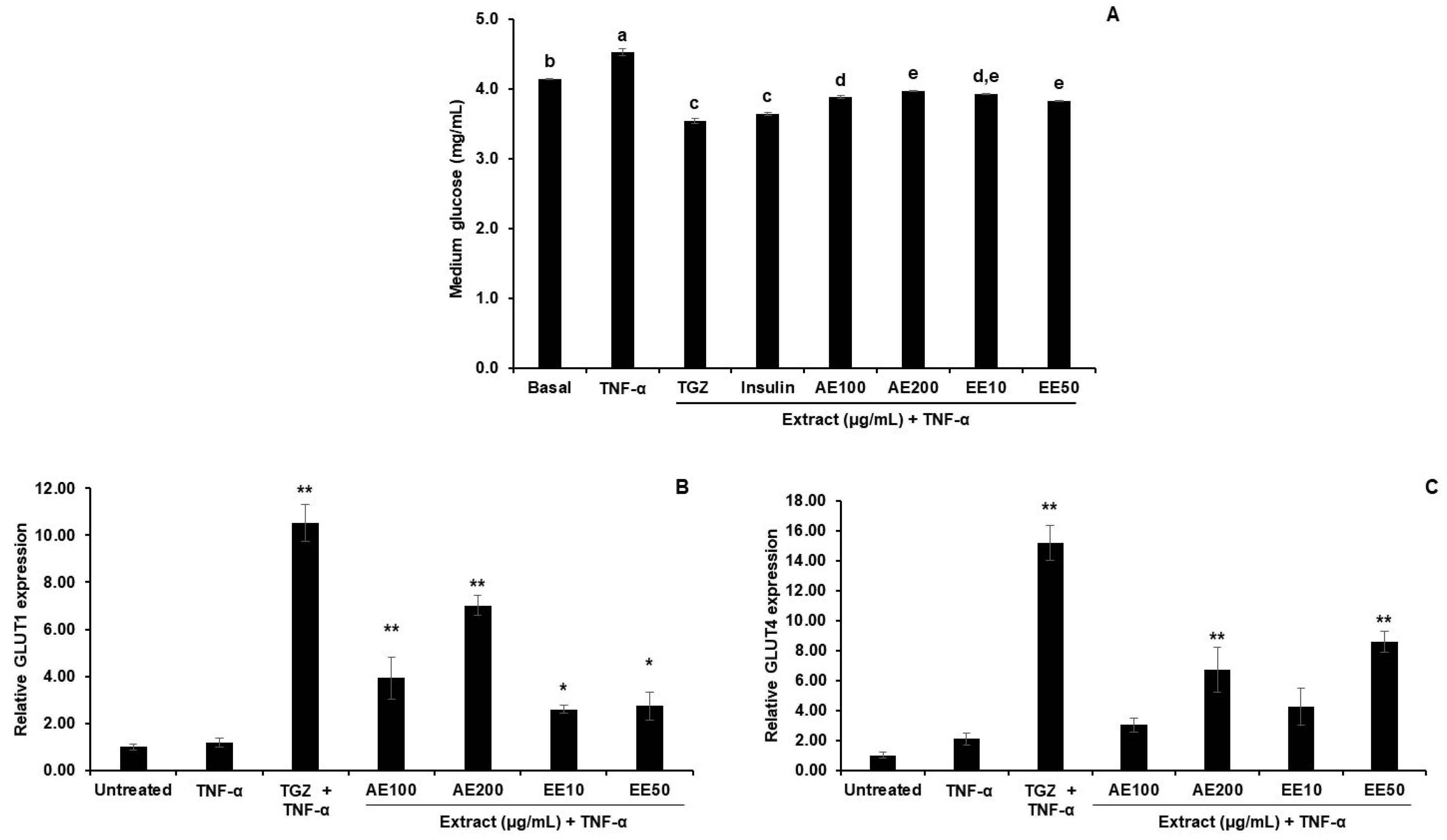
3.4.2. Effect of TPDM6315 Extracts on Glucose Uptake in TNF-α-Induced 3T3-L1 Adipocytes
4. Discussion
4.1. Characterization of the Extracts
4.2. Anti-Inflammation Effect of TPDM6315 Extracts
4.3. Effect of AE and EE on Lipid Accumulation of 3T3-L1 Adipocyte
4.4. Effect of AE and EE on TNF-α-Induced 3T3-L1 Adipocytes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zampronio, A.R.; Soares, D.M.; Souza, G.E. Central mediators involved in the febrile response: Effects of antipyretic drugs. Temperature 2015, 2, 506–521. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.E.; Schmidt, H.; Lai, B.; Ge, K. Transcriptional and epigenomic regulation of adipogenesis. Mol. Cell. Biol. 2019, 39, e00601-18. [Google Scholar] [CrossRef] [Green Version]
- da Cruz Nascimento, S.S.; Carvalho de Queiroz, J.L.; Fernandes de Medeiros, A.; de França Nunes, A.C.; Piuvezam, G.; Lima Maciel, B.L.; Souza Passos, T.; Morais, A.H.D.A. Anti-inflammatory agents as modulators of the inflammation in adipose tissue: A systematic review. PLoS ONE 2022, 17, e0273942. [Google Scholar] [CrossRef]
- Stephens, J.M.; Lee, J.; Pilch, P.F. Tumor necrosis factor-alpha-induced insulin resistance in 3T3-L1 adipocytes is accompanied by a loss of insulin receptor substrate-1 and GLUT4 expression without a loss of insulin receptor-mediated signal transduction. J. Biol. Chem. 1997, 272, 971–976. [Google Scholar] [CrossRef] [Green Version]
- Budluang, P.; Pitchakarn, P.; Ting, P.; Temviriyanukul, P.; Wongnoppawich, A.; Imsumrna, A. Anti-inflammatory and anti-insulin resistance activities of aqueous extract from Anoectochilus burmannicus. Food Sci. Nutr. 2017, 5, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Chang, W.C.; Yang, H.J.; Wu, J.S.B.; Shen, S.C. Gallic acid alleviates hypertriglyceridemia and fat accumulation via modulating glycolysis and lipolysis pathways in perirenal adipose tissues of rats fed a high-fructose diet. Int. J. Mol. Sci. 2018, 19, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.Q.; Song, Y.L.; Zhu, Z.X.; Su, C.; Zhang, X.; Wang, J.; Shi, S.P.; Tu, P.F. Anti-inflammatory dimeric furanocoumarins from the roots of Angelica dahurica. Fitoterapia 2015, 105, 187–193. [Google Scholar] [CrossRef]
- Lu, X.; Yuan, Z.Y.; Yan, X.J.; Lei, F.; Jiang, J.F.; Yu, X.; Yang, X.W.; Xing, D.M.; Du, L.J. Effects of Angelica dahurica on obesity and fatty liver in mice. Chin. J. Nat. Med. 2016, 14, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.S.; Hong, Y.; Lee, H.H.; Ryu, B.; Cho, Y.W.; Kim, N.J.; Jang, D.S.; Lee, K.T. Fulgidic Acid Isolated from the rhizomes of Cyperus rotundus suppresses LPS-Induced iNOS, COX-2, TNF-α, and IL-6 expression by AP-1 inactivation in RAW264.7 macrophages. Biol. Pharm. Bull. 2015, 38, 1081–1086. [Google Scholar] [CrossRef] [Green Version]
- Wongchum, N.; Dechakhamphu, A.; Panya, P.; Pinlaor, S.; Pinmongkhonkul, S.; Tanomtong, A. Hydroethanolic Cyperus rotundus L. extract exhibits antiobesity property and increases lifespan expectancy in Drosophila melanogaster fed a high-fat diet. J. Herbmed. Pharmacol. 2022, 11, 296–304. [Google Scholar] [CrossRef]
- Majeed, M.; Nagabhushanam, K.; Bhat, B.; Ansari, M.; Pandey, A.; Bani, S.; Mundkur, L. The anti-obesity potential of Cyperus rotundus extract containing piceatannol, scirpusin A and scirpusin B from rhizomes: Preclinical and clinical evaluations. Diabetes Metab. Syndr. Obes. Targets Ther. 2022, 15, 369–382. [Google Scholar] [CrossRef]
- Reanmongkol, W.; Subhadhirasakul, S.; Bouking, P. Antinociceptive and antipyretic activities of extracts and fractions from Dracaena loureiri in experimental animals. Songklanakarin J. Sci. Technol. 2003, 25, 467–476. [Google Scholar]
- Jaijoy, K.; Soonthornchareonnon, N.; Panthong, A.; Sireeratawong, S. Anti-inflammatory and analgesic activities of the water extract from the fruit of Phyllanthus emblica Linn. Int. J. Appl. Res. Nat. Prod. 2010, 3, 28–35. [Google Scholar]
- Balusamy, S.R.; Veerappan, K.; Ranjan, A.; Kim, Y.-J.; Chellappan, D.K.; Dua, K.; Lee, J.; Perumalsamy, H. Phyllanthus emblica fruit extract attenuates lipid metabolism in 3T3-L1 adipocytes via activating apoptosis mediated cell death. Phytomedicine 2019, 66, 153129. [Google Scholar] [CrossRef]
- Kumar, R.; Gupta, Y.K.; Singh, S.; Raj, A. Anti-inflammatory effect of Picrorhiza kurroa in experimental models of inflammation. Planta Med. 2016, 82, 1403–1409. [Google Scholar] [CrossRef]
- Khandekar, S.; Pansare, T.; Pachpor, A.G.; Maurya, S. Role of katuka (Picrorhiza kurroa Royle ex Benth.) in obesity W.S.R to Ayurvedic and modern aspect: A review. Int. J. Herb. Med. 2019, 7, 31–35. [Google Scholar]
- Deb, P.K.; Ghosh, R.; Chakraverty, R.; Debnath, R.; Das, L.; Bhakta, T. Phytochemical and pharmacological evaluation of fruits of Solanum indicum L. Int. J. Pharm. Sci. Rev. Res. 2014, 25, 28–32. [Google Scholar]
- Epoh, N.J.; Dongmo, O.L.M.; Mache, R.A.; Telefo, P.B.; Tchouanguep, F.M. Evaluation of anti-obesity effect of aqueous extract of Solanum indicum L. fruits on high fat diet-induced obese rat. Eur. J. Pharm. Med. Res. 2020, 7, 118–128. [Google Scholar]
- Ganesan, K.; Ramasamy, M.; Gani, S.B. Antihyperlipideamic effect of Solanum trilobatum L. leaves extract on streptozotocin induced diabetic rats. Asian J. Biomed. Pharm. Sci. 2013, 3, 51–57. [Google Scholar]
- Jayesh, K.; Karishma, R.; Vysakh, A.; Gopika, P.; Latha, M.S. Terminalia bellirica (Gaertn) Roxb fruit exerts anti-inflammatory effect via regulating arachidonic acid pathway and pro-inflammatory cytokines in lipopolysaccharide-induced RAW 264.7 macrophages. Inflammopharmacology 2020, 28, 265–274. [Google Scholar]
- Das, C.M.S.; Devi, S.G. Antiobesity activity of ethanolic extract of fruits of Terminalia bellirica on atherogenic diet induced obesity in experimental rats. J. Chem. Pharm. Res. 2016, 8, 191–197. [Google Scholar]
- Shendge, A.K.; Sarkar, R.; Mandal, N. Potent anti-inflammatory Terminalia chebula fruit showed in vitro anticancer activity on lung and breast carcinoma cells through the regulation of Bax/Bcl-2 and caspase-cascade pathways. J. Food Biochem. 2020, 44, e13521. [Google Scholar] [CrossRef]
- Subramanian, G.; Shanmugamprema, D.; Subramani, R.; Muthuswamy, K.; Ponnusamy, V.; Tankay, K.; Velusamy, T.; Krishnan, V.; Subramaniam, S. Anti-obesity effect of T. chebula fruit extract on high fat diet induced obese mice: A possible alternative therapy. Mol. Nutr. Food Res. 2021, 65, 2001224. [Google Scholar] [CrossRef] [PubMed]
- Yokozawa, T.; Wang, T.S.; Chen, C.P.; Hattori, M. Inhibition of nitric oxide release by an aqueous extract of Tinospora tuberculata. Phytother. Res. 2000, 14, 51–53. [Google Scholar] [CrossRef]
- Li, F.; Nitteranon, V.; Tang, X.; Liang, J.; Zhang, G.; Parkin, K.L.; Hu, Q. In vitro antioxidant and anti-inflammatory activities of 1-dehydro-[6]-gingerdione, 6-shogaol, 6-dehydroshogaol and hexahydrocurcumin. Food Chem. 2012, 135, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.H.; Fang, F.; Kang, I. Ginger (Zingiber officinale) attenuates obesity and adipose tissue remodeling in high-fat diet-fed C57BL/6 Mice. Int. J. Environ. Res. Public Health 2021, 18, 631. [Google Scholar] [CrossRef]
- Nualkaew, S. Applied Thai Traditional Pharmacy, 1st ed.; KKU Printing House: Khon Kaen, Thailand, 2020; pp. 178–229. [Google Scholar]
- Srisopon, S.; Burana-Osot, J.; Sotanaphun, U. Botanical identification of Chan-Khao and Chan-Thana by thin-layer chromatography. Thai Pharm. Health Sci. J. 2015, 10, 19–24. [Google Scholar]
- Sabuhom, P.; Subin, P.; Luecha, P.; Nualkaew, S.; Nualkaew, N. Effects of plant part substitution in a Thai traditional recipe on α-glucosidase inhibition. Trop. J. Nat. Prod. Res. 2023, 7, 2919–2925. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Waltenberger, B.; Schuster, D.; Paramapojn, S.; Gritsanapan, W.; Wolber, G.; Rollinger, J.M.; Stuppner, H. Predicting cyclooxygenase inhibition by three-dimensional pharmacophoric profiling. Part II: Identification of enzyme inhibitors from Prasaplai, a Thai traditional medicine. Phytomedicine 2011, 18, 119–133. [Google Scholar] [CrossRef] [Green Version]
- Vannabhum, M.; Poopong, S.; Wongwananuruk, T.; Nimmannit, A.; Suwannatrai, U.; Dangrat, C.; Apichartvorakit, A.; Booranasubkajorn, S.; Laohapand, T.; Akaraserenont, P. The efficacy of Thai herbal Prasaplai formula for treatment of primary dysmenorrhea: A short-term randomized controlled trial. Evid. Based Complement. Altern. Med. 2016, 2016, 2096797. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Kishimoto, Y.; Sasaki, M.; Sato, A.; Kamiya, T.; Kondo, K.; Iida, K. Terminalia bellirica (Gaertn.) Roxb. extract and gallic acid attenuate LPS-induced inflammation and oxidative stress via MAPK/NF- κ B and Akt/AMPK/Nrf2 pathways. Oxid. Med. Cell. Longev. 2018, 1, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Zhang, Y.; Tang, C.; Hou, Y.; Ai, X.; Chen, X.; Zhang, Y.; Wang, X.; Meng, X. Gallic acid: Pharmacological activities and molecular mechanisms involved in inflammation-related diseases. Biomed. Pharmacother. 2020, 133, 110985. [Google Scholar] [CrossRef]
- Behera, P.K.; Devi, S.; Mittal, N. Therapeutic potential of gallic acid in obesity: Considerable shift! Obes. Med. 2023, 37, 100473. [Google Scholar] [CrossRef]
- Reddy, D.B.; Reddanna, P. Chebulagic acid (CA) attenuates LPS-induced inflammation by suppressing NF-κB and MAPK activation in RAW 264.7 macrophages. Biochem. Biophys. Res. Commun. 2009, 381, 112–117. [Google Scholar] [CrossRef]
- Shyni, G.L.; Kavitha, S.; Indu, S.; Das Arya, A.; Anusree, S.S.; Vineetha, V.P.; Vandana, S.; Sundaresan, A.; Raghu, K.G. Chebulagic acid from Terminalia chebula enhances insulin mediated glucose uptake in 3T3-L1 adipocytes via PPARγ signaling pathway. Biofactors 2014, 40, 646–657. [Google Scholar]
- Gupta, A.; Kumar, R.; Ganguly, R.; Singh, A.K.; Rana, H.K.; Pandey, A.K. Antioxidant, anti-inflammatory and hepatoprotective activities of Terminalia bellirica and its bioactive component ellagic acid against diclofenac induced oxidative stress and hepatotoxicity. Toxicol. Rep. 2021, 8, 44–52. [Google Scholar] [CrossRef]
- Shiojima, Y.; Takahashi, M.; Kikuchi, M.; Akanuma, M. Effect of ellagic acid on body fat and triglyceride reduction in healthy overweight volunteers: A randomized, double-blind, placebo-controlled parallel group study. J. Funct. Food Health Dis. 2020, 10, 180–194. [Google Scholar] [CrossRef]
- Ghadimi, M.; Foroughi, F.; Hashemipour, S.; Nooshabadi, M.R.; Ahmadi, M.H.; Yari, M.G.; Kavianpour, M.; Haghighian, H.K. Decreased insulin resistance in diabetic patients by influencing Sirtuin1 and Fetuin-A following supplementation with ellagic acid: A randomized controlled trial. Diabetol. Metab. Syndr. 2021, 13, 16. [Google Scholar] [CrossRef]
- Sharma, K.; Kumar, S.; Prakash, R.; Khanka, S.; Mishra, T.; Rathur, R.; Biswas, A.; Verma, S.K.; Bhatta, R.S.; Narender, T.; et al. Chebulinic acid alleviates LPS-induced inflammatory bone loss by targeting the crosstalk between reactive oxygen species/NFκB signaling in osteoblast cells. Free Radic. Biol. Med. 2023, 194, 99–113. [Google Scholar] [CrossRef]
- Kim, J.; Ahn, D.; Chung, S.J. Chebulinic acid suppresses adipogenesis in 3T3-L1 preadipocytes by inhibiting PPP1CB activity. Int. J. Mol. Sci. 2022, 23, 865. [Google Scholar] [CrossRef]
- Yoon, S.-Y.; Kang, H.J.; Ahn, D.; Hwang, J.Y.; Kwon, S.J.; Chung, S.J. Identification of chebulinic acid as a dual targeting inhibitor of protein tyrosine phosphatases. Bioorg. Chem. 2019, 90, 103087. [Google Scholar] [CrossRef]
- Liang, W.H.; Chang, T.W.; Charng, Y.C. Effects of drying methods on contents of bioactive compounds and antioxidant activities of Angelica dahurica. Food Sci. Biotechnol. 2018, 27, 1085–1092. [Google Scholar] [CrossRef]
- Lee, T.Y.; Lee, K.C.; Chen, S.Y.; Chang, H.H. 6-Gingerol inhibits ROS and iNOS through the suppression of PKC-alpha and NF-kappaB pathways in lipopolysaccharide-stimulated mouse macrophages. Biochem. Biophys. Res. Commun. 2009, 382, 134–139. [Google Scholar] [CrossRef]
- Gunawan, S.; Munika, E.; Wulandari, E.T.; Ferdinal, F.; Purwaningsih, E.H.; Wuyung, P.E.; Louisa, M.; Soetikno, V. 6-gingerol ameliorates weight gain and insulin resistance in metabolic syndrome rats by regulating adipocytokines. Saudi Pharm. J. 2023, 31, 351–358. [Google Scholar] [CrossRef]
- Bachar, O.; Adner, M.; Uddman, R.; Cardell, L.O. Toll-like receptor stimulation induces airway hyper-responsiveness to bradykinin, an effect mediated by JNK and NF-kappa B signaling pathways. Eur. J. Immunol. 2004, 34, 1196–1207. [Google Scholar] [CrossRef]
- Aronoff, D.M.; Neilson, E.G. Antipyretics mechanisms of action and clinical use in fever suppression. Am. J. Med. 2001, 111, 304–315. [Google Scholar] [CrossRef]
- Jin, L.; Shi, G.; Ning, G.; Li, X.; Zhang, Z. Andrographolide attenuates tumor necrosis factor alpha induced insulin resistance in 3T3-L1 adipocytes. Mol. Cell. Endocrinol. 2011, 332, 134–139. [Google Scholar] [CrossRef]
- Baek, S.H.; Park, T.; Kang, M.G.; Park, D. Anti-inflammatory activity and ROS regulation effect of sinapaldehyde in LPS-stimulated RAW 264.7 macrophages. Molecules 2020, 25, 4089. [Google Scholar] [CrossRef]
- Guan, F.; Wang, H.; Shan, Y.; Chen, Y.; Wang, M.; Wang, Q.; Yin, M.; Zhao, Y.; Feng, X.; Zhang, J. Inhibition of COX-2 and PGE2 in LPS-stimulated RAW264.7 cells by lonimacranthoide VI, a chlorogenic acid ester saponin. Biomed. Rep. 2014, 2, 760–764. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.I.; Pero, R.S.; Scheck, A.C.; Romanovsky, A.A. Prostaglandin E2-synthesizing enzymes in fever: Differential transcriptional regulation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R1104–R1117. [Google Scholar] [CrossRef] [Green Version]
- Roth, J.; Blatteis, C.M. Mechanisms of fever production and lysis: Lessons from experimental LPS fever. Compr. Physiol. 2014, 4, 1563–1604. [Google Scholar] [CrossRef]
- BenSaad, L.A.; Kim, K.H.; Quah, C.C.; Kim, W.R.; Shahimi, M. Anti-inflammatory potential of ellagic acid, gallic acid and punicalagin A and B isolated from Punica granatum. BMC Complement. Altern. Med. 2017, 17, 47. [Google Scholar] [CrossRef] [Green Version]
- Juckmeta, T.; Itharat, A. Anti- inflammatory and antioxidant activities of Thai traditional remedy called “Ya-ha-rak”. J. Health Res. 2012, 26, 205–210. [Google Scholar]
- Prommee, N.; Itharat, A.; Thongdeeying, P.; Makchuchit, S.; Pipatrattanaseree, W.; Tasanarong, A.; Ooraikul, B.; Davies, N.M. Exploring in vitro anti-proliferative and anti-inflammatory activities of Prasachandaeng remedy, and its bioactive compounds. BMC Complement. Med. Ther. 2022, 22, 217. [Google Scholar] [CrossRef]
- Sireeratawong, S.; Khonsung, P.; Piyabhan, P.; Nanna, U.; Soonthornchareonnon, N.; Jaijoy, K. Anti-inflammatory and anti-ulcerogenic activities of Chantaleela recipe. Afr. J. Tradit. Complement. Altern. Med. 2012, 9, 485–494. [Google Scholar] [CrossRef]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar] [CrossRef]
- Cave, E.; Crowther, N.J. The Use of 3T3-L1 Murine Preadipocytes as a Model of Adipogenesis. In Pre-Clinical Models: Techniques and Protocols, Methods in Molecular Biology; Guest Paul, C., Ed.; Springer Science Business Media: Berlin/Heidelberg, Germany, 2019; p. 1916. [Google Scholar] [CrossRef]
- Wong-A-Nan, N.; Inthanon, K.; Saiai, A.; Inta, A.; Nimlamool, W.; Chomdej, S.; Kittakoop, P.; Wongkham, W. Lipogenesis inhibition and adipogenesis regulation via PPARc pathway in 3T3-L1 cells by Zingiber cassumunar Roxb. rhizome extracts. Egypt. J. Basic Appl. Sci. 2018, 5, 289–297. [Google Scholar]
- Saraphanchotiwitthaya, A.; Sripalakit, P. Jatupalathika herbal formula inhibits lipid accumulation and induces lipolysis in 3T3-L1 adipocytes. Sci. Asia 2022, 48, 1–7. [Google Scholar] [CrossRef]
- Wang, L.; Li, L.; Ran, X.; Long, M.; Zhang, M.; Tao, Y.; Luo, X.; Wang, Y.; Ma, X.; Halmurati, U.; et al. Ellagic acid reduces adipogenesis through inhibition of differentiation-prevention of the induction of Rb phosphorylation in 3T3-L1 adipocytes. Evid. Based Complement. Altern. Med. 2013, 2013, 287534. [Google Scholar] [CrossRef] [Green Version]
- Tzeng, T.F.; Liu, I.M. 6-Gingerol prevents adipogenesis and the accumulation of cytoplasmic lipid droplets in 3T3-L1 cells. Phytomedicine 2013, 20, 481–487. [Google Scholar] [CrossRef]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic adipose tissue inflammation linking obesity to insulin resistance and type 2 diabetes. Front. Physiol. 2020, 10, 1607. [Google Scholar] [CrossRef]
- Yuan, M.; Konstantopoulos, N.; Lee, J.; Hansen, L.; Li, Z.W.; Karin, M.; Shoelson, S.E. Reversal of Obesity- and Diet-Induced Insulin Resistance with Salicylates or Targeted Disruption of Ikkbeta. Science 2001, 293, 1673–1677. [Google Scholar] [CrossRef]
- Jin, D.; Sun, J.; Huang, J.; He, Y.; Yu, A.; Yu, X.; Yang, Z. TNF-α reduces g0s2 expression and stimulates lipolysis through PPAR-γ inhibition in 3T3-L1 adipocytes. Cytokine 2014, 69, 196–205. [Google Scholar] [CrossRef]
- Xing, H.; Northrop, J.P.; Russell Grove, J.; Kilpatrick, K.E.; Jui-Lan, S.U.; Ringold, G.M. TNFα-mediated inhibition and reversal of adipocyte differentiation is accompanied by suppressed expression of PPARγ without effects on Pref-1 expression. Endocrinology 1997, 138, 2776–2783. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, H.; Hegde, V.; Dubuisson, O.; Gao, Z.; Dhurandhar, N.V.; Ye, J. Interplay of pro- and anti-inflammatory cytokines to determine lipid accretion in adipocytes. Int. J. Obes. 2013, 37, 1490–1498. [Google Scholar] [CrossRef] [Green Version]
- Qiao, L.; Kinney, B.; Schaack, J.; Shao, J. Adiponectin inhibits lipolysis in mouse adipocytes. Diabetes 2011, 60, 1519–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aljada, A.; Garg, R.; Ghanim, H.; Mohanty, P.; Hamouda, W.; Assian, E.; Dandona, P. Nuclear factor-kappaB suppressive and inhibitor-kappaB stimulatory effects of troglitazone in obese patients with type 2 diabetes: Evidence of an antiinflammatory action? J. Clin. Endocrinol. Metab. 2001, 86, 3250–3256. [Google Scholar] [PubMed] [Green Version]
- Małodobra-Mazur, M.; Cierzniak, A.; Ryba, M.; Sozański, T.; Piórecki, N.; Kucharska, A.Z. Increases glucose uptake and the expression of PPARG in insulin-resistant adipocytes. Nutrients 2022, 14, 2307. [Google Scholar] [CrossRef]
- Szalkowski, D.; White-Carrington, S.; Berger, J.; Zhang, B. Antidiabetic thiazolidinediones block the inhibitory effect of tumor necrosis factor-alpha on differentiation, insulin-stimulated glucose uptake, and gene expression in 3T3-L1 cells. Endocrinology 1995, 136, 1474–1481. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Botanical Name | Part | Traditional Use as Anti-Fever * | Anti-Inflammation and Antipyretic | Anti-Obesity |
|---|---|---|---|---|
| Angelica dahurica (Fisch. Ex Hoffm.) Benth. & Hook. f. ex Franch. & Sav. | Root | / | [7] | [8] |
| Cyperus rotundus L. | Rhizome | - | [9] | [10,11] |
| Digitaria ciliaris (Retz.) Koeler | Whole plant | / | - | - |
| Dracaena loureiroi Gagnep. | Stem | / | [12] | - |
| Gymnopetalum chinense (Lour.) Merr. | Whole fruit | / | - | - |
| Gymnopetalum integrifolium Kurz | Root | / | - | - |
| Phyllanthus emblica L | Whole fruit | - | [13] | [14] |
| Picrorhiza kurrooa Royle ex Benth. | Root and rhizome | / | [15] | [16] |
| Santalum spicatum L. | Stem | / | - | - |
| Solanum indicum L. | Root | / | [17] | [18] |
| Solanum trilobatum L. | Root | / | - | [19] |
| Terminalia bellirica (Gaertn.) Roxb. | Whole fruit | / | [20] | [21] |
| Terminalia chebula Retz. | Whole fruit | / | [22] | [23] |
| Tinospora crispa (L.) Miers ex Hook. f. & Thomson | Stem | / | [24] | - |
| Zingiber officinale Roscoe. | Rhizome | / | [25] | [26] |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| iNOS (112 bp) | 5′ CTGCCAGGGTCACAACTTTAC 3′ | 5′ AACAGCTCAGTCCCTTCACC 3′ |
| PGE2 (143 bp) | 5′ TGACAGCCGTGGGTAAAGAC 3′ | 5′ CCAAGGCTGGATGTGTGAGT 3′ |
| TNFα (128 bp) | 5′ GATCGGTCCCCAAAGGGATG 3′ | 5′ TTGCTACGACGTGGGCTAC 3′ |
| IL6 (154 bp) | 5′ GTCCTTCCTACCCCAATTTCCA 3′ | 5′ TAACGCACTAGGTTTGCCGA 3′ |
| Adiponectin (146 bp) | 5′ TGACGACACCAAAAGGGCTC 3′ | 5′ ACCTGCACAAGTTCCCTTGG 3′ |
| GLUT1 (139 bp) | 5′ CAATGGCGGCGGTCCTATAA 3′ | 5′ TGTAACTATGCGTCTCCCGC 3′ |
| GLUT4 (200 bp) | 5′ TCTGACGTAAGGATGGGGAAC 3′ | 5′ TTGTGGGATGGAATCCGGTC 3′ |
| β-Actin (196 bp) | 5′ GACACGAGTTGGTTGGAGCA 3′ | 5′ GCGACCATCCTCCTCTTAGG 3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subin, P.; Sabuhom, P.; Naladta, A.; Luecha, P.; Nualkaew, S.; Nualkaew, N. An Evaluation of the Anti-Inflammatory Effects of a Thai Traditional Polyherbal Recipe TPDM6315 in LPS-Induced RAW264.7 Macrophages and TNF-α-Induced 3T3-L1 Adipocytes. Curr. Issues Mol. Biol. 2023, 45, 4891-4907. https://doi.org/10.3390/cimb45060311
Subin P, Sabuhom P, Naladta A, Luecha P, Nualkaew S, Nualkaew N. An Evaluation of the Anti-Inflammatory Effects of a Thai Traditional Polyherbal Recipe TPDM6315 in LPS-Induced RAW264.7 Macrophages and TNF-α-Induced 3T3-L1 Adipocytes. Current Issues in Molecular Biology. 2023; 45(6):4891-4907. https://doi.org/10.3390/cimb45060311
Chicago/Turabian StyleSubin, Phetpawi, Pattraporn Sabuhom, Alisa Naladta, Prathan Luecha, Somsak Nualkaew, and Natsajee Nualkaew. 2023. "An Evaluation of the Anti-Inflammatory Effects of a Thai Traditional Polyherbal Recipe TPDM6315 in LPS-Induced RAW264.7 Macrophages and TNF-α-Induced 3T3-L1 Adipocytes" Current Issues in Molecular Biology 45, no. 6: 4891-4907. https://doi.org/10.3390/cimb45060311