
Elicitation of Inhibitory Effects for AGE-Induced Oxidative Stress in Rotator Cuff-Derived Cells by Apocynin
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Preparation and Treatment of Human Rotator Cuff-Derived Cells
2.3. Quantitative Real-Time Polymerase Chain Reaction (PCR)
2.4. Cell Viability Assay
2.5. Apoptotic Cells Analysis
2.6. Detection of ROS Production
2.7. Statistical Analysis
3. Results
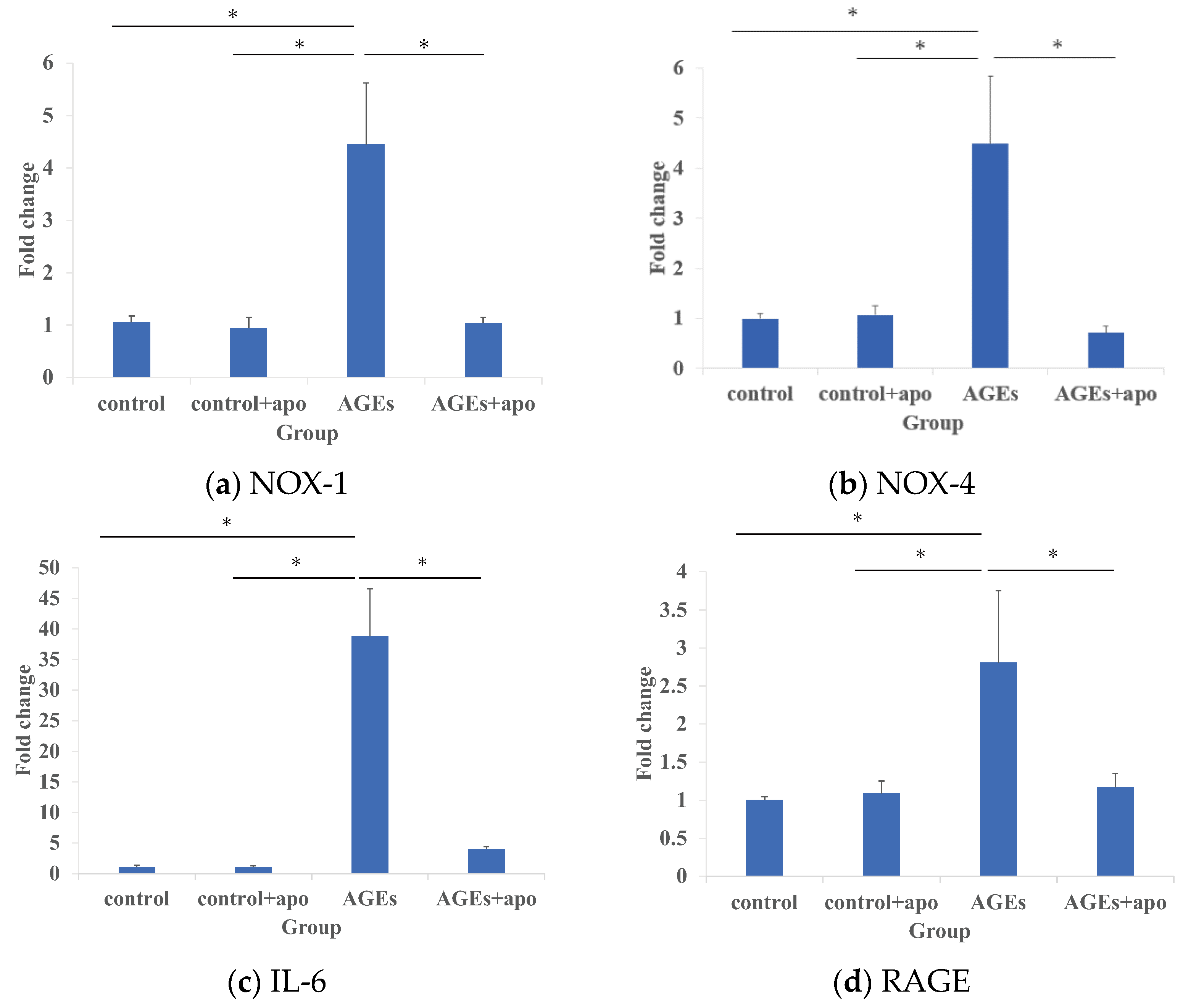
3.1. Modification of AGE-Related Markers in RC-Derived Cells by Apocynin Treatment
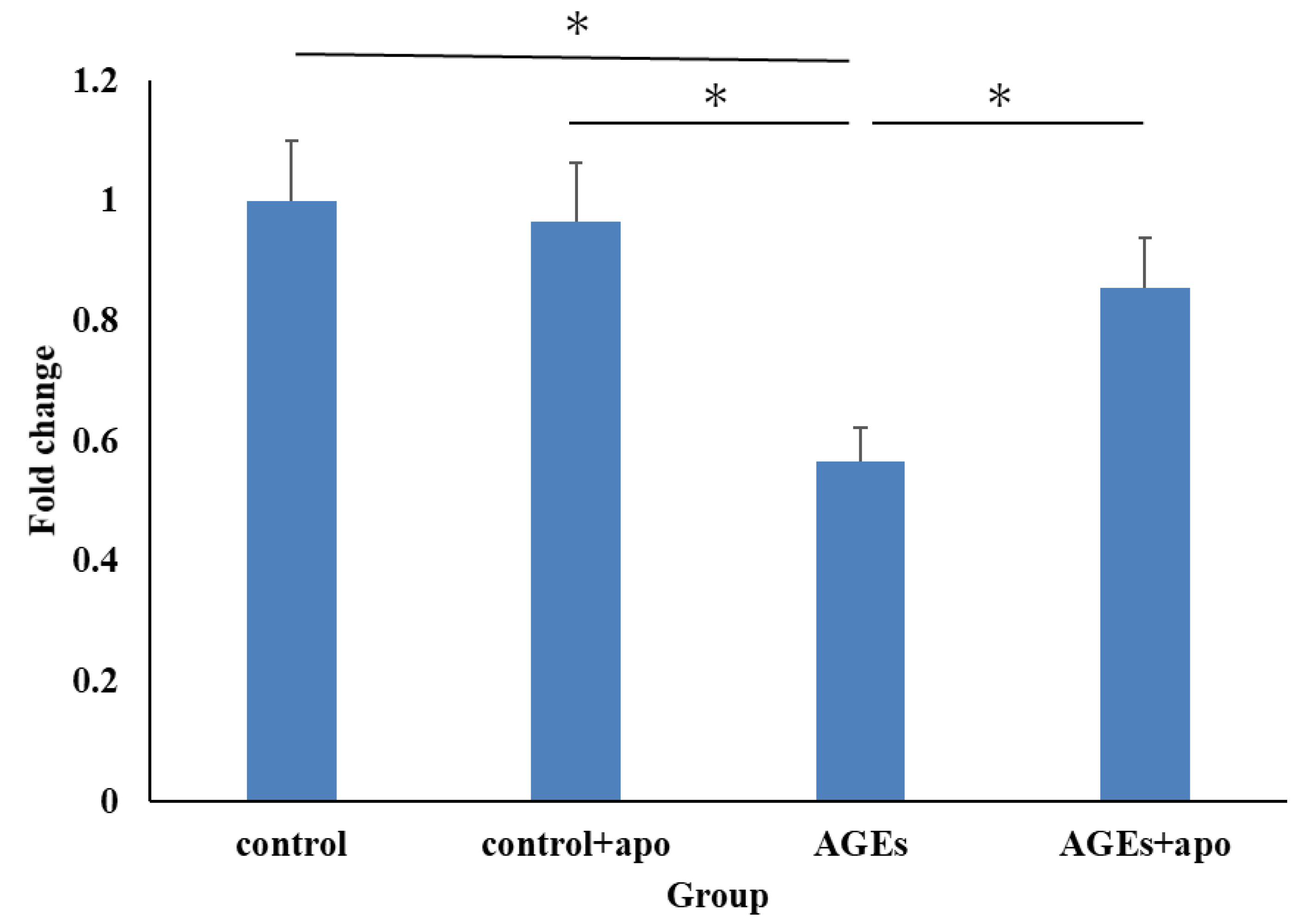
3.2. Apocynin Resulted in Inhibition of Decreased RC-Derived Cell Viability with AGEs Stimulation
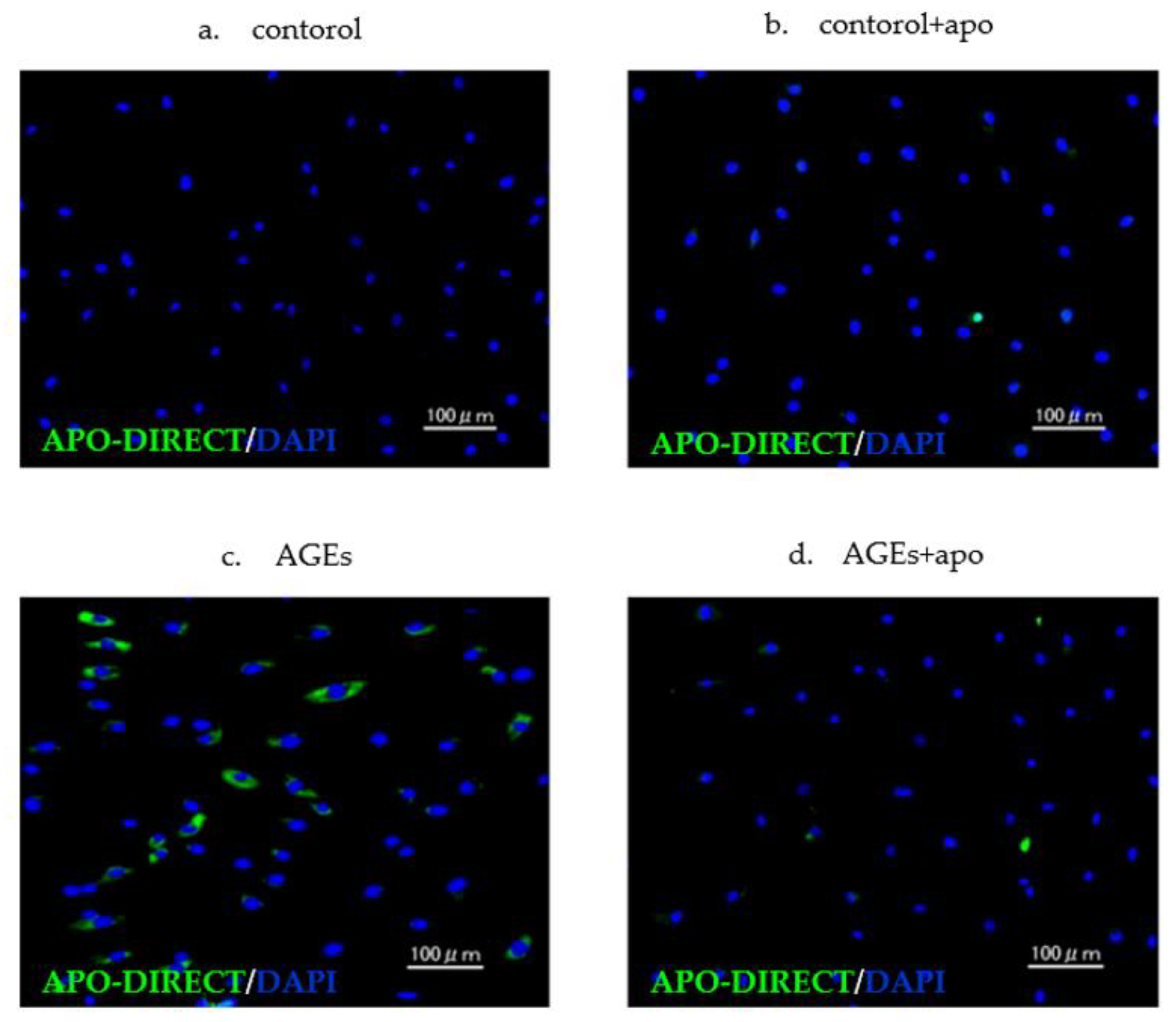
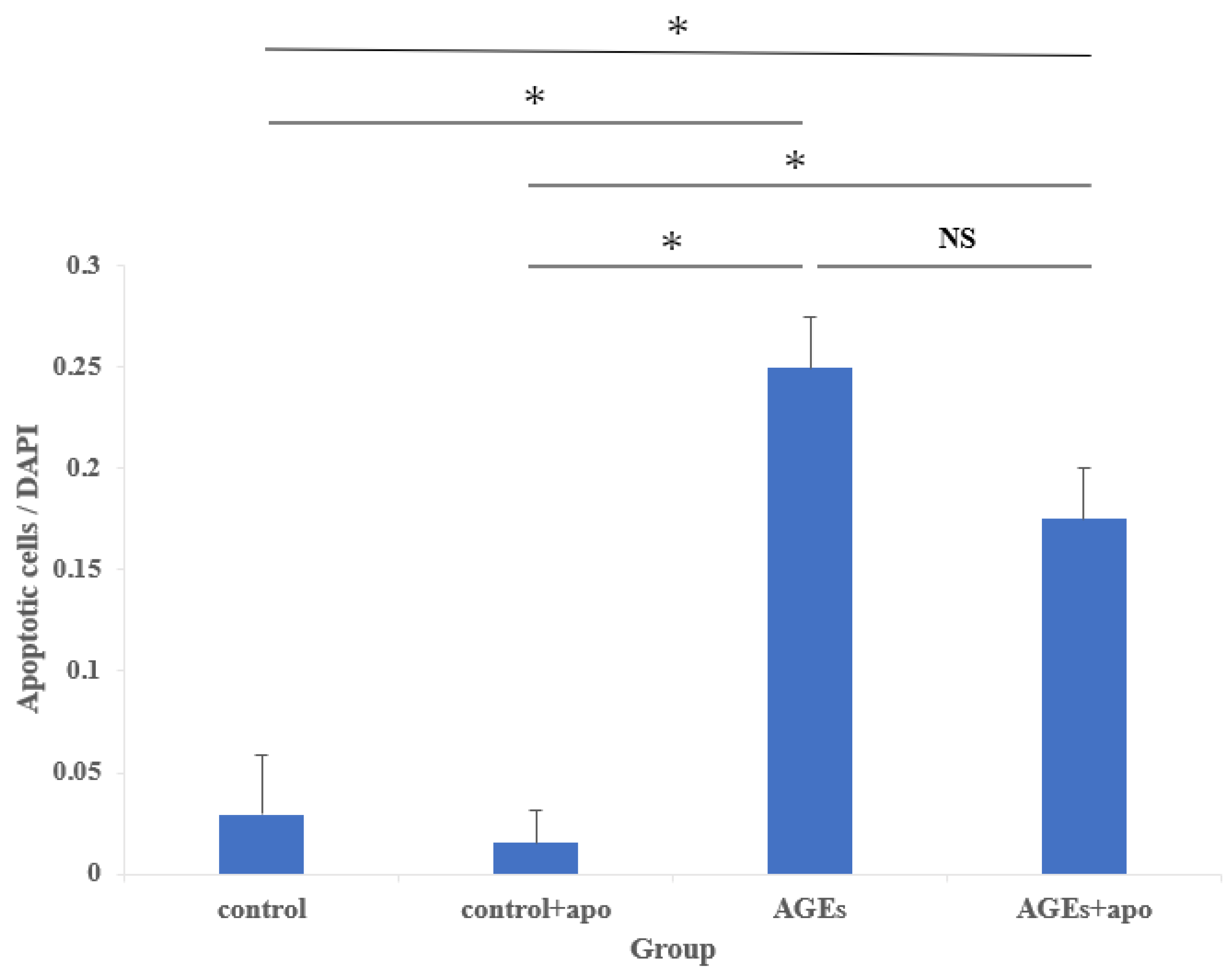
3.3. Cell Damage of AGEs to RC-Derived Cells and Their Inhibition Tendency with Apocynin
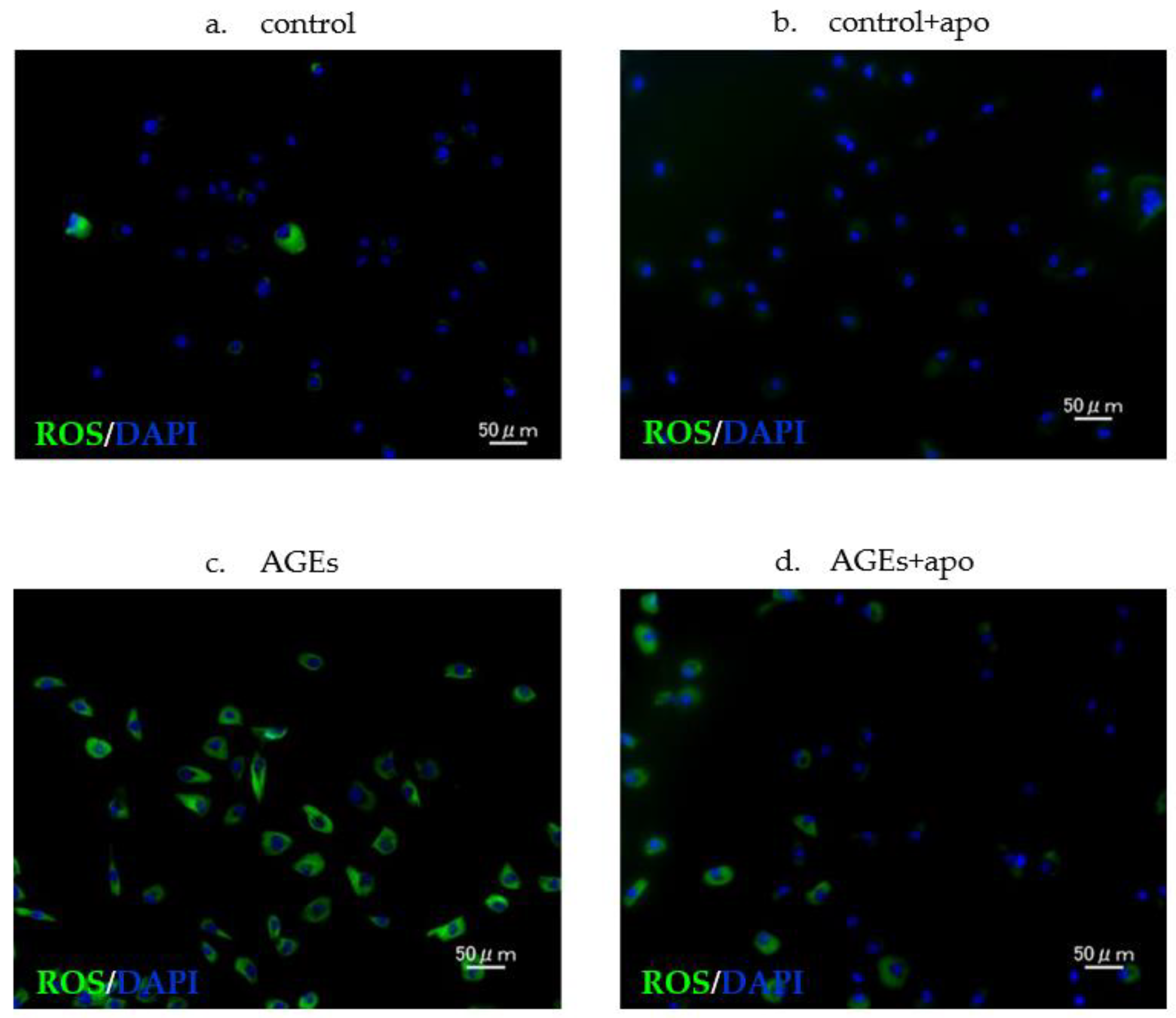
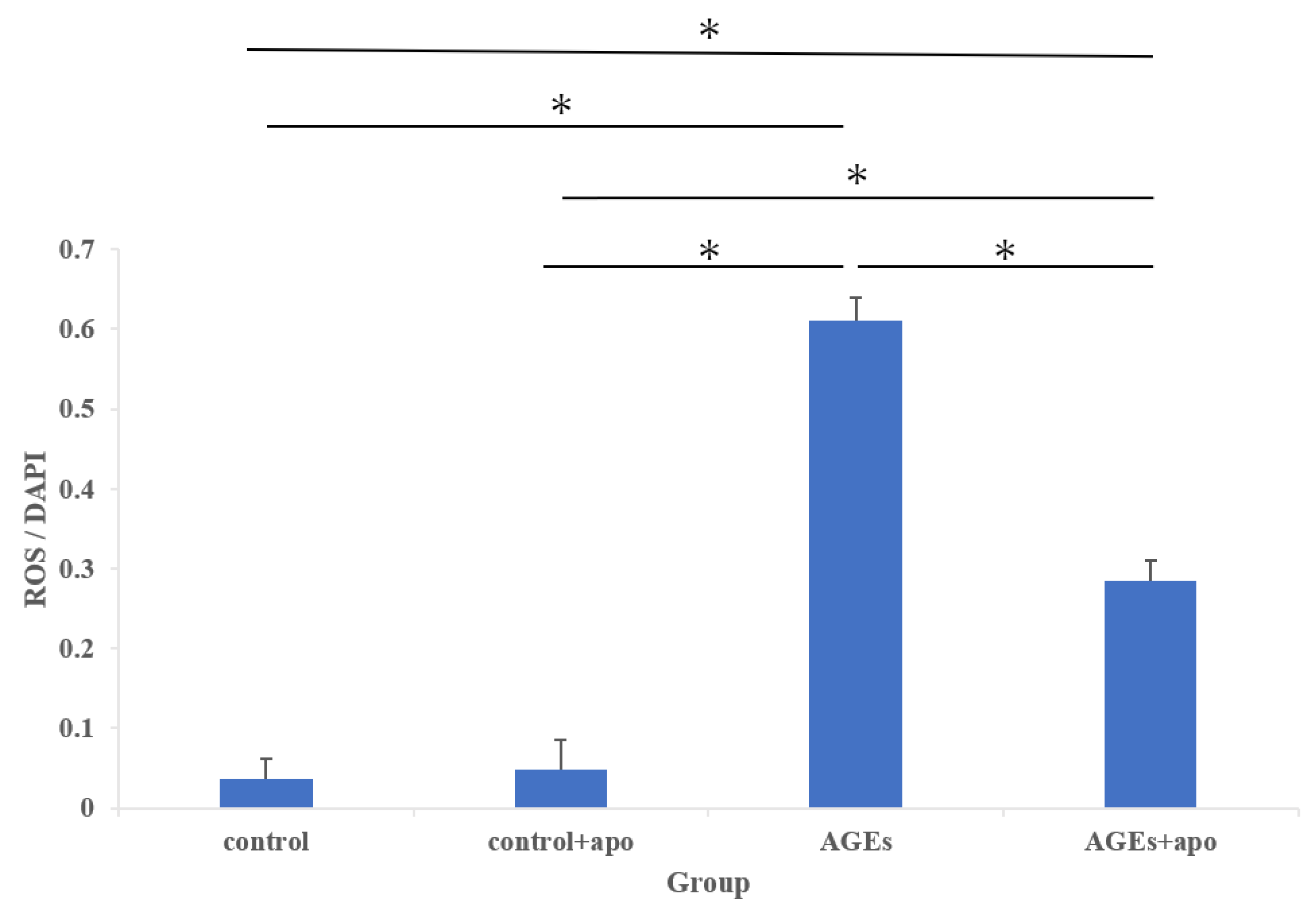
3.4. Downregulation of AGE-Related ROS Generation Was Observed in Apocynin-Treated RC Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamamoto, A.; Takagishi, K.; Osawa, T.; Yanagawa, T.; Nakajima, D.; Shitara, H.; Kobayashi, T. Prevalence and risk factors of a rotator cuff tear in the general population. J. Shoulder Elb. Surg. 2010, 19, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Abboud, J.A.; Kim, J.S. The effect of hypercholesterolemia on rotator cuff disease. Clin. Orthop. Relat. Res. 2010, 468, 1493–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDermott, F.T. Repetition strain injury: A review of current understanding. Med. J. Aust. 1986, 144, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Jahan, H.; Choudhary, M.I. Glycation, carbonyl stress and AGEs inhibitors: A patent review. Expert Opin. Ther. Pat. 2015, 25, 1267–1284. [Google Scholar] [PubMed]
- Stinghen, A.E.; Massy, Z.A.; Vlassara, H.; Striker, G.E.; Boullier, A. Uremic Toxicity of Advanced Glycation End Products in CKD. J. Am. Soc. Nephrol. 2016, 27, 354–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, M.; Marumo, K. Collagen cross-links as a determinant of bone quality: A possible explanation for bone fragility in aging, osteoporosis, and diabetes mellitus. Osteoporos. Int. 2010, 21, 195–214. [Google Scholar] [CrossRef]
- Remigante, A.; Spinelli, S.; Basile, N.; Caruso, D.; Falliti, G.; Dossena, S.; Marino, A.; Morabito, R. Oxidation Stress as a Mechanism of Aging in Human Erythrocytes: Protective Effect of Quercetin. Int. J. Mol. Sci. 2022, 23, 7791. [Google Scholar] [CrossRef]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’nikova, T.I.; Porozov, Y.B.; Terentiev, A.A. Oxidative Stress and Advanced Lipoxidation and Glycation End Products (ALEs and AGEs) in Aging and Age-Related Diseases. Oxidative Med. Cell. Longev. 2019, 2019, 3085756. [Google Scholar] [CrossRef] [Green Version]
- Fuentealba, D.; Friguet, B.; Silva, E. Advanced glycation endproducts induce photocrosslinking and oxidation of bovine lens proteins through type-I mechanism. Photochem. Photobiol. 2009, 85, 185–194. [Google Scholar] [CrossRef]
- Yan, H.D.; Li, X.Z.; Xie, J.M.; Li, M. Effects of advanced glycation end products on renal fibrosis and oxidative stress in cultured NRK-49F cells. Chin. Med. J. 2007, 120, 787–793. [Google Scholar] [CrossRef]
- Grossin, N.; Auger, F.; Niquet-Leridon, C.; Durieux, N.; Montaigne, D.; Schmidt, A.M.; Susen, S.; Jacolot, P.; Beuscart, J.B.; Tessier, F.J.; et al. Dietary CML-enriched protein induces functional arterial aging in a RAGE-dependent manner in mice. Mol. Nutr. Food Res. 2015, 59, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Siddiqui, Z.; Rehman, S.; Khan, M.Y.; Khan, H.; Khanum, S.; Alouffi, S.; Saeed, M. A Glycation Angle to Look into the Diabetic Vasculopathy: Cause and Cure. Curr. Vasc. Pharmacol. 2017, 15, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Basta, G. Receptor for advanced glycation endproducts and atherosclerosis: From basic mechanisms to clinical implications. Atherosclerosis 2008, 196, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.M.; Smith, D.M.; Alt, N.; Thorpe, S.R.; Baynes, J.W. Tissue-specific variation in glycation of proteins in diabetes: Evidence for a functional role of amadoriase enzymes. Ann. N. Y. Acad. Sci. 2005, 1043, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Shakeel, M. Recent advances in understanding the role of oxidative stress in diabetic neuropathy. Diabetes Metab. Syndr. 2015, 9, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Portal-Nunez, S.; Ardura, J.A.; Lozano, D.; Martinez de Toda, I.; De la Fuente, M.; Herrero-Beaumont, G.; Largo, R.; Esbrit, P. Parathyroid hormone-related protein exhibits antioxidant features in osteoblastic cells through its N-terminal and osteostatin domains. Bone Jt. Res. 2018, 7, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Lagoda, G.; Leite, R.; Webb, R.C.; Burnett, A.L. NADPH oxidase activation: A mechanism of hypertension-associated erectile dysfunction. J. Sex. Med. 2008, 5, 544–551. [Google Scholar] [CrossRef]
- Taye, A.; Saad, A.H.; Kumar, A.H.; Morawietz, H. Effect of apocynin on NADPH oxidase-mediated oxidative stress-LOX-1-eNOS pathway in human endothelial cells exposed to high glucose. Eur. J. Pharmacol. 2010, 627, 42–48. [Google Scholar] [CrossRef]
- Mifune, Y.; Inui, A.; Muto, T.; Nishimoto, H.; Kataoka, T.; Kurosawa, T.; Yamaura, K.; Mukohara, S.; Niikura, T.; Kokubu, T.; et al. Influence of advanced glycation end products on rotator cuff. J. Shoulder Elb. Surg. 2019, 28, 1490–1496. [Google Scholar] [CrossRef]
- Heumuller, S.; Wind, S.; Barbosa-Sicard, E.; Schmidt, H.H.; Busse, R.; Schroder, K.; Brandes, R.P. Apocynin is not an inhibitor of vascular NADPH oxidases but an antioxidant. Hypertension 2008, 51, 211–217. [Google Scholar] [CrossRef]
- Gimenes, R.; Gimenes, C.; Rosa, C.M.; Xavier, N.P.; Campos, D.H.S.; Fernandes, A.A.H.; Cezar, M.D.M.; Guirado, G.N.; Pagan, L.U.; Chaer, I.D.; et al. Influence of apocynin on cardiac remodeling in rats with streptozotocin-induced diabetes mellitus. Cardiovasc. Diabetol. 2018, 17, 15. [Google Scholar] [CrossRef] [PubMed]
- Yokota, T.; Kinugawa, S.; Hirabayashi, K.; Matsushima, S.; Inoue, N.; Ohta, Y.; Hamaguchi, S.; Sobirin, M.A.; Ono, T.; Suga, T.; et al. Oxidative stress in skeletal muscle impairs mitochondrial respiration and limits exercise capacity in type 2 diabetic mice. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1069–H1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Duarte, S.; Montoya-Perez, R.; Marquez-Gamino, S.; Vera-Delgado, K.S.; Caudillo-Cisneros, C.; Sotelo-Barroso, F.; Sanchez-Briones, L.A.; Sanchez-Duarte, E. Apocynin Attenuates Diabetes-Induced Skeletal Muscle Dysfunction by Mitigating ROS Generation and Boosting Antioxidant Defenses in Fast-Twitch and Slow-Twitch Muscles. Life 2022, 12, 674. [Google Scholar] [CrossRef] [PubMed]
- Legerlotz, K.; Riley, G.P.; Screen, H.R. Specimen dimensions influence the measurement of material properties in tendon fascicles. J. Biomech. 2010, 43, 2274–2280. [Google Scholar] [CrossRef] [Green Version]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef]
- Hancock, J.T.; Desikan, R.; Neill, S.J. Role of reactive oxygen species in cell signalling pathways. Biochem. Soc. Trans. 2001, 29, 345–350. [Google Scholar] [CrossRef]
- Sumimoto, H. Structure, regulation and evolution of Nox-family NADPH oxidases that produce reactive oxygen species. FEBS J. 2008, 275, 3249–3277. [Google Scholar] [CrossRef]
- Koike, S.; Yano, S.; Tanaka, S.; Sheikh, A.M.; Nagai, A.; Sugimoto, T. Advanced Glycation End-Products Induce Apoptosis of Vascular Smooth Muscle Cells: A Mechanism for Vascular Calcification. Int. J. Mol. Sci. 2016, 17, 1567. [Google Scholar] [CrossRef] [Green Version]
- Ueda, Y.; Inui, A.; Mifune, Y.; Sakata, R.; Muto, T.; Harada, Y.; Takase, F.; Kataoka, T.; Kokubu, T.; Kuroda, R. The effects of high glucose condition on rat tenocytes in vitro and rat Achilles tendon in vivo. Bone Jt. Res. 2018, 7, 362–372. [Google Scholar] [CrossRef]
- Ott, C.; Jacobs, K.; Haucke, E.; Navarrete Santos, A.; Grune, T.; Simm, A. Role of advanced glycation end products in cellular signaling. Redox Biol. 2014, 2, 411–429. [Google Scholar] [CrossRef] [Green Version]
- Olukman, M.; Orhan, C.E.; Celenk, F.G.; Ulker, S. Apocynin restores endothelial dysfunction in streptozotocin diabetic rats through regulation of nitric oxide synthase and NADPH oxidase expressions. J. Diabetes Complicat. 2010, 24, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Thallas-Bonke, V.; Thorpe, S.R.; Coughlan, M.T.; Fukami, K.; Yap, F.Y.; Sourris, K.C.; Penfold, S.A.; Bach, L.A.; Cooper, M.E.; Forbes, J.M. Inhibition of NADPH oxidase prevents advanced glycation end product-mediated damage in diabetic nephropathy through a protein kinase C-alpha-dependent pathway. Diabetes 2008, 57, 460–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahali, S.; Raviprakash, N.; Raghavendra, P.B.; Manna, S.K. Advanced glycation end products (AGEs) induce apoptosis via a novel pathway: Involvement of Ca2+ mediated by interleukin-8 protein. J. Biol. Chem. 2011, 286, 34903–34913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushal, N.; Bansal, M.P. Dietary selenium variation-induced oxidative stress modulates CDC2/cyclin B1 expression and apoptosis of germ cells in mice testis. J. Nutr. Biochem. 2007, 18, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.Y.; Zhang, Q.; Dai, D.Z.; Ying, H.J.; Wang, Q.J.; Dai, Y. Effects of strontium fructose 1,6-diphosphate on expression of apoptosis-related genes and oxidative stress in testes of diabetic rats. Int. J. Urol. 2008, 15, 251–256. [Google Scholar] [CrossRef]
- Yuan, J.; Murrell, G.A.; Trickett, A.; Landtmeters, M.; Knoops, B.; Wang, M.X. Overexpression of antioxidant enzyme peroxiredoxin 5 protects human tendon cells against apoptosis and loss of cellular function during oxidative stress. Biochim. Biophys. Acta 2004, 1693, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Liu, Z.; Zhuan, L.; Wang, T.; Guo, S.; Wang, S.; Liu, J.; Ye, Z. Effects of apocynin on oxidative stress and expression of apoptosis-related genes in testes of diabetic rats. Mol. Med. Rep. 2013, 7, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Kameyama, M.; Chen, K.R.; Mukai, K.; Shimada, A.; Atsumi, Y.; Yanagimoto, S. Histopathological characteristics of stenosing flexor tenosynovitis in diabetic patients and possible associations with diabetes-related variables. J. Hand Surg. Am 2013, 38, 1331–1339. [Google Scholar] [CrossRef]
- Tsai, W.C.; Liang, F.C.; Cheng, J.W.; Lin, L.P.; Chang, S.C.; Chen, H.H.; Pang, J.H. High glucose concentration up-regulates the expression of matrix metalloproteinase-9 and -13 in tendon cells. BMC Musculoskelet. Disord. 2013, 14, 255. [Google Scholar] [CrossRef] [Green Version]
- Kislinger, T.; Fu, C.; Huber, B.; Qu, W.; Taguchi, A.; Du Yan, S.; Hofmann, M.; Yan, S.F.; Pischetsrieder, M.; Stern, D.; et al. N(epsilon)-(carboxymethyl)lysine adducts of proteins are ligands for receptor for advanced glycation end products that activate cell signaling pathways and modulate gene expression. J. Biol. Chem. 1999, 274, 31740–31749. [Google Scholar] [CrossRef] [Green Version]
- Oreff, G.L.; Fenu, M.; Vogl, C.; Ribitsch, I.; Jenner, F. Species variations in tenocytes’ response to inflammation require careful selection of animal models for tendon research. Sci. Rep. 2021, 11, 12451. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.Y.; Cooper, M.E. Clinical review: The role of advanced glycation end products in progression and complications of diabetes. J. Clin. Endocrinol. Metab. 2008, 93, 1143–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, L.; Bestwick, C.S.; Bestwick, L.A.; Maffulli, N.; Aspden, R.M. Phenotypic drift in human tenocyte culture. Tissue Eng. 2006, 12, 1843–1849. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Patients Registered (n = 12) |
|---|---|
| Age (y), mean ± SD | 64.8 ± 11.7 SD |
| Sex (%) | |
| Men | 4 (33) |
| Women | 8 (66) |
| Complications (%) | |
| Diabetes | 2 (17) |
| Hypertension | 3 (17) |
| Rotator cuff tear size (%) (Cofield classification) | Small: 6 (50); Medium: 5 (42); Large: 1 (17) |
| Fatty degeneration (%) (Goutallier classification) | Stage1: 5 (42); Stage2: 6 (50); Stage3: 1 (17) |
| Gene | Oligonucleotide sequence |
|---|---|
| NOX-1 | Forward 5′ GGTTTTACCGCTCCCAGCAGAA 3′ |
| Reverse 5′ CTTCCATGCTGAAGCCACGCTT 3′ | |
| NOX-4 | Forward 5′ GCCAGAGTATCACTACCTCCAC 3′ |
| Reverse 5′ CTCGGAGGTAAGCCAAGAGTGT 3′ | |
| IL-6 | Forward 5′ AGACAGCCACTCACCTCTTCAG 3′ |
| Reverse 5′ TTCTGCCAGTGCCTCTTTGCTG 3′ | |
| RAGE | Forward 5′ CACCTTCTCCTGTAGCTTCAGC 3′ |
| Reverse 5′ AGGAGCTACTGCTCCACCTTCT 3′ | |
| GAPDH | Forward 5′ GTCTCCTCTGACTTCAACAGCG 3′ |
| Reverse 5′ ACCACCCTGTTGCTGTAGCCAA 3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furukawa, T.; Kurosawa, T.; Mifune, Y.; Inui, A.; Nishimoto, H.; Ueda, Y.; Kataoka, T.; Yamaura, K.; Mukohara, S.; Yoshikawa, T.; et al. Elicitation of Inhibitory Effects for AGE-Induced Oxidative Stress in Rotator Cuff-Derived Cells by Apocynin. Curr. Issues Mol. Biol. 2023, 45, 3434-3445. https://doi.org/10.3390/cimb45040225
Furukawa T, Kurosawa T, Mifune Y, Inui A, Nishimoto H, Ueda Y, Kataoka T, Yamaura K, Mukohara S, Yoshikawa T, et al. Elicitation of Inhibitory Effects for AGE-Induced Oxidative Stress in Rotator Cuff-Derived Cells by Apocynin. Current Issues in Molecular Biology. 2023; 45(4):3434-3445. https://doi.org/10.3390/cimb45040225
Chicago/Turabian StyleFurukawa, Takahiro, Takashi Kurosawa, Yutaka Mifune, Atsuyuki Inui, Hanako Nishimoto, Yasuhiro Ueda, Takeshi Kataoka, Kohei Yamaura, Shintaro Mukohara, Tomoya Yoshikawa, and et al. 2023. "Elicitation of Inhibitory Effects for AGE-Induced Oxidative Stress in Rotator Cuff-Derived Cells by Apocynin" Current Issues in Molecular Biology 45, no. 4: 3434-3445. https://doi.org/10.3390/cimb45040225