
The Involvement of Hypoxia in the Response of Neuroblastoma Cells to the Exposure of Atorvastatin


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Treatments
2.3. Hypoxia Induction
2.4. Cell Culture
2.5. Neutral Red Assay
2.6. Thiazolyl Blue Tetrazolium Bromide Assay
2.7. Cell Morphology Assessment
2.8. Statistical Analyses
2.9. In Silico ADMET Modeling
3. Results and Discussion
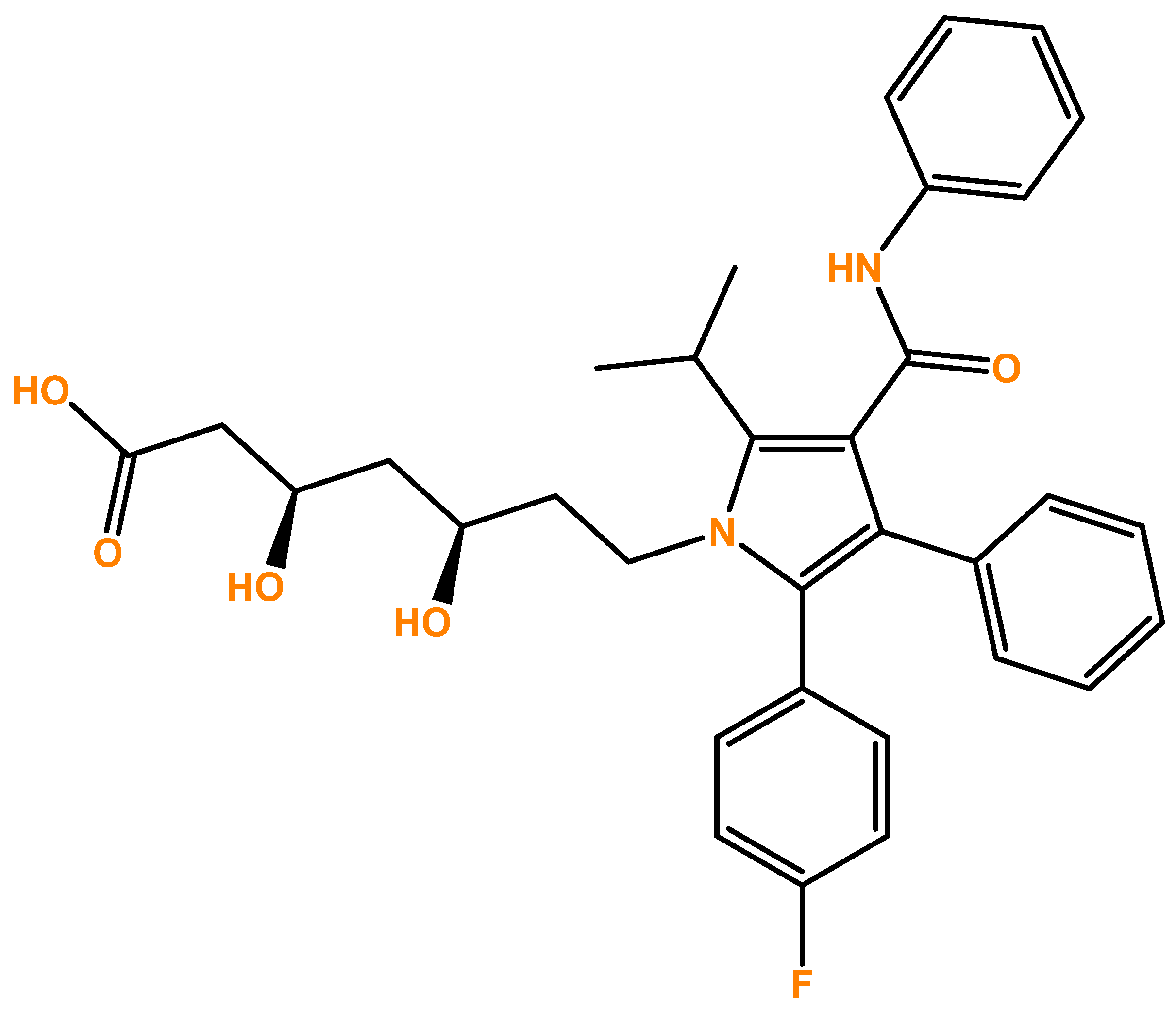
3.1. The Effect of Atorvastatin on the Viability and Morphology of SH-SY5Y Cells
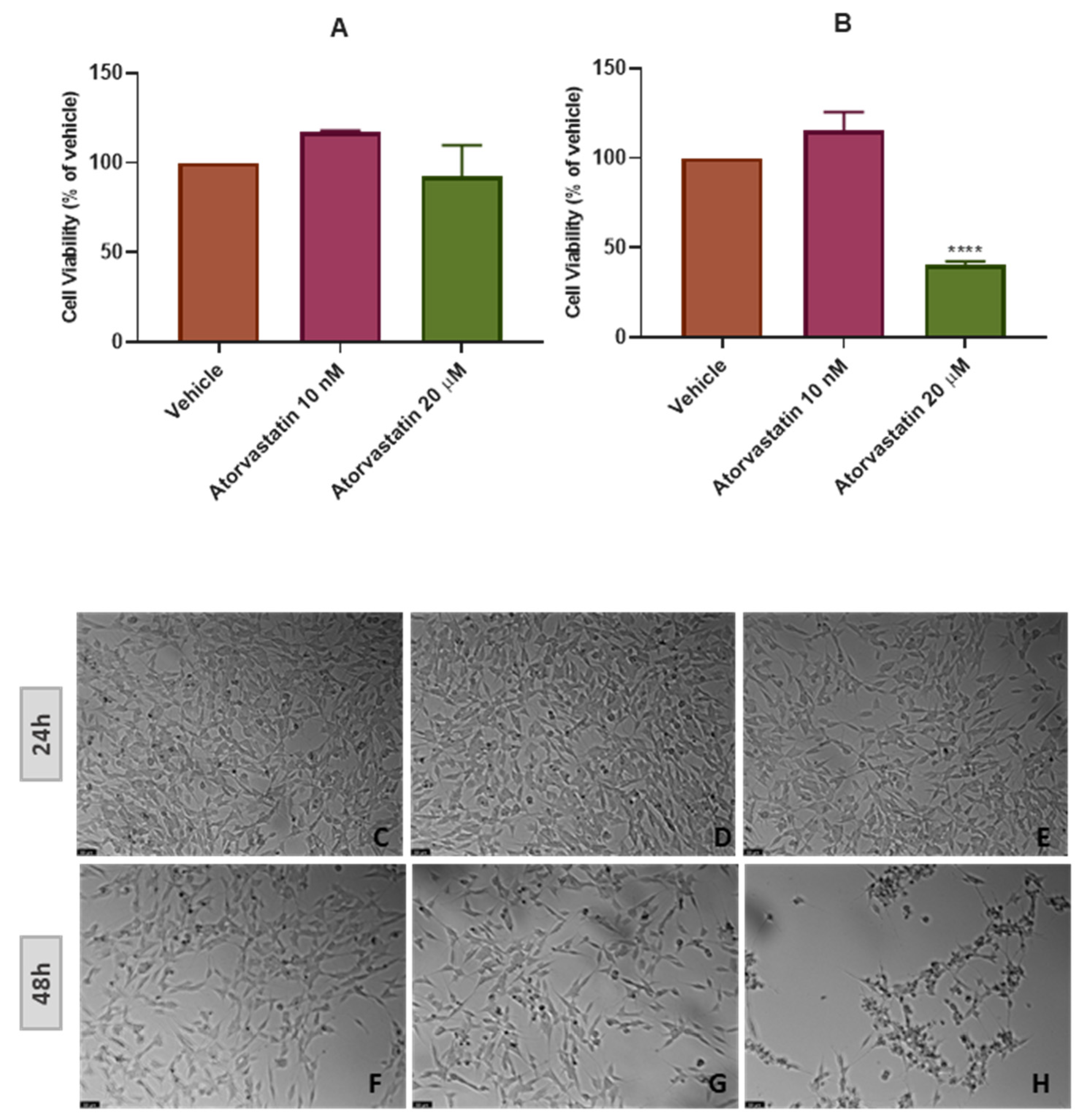
3.2. The Effect of Cobalt Chloride Induced Hypoxia on SH-SY5Y Cellular Viability and Morphology after Atorvastatin Application
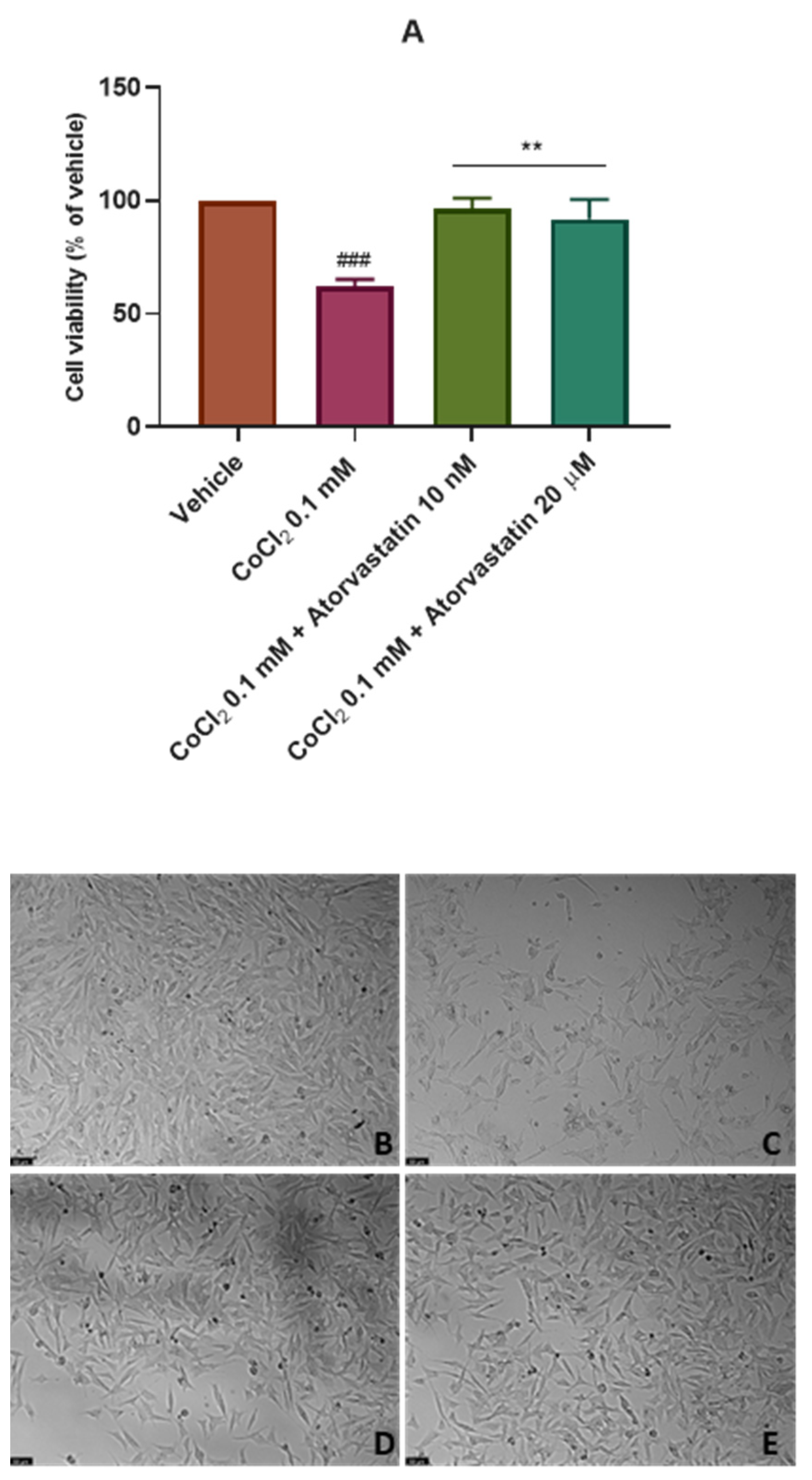
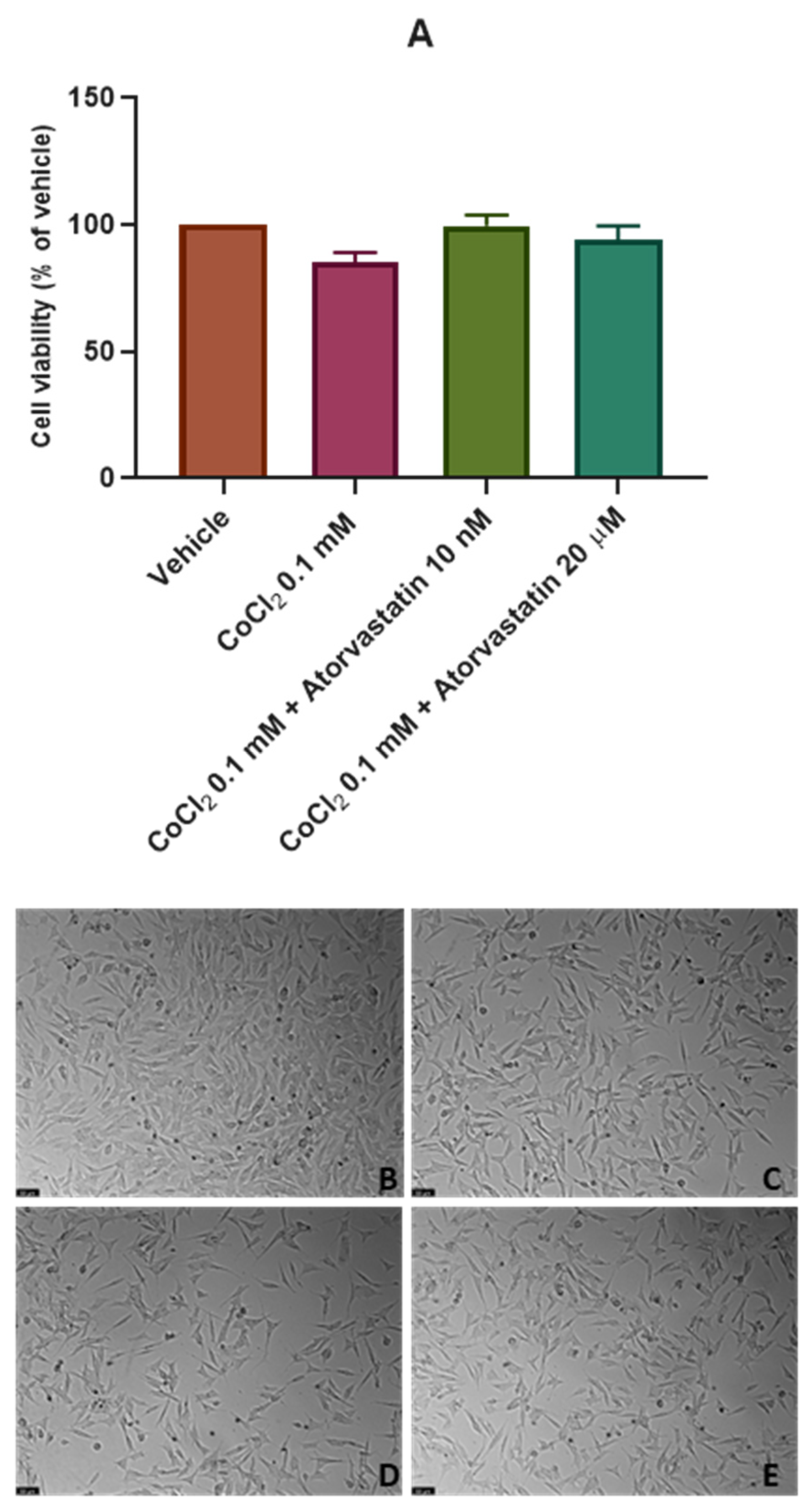
3.3. The Effect of the Combination of Echinomycin with Cobalt Chloride on SH-SY5Y Cellular Viability and Morphology after Atorvastatin Application
3.4. In Silico Validation of the Effect of the Combination of Atorvastatin and Echinomycin
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 20 February 2023).
- Correia, A.; Silva, D.; Correia, A.; Vilanova, M.; Gärtner, F.; Vale, N. Study of New Therapeutic Strategies to Combat Breast Cancer Using Drug Combinations. Biomolecules 2018, 8, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zhou, L.; Xie, N.; Nice, E.C.; Zhang, T.; Cui, Y.; Huang, C. Overcoming cancer therapeutic bottleneck by drug repurposing. Signal Transduct. Target. Ther. 2020, 5, 113. [Google Scholar] [CrossRef]
- Ishida, J.; Konishi, M.; Ebner, N.; Springer, J. Repurposing of approved cardiovascular drugs. J. Transl. Med. 2016, 14, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, D.; Vale, N. New Trends for Antimalarial Drugs: Synergism between Antineoplastics and Antimalarials on Breast Cancer Cells. Biomolecules 2020, 10, 1623. [Google Scholar] [CrossRef]
- Wu, H.; Huang, D.; Zhou, H.; Sima, X.; Wu, Z.; Sun, Y.; Wang, L.; Ruan, Y.; Wu, Q.; Wu, F.; et al. Metformin: A promising drug for human cancers (Review). Oncol. Lett. 2022, 24, 204. [Google Scholar] [CrossRef]
- Ekinci, E.; Rohondia, S.; Khan, R.; Dou, Q.P. Repurposing Disulfiram as An Anti-Cancer Agent: Updated Review on Literature and Patents. Recent Pat. Anti-Cancer Drug Discov. 2019, 14, 113–132. [Google Scholar] [CrossRef] [PubMed]
- Latif, T.; Chauhan, N.; Khan, R.; Moran, A.; Usmani, S.Z. Thalidomide and its analogues in the treatment of Multiple Myeloma. Exp. Hematol. Oncol. 2012, 1, 27. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Hu, J.-W.; He, X.-R.; Jin, W.-L.; He, X.-Y. Statins: A repurposed drug to fight cancer. J. Exp. Clin. Cancer Res. 2021, 40, 241. [Google Scholar] [CrossRef]
- Barbalata, C.I.; Tefas, L.R.; Achim, M.; Tomuta, I.; Porfire, A.S. Statins in risk-reduction and treatment of cancer. World J. Clin. Oncol. 2020, 11, 573–588. [Google Scholar] [CrossRef]
- Farina, H.G.; Bublik, D.R.; Alonso, D.F.; Gomez, D.E. Lovastatin alters cytoskeleton organization and inhibits experimental metastasis of mammary carcinoma cells. Clin. Exp. Metastasis 2002, 19, 551–559. [Google Scholar] [CrossRef]
- Muz, B.; de la Puente, P.; Azab, F.; Azab, A.K. The role of hypoxia in cancer progression, angiogenesis, metastasis, and resistance to therapy. Hypoxia 2015, 3, 83. [Google Scholar] [CrossRef] [Green Version]
- d’Hose, D.; Mignion, L.; Hamelin, L.; Sonveaux, P.; Jordan, B.F.; Gallez, B. Statins Alleviate Tumor Hypoxia in Prostate Cancer Models by Decreasing Oxygen Consumption: An Opportunity for Radiosensitization? Biomolecules 2022, 12, 1418. [Google Scholar] [CrossRef]
- Marti, J.L.G.; Beckwitt, C.H.; Clark, A.M.; Wells, A. Atorvastatin facilitates chemotherapy effects in metastatic triple-negative breast cancer. Br. J. Cancer 2021, 125, 1285–1298. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Cao, Y.; Liu, L.; Zhao, Z.; Yin, H.; Wang, H. Atorvastatin regulates the migration and invasion of prostate cancer through the epithelial-mesenchymal transformation and matrix metalloproteinase pathways. Investig. Clin. Urol. 2022, 63, 350. [Google Scholar] [CrossRef] [PubMed]
- Bakar-Ates, F.; Ozkan, E. Atorvastatin induces downregulation of matrix metalloproteinase-2/9 in MDA-MB-231 triple negative breast cancer cells. Med. Oncol. 2022, 40, 22. [Google Scholar] [CrossRef] [PubMed]
- Zipinotti dos Santos, D.; Santos Guimaraes, I.D.; Hakeem-Sanni, M.F.; Cochran, B.J.; Rye, K.A.; Grewal, T.; Hoy, A.J.; Rangel, L.B.A. Atorvastatin improves cisplatin sensitivity through modulation of cholesteryl ester homeostasis in breast cancer cells. Discov. Oncol. 2022, 13, 135. [Google Scholar] [CrossRef] [PubMed]
- Choe, E.-J.; Lee, C.-H.; Bae, J.-H.; Park, J.-M.; Park, S.-S.; Baek, M.-C. Atorvastatin Enhances the Efficacy of Immune Checkpoint Therapy and Suppresses the Cellular and Extracellular Vesicle PD-L1. Pharmaceutics 2022, 14, 1660. [Google Scholar] [CrossRef] [PubMed]
- Colon, N.C.; Chung, D.H. Neuroblastoma. Adv. Pediatr. 2011, 58, 297–311. [Google Scholar] [CrossRef] [Green Version]
- Kovalevich, J.; Langford, D. Considerations for the Use of SH-SY5Y Neuroblastoma Cells in Neurobiology. In Methods in Molecular Biology (Clifton, N.J.); NIH Public Access: Clifton, NJ, USA, 2013; Volume 1078, pp. 9–21. ISBN 9781627036399. [Google Scholar]
- Muñoz-Sánchez, J.; Chánez-Cárdenas, M.E. The use of cobalt chloride as a chemical hypoxia model. J. Appl. Toxicol. 2019, 39, 556–570. [Google Scholar] [CrossRef]
- Hellwig-Bürgel, T.; Stiehl, D.P.; Wagner, A.E.; Metzen, E.; Jelkmann, W. Review: Hypoxia-inducible factor-1 (HIF-1): A novel transcription factor in immune reactions. J. Interferon Cytokine Res. 2005, 25, 297–310. [Google Scholar] [CrossRef]
- Lee, J.W.; Bae, S.H.; Jeong, J.W.; Kim, S.H.; Kim, K.W. Hypoxia-inducible factor (HIF-1)alpha: Its protein stability and biological functions. Exp. Mol. Med. 2004, 36, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakkola, J.; Hukkanen, J.; Turpeinen, M.; Pelkonen, O. Inhibition and induction of CYP enzymes in humans: An update. Arch. Toxicol. 2020, 94, 3671–3722. [Google Scholar] [CrossRef]
- Park, J.-E.; Kim, K.-B.; Bae, S.K.; Moon, B.-S.; Liu, K.-H.; Shin, J.-G. Contribution of cytochrome P450 3A4 and 3A5 to the metabolism of atorvastatin. Xenobiotica 2008, 38, 1240–1251. [Google Scholar] [CrossRef]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, M.; Turnbull, T.; Sebastian, S.; Kempson, I. The MTT Assay: Utility, Limitations, Pitfalls, and Interpretation in Bulk and Single-Cell Analysis. Int. J. Mol. Sci. 2021, 22, 12827. [Google Scholar] [CrossRef]
- Correia, A.S.; Fraga, S.; Teixeira, J.P.; Vale, N. Cell Model of Depression: Reduction of Cell Stress with Mirtazapine. Int. J. Mol. Sci. 2022, 23, 4942. [Google Scholar] [CrossRef]
- Saraste, A.; Pulkki, K. Morphologic and biochemical hallmarks of apoptosis. Cardiovasc. Res. 2000, 45, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Fatehi Hassanabad, A. Current perspectives on statins as potential anti-cancer therapeutics: Clinical outcomes and underlying molecular mechanisms. Transl. Lung Cancer Res. 2019, 8, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Butterick, T.A.; Igbavboa, U.; Eckert, G.P.; Sun, G.Y.; Weisman, G.A.; Müller, W.E.; Wood, W.G. Simvastatin Stimulates Production of the Antiapoptotic Protein Bcl-2 via Endothelin-1 and NFATc3 in SH-SY5Y Cells. Mol. Neurobiol. 2010, 41, 384–391. [Google Scholar] [CrossRef] [Green Version]
- Atil, B.; Sieczkowski, E.; Hohenegger, M. Statins reduce endogenous dolichol levels in the neuroblastoma cell line SH-SY5Y. BMC Pharmacol. Toxicol. 2012, 13, A51. [Google Scholar] [CrossRef] [Green Version]
- Sebestyén, A.; Kopper, L.; Dankó, T.; Tímár, J. Hypoxia Signaling in Cancer: From Basics to Clinical Practice. Pathol. Oncol. Res. 2021, 27, 1609802. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, Y.; Miyagi-Shiohira, C.; Noguchi, H.; Omasa, T. Atorvastatin Inhibits the HIF1α-PPAR Axis, Which Is Essential for Maintaining the Function of Human Induced Pluripotent Stem Cells. Mol. Ther. 2018, 26, 1715–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Zhang, L.; Zhang, H. Atorvastatin combined with routine therapy on HIF-1, VEGF concentration and cardiac function in rats with acute myocardial infarction. Exp. Ther. Med. 2020, 19, 2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atorvastatin | C33H35FN2O5. PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Atorvastatin (accessed on 2 March 2023).
- AID 781329—pKa (Acid-Base Dissociation Constant) as Determined by Other Workers. PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/bioassay/781329#sid=103554720 (accessed on 2 March 2023).
- Atorvastatin: Uses, Interactions, Mechanism of Action. DrugBank Online. Available online: https://go.drugbank.com/drugs/DB01076 (accessed on 2 March 2023).
- McDonnell, A.M.; Dang, C.H. Basic Review of the Cytochrome P450 System. J. Adv. Pract. Oncol. 2013, 4, 263–268. [Google Scholar] [CrossRef]
- Valentovic, M. Atorvastatin. In xPharm: The Comprehensive Pharmacology Reference; Elsevier: Amsterdam, The Netherlands, 2007; pp. 1–6. ISBN 9780080552323. [Google Scholar]
- Hirota, T.; Fujita, Y.; Ieiri, I. An updated review of pharmacokinetic drug interactions and pharmacogenetics of statins. Expert Opin. Drug Metab. Toxicol. 2020, 16, 809–822. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Predicated Value | Observed Value | Reference |
|---|---|---|---|
| Molecular Weight (g/mol) | 558.65 | 558.64 | [36] |
| Ionization constant (pKa) | 4.71 | 4.46 | [37] |
| LogP | 4.276 | 6.360 | [38] |
| LogD | 1.640 | ND | ADMET Predictor |
| Solubility (mg/mL) | 0.185 | 1.120 | [36] |
| Permeability measure (cm/s × 104) | 1.728 | ND | ADMET Predictor |
| Blood-brain barrier permeability | Low likelihood | ||
| Unbound fraction (fu) (%) | 5.037 | ||
| Blood/plasma ratio | 0.667 | ||
| Volume of distribution (Vd) (L/kg) | 0.317 | ||
| Cmax (ng/mL) * | 301.630 | ||
| Tmax (h) * | 1.760 | ||
| AUCinf (ng·h/mL) * | 1366.220 | ||
| T1/2 (h) * | 1.730 | ||
| Total clearance (CL) (L/h) * | 6.560 |
| Drug | CYP Enzymes | Inhibitor | Substrate | Km (μM) | Vmax (nmol/min/nmol Enzyme) | Clint (μL/min/mg HLM Protein) | Sites of Metabolism |
|---|---|---|---|---|---|---|---|
| Atorvastatin | 2C9 | Yes (77%) | Yes (45%) | 6.096 | 1.876 | 22.467 | C33, C19, C3, C5, C17, C2 |
| 2C8 | ND | Yes (91%) | ND | ND | ND | C18, C19, C33, C1, C17 | |
| 3A4 | Yes (80%) | Yes (72%) | 17.800 | 77.080 | 480.670 | C19, C33, C1, C17, C18, C28 | |
| Echinomycin | 2C8 | ND | Yes (91%) | ND | ND | ND | S53, C48, C25, C58, S51, C32, C33 |
| 3A4 | Yes (80%) | Yes (81%) | 0.621 | 0.291 | 52.014 | S53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correia, A.S.; Marques, L.; Vale, N. The Involvement of Hypoxia in the Response of Neuroblastoma Cells to the Exposure of Atorvastatin. Curr. Issues Mol. Biol. 2023, 45, 3333-3346. https://doi.org/10.3390/cimb45040218
Correia AS, Marques L, Vale N. The Involvement of Hypoxia in the Response of Neuroblastoma Cells to the Exposure of Atorvastatin. Current Issues in Molecular Biology. 2023; 45(4):3333-3346. https://doi.org/10.3390/cimb45040218
Chicago/Turabian StyleCorreia, Ana Salomé, Lara Marques, and Nuno Vale. 2023. "The Involvement of Hypoxia in the Response of Neuroblastoma Cells to the Exposure of Atorvastatin" Current Issues in Molecular Biology 45, no. 4: 3333-3346. https://doi.org/10.3390/cimb45040218