
Transcriptome Sequencing Reveals Salmonella Flagellin Activation of Interferon-β-Related Immune Responses in Macrophages
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice and Ethics Statement
2.2. Extraction and Characterization of Flagellin (FliC)
2.3. TLR5 Ligand Activity of Flagellin
2.4. Flagellin Stimulation of RAW264.7 Cells at Different Concentrations
2.5. RNA-Seq and Data Analysis
2.6. Verification of RNA-Seq
2.7. Flagellin Stimulation of TLR4-Deficient HCT116-Dual Cells
2.8. Expression Levels of IFN-β in RAW264.7 Cells Stimulated by Polymyxin B (PMB)-Treated FliC
2.9. Flagellin Stimulation of Human THP-1 Cells
2.10. Isolation of Bone Marrow-Derived Macrophages (BMMs) and Dendritic Cells (BMDCs)
2.11. Flagellin Stimulation of BMMs and BMDCs
2.12. RNA Extraction, Reverse Transcription, and qRT-PCR
2.13. ELISA to Detect the Secretion Level of IFN-β
2.14. Statistical Analysis
3. Results
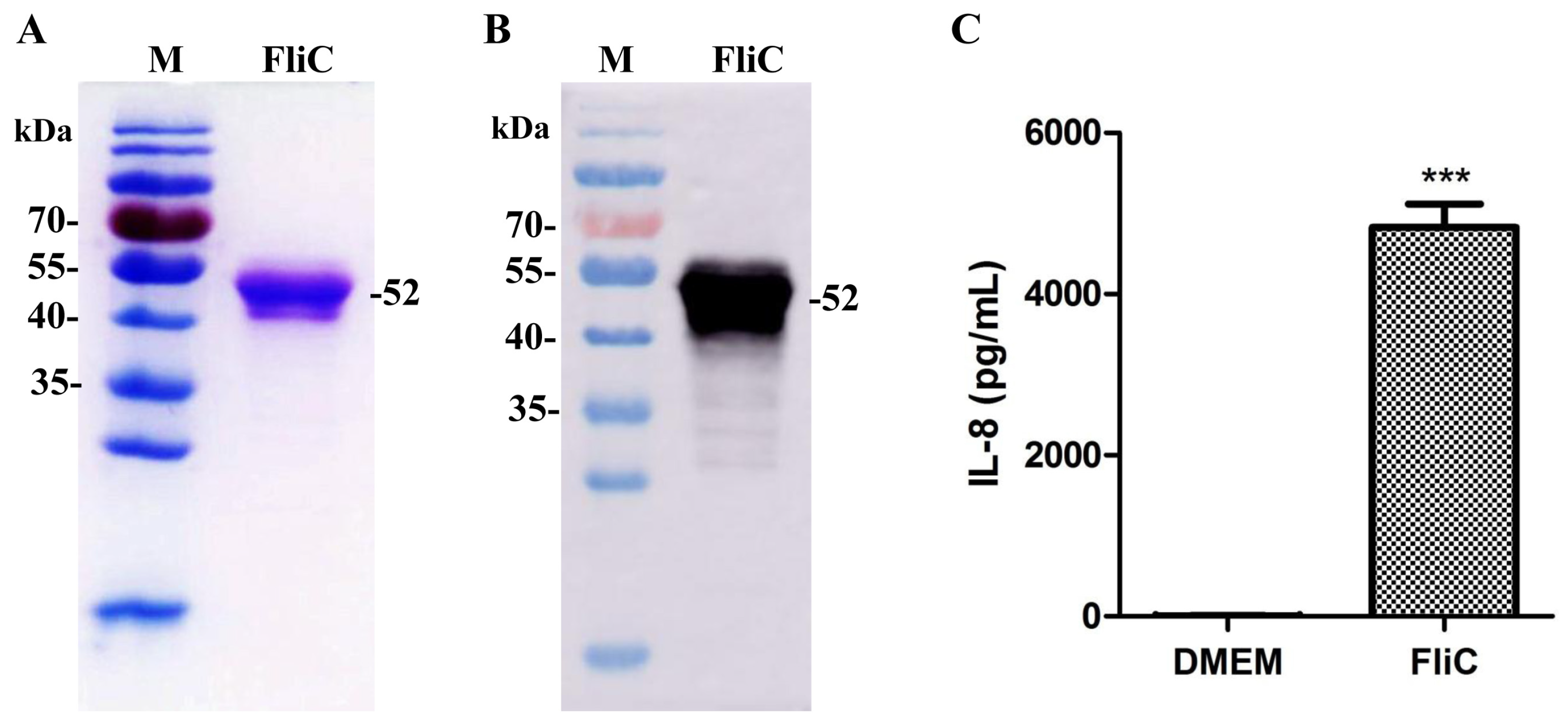
3.1. Identification of the Flagellin from Salmonella typhimurium
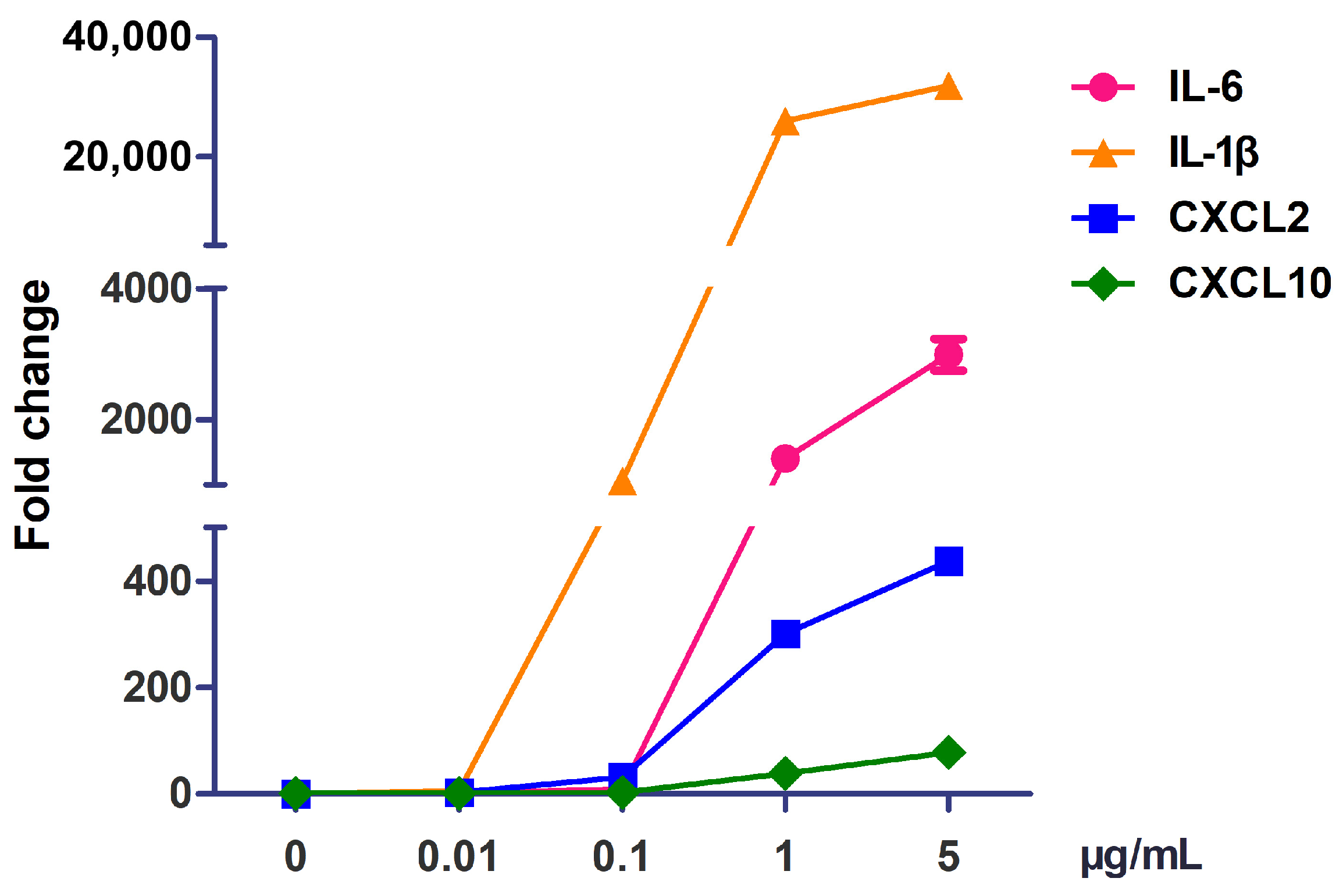
3.2. The Optimal Concentration of Flagellin to Activate RAW264.7 Cells
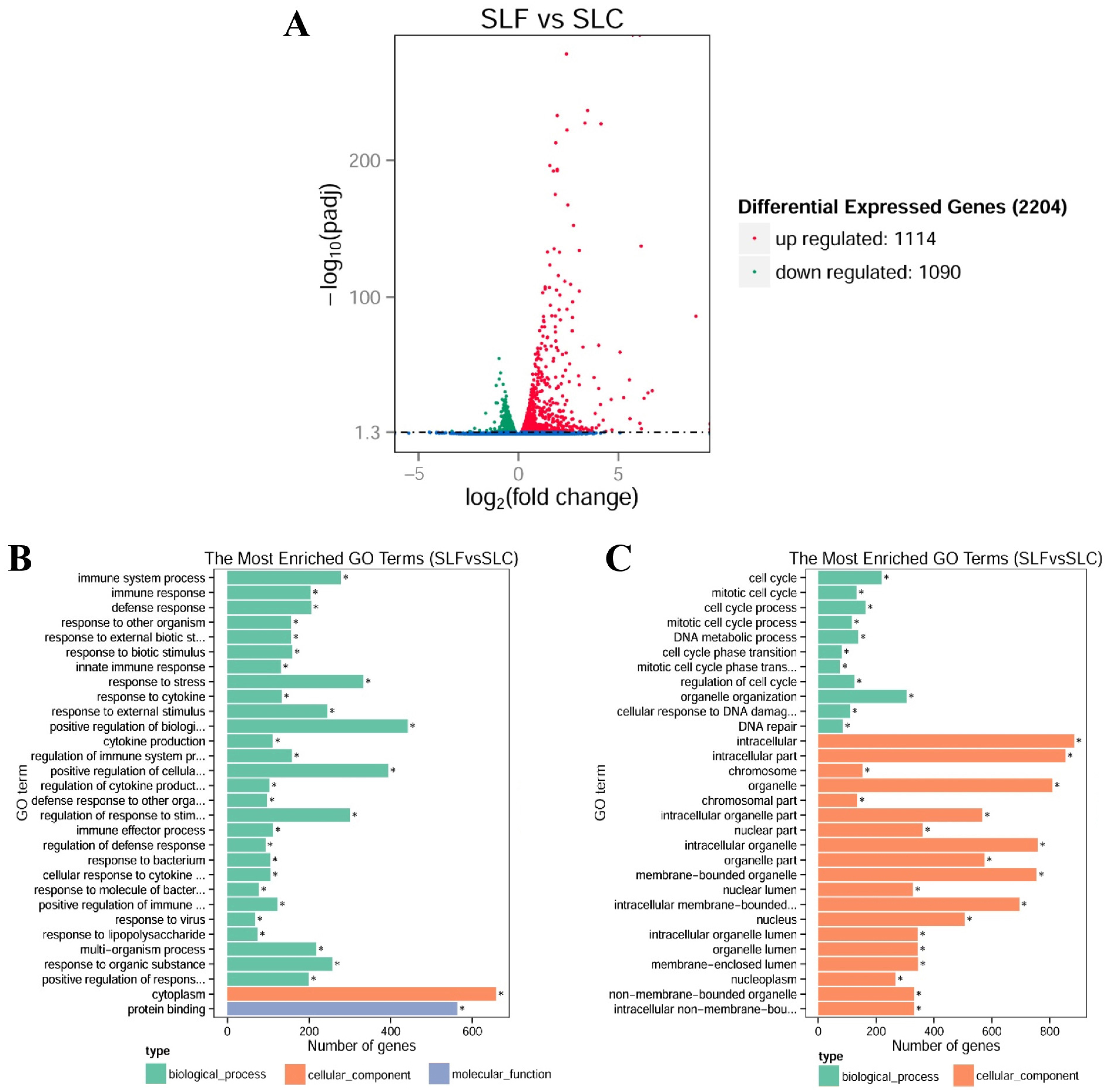
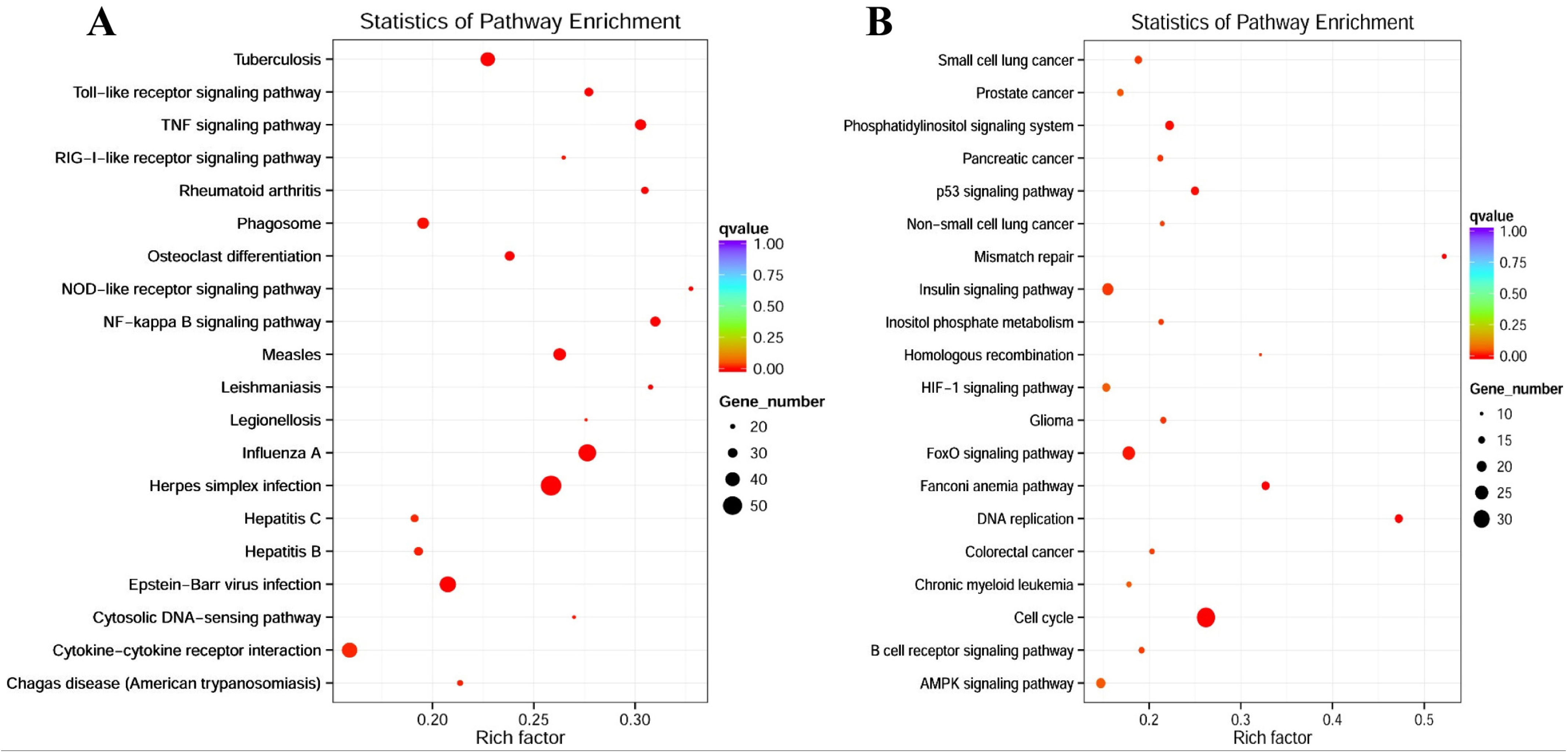
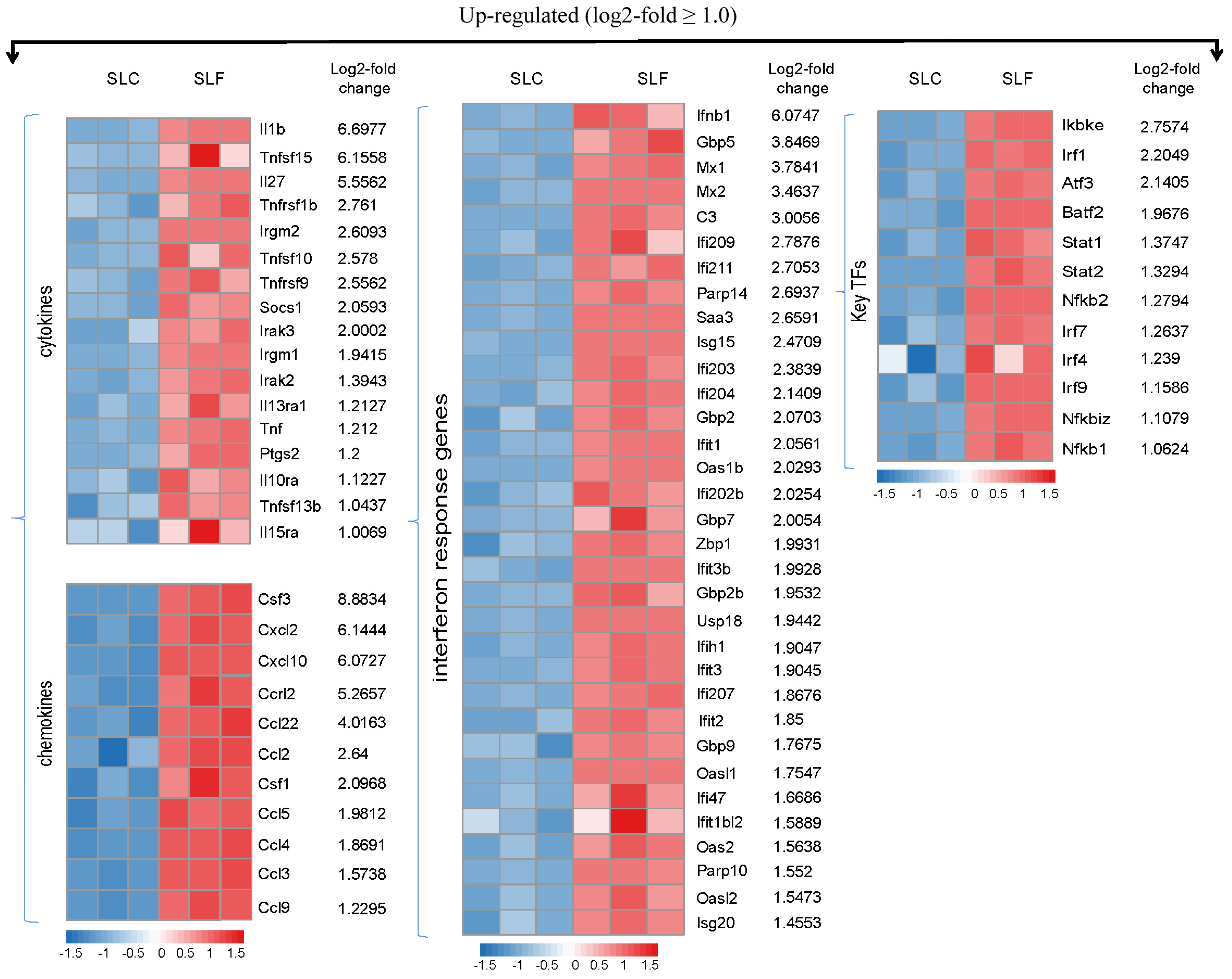
3.3. RNA-Seq Transcriptional Profiling in FliC-Stimulated RAW264.7 Cells
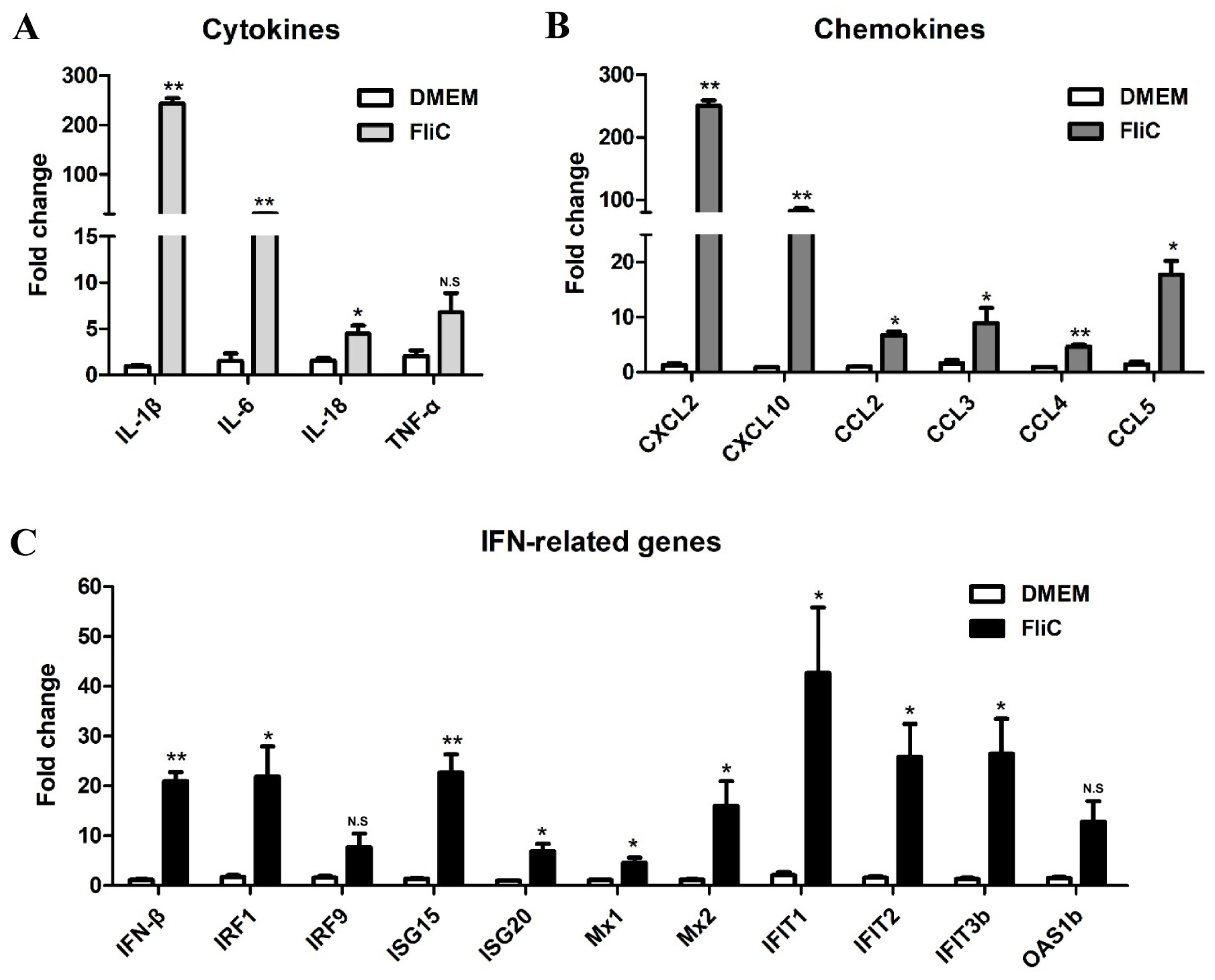
3.4. Verification of DEGs Identified by RNA-Seq by qRT-PCR
3.5. Expression and Secretion Levels of IFN-β in RAW264.7 Cells Stimulated by Trypsin-Treated FliC
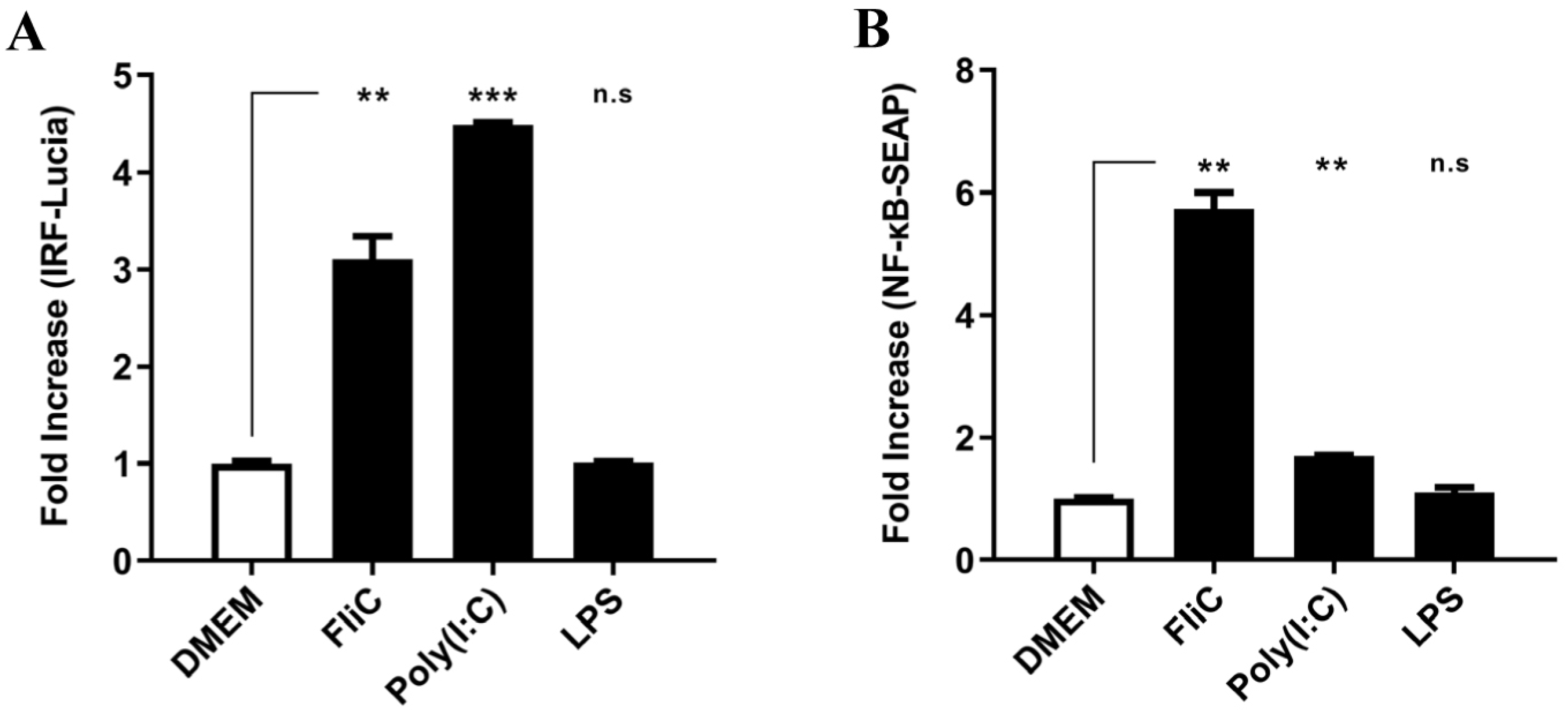
3.6. Activation of IRF Pathway in TLR4-Deficient HCT116-Dual Cells Stimulated by FliC
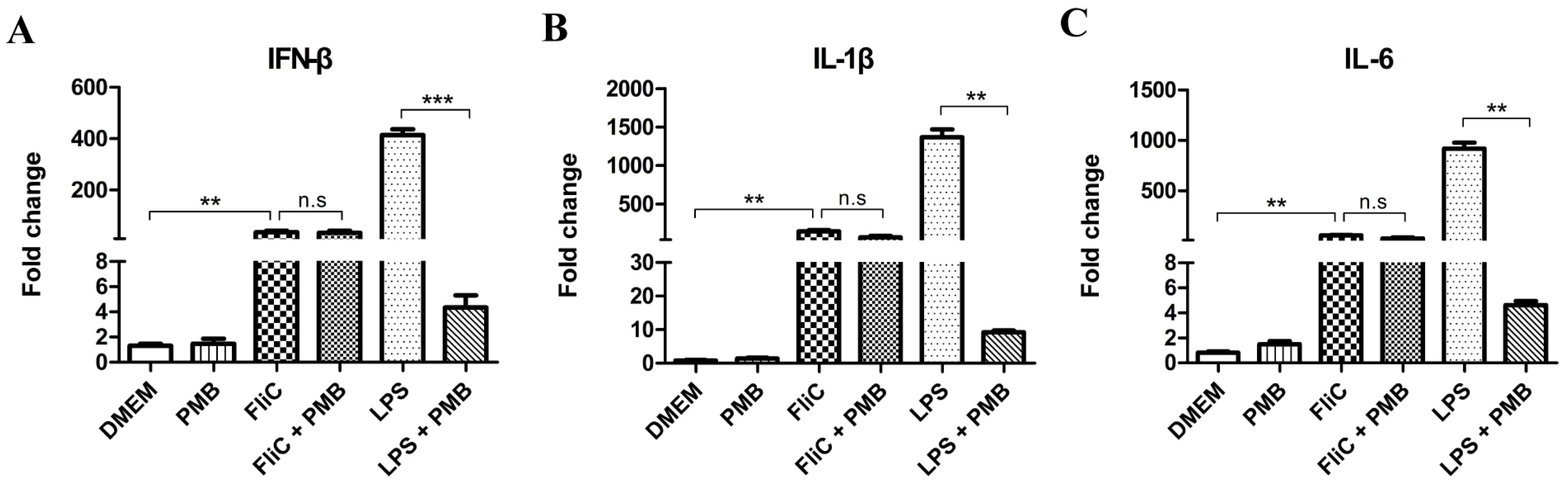
3.7. Induction of IFN-β in RAW264.7 Cells Stimulated by Polymyxin B-Treated FliC
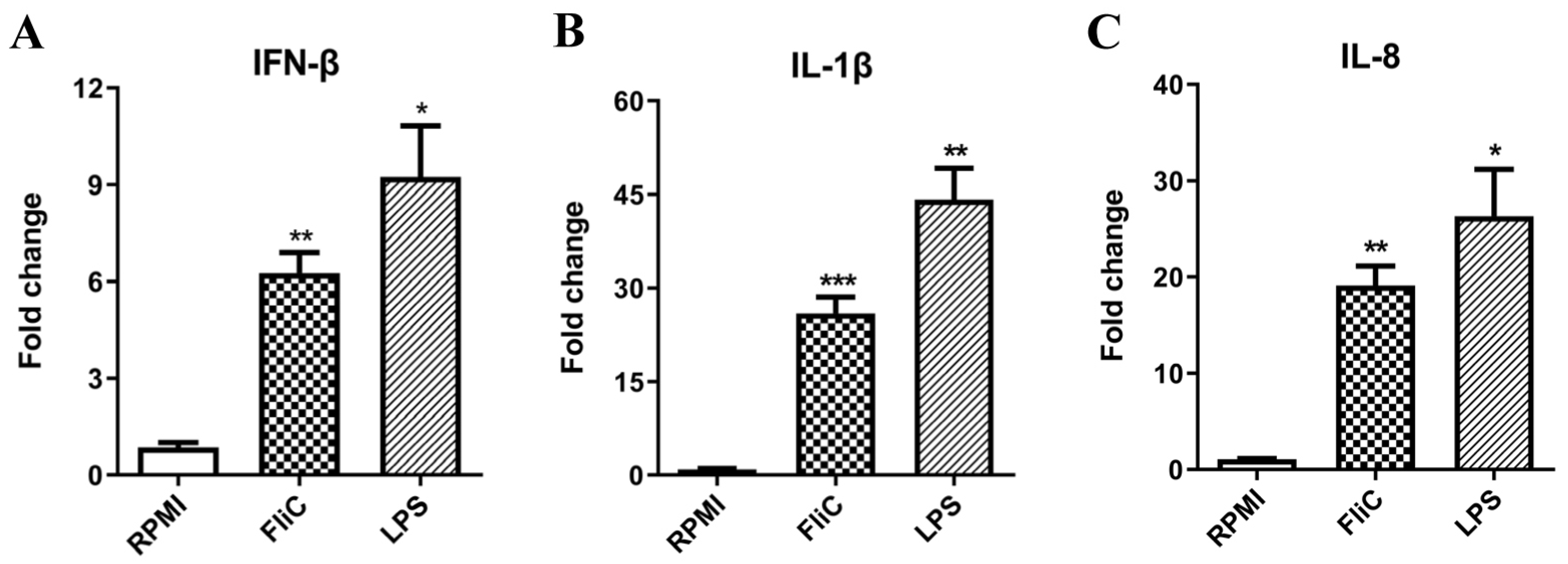
3.8. Flagellin Stimulates IFN-β Expression in Human THP-1 Cells
3.9. Flagellin Stimulates IFN-β Expression and Secretion in BMMs and BMDCs
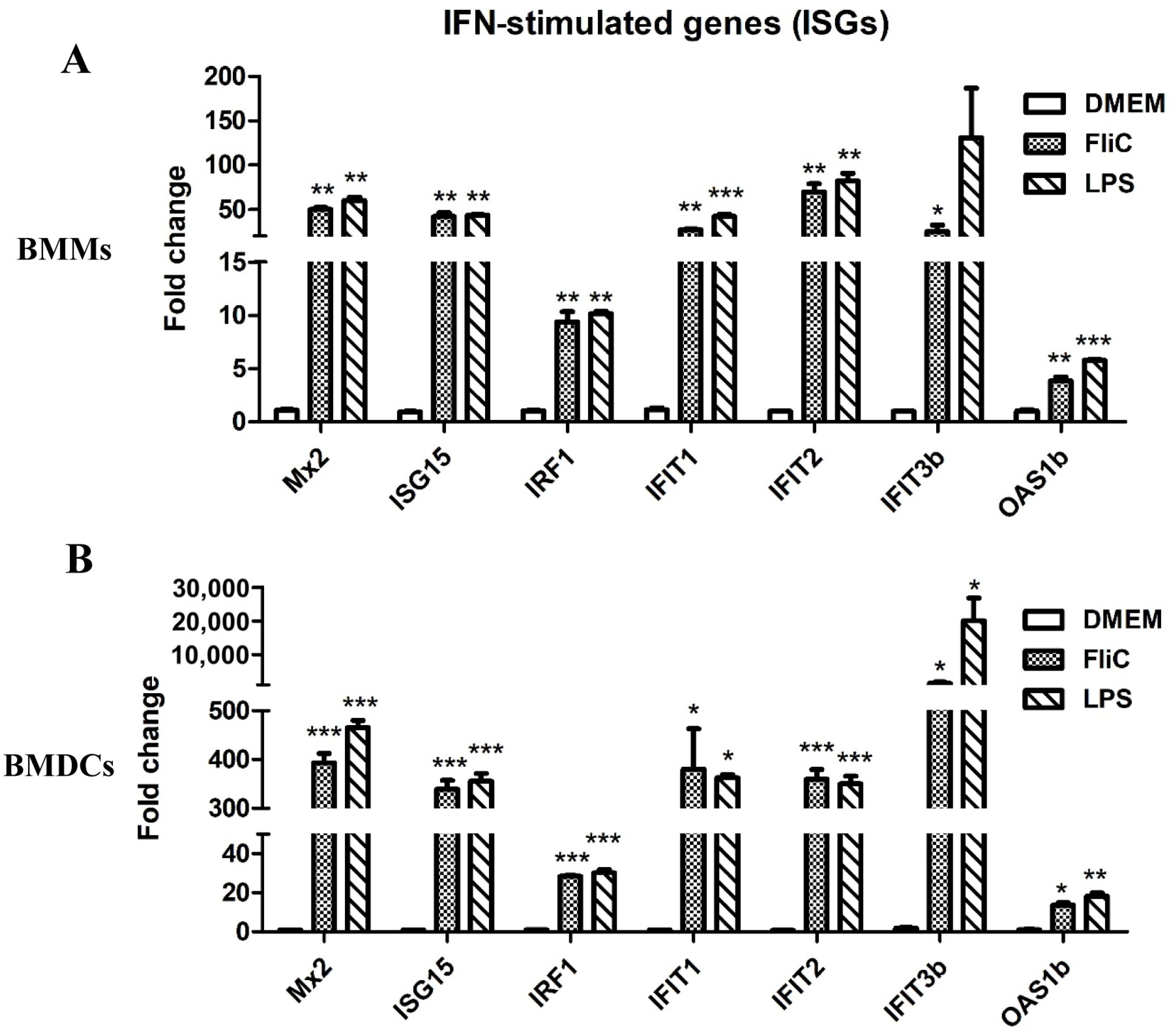
3.10. Flagellin Promotes ISG Expression in BMMs and BMDCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Le, V.P.; Jeong, D.G.; Yoon, S.W.; Kwon, H.M.; Trinh, T.B.N.; Nguyen, T.L.; Bui, T.T.N.; Oh, J.; Kim, J.B.; Cheong, K.M.; et al. Outbreak of African swine fever, Vietnam, 2019. Emerg. Infect. Dis. 2019, 25, 1433–1435. [Google Scholar] [CrossRef] [PubMed]
- Philippon, D.A.M.; Wu, P.; Cowling, B.J.; Lau, E.H.Y. Avian influenza human infections at the human-animal interface. J. Infect. Dis. 2020, 222, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Hajam, I.A.; Dar, P.A.; Shahnawaz, I.; Jaume, J.C.; Lee, J.H. Bacterial flagellin-a potent immunomodulatory agent. Exp. Mol. Med. 2017, 49, e373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.D.; Andersen-Nissen, E.; Hayashi, F.; Strobe, K.; Bergman, M.A.; Barrett, S.L.; Cookson, B.T.; Aderem, A. Toll-like receptor 5 recognizes a conserved site on flagellin required for protofilament formation and bacterial motility. Nat. Immunol. 2003, 4, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Kay, C.; Wang, R.; Kirkby, M.; Man, S.M. Molecular mechanisms activating the NAIP-NLRC4 inflammasome: Implications in infectious disease, autoinflammation, and cancer. Immunol. Rev. 2020, 297, 67–82. [Google Scholar] [CrossRef]
- Tran, H.Q.; Ley, R.E.; Gewirtz, A.T.; Chassaing, B. Flagellin-elicited adaptive immunity suppresses flagellated microbiota and vaccinates against chronic inflammatory diseases. Nat. Commun. 2019, 10, 5650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, L.; Xiong, D.; Song, H.; Wu, L.; Zhang, M.; Kang, X.; Pan, Z.; Jiao, X. Mucosal and systemic immune responses to influenza H7N9 antigen HA1-2 co-delivered intranasally with flagellin or polyethyleneimine in mice and chickens. Front. Immunol. 2017, 8, 326. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Xiong, D.; Hu, M.; Kang, X.; Pan, Z.; Jiao, X. Enhanced humoural and cellular immune responses to influenza H7N9 antigen HA1-2 fused with flagellin in chickens. BMC Vet. Res. 2017, 13, 190. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr. Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb. Symp. Quant. Biol. 1989, 54, 1–13. [Google Scholar] [CrossRef]
- Duan, T.; Du, Y.; Xing, C.; Wang, H.Y.; Wang, R.F. Toll-like receptor signaling and its role in cell-mediated immunity. Front. Immunol. 2022, 13, 812774. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 2004, 5, 987–995. [Google Scholar] [CrossRef]
- Zhang, L.; Pan, Z.; Kang, X.; Yang, Y.; Kang, H.; Zhang, N.; Rosati, J.M.; Jiao, X. Amino acids 89-96 of Salmonella typhimurium flagellin represent the major domain responsible for TLR5-independent adjuvanticity in the humoral immune response. Cell. Mol. Immunol. 2015, 12, 625–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Yglesias, A.H.; Zhao, X.; Quarles, E.K.; Lai, M.A.; VandenBos, T.; Strong, R.K.; Smith, K.D. Flagellin induces antibody responses through a TLR5- and inflammasome-independent pathway. J. Immunol. 2014, 192, 1587–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilhelm, B.T.; Marguerat, S.; Watt, S.; Schubert, F.; Wood, V.; Goodhead, I.; Penkett, C.J.; Rogers, J.; Bähler, J. Dynamic repertoire of a eukaryotic transcriptome surveyed at single-nucleotide resolution. Nature 2008, 453, 1239–1243. [Google Scholar] [CrossRef]
- Ibrahim, G.F.; Fleet, G.H.; Lyons, M.J.; Walker, R.A. Method for the isolation of highly purified Salmonella flagellins. J. Clin. Microbiol. 1985, 22, 1040–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, D.; Song, L.; Geng, S.; Jiao, Y.; Zhou, X.; Song, H.; Kang, X.; Zhou, Y.; Xu, X.; Sun, J.; et al. Salmonella coiled-coil- and TIR-containing TcpS evades the innate immune system and subdues inflammation. Cell Rep. 2019, 28, 804–818.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.Y.; Lu, J.H. Autophagy and macrophage functions: Inflammatory response and phagocytosis. Cells 2019, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Scheibel, M.; Klein, B.; Merkle, H.; Schulz, M.; Fritsch, R.; Greten, F.R.; Arkan, M.C.; Schneider, G.; Schmid, R.M. IkappaBbeta is an essential co-activator for LPS-induced IL-1beta transcription in vivo. J. Exp. Med. 2010, 207, 2621–2630. [Google Scholar] [CrossRef] [Green Version]
- Racioppi, L.; Noeldner, P.K.; Lin, F.; Arvai, S.; Means, A.R. Calcium/calmodulin-dependent protein kinase kinase 2 regulates macrophage-mediated inflammatory responses. J. Biol. Chem. 2012, 287, 11579–11591. [Google Scholar] [CrossRef] [Green Version]
- Huynh, K.K.; Kay, J.G.; Stow, J.L.; Grinstein, S. Fusion, fission, and secretion during phagocytosis. Physiology 2007, 22, 366–372. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.; Park, A.; Huh, J.W.; You, G.; Jung, D.J.; Song, M.; Lee, H.K.; Kim, Y.M. Flagellin-stimulated production of interferon-β promotes anti-flagellin IgG2c and IgA responses. Mol. Cells 2020, 43, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Higa-Nakamine, S.; Noguchi, N.; Maeda, N.; Toku, S.; Isohama, Y.; Sugahara, K.; Kukita, I.; Yamamoto, H. Induction of epithelial-mesenchymal transition by flagellin in cultured lung epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 303, L1057–69. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.G.; Kim, K.H.; Basom, R.S.; Gharib, S.A. Plasticity of airway epithelial cell transcriptome in response to flagellin. PLoS ONE 2015, 10, e0115486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, E.; Jung, J.; Rhee, S.H. Toll-like receptor 5 engagement induces interleukin-17C expression in intestinal epithelial cells. J. Interferon Cytokine Res. 2012, 32, 583–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, N.; Sang Yoon, G.; Liu, X.; Mi, X.; Chen, W.; Standiford, T.J.; Yu, F.S. Genome-wide transcriptional analysis of differentially expressed genes in flagellin-pretreated mouse corneal epithelial cells in response to Pseudomonas aeruginosa: Involvement of S100A8/A9. Mucosal Immunol. 2013, 6, 993–1005. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.X.; Tseng, J.C.; Yu, G.Y.; Luo, Y.; Huang, C.F.; Hong, Y.R.; Chuang, T.H. Recent advances in the development of toll-like receptor agonist-based vaccine adjuvants for infectious diseases. Pharmaceutics 2022, 14, 423. [Google Scholar] [CrossRef]
- Vijayan, A.; Rumbo, M.; Carnoy, C.; Sirard, J.C. Compartmentalized antimicrobial defenses in response to flagellin. Trends Microbiol. 2018, 26, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Sierro, F.; Dubois, B.; Coste, A.; Kaiserlian, D.; Kraehenbuhl, J.P.; Sirard, J.C. Flagellin stimulation of intestinal epithelial cells triggers CCL20-mediated migration of dendritic cells. Proc. Natl. Acad. Sci. USA 2001, 98, 13722–13727. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yang, J.; Shi, J.; Gong, Y.N.; Lu, Q.; Xu, H.; Liu, L.; Shao, F. The NLRC4 inflammasome receptors for bacterial flagellin and type III secretion apparatus. Nature 2011, 477, 596–600. [Google Scholar] [CrossRef]
- Gram, A.M.; Wright, J.A.; Pickering, R.J.; Lam, N.L.; Booty, L.M.; Webster, S.J.; Bryant, C.E. Salmonella flagellin activates NAIP/NLRC4 and canonical NLRP3 inflammasomes in human macrophages. J. Immunol. 2021, 206, 631–640. [Google Scholar] [CrossRef]
- Yoon, S.I.; Kurnasov, O.; Natarajan, V.; Hong, M.; Gudkov, A.V.; Osterman, A.L.; Wilson, I.A. Structural basis of TLR5-flagellin recognition and signaling. Science 2012, 335, 859–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijay-Kumar, M.; Aitken, J.D.; Gewirtz, A.T. Toll like receptor-5: Protecting the gut from enteric microbes. Semin. Immunopathol. 2008, 30, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Kofoed, E.M.; Vance, R.E. Innate immune recognition of bacterial ligands by NAIPs determines inflammasome specificity. Nature 2011, 477, 592–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lightfield, K.L.; Persson, J.; Brubaker, S.W.; Witte, C.E.; von Moltke, J.; Dunipace, E.A.; Henry, T.; Sun, Y.H.; Cado, D.; Dietrich, W.F.; et al. Critical function for Naip5 in inflammasome activation by a conserved carboxy-terminal domain of flagellin. Nat. Immunol. 2008, 9, 1171–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, E.A.; Alpuche-Aranda, C.M.; Dors, M.; Clark, A.E.; Bader, M.W.; Miller, S.I.; Aderem, A. Cytoplasmic flagellin activates caspase-1 and secretion of interleukin 1beta via Ipaf. Nat. Immunol. 2006, 7, 569–575. [Google Scholar] [CrossRef]
- Lin, Y.J.; Papp, G.; Miskey, C.; Fiedler, A.; Goretzki, A.; Wolfheimer, S.; Zimmermann, J.; Crauwels, P.; Ivics, Z.; van Zandbergen, G.; et al. The flagellin: Allergen fusion protein rFlaA:Betv1 induces a MyD88- and MAPK-dependent activation of glucose metabolism in macrophages. Cells 2021, 10, 2614. [Google Scholar] [CrossRef]
- Kortmann, J.; Brubaker, S.W.; Monack, D.M. Cutting edge: Inflammasome activation in primary human macrophages is dependent on flagellin. J. Immunol. 2015, 195, 815–819. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Wang, X.; Saredy, J.; Yuan, Z.; Yang, X.; Wang, H. Innate-adaptive immunity interplay and redox regulation in immune response. Redox Biol. 2020, 37, 101759. [Google Scholar] [CrossRef]
- George, P.J.; Marches, R.; Nehar-Belaid, D.; Banchereau, J.; Lustigman, S. The Th1/Tfh-like biased responses elicited by the rASP-1 innate adjuvant are dependent on TRIF and Type I IFN receptor pathways. Front. Immunol. 2022, 13, 961094. [Google Scholar] [CrossRef]
- Le Bon, A.; Schiavoni, G.; D’Agostino, G.; Gresser, I.; Belardelli, F.; Tough, D.F. Type i interferons potently enhance humoral immunity and can promote isotype switching by stimulating dendritic cells in vivo. Immunity 2001, 14, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Duong, E.; Fessenden, T.B.; Lutz, E.; Dinter, T.; Yim, L.; Blatt, S.; Bhutkar, A.; Wittrup, K.D.; Spranger, S. Type I interferon activates MHC class I-dressed CD11b+ conventional dendritic cells to promote protective anti-tumor CD8+ T cell immunity. Immunity 2022, 55, 308–323.e9. [Google Scholar] [CrossRef] [PubMed]
- Coccia, E.M.; Battistini, A. Early IFN type I response: Learning from microbial evasion strategies. Semin. Immunol. 2015, 27, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Sugrue, J.A.; Bourke, N.M.; O’Farrelly, C. Type I interferon and the spectrum of susceptibility to viral infection and autoimmune disease: A shared genomic signature. Front. Immunol. 2021, 12, 757249. [Google Scholar] [CrossRef] [PubMed]
- Crouse, J.; Kalinke, U.; Oxenius, A. Regulation of antiviral T cell responses by type I interferons. Nat. Rev. Immunol. 2015, 15, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Horisberger, M.A.; Staeheli, P.; Haller, O. Interferon induces a unique protein in mouse cells bearing a gene for resistance to influenza virus. Proc. Natl. Acad. Sci. USA 1983, 80, 1910–1914. [Google Scholar] [CrossRef] [Green Version]
- Diamond, M.S.; Farzan, M. The broad-spectrum antiviral functions of IFIT and IFITM proteins. Nat. Rev. Immunol. 2013, 13, 46–57. [Google Scholar] [CrossRef]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef]
- Steinman, R.M.; Banchereau, J. Taking dendritic cells into medicine. Nature 2007, 449, 419–426. [Google Scholar] [CrossRef]
- Patel, A.A.; Ginhoux, F.; Yona, S. Monocytes, macrophages, dendritic cells and neutrophils: An update on lifespan kinetics in health and disease. Immunology 2021, 163, 250–261. [Google Scholar] [CrossRef]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of type I interferon responses. Nat. Rev. Immunol. 2014, 14, 36–49. [Google Scholar] [CrossRef] [Green Version]
- Schoggins, J.W. Interferon-stimulated genes: What do they all do? Annu. Rev. Virol. 2019, 6, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Xia, P.; Li, S.; Zhang, T.; Wang, T.T.; Zhu, J. RNA sensors of the innate immune system and their detection of pathogens. IUBMB Life 2017, 69, 297–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Product Size (bp) | Accession No. |
|---|---|---|---|---|
| IL-1β | gcccatcctctgtgactcat | aggccacaggtattttgtcg | 230 | NM_008361.4 |
| IL-18 | gacagcctgtgttcgaggat | tggatccatttcctcaaagg | 188 | NM_008360.2 |
| IL-6 | agttgccttcttgggactga | tccacgatttcccagagaac | 159 | NM_031168.2 |
| TNF-α | agcccccagtctgtatcctt | ctccctttgcagaactcagg | 212 | NM_013693.3 |
| CCL2 | aggtccctgtcatgcttctg | tctggacccattccttcttg | 249 | NM_011333.3 |
| CCL3 | agattccacgccaattcatc | ctcaagcccctgctctacac | 223 | NM_011337.2 |
| CCL4 | cccacttcctgctgtttctc | gaggaggcctctcctgaagt | 238 | NM_013652.2 |
| CCL5 | tgccaacccagagaagaagt | agatgcccattttcccagga | 184 | NM_013653.3 |
| CXCL2 | aagtttgccttgaccctgaa | aggcacatcaggtacgatcc | 180 | NM_009140.2 |
| CXCL10 | ggatggctgtcctagctctg | ataaccccttgggaagatgg | 208 | NM_008176.3 |
| IFN-β | ccctatggagatgacggaga | ctgtctgctggtggagttca | 161 | NM_010510.1 |
| IRF1 | gcaaaaccaagaggaagctg | cagagagactgctgctgacg | 186 | NM_008390.2 |
| IRF9 | gtctggaagactcgcctacg | tggtcctcccattttccata | 220 | NM_001159417.1 |
| Mx1 | tctgtgcaggcactatgagg | gcctctccactcctctcctt | 247 | NM_010846.1 |
| Mx2 | cccagaggcagtggtattgt | acatttggggagctgacatc | 227 | NM_013606.1 |
| ISG15 | aagaagcagattgcccagaa | tctgcgtcagaaagacctca | 217 | NM_015783.3 |
| ISG20 | ccatggactgtgagatggtg | agcttgcctttcagaagctg | 229 | NM_020583.5 |
| IFIT1 | aggctggagtgtgctgagat | tctggatttaaccggacagc | 224 | NM_008331.3 |
| IFIT2 | caccttcggtatggcaactt | gcaaggcctcagaatcagac | 181 | NM_008332.3 |
| IFIT3b | cgagcaaaaatgtgctttga | gctccccttcagcttcttct | 190 | NM_001005858.3 |
| OAS1b | accgtcttggaactggtcac | atgttccttgttgggtcagc | 155 | NM_001083925.1 |
| GAPDH | aactttggcattgtggaagg | acacattgggggtaggaaca | 223 | NM_001289726.1 |
| hIFN-β | cattacctgaaggccaagga | cagcatctgctggttgaaga | 178 | NM_002176.4 |
| hIL-1β | gggcctcaaggaaaagaatc | ttctgcttgagaggtgctga | 205 | NM_000576.3 |
| hIL-8 | gtgcagttttgccaaggagt | ctctgcacccagttttcctt | 196 | BC013615.1 |
| hGAPDH | gagtcaacggatttggtcgt | ttgattttggagggatctcg | 238 | NM_002046.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, L.; Xiong, D.; Wen, Y.; Tan, R.; Kang, X.; Jiao, X.; Pan, Z. Transcriptome Sequencing Reveals Salmonella Flagellin Activation of Interferon-β-Related Immune Responses in Macrophages. Curr. Issues Mol. Biol. 2023, 45, 2798-2816. https://doi.org/10.3390/cimb45040183
Song L, Xiong D, Wen Y, Tan R, Kang X, Jiao X, Pan Z. Transcriptome Sequencing Reveals Salmonella Flagellin Activation of Interferon-β-Related Immune Responses in Macrophages. Current Issues in Molecular Biology. 2023; 45(4):2798-2816. https://doi.org/10.3390/cimb45040183
Chicago/Turabian StyleSong, Li, Dan Xiong, Yaya Wen, Ruimeng Tan, Xilong Kang, Xinan Jiao, and Zhiming Pan. 2023. "Transcriptome Sequencing Reveals Salmonella Flagellin Activation of Interferon-β-Related Immune Responses in Macrophages" Current Issues in Molecular Biology 45, no. 4: 2798-2816. https://doi.org/10.3390/cimb45040183