
Origin and Emergence of Microglia in the CNS—An Interesting (Hi)story of an Eccentric Cell

 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
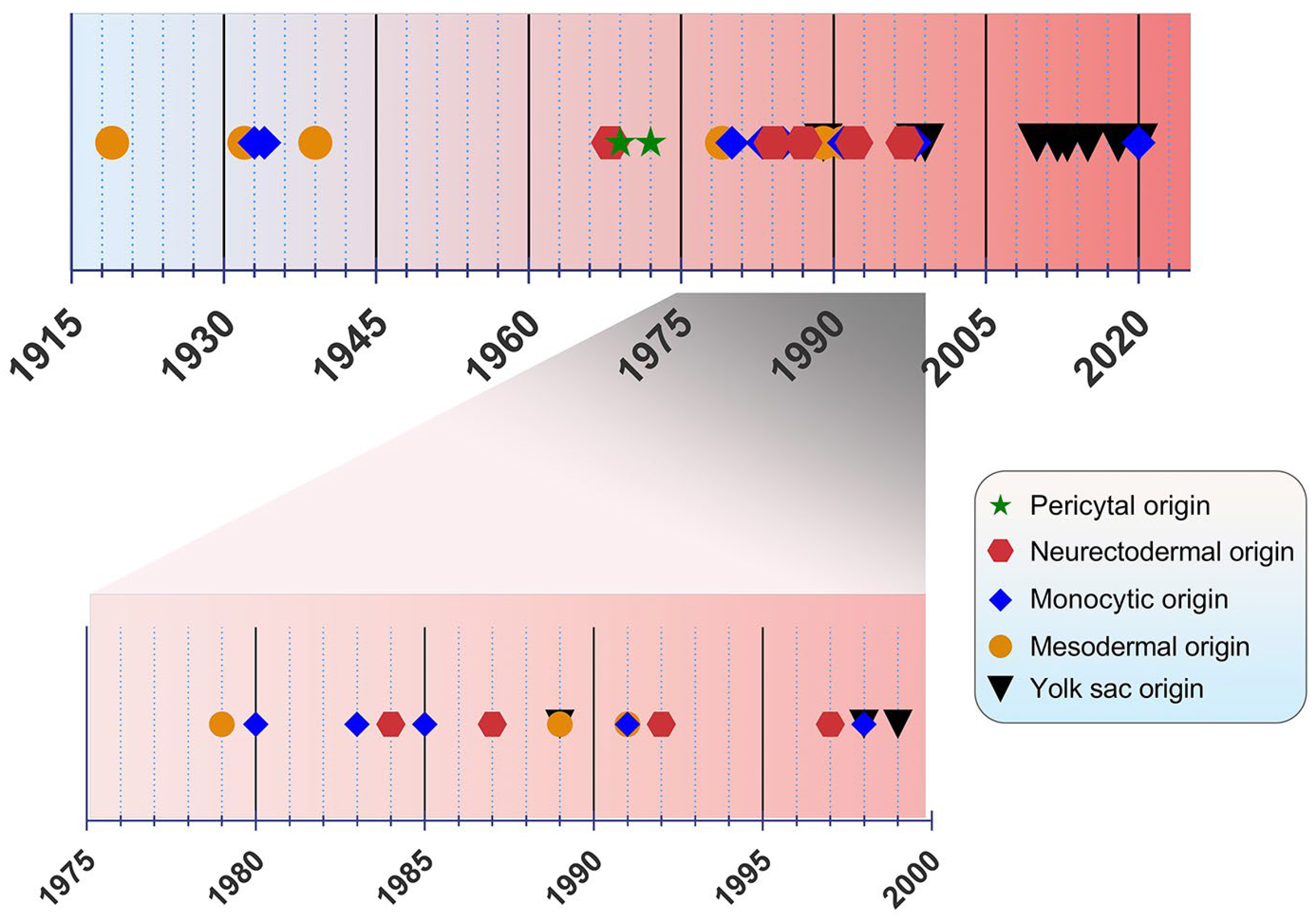
2. Discovery and Ontogeny of Microglia
3. Molecular Cues Orchestrating Microgliogenesis
4. Spatiotemporal Distribution in Various Species
5. Proliferation in the Adult Compromised CNS
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AGM | Aorta-gonad-mesonephros |
| Akt | Protein kinase B |
| ALS | Amyotrophic lateral sclerosis |
| ARID1A | AT-rich interaction domain 1A |
| BAMs | Border-associated macrophages |
| BATF3 | Basic leucine zipper transcriptional factor ATF-like 3 |
| BBB | Blood–brain barrier |
| BDNF | Brain-derived neurotrophic factor |
| C/EBP-α | CCAAT/enhancer-binding protein alpha |
| CCL1 | Chemokine (C-C motif) ligand 1 |
| CCL2 | Chemokine (C-C motif) ligand 2 |
| CCL3 | Chemokine (C-C motif) ligand 3 |
| CCR2 | C-C chemokine receptor type 2 |
| CD11b | Cluster of differentiation molecule 11B |
| CD206 | Cluster of differentiation molecule 206 |
| CD45 | Cluster of differentiation molecule 45 |
| CD64 | Cluster of differentiation molecule 64 |
| CD68 | Cluster of differentiation molecule 68 |
| CNS | Central nervous system |
| CSF-1 | Colony stimulating factor-1 |
| CSF1R | Colony stimulating factor 1 receptor |
| CX3CL1 | Chemokine (C-X3-C motif) ligand 1 |
| CX3CR1 | CX3C motif chemokine receptor 1 |
| CXCL12 | C-X-C motif chemokine ligand 12 |
| CXCR4 | C-X-C chemokine receptor type 4 |
| CXCR7 | C-X-C chemokine receptor type 7 |
| DAP12 | DNAX-activating protein of 12 kDa |
| E | Embryonic day |
| EED | Embryonic ectoderm development |
| EMPs | Erythro-myeloid progenitors |
| Erk1/2 | Extracellular signal-regulated kinase 1/2 |
| G-CSF | Granulocyte colony stimulating factor |
| G-CSFR | Granulocyte colony stimulating factor receptor |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| GM-CSFR | Granulocyte-macrophage colony-stimulating factor receptor |
| GSK3β | Glycogen synthase kinase-3 beta |
| GW | Gestational week |
| HDAC1 | Histone Deacetylase 1 |
| HDAC2 | Histone Deacetylase 2 |
| HOXB8 | Homeobox B8 |
| HSC | Hematopoietic stem cells |
| IBA1 | Ionized calcium binding adaptor molecule 1 |
| ID2 | Inhibitor of DNA binding 2 |
| IL-33 | Interleukin 33 |
| IL-34 | Interleukin 34 |
| IL-5 | Interleukin 5 |
| iNOS | Inducible nitric oxide synthase |
| IRF8 | Interferon regulatory factor 8 |
| KLF4 | Krüppel-like factor 4 |
| M-CSF | Macrophage colony-stimulating factor |
| M-CSFR | Macrophage colony-stimulating factor receptor |
| MAFB | MAF bZIP transcription factor B |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MECP2 | Methyl-CpG binding protein 2 |
| MEIS3 | Meis homeobox 3 |
| MHC II | Major histocompatibility complex class II |
| MIF | Macrophage migration inhibitory factor |
| MIP-1α | Macrophage inflammatory protein-1 alpha |
| miR-124 | microRNA 124 |
| MMP8 | Matrix Metallopeptidase 8 |
| MMP9 | Matrix Metallopeptidase 9 |
| MPO | Myeloperoxidase |
| MYB | MYB proto-oncogene, transcription factor |
| NF-kB | Nuclear factor kappa B |
| NGF | Nerve growth factor |
| NOX2 | NADPH oxidase 2 |
| NR4A1 | Nuclear receptor subfamily 4 group A member 1 |
| NT-3 | Neurotrophin-3 |
| NT-4 | Neurotrophin-4 |
| P2RX7 | P2X purinoceptor 7 |
| P2RY12 | Purinergic receptor P2Y12 |
| PGRN | Progranulin |
| qRT-PCR | Quantitative reverse transcriptase polymerase chain reaction |
| RCA-1 | Ricinus communis agglutinin-1 |
| RUNX1 | Runt-related transcription factor 1 |
| SALL1 | Spalt like transcription factor 1 |
| SALL3 | Spalt like transcription factor 3 |
| SCF | Stem cell factor |
| SVZ | Subventricular zone |
| TGF-β1 | Transforming growth factor beta 1 |
| TGM2 | Transglutaminase 2 |
| TMEM119 | Transmembrane Protein 119 |
| TREM2 | Triggering receptor expressed on myeloid cells 2 |
| VEGFR1 | Vascular endothelial growth factor receptor-1 |
| YS | Yolk sac |
| αVβ8 integrin | Integrin subunit alpha V and beta 8 |
References
- Davies, L.C.; Jenkins, S.J.; Allen, J.E.; Taylor, P.R. Tissue-Resident Macrophages. Nat. Immunol. 2013, 14, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Soulet, D.; Rivest, S. Microglia. Curr. Biol. CB 2008, 18, R506–R508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.; Eo, J.-C.; Lee, C.; Yu, J.-W. Distinct Features of Brain-Resident Macrophages: Microglia and Non-Parenchymal Brain Macrophages. Mol. Cells 2021, 44, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, S.E.; Medeiros, M.; Porfirio, J.; Tavares, W.; Pessôa, L.; Grinberg, L.; Leite, R.E.P.; Ferretti-Rebustini, R.E.L.; Suemoto, C.K.; Filho, W.J.; et al. Similar Microglial Cell Densities across Brain Structures and Mammalian Species: Implications for Brain Tissue Function. J. Neurosci. 2020, 40, 4622–4643. [Google Scholar] [CrossRef] [PubMed]
- Paolicelli, R.C.; Sierra, A.; Stevens, B.; Tremblay, M.-E.; Aguzzi, A.; Ajami, B.; Amit, I.; Audinat, E.; Bechmann, I.; Bennett, M.; et al. Microglia States and Nomenclature: A Field at Its Crossroads. Neuron 2022, 110, 3458–3483. [Google Scholar] [CrossRef]
- Lenz, K.M.; Nelson, L.H. Microglia and Beyond: Innate Immune Cells As Regulators of Brain Development and Behavioral Function. Front. Immunol. 2018, 9, 698. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, M.-È.; Stevens, B.; Sierra, A.; Wake, H.; Bessis, A.; Nimmerjahn, A. The Role of Microglia in the Healthy Brain. J. Neurosci. 2011, 31, 16064–16069. [Google Scholar] [CrossRef] [Green Version]
- Neurons and Neuroglia-ClinicalKey. Available online: https://www.clinicalkey.com/#!/content/book/3-s2.0-B978032328782100438X (accessed on 28 December 2022).
- Chen, Z.; Trapp, B.D. Microglia and Neuroprotection. J. Neurochem. 2016, 136 (Suppl. 1), 10–17. [Google Scholar] [CrossRef]
- Bachiller, S.; Jiménez-Ferrer, I.; Paulus, A.; Yang, Y.; Swanberg, M.; Deierborg, T.; Boza-Serrano, A. Microglia in Neurological Diseases: A Road Map to Brain-Disease Dependent-Inflammatory Response. Front. Cell. Neurosci. 2018, 12, 488. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, B.L.; Sicotte, N.L. Microglia in Multiple Sclerosis: Friend or Foe? Front. Immunol. 2020, 11, 374. [Google Scholar] [CrossRef]
- Clarke, B.E.; Patani, R. The Microglial Component of Amyotrophic Lateral Sclerosis. Brain J. Neurol. 2020, 143, 3526–3539. [Google Scholar] [CrossRef]
- Sierra, A.; de Castro, F.; Del Río-Hortega, J.; Rafael Iglesias-Rozas, J.; Garrosa, M.; Kettenmann, H. The “Big-Bang” for Modern Glial Biology: Translation and Comments on Pío Del Río-Hortega 1919 Series of Papers on Microglia. Glia 2016, 64, 1801–1840. [Google Scholar] [CrossRef]
- Colonna, M.; Butovsky, O. Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annu. Rev. Immunol. 2017, 35, 441–468. [Google Scholar] [CrossRef]
- Sohet, F.; Lin, C.; Munji, R.N.; Lee, S.Y.; Ruderisch, N.; Soung, A.; Arnold, T.D.; Derugin, N.; Vexler, Z.S.; Yen, F.T.; et al. LSR/Angulin-1 Is a Tricellular Tight Junction Protein Involved in Blood–Brain Barrier Formation. J. Cell Biol. 2015, 208, 703–711. [Google Scholar] [CrossRef] [Green Version]
- Hoeffel, G.; Ginhoux, F. Ontogeny of Tissue-Resident Macrophages. Front. Immunol. 2015, 6, 486. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Hirschi, K.K. Tissue-Resident Macrophage Development and Function. Front. Cell Dev. Biol. 2020, 8, 617879. [Google Scholar] [CrossRef]
- Vink, C.S.; Mariani, S.A.; Dzierzak, E. Embryonic Origins of the Hematopoietic System: Hierarchies and Heterogeneity. HemaSphere 2022, 6, e737. [Google Scholar] [CrossRef]
- Yoder, M.C. Inducing Definitive Hematopoiesis in a Dish. Nat. Biotechnol. 2014, 32, 539–541. [Google Scholar] [CrossRef]
- Ling, E.A. Transformation of Monocytes into Amoeboid Microglia in the Corpus Callosum of Postnatal Rats, as Shown by Labelling Monocytes by Carbon Particles. J. Anat. 1979, 128, 847–858. [Google Scholar]
- Rio-Hortega, P. The Microglia. Lancet 1939, 233, 1023–1026. [Google Scholar] [CrossRef]
- Kershman, J. Genesis of Microglia in the Human Brain. Arch. Neurol. Psychiatry 1939, 41, 24–50. [Google Scholar] [CrossRef]
- Boya, J.; Calvo, J.; Prado, A. The Origin of Microglial Cells. J. Anat. 1979, 129, 177–186. [Google Scholar] [PubMed]
- Boya, J.; Calvo, J.L.; Carbonell, A.L.; Borregon, A. A Lectin Histochemistry Study on the Development of Rat Microglial Cells. J. Anat. 1991, 175, 229–236. [Google Scholar] [PubMed]
- Dalmau, I.; Finsen, B.; Tønder, N.; Zimmer, J.; González, B.; Castellano, B. Development of Microglia in the Prenatal Rat Hippocampus. J. Comp. Neurol. 1997, 377, 70–84. [Google Scholar] [CrossRef]
- Barón, M.; Gallego, A. The Relation of the Microglia with the Pericytes in the Cat Cerebral Cortex. Z. Zellforsch. Mikrosk. Anat. 1972, 128, 42–57. [Google Scholar] [CrossRef]
- Mori, S.; Leblond, C.P. Identification of Microglia in Light and Electron Microscopy. J. Comp. Neurol. 1969, 135, 57–80. [Google Scholar] [CrossRef]
- Juba, A. Untersuchungen Über Die Entwicklung Der Hortegaschen Microglia Des Menschen. Arch. Psychiat. Νervenkr. 1933, 101, 577–592. [Google Scholar] [CrossRef]
- Santha, K.; Juba, A. Weitere Untersuchungen Über Die Entwicklung Der Hortegaschen Mikroglia. Arch. Psychiat. Νervenkr. 1933, 98, 598–613. [Google Scholar] [CrossRef]
- Perry, V.H.; Hume, D.A.; Gordon, S. Immunohistochemical Localization of Macrophages and Microglia in the Adult and Developing Mouse Brain. Neuroscience 1985, 15, 313–326. [Google Scholar] [CrossRef]
- Ling, E.A.; Penney, D.; Leblond, C.P. Use of Carbon Labeling to Demonstrate the Role of Blood Monocytes as Precursors of the “ameboid Cells” Present in the Corpus Callosum of Postnatal Rats. J. Comp. Neurol. 1980, 193, 631–657. [Google Scholar] [CrossRef]
- Murabe, Y.; Sano, Y. Morphological Studies on Neuroglia. VII. Distribution of “Brain Macrophages” in Brains of Neonatal and Adult Rats, as Determined by Means of Immunohistochemistry. Cell Tissue Res. 1983, 229, 85–95. [Google Scholar] [CrossRef]
- Ling, E.A.; Kaur, C.; Wong, W.C. Expression of Major Histocompatibility Complex and Leukocyte Common Antigens in Amoeboid Microglia in Postnatal Rats. J. Anat. 1991, 177, 117–126. [Google Scholar]
- Oehmichen, M.; Wiethölter, H.; Greaves, M.F. Immunological Analysis of Human Microglia: Lack of Monocytic and Lymphoid Membrane Differentiation Antigens. J. Neuropathol. Exp. Neurol. 1979, 38, 99–103. [Google Scholar] [CrossRef]
- Lewis, P.D. The Fate of the Subependymal Cell in the Adult Rat Brain, with a Note on the Origin of Microglia. Brain J. Neurol. 1968, 91, 721–736. [Google Scholar] [CrossRef]
- Kitamura, T.; Miyake, T.; Fujita, S. Genesis of Resting Microglia in the Gray Matter of Mouse Hippocampus. J. Comp. Neurol. 1984, 226, 421–433. [Google Scholar] [CrossRef]
- Fedoroff, S.; Zhai, R.; Novak, J.P. Microglia and Astroglia Have a Common Progenitor Cell. J. Neurosci. Res. 1997, 50, 477–486. [Google Scholar] [CrossRef]
- de Groot, C.J.; Huppes, W.; Sminia, T.; Kraal, G.; Dijkstra, C.D. Determination of the Origin and Nature of Brain Macrophages and Microglial Cells in Mouse Central Nervous System, Using Non-Radioactive in Situ Hybridization and Immunoperoxidase Techniques. Glia 1992, 6, 301–309. [Google Scholar] [CrossRef]
- Schelper, R.L.; Adrian, E.K. Monocytes Become Macrophages; They Do Not Become Microglia: A Light and Electron Microscopic Autoradiographic Study Using 125-Iododeoxyuridine. J. Neuropathol. Exp. Neurol. 1986, 45, 1–19. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Fujiwara, M. Absence of Donor-Type Major Histocompatibility Complex Class I Antigen-Bearing Microglia in the Rat Central Nervous System of Radiation Bone Marrow Chimeras. J. Neuroimmunol. 1987, 17, 71–82. [Google Scholar] [CrossRef]
- Hutchins, K.D.; Dickson, D.W.; Rashbaum, W.K.; Lyman, W.D. Localization of Morphologically Distinct Microglial Populations in the Developing Human Fetal Brain: Implications for Ontogeny. Brain Res. Dev. Brain Res. 1990, 55, 95–102. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamamura, F.; Naito, M. Differentiation, Maturation, and Proliferation of Macrophages in the Mouse Yolk Sac: A Light-Microscopic, Enzyme-Cytochemical, Immunohistochemical, and Ultrastructural Study. J. Leukoc. Biol. 1989, 45, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, E.; Miki, A.; Mizoguti, H. Histochemical Study of the Differentiation of Microglial Cells in the Developing Human Cerebral Hemispheres. J. Anat. 1989, 166, 253–264. [Google Scholar] [PubMed]
- Kurz, H.; Christ, B. Embryonic CNS Macrophages and Microglia Do Not Stem from Circulating, but from Extravascular Precursors. Glia 1998, 22, 98–102. [Google Scholar] [CrossRef]
- Andjelkovic, A.V.; Nikolic, B.; Pachter, J.S.; Zecevic, N. Macrophages/Microglial Cells in Human Central Nervous System during Development: An Immunohistochemical Study. Brain Res. 1998, 814, 13–25. [Google Scholar] [CrossRef]
- Alliot, F.; Godin, I.; Pessac, B. Microglia Derive from Progenitors, Originating from the Yolk Sac, and Which Proliferate in the Brain. Brain Res. Dev. Brain Res. 1999, 117, 145–152. [Google Scholar] [CrossRef]
- Ginhoux, F.; Greter, M.; Leboeuf, M.; Nandi, S.; See, P.; Gokhan, S.; Mehler, M.F.; Conway, S.J.; Ng, L.G.; Stanley, E.R.; et al. Fate Mapping Analysis Reveals That Adult Microglia Derive from Primitive Macrophages. Science 2010, 330, 841–845. [Google Scholar] [CrossRef] [Green Version]
- Mizutani, M.; Pino, P.A.; Saederup, N.; Charo, I.F.; Ransohoff, R.M.; Cardona, A.E. The Fractalkine Receptor but Not CCR2 Is Present on Microglia from Embryonic Development throughout Adulthood. J. Immunol. 2012, 188, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Schulz, C.; Gomez Perdiguero, E.; Chorro, L.; Szabo-Rogers, H.; Cagnard, N.; Kierdorf, K.; Prinz, M.; Wu, B.; Jacobsen, S.E.W.; Pollard, J.W.; et al. A Lineage of Myeloid Cells Independent of Myb and Hematopoietic Stem Cells. Science 2012, 336, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Kierdorf, K.; Erny, D.; Goldmann, T.; Sander, V.; Schulz, C.; Perdiguero, E.G.; Wieghofer, P.; Heinrich, A.; Riemke, P.; Hölscher, C.; et al. Microglia Emerge from Erythromyeloid Precursors via Pu.1- and Irf8-Dependent Pathways. Nat. Neurosci. 2013, 16, 273–280. [Google Scholar] [CrossRef]
- Hoeffel, G.; Chen, J.; Lavin, Y.; Low, D.; Almeida, F.F.; See, P.; Beaudin, A.E.; Lum, J.; Low, I.; Forsberg, E.C.; et al. C-Myb(+) Erythro-Myeloid Progenitor-Derived Fetal Monocytes Give Rise to Adult Tissue-Resident Macrophages. Immunity 2015, 42, 665–678. [Google Scholar] [CrossRef] [Green Version]
- Gomez Perdiguero, E.; Klapproth, K.; Schulz, C.; Busch, K.; Azzoni, E.; Crozet, L.; Garner, H.; Trouillet, C.; de Bruijn, M.F.; Geissmann, F.; et al. Tissue-Resident Macrophages Originate from Yolk-Sac-Derived Erythro-Myeloid Progenitors. Nature 2015, 518, 547–551. [Google Scholar] [CrossRef]
- Sheng, J.; Ruedl, C.; Karjalainen, K. Most Tissue-Resident Macrophages Except Microglia Are Derived from Fetal Hematopoietic Stem Cells. Immunity 2015, 43, 382–393. [Google Scholar] [CrossRef] [Green Version]
- De, S.; Van Deren, D.; Peden, E.; Hockin, M.; Boulet, A.; Titen, S.; Capecchi, M.R. Two Distinct Ontogenies Confer Heterogeneity to Mouse Brain Microglia. Dev. Camb. Engl. 2018, 145, dev152306. [Google Scholar] [CrossRef] [Green Version]
- Fehrenbach, M.K.; Tjwa, M.; Bechmann, I.; Krueger, M. Decreased Microglial Numbers in Vav1-Cre+:Dicer Knock-out Mice Suggest a Second Source of Microglia beyond Yolk Sac Macrophages. Ann. Anat. Anat. Anz. 2018, 218, 190–198. [Google Scholar] [CrossRef]
- Utz, S.G.; See, P.; Mildenberger, W.; Thion, M.S.; Silvin, A.; Lutz, M.; Ingelfinger, F.; Rayan, N.A.; Lelios, I.; Buttgereit, A.; et al. Early Fate Defines Microglia and Non-Parenchymal Brain Macrophage Development. Cell 2020, 181, 557–573.e18. [Google Scholar] [CrossRef]
- Chen, H.-R.; Sun, Y.-Y.; Chen, C.-W.; Kuo, Y.-M.; Kuan, I.S.; Li, Z.-R.T.; Short-Miller, J.C.; Smucker, M.R.; Kuan, C.-Y. Fate Mapping via CCR2-CreER Mice Reveals Monocyte-to-Microglia Transition in Development and Neonatal Stroke. Sci. Adv. 2020, 6, eabb2119. [Google Scholar] [CrossRef]
- Bian, Z.; Gong, Y.; Huang, T.; Lee, C.Z.W.; Bian, L.; Bai, Z.; Shi, H.; Zeng, Y.; Liu, C.; He, J.; et al. Deciphering Human Macrophage Development at Single-Cell Resolution. Nature 2020, 582, 571–576. [Google Scholar] [CrossRef]
- Zusso, M.; Methot, L.; Lo, R.; Greenhalgh, A.D.; David, S.; Stifani, S. Regulation of Postnatal Forebrain Amoeboid Microglial Cell Proliferation and Development by the Transcription Factor Runx1. J. Neurosci. 2012, 32, 11285–11298. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Zhang, P.; Hirai, H.; Elf, S.; Yan, X.; Chen, Z.; Koschmieder, S.; Okuno, Y.; Dayaram, T.; Growney, J.D.; et al. PU. 1 Is a Major Downstream Target of AML1 (RUNX1) in Adult Mouse Hematopoiesis. Nat. Genet. 2008, 40, 51–60. [Google Scholar] [CrossRef]
- Goldmann, T.; Wieghofer, P.; Jordão, M.J.C.; Prutek, F.; Hagemeyer, N.; Frenzel, K.; Amann, L.; Staszewski, O.; Kierdorf, K.; Krueger, M.; et al. Origin, Fate and Dynamics of Macrophages at Central Nervous System Interfaces. Nat. Immunol. 2016, 17, 797–805. [Google Scholar] [CrossRef]
- Olson, M.C.; Scott, E.W.; Hack, A.A.; Su, G.H.; Tenen, D.G.; Singh, H.; Simon, M.C. PU. 1 Is Not Essential for Early Myeloid Gene Expression but Is Required for Terminal Myeloid Differentiation. Immunity 1995, 3, 703–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Xue, R.; Hassan, S.; Nguyen, T.M.L.; Wang, T.; Pan, H.; Xu, J.; Liu, Q.; Zhang, W.; Wen, Z. Il34-Csf1r Pathway Regulates the Migration and Colonization of Microglial Precursors. Dev. Cell 2018, 46, 552–563.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buttgereit, A.; Lelios, I.; Yu, X.; Vrohlings, M.; Krakoski, N.R.; Gautier, E.L.; Nishinakamura, R.; Becher, B.; Greter, M. Sall1 Is a Transcriptional Regulator Defining Microglia Identity and Function. Nat. Immunol. 2016, 17, 1397–1406. [Google Scholar] [CrossRef] [PubMed]
- Butovsky, O.; Jedrychowski, M.P.; Moore, C.S.; Cialic, R.; Lanser, A.J.; Gabriely, G.; Koeglsperger, T.; Dake, B.; Wu, P.M.; Doykan, C.E.; et al. Identification of a Unique TGF-β–Dependent Molecular and Functional Signature in Microglia. Nat. Neurosci. 2014, 17, 131–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mass, E.; Ballesteros, I.; Farlik, M.; Halbritter, F.; Günther, P.; Crozet, L.; Jacome-Galarza, C.E.; Händler, K.; Klughammer, J.; Kobayashi, Y.; et al. Specification of Tissue-Resident Macrophages during Organogenesis. Science 2016, 353, aaf4238. [Google Scholar] [CrossRef] [Green Version]
- Nataf, S.; Anginot, A.; Vuaillat, C.; Malaval, L.; Fodil, N.; Chereul, E.; Langlois, J.-B.; Dumontel, C.; Cavillon, G.; Confavreux, C.; et al. Brain and Bone Damage in KARAP/DAP12 Loss-of-Function Mice Correlate with Alterations in Microglia and Osteoclast Lineages. Am. J. Pathol. 2005, 166, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Otero, K.; Turnbull, I.R.; Poliani, P.L.; Vermi, W.; Cerutti, E.; Aoshi, T.; Tassi, I.; Takai, T.; Stanley, S.L.; Miller, M.; et al. Macrophage Colony-Stimulating Factor Induces the Proliferation and Survival of Macrophages via a Pathway Involving DAP12 and Beta-Catenin. Nat. Immunol. 2009, 10, 734–743. [Google Scholar] [CrossRef] [Green Version]
- Lelli, A.; Gervais, A.; Colin, C.; Chéret, C.; de Almodovar, C.R.; Carmeliet, P.; Krause, K.-H.; Boillée, S.; Mallat, M. The NADPH Oxidase Nox2 Regulates VEGFR1/CSF-1R-Mediated Microglial Chemotaxis and Promotes Early Postnatal Infiltration of Phagocytes in the Subventricular Zone of the Mouse Cerebral Cortex. Glia 2013, 61, 1542–1555. [Google Scholar] [CrossRef]
- Arnò, B.; Grassivaro, F.; Rossi, C.; Bergamaschi, A.; Castiglioni, V.; Furlan, R.; Greter, M.; Favaro, R.; Comi, G.; Becher, B.; et al. Neural Progenitor Cells Orchestrate Microglia Migration and Positioning into the Developing Cortex. Nat. Commun. 2014, 5, 5611. [Google Scholar] [CrossRef] [Green Version]
- Lipfert, J.; Ödemis, V.; Wagner, D.-C.; Boltze, J.; Engele, J. CXCR4 and CXCR7 Form a Functional Receptor Unit for SDF-1/CXCL12 in Primary Rodent Microglia. Neuropathol. Appl. Neurobiol. 2013, 39, 667–680. [Google Scholar] [CrossRef]
- Hoshiko, M.; Arnoux, I.; Avignone, E.; Yamamoto, N.; Audinat, E. Deficiency of the Microglial Receptor CX3CR1 Impairs Postnatal Functional Development of Thalamocortical Synapses in the Barrel Cortex. J. Neurosci. 2012, 32, 15106–15111. [Google Scholar] [CrossRef] [Green Version]
- Rezaie, P.; Trillo-Pazos, G.; Greenwood, J.; Everall, I.P.; Male, D.K. Motility and Ramification of Human Fetal Microglia in Culture: An Investigation Using Time-Lapse Video Microscopy and Image Analysis. Exp. Cell Res. 2002, 274, 68–82. [Google Scholar] [CrossRef]
- Ponomarev, E.D.; Veremeyko, T.; Barteneva, N.; Krichevsky, A.M.; Weiner, H.L. MicroRNA-124 Promotes Microglia Quiescence and Suppresses EAE by Deactivating Macrophages via the C/EBP-α-PU.1 Pathway. Nat. Med. 2011, 17, 64–70. [Google Scholar] [CrossRef]
- Gong, M.; Shi, R.; Liu, Y.; Ke, J.; Liu, X.; Du, H.-Z.; Liu, C.-M. Abnormal Microglial Polarization Induced by Arid1a Deletion Leads to Neuronal Differentiation Deficits. Cell Prolif. 2022, 55, e13314. [Google Scholar] [CrossRef]
- Su, L.; Zhang, M.; Ji, F.; Zhao, J.; Wang, Y.; Wang, W.; Zhang, S.; Ma, H.; Wang, Y.; Jiao, J. Microglia Homeostasis Mediated by Epigenetic ARID1A Regulates Neural Progenitor Cells Response and Leads to Autism-like Behaviors. Mol. Psychiatry 2022, 5, 1–5. [Google Scholar] [CrossRef]
- Walsh, C.E.; Hitchcock, P.F. Progranulin Regulates Neurogenesis in the Developing Vertebrate Retina. Dev. Neurobiol. 2017, 77, 1114–1129. [Google Scholar] [CrossRef] [Green Version]
- Arnold, T.D.; Lizama, C.O.; Cautivo, K.M.; Santander, N.; Lin, L.; Qiu, H.; Huang, E.J.; Liu, C.; Mukouyama, Y.; Reichardt, L.F.; et al. Impaired AVβ8 and TGFβ Signaling Lead to Microglial Dysmaturation and Neuromotor Dysfunction. J. Exp. Med. 2019, 216, 900–915. [Google Scholar] [CrossRef]
- Datta, M.; Staszewski, O.; Raschi, E.; Frosch, M.; Hagemeyer, N.; Tay, T.L.; Blank, T.; Kreutzfeldt, M.; Merkler, D.; Ziegler-Waldkirch, S.; et al. Histone Deacetylases 1 and 2 Regulate Microglia Function during Development, Homeostasis, and Neurodegeneration in a Context-Dependent Manner. Immunity 2018, 48, 514–529.e6. [Google Scholar] [CrossRef] [Green Version]
- Jurga, A.M.; Paleczna, M.; Kuter, K.Z. Overview of General and Discriminating Markers of Differential Microglia Phenotypes. Front. Cell. Neurosci. 2020, 14, 198. [Google Scholar] [CrossRef]
- Bohnert, S.; Seiffert, A.; Trella, S.; Bohnert, M.; Distel, L.; Ondruschka, B.; Monoranu, C.-M. TMEM119 as a Specific Marker of Microglia Reaction in Traumatic Brain Injury in Postmortem Examination. Int. J. Leg. Med. 2020, 134, 2167–2176. [Google Scholar] [CrossRef]
- van Wageningen, T.A.; Vlaar, E.; Kooij, G.; Jongenelen, C.A.M.; Geurts, J.J.G.; van Dam, A.-M. Regulation of Microglial TMEM119 and P2RY12 Immunoreactivity in Multiple Sclerosis White and Grey Matter Lesions Is Dependent on Their Inflammatory Environment. Acta Neuropathol. Commun. 2019, 7, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vankriekelsvenne, E.; Chrzanowski, U.; Manzhula, K.; Greiner, T.; Wree, A.; Hawlitschka, A.; Llovera, G.; Zhan, J.; Joost, S.; Schmitz, C.; et al. Transmembrane Protein 119 Is Neither a Specific nor a Reliable Marker for Microglia. Glia 2022, 70, 1170–1190. [Google Scholar] [CrossRef] [PubMed]
- Konishi, H.; Kobayashi, M.; Kunisawa, T.; Imai, K.; Sayo, A.; Malissen, B.; Crocker, P.R.; Sato, K.; Kiyama, H. Siglec-H Is a Microglia-Specific Marker That Discriminates Microglia from CNS-Associated Macrophages and CNS-Infiltrating Monocytes. Glia 2017, 65, 1927–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, T.; Amann, L.; Sankowski, R.; Staszewski, O.; Lenz, M.; Errico, P.D.; Snaidero, N.; Costa Jordão, M.J.; Böttcher, C.; Kierdorf, K.; et al. Novel Hexb-Based Tools for Studying Microglia in the CNS. Nat. Immunol. 2020, 21, 802–815. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.L.; Bennett, F.C.; Liddelow, S.A.; Ajami, B.; Zamanian, J.L.; Fernhoff, N.B.; Mulinyawe, S.B.; Bohlen, C.J.; Adil, A.; Tucker, A.; et al. New Tools for Studying Microglia in the Mouse and Human CNS. Proc. Natl. Acad. Sci. USA 2016, 113, E1738–E1746. [Google Scholar] [CrossRef] [Green Version]
- Butovsky, O.; Siddiqui, S.; Gabriely, G.; Lanser, A.J.; Dake, B.; Murugaiyan, G.; Doykan, C.E.; Wu, P.M.; Gali, R.R.; Iyer, L.K.; et al. Modulating Inflammatory Monocytes with a Unique MicroRNA Gene Signature Ameliorates Murine ALS. J. Clin. Investig. 2012, 122, 3063–3087. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Jia, L.; Liu, C.-C.; Rong, Z.; Zhong, L.; Yang, L.; Chen, X.-F.; Fryer, J.D.; Wang, X.; Zhang, Y.-W.; et al. TREM2 Promotes Microglial Survival by Activating Wnt/β-Catenin Pathway. J. Neurosci. 2017, 37, 1772–1784. [Google Scholar] [CrossRef] [Green Version]
- Matcovitch-Natan, O.; Winter, D.R.; Giladi, A.; Vargas Aguilar, S.; Spinrad, A.; Sarrazin, S.; Ben-Yehuda, H.; David, E.; Zelada González, F.; Perrin, P.; et al. Microglia Development Follows a Stepwise Program to Regulate Brain Homeostasis. Science 2016, 353, aad8670. [Google Scholar] [CrossRef]
- Wittrahm, R.; Takalo, M.; Marttinen, M.; Kuulasmaa, T.; Mäkinen, P.; Kemppainen, S.; Martiskainen, H.; Rauramaa, T.; Pike, I.; Leinonen, V.; et al. MECP2 Increases the Pro-Inflammatory Response of Microglial Cells and Phosphorylation at Serine 423 Regulates Neuronal Gene Expression upon Neuroinflammation. Cells 2021, 10, 860. [Google Scholar] [CrossRef]
- Cronk, J.C.; Derecki, N.C.; Ji, E.; Xu, Y.; Lampano, A.E.; Smirnov, I.; Baker, W.; Norris, G.T.; Marin, I.; Coddington, N.; et al. Methyl-CpG Binding Protein 2 Regulates Microglia and Macrophage Gene Expression in Response to Inflammatory Stimuli. Immunity 2015, 42, 679–691. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-Y.; Deng, Y.-S.; Dai, S.-K.; Mi, T.-W.; Li, R.-Y.; Liu, P.-P.; Liu, C.; He, B.-D.; He, X.-C.; Du, H.-Z.; et al. Loss of Microglial EED Impairs Synapse Density, Learning, and Memory. Mol. Psychiatry 2022, 27, 2999–3009. [Google Scholar] [CrossRef]
- Liu, C.; Gao, X.; Shi, R.-X.; Wang, Y.-Y.; He, X.-C.; Du, H.-Z.; Hu, B.; Jiao, J.; Liu, C.-M.; Teng, Z.-Q. Microglial Transglutaminase 2 Deficiency Causes Impaired Synaptic Remodelling and Cognitive Deficits in Mice. Cell Prolif. 2023, e13439. [Google Scholar] [CrossRef]
- Rigato, C.; Swinnen, N.; Buckinx, R.; Couillin, I.; Mangin, J.-M.; Rigo, J.-M.; Legendre, P.; Le Corronc, H. Microglia Proliferation Is Controlled by P2X7 Receptors in a Pannexin-1-Independent Manner during Early Embryonic Spinal Cord Invasion. J. Neurosci. 2012, 32, 11559–11573. [Google Scholar] [CrossRef] [Green Version]
- Yasuoka, S.; Kawanokuchi, J.; Parajuli, B.; Jin, S.; Doi, Y.; Noda, M.; Sonobe, Y.; Takeuchi, H.; Mizuno, T.; Suzumura, A. Production and Functions of IL-33 in the Central Nervous System. Brain Res. 2011, 1385, 8–17. [Google Scholar] [CrossRef]
- Bartolini, A.; Vigliani, M.-C.; Magrassi, L.; Vercelli, A.; Rossi, F. G-CSF Administration to Adult Mice Stimulates the Proliferation of Microglia but Does Not Modify the Outcome of Ischemic Injury. Neurobiol. Dis. 2011, 41, 640–649. [Google Scholar] [CrossRef]
- Lee, S.C.; Liu, W.; Brosnan, C.F.; Dickson, D.W. GM-CSF Promotes Proliferation of Human Fetal and Adult Microglia in Primary Cultures. Glia 1994, 12, 309–318. [Google Scholar] [CrossRef]
- Liva, S.M.; de Vellis, J. IL-5 Induces Proliferation and Activation of Microglia via an Unknown Receptor. Neurochem. Res. 2001, 26, 629–637. [Google Scholar] [CrossRef]
- Akimoto, N.; Ifuku, M.; Mori, Y.; Noda, M. Effects of Chemokine (C–C Motif) Ligand 1 on Microglial Function. Biochem. Biophys. Res. Commun. 2013, 436, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ma, P.; Guan, Q.; Meng, L.; Su, L.; Wang, L.; Yuan, B. Effect of Chemokine CC Ligand 2 (CCL2) on A-synuclein-induced Microglia Proliferation and Neuronal Apoptosis. Mol. Med. Rep. 2018, 18, 4213–4218. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Geula, C.; Lu, C.; Koziel, H.; Hatcher, L.M.; Roisen, F.J. Neurotrophins Regulate Proliferation and Survival of Two Microglial Cell Lines in Vitro. Exp. Neurol. 2003, 183, 469–481. [Google Scholar] [CrossRef]
- Terashima, T.; Nakae, Y.; Katagi, M.; Okano, J.; Suzuki, Y.; Kojima, H. Stem Cell Factor Induces Polarization of Microglia to the Neuroprotective Phenotype in Vitro. Heliyon 2018, 4, e00837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dermitzakis, I.; Manthou, M.E.; Meditskou, S.; Miliaras, D.; Kesidou, E.; Boziki, M.; Petratos, S.; Grigoriadis, N.; Theotokis, P. Developmental Cues and Molecular Drivers in Myelinogenesis: Revisiting Early Life to Re-Evaluate the Integrity of CNS Myelin. Curr. Issues Mol. Biol. 2022, 44, 3208–3237. [Google Scholar] [CrossRef] [PubMed]
- Lituma, P.J.; Woo, E.; O’Hara, B.F.; Castillo, P.E.; Sibinga, N.E.S.; Nandi, S. Altered Synaptic Connectivity and Brain Function in Mice Lacking Microglial Adapter Protein Iba1. Proc. Natl. Acad. Sci. USA 2021, 118, e2115539118. [Google Scholar] [CrossRef] [PubMed]
- Maksoud, M.J.E.; Tellios, V.; Lu, W.-Y. Nitric Oxide Attenuates Microglia Proliferation by Sequentially Facilitating Calcium Influx through TRPV2 Channels, Activating NFATC2, and Increasing P21 Transcription. Cell Cycle 2021, 20, 417–433. [Google Scholar] [CrossRef] [PubMed]
- The Human Protein Atlas. Available online: https://www.proteinatlas.org/ (accessed on 15 January 2023).
- Home-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/ (accessed on 15 January 2023).
- Ashwell, K. The Distribution of Microglia and Cell Death in the Fetal Rat Forebrain. Brain Res. Dev. Brain Res. 1991, 58, 1–12. [Google Scholar] [CrossRef]
- Sorokin, S.P.; Hoyt, R.F.; Blunt, D.G.; McNelly, N.A. Macrophage Development: II. Early Ontogeny of Macrophage Populations in Brain, Liver, and Lungs of Rat Embryos as Revealed by a Lectin Marker. Anat. Rec. 1992, 232, 527–550. [Google Scholar] [CrossRef]
- Wang, C.C.; Wu, C.H.; Shieh, J.Y.; Wen, C.Y.; Ling, E.A. Immunohistochemical Study of Amoeboid Microglial Cells in Fetal Rat Brain. J. Anat. 1996, 189, 567–574. [Google Scholar]
- Swinnen, N.; Smolders, S.; Avila, A.; Notelaers, K.; Paesen, R.; Ameloot, M.; Brône, B.; Legendre, P.; Rigo, J.-M. Complex Invasion Pattern of the Cerebral Cortex Bymicroglial Cells during Development of the Mouse Embryo. Glia 2013, 61, 150–163. [Google Scholar] [CrossRef]
- Stremmel, C.; Schuchert, R.; Wagner, F.; Thaler, R.; Weinberger, T.; Pick, R.; Mass, E.; Ishikawa-Ankerhold, H.C.; Margraf, A.; Hutter, S.; et al. Yolk Sac Macrophage Progenitors Traffic to the Embryo during Defined Stages of Development. Nat. Commun. 2018, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Kaur, C.; Rathnasamy, G.; Ling, E.-A. Biology of Microglia in the Developing Brain. J. Neuropathol. Exp. Neurol. 2017, 76, 736–753. [Google Scholar] [CrossRef] [Green Version]
- Ling, E.A.; Wong, W.C. The Origin and Nature of Ramified and Amoeboid Microglia: A Historical Review and Current Concepts. Glia 1993, 7, 9–18. [Google Scholar] [CrossRef]
- Esiri, M.M.; al Izzi, M.S.; Reading, M.C. Macrophages, Microglial Cells, and HLA-DR Antigens in Fetal and Infant Brain. J. Clin. Pathol. 1991, 44, 102–106. [Google Scholar] [CrossRef] [Green Version]
- Rezaie, P.; Male, D. Colonisation of the Developing Human Brain and Spinal Cord by Microglia: A Review. Microsc. Res. Tech. 1999, 45, 359–382. [Google Scholar] [CrossRef]
- Rezaie, P.; Dean, A.; Male, D.; Ulfig, N. Microglia in the Cerebral Wall of the Human Telencephalon at Second Trimester. Cereb. Cortex 2005, 15, 938–949. [Google Scholar] [CrossRef] [Green Version]
- Monier, A.; Evrard, P.; Gressens, P.; Verney, C. Distribution and Differentiation of Microglia in the Human Encephalon during the First Two Trimesters of Gestation. J. Comp. Neurol. 2006, 499, 565–582. [Google Scholar] [CrossRef]
- Monier, A.; Adle-Biassette, H.; Delezoide, A.-L.; Evrard, P.; Gressens, P.; Verney, C. Entry and Distribution of Microglial Cells in Human Embryonic and Fetal Cerebral Cortex. J. Neuropathol. Exp. Neurol. 2007, 66, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Verney, C.; Monier, A.; Fallet-Bianco, C.; Gressens, P. Early Microglial Colonization of the Human Forebrain and Possible Involvement in Periventricular White-Matter Injury of Preterm Infants. J. Anat. 2010, 217, 436–448. [Google Scholar] [CrossRef]
- Cuadros, M.A.; Martin, C.; Coltey, P.; Almendros, A.; Navascués, J. First Appearance, Distribution, and Origin of Macrophages in the Early Development of the Avian Central Nervous System. J. Comp. Neurol. 1993, 330, 113–129. [Google Scholar] [CrossRef]
- Cuadros, M.A.; Rodríguez-Ruiz, J.; Calvente, R.; Almendros, A.; Marín-Teva, J.L.; Navascués, J. Microglia Development in the Quail Cerebellum. J. Comp. Neurol. 1997, 389, 390–401. [Google Scholar] [CrossRef]
- Krall, W.J.; Challita, P.M.; Perlmutter, L.S.; Skelton, D.C.; Kohn, D.B. Cells Expressing Human Glucocerebrosidase from a Retroviral Vector Repopulate Macrophages and Central Nervous System Microglia after Murine Bone Marrow Transplantation. Blood 1994, 83, 2737–2748. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, D.W.; Abkowitz, J.L. Kinetics of Central Nervous System Microglial and Macrophage Engraftment: Analysis Using a Transgenic Bone Marrow Transplantation Model. Blood 1997, 90, 986–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eglitis, M.A.; Mezey, É. Hematopoietic Cells Differentiate into Both Microglia and Macroglia in the Brains of Adult Mice. Proc. Natl. Acad. Sci. USA 1997, 94, 4080–4085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priller, J.; Flügel, A.; Wehner, T.; Boentert, M.; Haas, C.A.; Prinz, M.; Fernández-Klett, F.; Prass, K.; Bechmann, I.; de Boer, B.A.; et al. Targeting Gene-Modified Hematopoietic Cells to the Central Nervous System: Use of Green Fluorescent Protein Uncovers Microglial Engraftment. Nat. Med. 2001, 7, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Simard, A.R.; Rivest, S. Bone Marrow Stem Cells Have the Ability to Populate the Entire Central Nervous System into Fully Differentiated Parenchymal Microglia. FASEB J. 2004, 18, 998–1000. [Google Scholar] [CrossRef] [Green Version]
- Asheuer, M.; Pflumio, F.; Benhamida, S.; Dubart-Kupperschmitt, A.; Fouquet, F.; Imai, Y.; Aubourg, P.; Cartier, N. Human CD34+ Cells Differentiate into Microglia and Express Recombinant Therapeutic Protein. Proc. Natl. Acad. Sci. USA 2004, 101, 3557–3562. [Google Scholar] [CrossRef] [Green Version]
- Beck, H.; Voswinckel, R.; Wagner, S.; Ziegelhoeffer, T.; Heil, M.; Helisch, A.; Schaper, W.; Acker, T.; Hatzopoulos, A.K.; Plate, K.H. Participation of Bone Marrow-Derived Cells in Long-Term Repair Processes after Experimental Stroke. J. Cereb. Blood Flow Metab. 2003, 23, 709–717. [Google Scholar] [CrossRef] [Green Version]
- Hess, D.C.; Abe, T.; Hill, W.D.; Studdard, A.M.; Carothers, J.; Masuya, M.; Fleming, P.A.; Drake, C.J.; Ogawa, M. Hematopoietic Origin of Microglial and Perivascular Cells in Brain. Exp. Neurol. 2004, 186, 134–144. [Google Scholar] [CrossRef]
- Djukic, M.; Mildner, A.; Schmidt, H.; Czesnik, D.; Brück, W.; Priller, J.; Nau, R.; Prinz, M. Circulating Monocytes Engraft in the Brain, Differentiate into Microglia and Contribute to the Pathology Following Meningitis in Mice. Brain J. Neurol. 2006, 129, 2394–2403. [Google Scholar] [CrossRef] [Green Version]
- Bechmann, I.; Goldmann, J.; Kovac, A.D.; Kwidzinski, E.; Simbürger, E.; Naftolin, F.; Dirnagl, U.; Nitsch, R.; Priller, J. Circulating Monocytic Cells Infiltrate Layers of Anterograde Axonal Degeneration Where They Transform into Microglia. FASEB J. 2005, 19, 647–649. [Google Scholar] [CrossRef]
- Rodriguez, M.; Alvarez-Erviti, L.; Blesa, F.J.; Rodríguez-Oroz, M.C.; Arina, A.; Melero, I.; Ramos, L.I.; Obeso, J.A. Bone-Marrow-Derived Cell Differentiation into Microglia: A Study in a Progressive Mouse Model of Parkinson’s Disease. Neurobiol. Dis. 2007, 28, 316–325. [Google Scholar] [CrossRef]
- Malm, T.M.; Koistinaho, M.; Pärepalo, M.; Vatanen, T.; Ooka, A.; Karlsson, S.; Koistinaho, J. Bone-Marrow-Derived Cells Contribute to the Recruitment of Microglial Cells in Response to Beta-Amyloid Deposition in APP/PS1 Double Transgenic Alzheimer Mice. Neurobiol. Dis. 2005, 18, 134–142. [Google Scholar] [CrossRef]
- Simard, A.R.; Soulet, D.; Gowing, G.; Julien, J.-P.; Rivest, S. Bone Marrow-Derived Microglia Play a Critical Role in Restricting Senile Plaque Formation in Alzheimer’s Disease. Neuron 2006, 49, 489–502. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.; Theotokis, P.; Lee, J.Y.; Kim, M.J.; Nheu, D.; Ellen, O.; Bedford, T.; Ramanujam, P.; Wright, D.K.; McDonald, S.J.; et al. NgR-Fc Delivered by Hematopoietic Cells Enhances Neurorepair in a Multiple Sclerosis Model. Brain Commun. 2023, in press. [Google Scholar]
- Flügel, A.; Bradl, M.; Kreutzberg, G.W.; Graeber, M.B. Transformation of Donor-Derived Bone Marrow Precursors into Host Microglia during Autoimmune CNS Inflammation and during the Retrograde Response to Axotomy. J. Neurosci. Res. 2001, 66, 74–82. [Google Scholar] [CrossRef]
- Priller, J.; Prinz, M.; Heikenwalder, M.; Zeller, N.; Schwarz, P.; Heppner, F.L.; Aguzzi, A. Early and Rapid Engraftment of Bone Marrow-Derived Microglia in Scrapie. J. Neurosci. 2006, 26, 11753–11762. [Google Scholar] [CrossRef] [Green Version]
- Vallières, L.; Sawchenko, P.E. Bone Marrow-Derived Cells That Populate the Adult Mouse Brain Preserve Their Hematopoietic Identity. J. Neurosci. 2003, 23, 5197–5207. [Google Scholar] [CrossRef] [Green Version]
- Massengale, M.; Wagers, A.J.; Vogel, H.; Weissman, I.L. Hematopoietic Cells Maintain Hematopoietic Fates upon Entering the Brain. J. Exp. Med. 2005, 201, 1579–1589. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Shi, X.Q.; Echeverry, S.; Mogil, J.S.; De Koninck, Y.; Rivest, S. Expression of CCR2 in Both Resident and Bone Marrow-Derived Microglia Plays a Critical Role in Neuropathic Pain. J. Neurosci. 2007, 27, 12396–12406. [Google Scholar] [CrossRef] [Green Version]
- Marchi, S.; Guilbaud, E.; Tait, S.W.G.; Yamazaki, T.; Galluzzi, L. Mitochondrial Control of Inflammation. Nat. Rev. Immunol. 2023, 23, 159–173. [Google Scholar] [CrossRef]
- Joshi, A.U.; Minhas, P.S.; Liddelow, S.A.; Haileselassie, B.; Andreasson, K.I.; Dorn, G.W.; Mochly-Rosen, D. Fragmented Mitochondria Released from Microglia Trigger A1 Astrocytic Response and Propagate Inflammatory Neurodegeneration. Nat. Neurosci. 2019, 22, 1635–1648. [Google Scholar] [CrossRef]
- Leong, S.K.; Ling, E.A. Amoeboid and Ramified Microglia: Their Interrelationship and Response to Brain Injury. Glia 1992, 6, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Lassmann, H.; Schmied, M.; Vass, K.; Hickey, W.F. Bone Marrow Derived Elements and Resident Microglia in Brain Inflammation. Glia 1993, 7, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Dalmau, I.; Vela, J.M.; González, B.; Finsen, B.; Castellano, B. Dynamics of Microglia in the Developing Rat Brain. J. Comp. Neurol. 2003, 458, 144–157. [Google Scholar] [CrossRef]
- Lawson, L.J.; Perry, V.H.; Gordon, S. Turnover of Resident Microglia in the Normal Adult Mouse Brain. Neuroscience 1992, 48, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Solomon, J.N.; Lewis, C.-A.B.; Ajami, B.; Corbel, S.Y.; Rossi, F.M.V.; Krieger, C. Origin and Distribution of Bone Marrow-Derived Cells in the Central Nervous System in a Mouse Model of Amyotrophic Lateral Sclerosis. Glia 2006, 53, 744–753. [Google Scholar] [CrossRef]
- Ajami, B.; Bennett, J.L.; Krieger, C.; Tetzlaff, W.; Rossi, F.M.V. Local Self-Renewal Can Sustain CNS Microglia Maintenance and Function throughout Adult Life. Nat. Neurosci. 2007, 10, 1538–1543. [Google Scholar] [CrossRef]
- Mildner, A.; Schmidt, H.; Nitsche, M.; Merkler, D.; Hanisch, U.-K.; Mack, M.; Heikenwalder, M.; Brück, W.; Priller, J.; Prinz, M. Microglia in the Adult Brain Arise from Ly-6ChiCCR2+ Monocytes Only under Defined Host Conditions. Nat. Neurosci. 2007, 10, 1544–1553. [Google Scholar] [CrossRef]
- Capotondo, A.; Milazzo, R.; Politi, L.S.; Quattrini, A.; Palini, A.; Plati, T.; Merella, S.; Nonis, A.; di Serio, C.; Montini, E.; et al. Brain Conditioning Is Instrumental for Successful Microglia Reconstitution Following Hematopoietic Stem Cell Transplantation. Proc. Natl. Acad. Sci. USA 2012, 109, 15018–15023. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Pang, S.; Yu, Y.; Wu, X.; Guo, J.; Zhang, S. Proliferation of Parenchymal Microglia Is the Main Source of Microgliosis after Ischaemic Stroke. Brain J. Neurol. 2013, 136, 3578–3588. [Google Scholar] [CrossRef] [Green Version]
- Bruttger, J.; Karram, K.; Wörtge, S.; Regen, T.; Marini, F.; Hoppmann, N.; Klein, M.; Blank, T.; Yona, S.; Wolf, Y.; et al. Genetic Cell Ablation Reveals Clusters of Local Self-Renewing Microglia in the Mammalian Central Nervous System. Immunity 2015, 43, 92–106. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhu, L.; He, S.; Wu, Y.; Jin, W.; Yu, T.; Qu, J.Y.; Wen, Z. Temporal-Spatial Resolution Fate Mapping Reveals Distinct Origins for Embryonic and Adult Microglia in Zebrafish. Dev. Cell 2015, 34, 632–641. [Google Scholar] [CrossRef] [Green Version]
- Tay, T.L.; Mai, D.; Dautzenberg, J.; Fernández-Klett, F.; Lin, G.; Sagar, N.; Datta, M.; Drougard, A.; Stempfl, T.; Ardura-Fabregat, A.; et al. A New Fate Mapping System Reveals Context-Dependent Random or Clonal Expansion of Microglia. Nat. Neurosci. 2017, 20, 793–803. [Google Scholar] [CrossRef]
- Cronk, J.C.; Filiano, A.J.; Louveau, A.; Marin, I.; Marsh, R.; Ji, E.; Goldman, D.H.; Smirnov, I.; Geraci, N.; Acton, S.; et al. Peripherally Derived Macrophages Can Engraft the Brain Independent of Irradiation and Maintain an Identity Distinct from Microglia. J. Exp. Med. 2018, 215, 1627–1647. [Google Scholar] [CrossRef] [Green Version]
- Lund, H.; Pieber, M.; Parsa, R.; Han, J.; Grommisch, D.; Ewing, E.; Kular, L.; Needhamsen, M.; Espinosa, A.; Nilsson, E.; et al. Competitive Repopulation of an Empty Microglial Niche Yields Functionally Distinct Subsets of Microglia-like Cells. Nat. Commun. 2018, 9, 4845. [Google Scholar] [CrossRef] [Green Version]
- Shemer, A.; Grozovski, J.; Tay, T.L.; Tao, J.; Volaski, A.; Süß, P.; Ardura-Fabregat, A.; Gross-Vered, M.; Kim, J.-S.; David, E.; et al. Engrafted Parenchymal Brain Macrophages Differ from Microglia in Transcriptome, Chromatin Landscape and Response to Challenge. Nat. Commun. 2018, 9, 5206. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Xu, Z.; Xiong, S.; Sun, F.; Qin, G.; Hu, G.; Wang, J.; Zhao, L.; Liang, Y.-X.; Wu, T.; et al. Repopulated Microglia Are Solely Derived from the Proliferation of Residual Microglia after Acute Depletion. Nat. Neurosci. 2018, 21, 530–540. [Google Scholar] [CrossRef]
- Elmore, M.R.P.; Najafi, A.R.; Koike, M.A.; Dagher, N.N.; Spangenberg, E.E.; Rice, R.A.; Kitazawa, M.; Matusow, B.; Nguyen, H.; West, B.L.; et al. Colony-Stimulating Factor 1 Receptor Signaling Is Necessary for Microglia Viability, Unmasking a Microglia Progenitor Cell in the Adult Brain. Neuron 2014, 82, 380–397. [Google Scholar] [CrossRef] [Green Version]
- Zhan, L.; Krabbe, G.; Du, F.; Jones, I.; Reichert, M.C.; Telpoukhovskaia, M.; Kodama, L.; Wang, C.; Cho, S.-H.; Sayed, F.; et al. Proximal Recolonization by Self-Renewing Microglia Re-Establishes Microglial Homeostasis in the Adult Mouse Brain. PLoS Biol. 2019, 17, e3000134. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Saito, T.; Saido, T.C.; Barron, A.M.; Ruedl, C. Microglia and CD206+ Border-Associated Mouse Macrophages Maintain Their Embryonic Origin during Alzheimer’s Disease. eLife 2021, 10, e71879. [Google Scholar] [CrossRef]
- Luo, C.; Jian, C.; Liao, Y.; Huang, Q.; Wu, Y.; Liu, X.; Zou, D.; Wu, Y. The Role of Microglia in Multiple Sclerosis. Neuropsychiatr. Dis. Treat. 2017, 13, 1661–1667. [Google Scholar] [CrossRef] [Green Version]
- Schetters, S.T.T.; Gomez-Nicola, D.; Garcia-Vallejo, J.J.; Van Kooyk, Y. Neuroinflammation: Microglia and T Cells Get Ready to Tango. Front. Immunol. 2018, 8, 1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goddery, E.N.; Fain, C.E.; Lipovsky, C.G.; Ayasoufi, K.; Yokanovich, L.T.; Malo, C.S.; Khadka, R.H.; Tritz, Z.P.; Jin, F.; Hansen, M.J.; et al. Microglia and Perivascular Macrophages Act as Antigen Presenting Cells to Promote CD8 T Cell Infiltration of the Brain. Front. Immunol. 2021, 12, 726421. [Google Scholar] [CrossRef] [PubMed]
- O’Loughlin, E.; Madore, C.; Lassmann, H.; Butovsky, O. Microglial Phenotypes and Functions in Multiple Sclerosis. Cold Spring Harb. Perspect. Med. 2018, 8, a028993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nheu, D.; Ellen, O.; Ye, S.; Ozturk, E.; Pagnin, M.; Kertadjaja, S.; Theotokis, P.; Grigoriadis, N.; McLean, C.; Petratos, S. Modulation of the Microglial Nogo-A/NgR Signaling Pathway as a Therapeutic Target for Multiple Sclerosis. Cells 2022, 11, 3768. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene | Locus | Protein | Species | Biological Role | Ref. |
|---|---|---|---|---|---|
| BDNF | 11p14.1 | Brain derived neurotrophic factor | Mice | Proliferation | [101] |
| CCL1 | 17q12 | C-C motif chemokine ligand 1 | Mice | Migration; Proliferation | [99] |
| CCL2 | 17q12 | C-C motif chemokine ligand 2 | Human; Mice | Migration; Proliferation; Terminal differentiation | [73,100] |
| CCL3 | 17q12 | C-C motif chemokine ligand 3 | Human | Migration; Terminal differentiation | [73] |
| CSF1 | 1p13.3 | Colony stimulating factor 1 | Mice | Proliferation; Terminal differentiation | [47,68] |
| CSF1R | 5q32 | Colony stimulating factor 1 receptor | Mice; Zebrafish | Migration; Terminal differentiation | [47,63] |
| CX3CL1 | 16q21 | C-X3-C motif chemokine ligand 1 | Mice | Migration | [72] |
| CX3CR1 | 3p22.2 | C-X3-C motif chemokine receptor 1 | Mice | Migration | [72] |
| CXCL12 | 10q11.21 | C-X-C motif chemokine ligand 12 | Mice; Rat | Migration; Proliferation | [70,71] |
| CXCR4 | 2q22.1 | C-X-C motif chemokine receptor 4 | Mice; Rat | Migration; Proliferation | [70,71] |
| CXCR7 | 2q37.3 | C-X-C chemokine receptor type 7 | Rat | Migration; Proliferation | [71] |
| DAP12 | 19q13.12 | DNAX-activating protein of 12 kDa | Mice | Proliferation; Terminal differentiation | [67,68] |
| G-CSF | 17q21.1 | Granulocyte colony-stimulating factor | Mice | Proliferation | [96] |
| GM-CSF | 5q31.1 | Granulocyte-macrophage colony-stimulating factor | Human | Proliferation | [97] |
| HDAC1 | 1p35.2–p35.1 | Histone deacetylase 1 | Mice | Proliferation | [79] |
| HDAC2 | 6q21 | Histone deacetylase 2 | Mice | Proliferation | [79] |
| IBA1 | 6p21.33 | Ionized calcium binding adaptor molecule 1 | Mice | Terminal differentiation | [104] |
| IL33 | 9p24.1 | Interleukin 33 | Mice | Proliferation | [95] |
| IL34 | 16q22.1 | Interleukin 34 | Mice; Zebrafish | Migration; Terminal differentiation | [47,63] |
| IL5 | 5q31.1 | Interleukin 5 | Rat | Proliferation | [98] |
| INOS | 19p13.11 | Inducible nitric oxide synthase | Mice | Proliferation | [105] |
| IRF8 | 16q24.1 | Interferon regulatory factor 8 | Mice | Early differentiation | [50] |
| ITGAV | 2q32.1 | Integrin subunit alpha V | Mice | Terminal differentiation | [78] |
| ITGB8 | 7p21.1 | Integrin subunit beta 8 | Mice | Terminal differentiation | [78] |
| MAFB | 20q12 | MAF bZIP transcription factor B | Mice | Terminal differentiation | [89] |
| MEIS3 | 19q13.32 | Meis homeobox 3 | Mice | Terminal differentiation | [66] |
| MIF | 22q11.23 | Macrophage migration inhibitory factor | Mice | Proliferation | [70] |
| MMP8 | 11q22.2 | Matrix metallopeptidase 8 | Mice | Migration | [50] |
| MMP9 | 20q13.12 | Matrix metallopeptidase 9 | Mice | Migration | [50] |
| NGF | 1p13.2 | Nerve growth factor | Mice | Proliferation | [101] |
| NOX2 | Xp21.1-p11.4 | NADPH oxidase 2 | Mice | Migration | [69] |
| P2RX7 | 12q24.31 | Purinergic receptor P2X 7 | Mice | Proliferation | [94] |
| PGRN | 17q21.31 | Progranulin | Zebrafish | Migration | [77] |
| RUNX1 | 21q22.12 | RUNX family transcription factor 1 | Mice | Proliferation; Early and terminal differentiation | [59,60] |
| SALL1 | 16q12.1 | Spalt like transcription factor 1 | Mice | Terminal differentiation | [66] |
| SALL3 | 18q23 | Spalt like transcription factor 3 | Mice | Terminal differentiation | [66] |
| SCF | 12q21.32 | Stem cell factor | Mice | Migration; Proliferation | [102] |
| SPI1 | 11p11.2 | Transcription factor PU.1 | Mice | Early differentiation | [50] |
| TGFB1 | 19q13.2 | Transforming growth factor beta 1 | Mice | Terminal differentiation | [65] |
| TREM2 | 6p21.1 | Triggering receptor expressed on myeloid cells 2 | Mice | Proliferation | [88] |
| VEGFR1 | 13q12.3 | Vascular endothelial growth factor receptor 1 | Mice | Migration | [69] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dermitzakis, I.; Manthou, M.E.; Meditskou, S.; Tremblay, M.-È.; Petratos, S.; Zoupi, L.; Boziki, M.; Kesidou, E.; Simeonidou, C.; Theotokis, P. Origin and Emergence of Microglia in the CNS—An Interesting (Hi)story of an Eccentric Cell. Curr. Issues Mol. Biol. 2023, 45, 2609-2628. https://doi.org/10.3390/cimb45030171
Dermitzakis I, Manthou ME, Meditskou S, Tremblay M-È, Petratos S, Zoupi L, Boziki M, Kesidou E, Simeonidou C, Theotokis P. Origin and Emergence of Microglia in the CNS—An Interesting (Hi)story of an Eccentric Cell. Current Issues in Molecular Biology. 2023; 45(3):2609-2628. https://doi.org/10.3390/cimb45030171
Chicago/Turabian StyleDermitzakis, Iasonas, Maria Eleni Manthou, Soultana Meditskou, Marie-Ève Tremblay, Steven Petratos, Lida Zoupi, Marina Boziki, Evangelia Kesidou, Constantina Simeonidou, and Paschalis Theotokis. 2023. "Origin and Emergence of Microglia in the CNS—An Interesting (Hi)story of an Eccentric Cell" Current Issues in Molecular Biology 45, no. 3: 2609-2628. https://doi.org/10.3390/cimb45030171