
Evaluation of Some Benzo[g]Quinazoline Derivatives as Antiviral Agents against Human Rotavirus Wa Strain: Biological Screening and Docking Study
 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
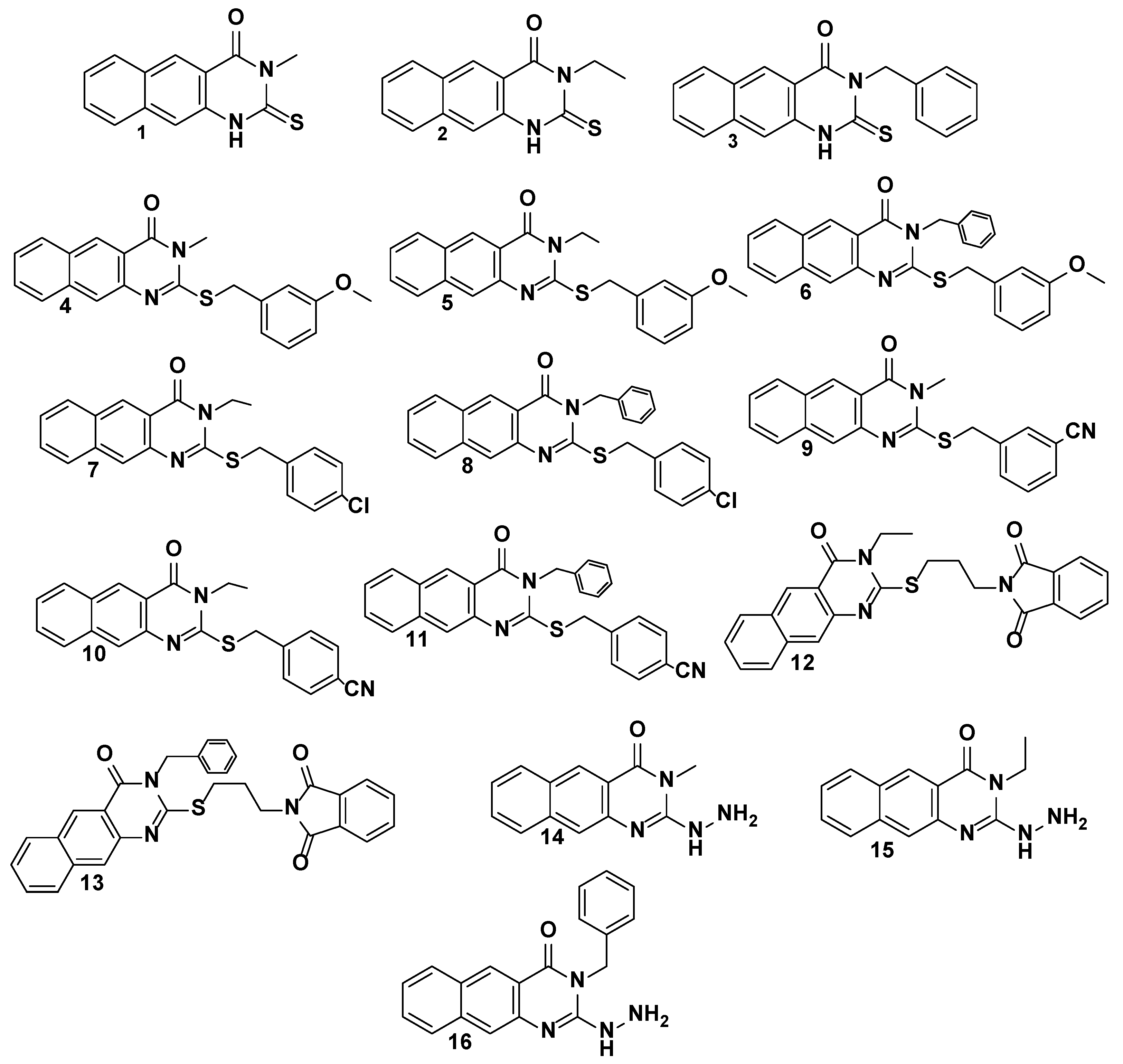
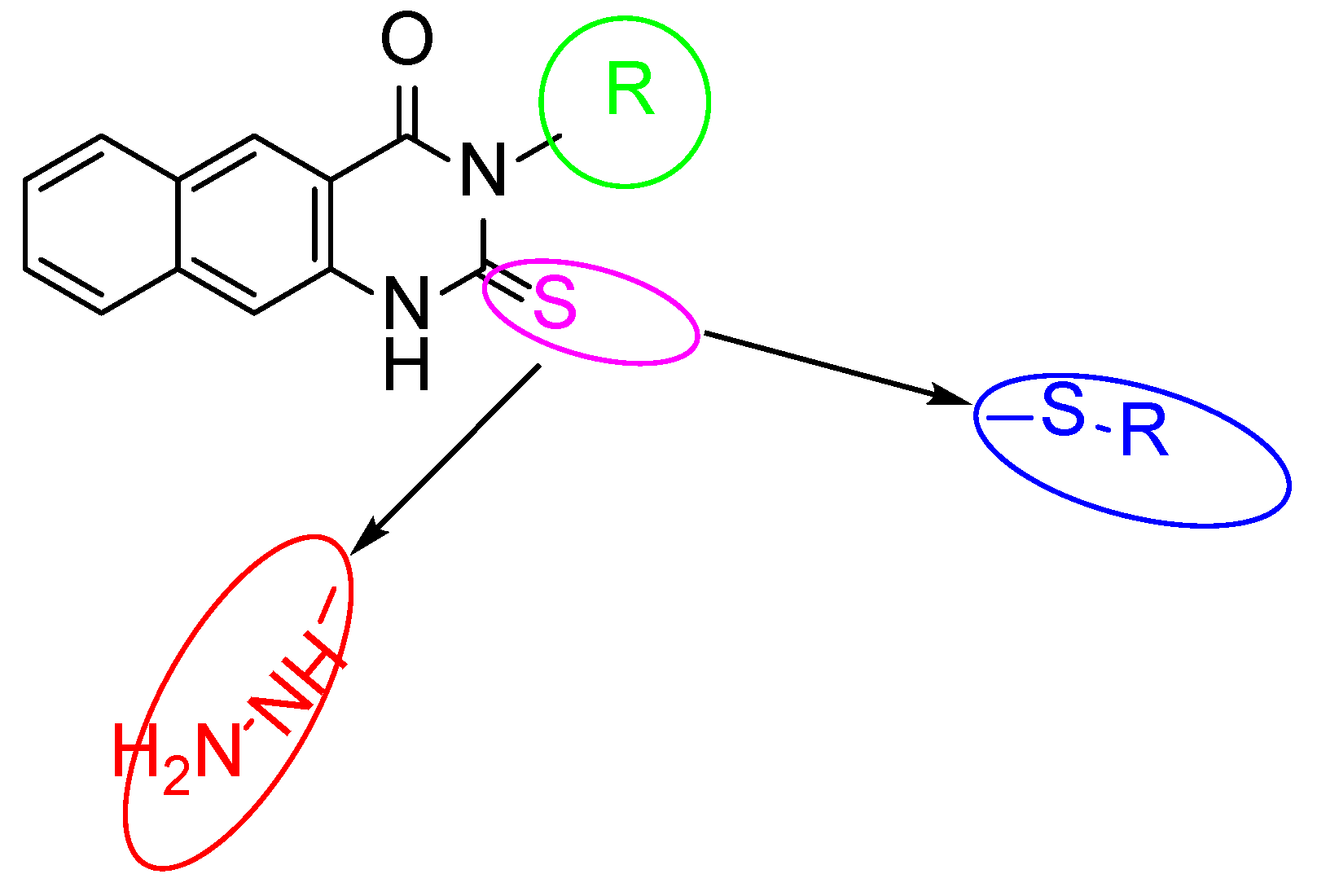
2.1. The Target 2-Thioxo-Benzo[g]quinazoline (1–16)
2.2. Cytotoxicity Test
2.3. Cell Morphology Evaluation by Inverted Light Microscopy
2.4. Cell Viability Assay
2.5. Determination of Rotavirus Titers Using Plaque Assay
2.6. Molecular Docking
2.7. Statistics
3. Results and Discussions
3.1. Biological Evaluation
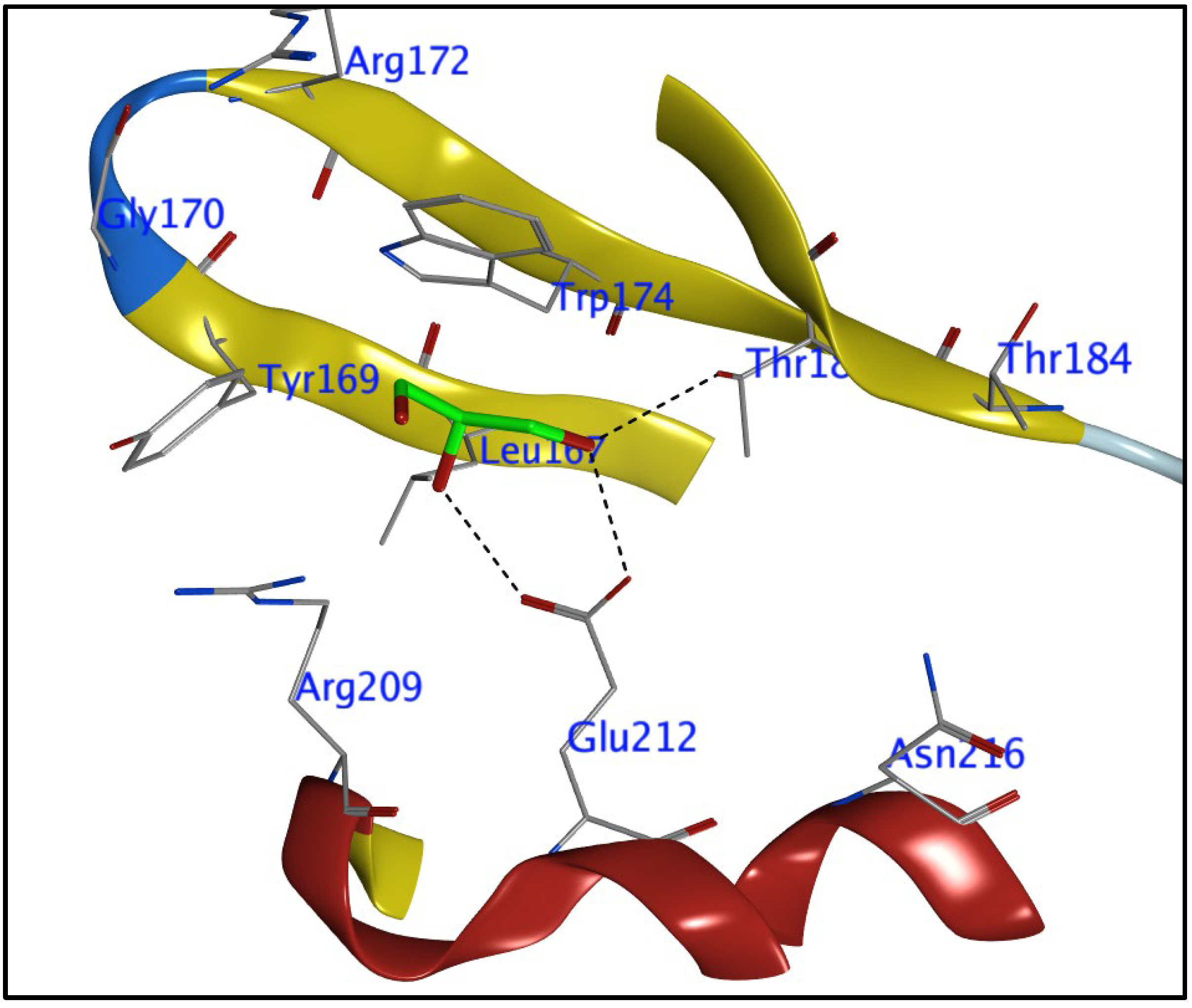
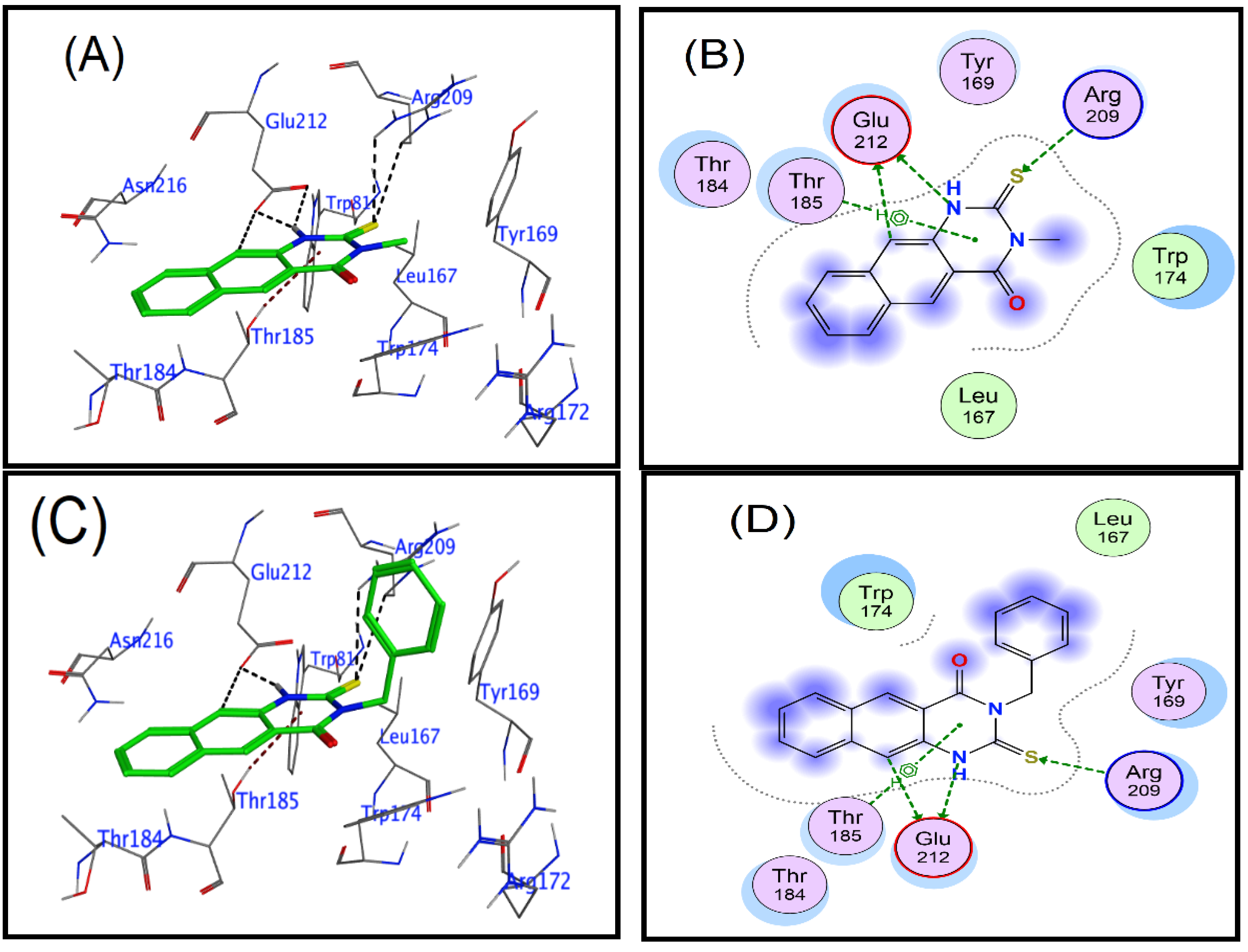
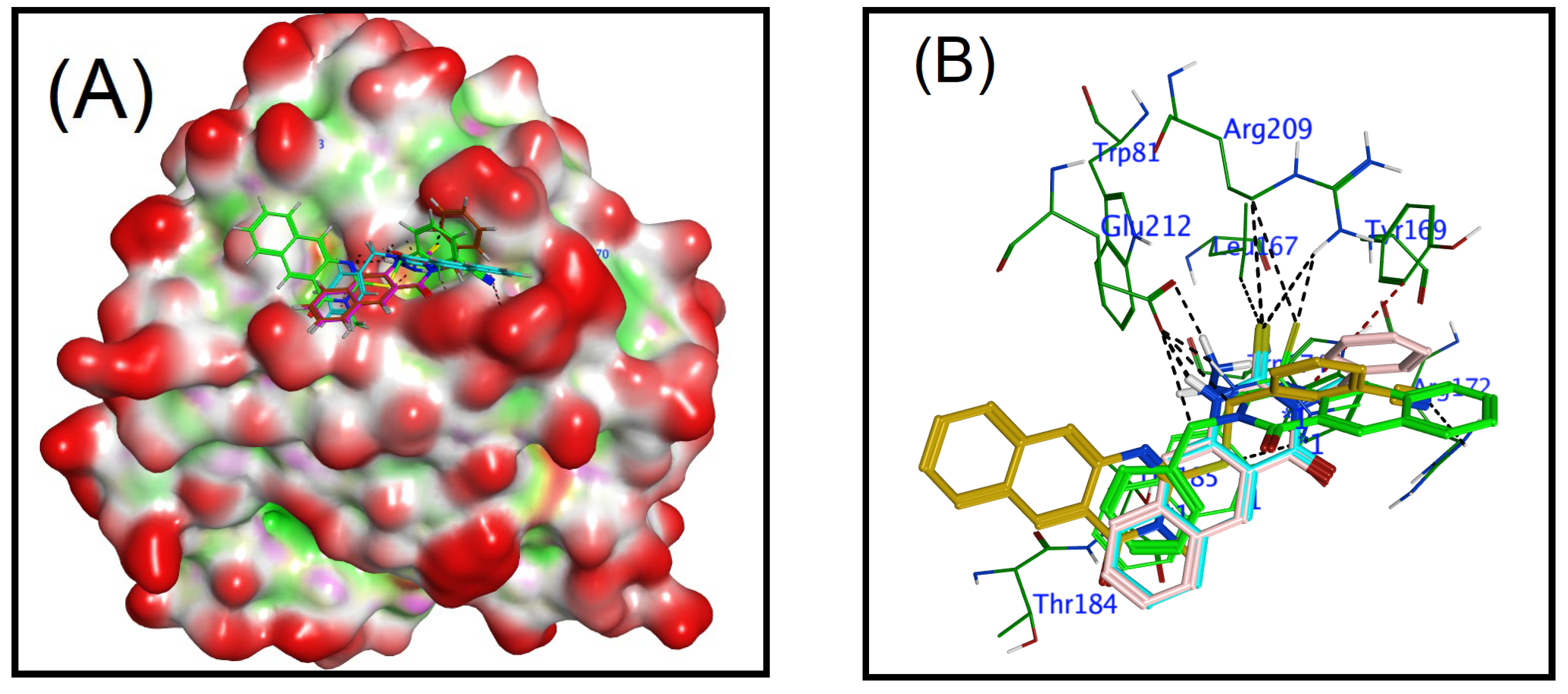
3.2. Docking Study
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yamashiro, R.; Sakudo, A.; Nagatsu, M. Efficient Recovery and Enrichment of Infectious Rotavirus Using Separation with Antibody-Integrated Graphite-Encapsulated Magnetic Nanobeads Produced by Argon/Ammonia Gas Plasma Technology. Int. J. Nanomed. 2019, 14, 1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atherly, D.E.; Lewis, K.D.C.; Tate, J.; Parashar, U.D.; Rheingans, R.D. Projected Health and Economic Impact of Rotavirus Vaccination in GAVI-Eligible Countries: 2011–2030. Vaccine 2012, 30, A7–A14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esona, M.D.; Gautam, R. Rotavirus. Clin. Lab. Med. 2015, 35, 363–391. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Otto, P.H.; Ciarlet, M.; Desselberger, U.; Van Ranst, M.; Johne, R. VP6-Sequence-Based Cutoff Values as a Criterion for Rotavirus Species Demarcation. Arch. Virol. 2012, 157, 1177–1182. [Google Scholar] [CrossRef]
- Chen, H.; Song, L.; Li, G.; Chen, W.; Zhao, S.; Zhou, R.; Shi, X.; Peng, Z.; Zhao, W. Human Rotavirus Strain Wa Downregulates NHE1 and NHE6 Expressions in Rotavirus-Infected Caco-2 Cells. Virus Genes 2017, 53, 367–376. [Google Scholar] [CrossRef]
- Kim, J.-H.; Kim, K.; Kim, W. Genipin Inhibits Rotavirus-Induced Diarrhea by Suppressing Viral Replication and Regulating Inflammatory Responses. Sci. Rep. 2020, 10, 15836. [Google Scholar] [CrossRef]
- La Frazia, S.; Ciucci, A.; Arnoldi, F.; Coira, M.; Gianferretti, P.; Angelini, M.; Belardo, G.; Burrone, O.R.; Rossignol, J.-F.; Santoro, M.G. Thiazolides, a New Class of Antiviral Agents Effective against Rotavirus Infection, Target Viral Morphogenesis, Inhibiting Viroplasm Formation. J. Virol. 2013, 87, 11096–11106. [Google Scholar] [CrossRef] [Green Version]
- Iosef, C.; Chang, K.; Azevedo, M.S.P.; Saif, L.J. Systemic and Intestinal Antibody Responses to NSP4 Enterotoxin of Wa Human Rotavirus in a Gnotobiotic Pig Model of Human Rotavirus Disease. J. Med. Virol. 2002, 68, 119–128. [Google Scholar] [CrossRef]
- Thiagarajah, J.R.; Verkman, A.S. CFTR Inhibitors for Treating Diarrheal Disease. Clin. Pharmacol. Ther. 2012, 92, 287–290. [Google Scholar] [CrossRef] [Green Version]
- Ko, E.-A.; Jin, B.-J.; Namkung, W.; Ma, T.; Thiagarajah, J.R.; Verkman, A.S. Chloride Channel Inhibition by a Red Wine Extract and a Synthetic Small Molecule Prevents Rotaviral Secretory Diarrhoea in Neonatal Mice. Gut 2014, 63, 1120–1129. [Google Scholar] [CrossRef] [Green Version]
- Kopic, S.; Geibel, J.P. Toxin Mediated Diarrhea in the 21st Century: The Pathophysiology of Intestinal Ion Transport in the Course of ETEC, V. Cholerae and Rotavirus Infection. Toxins 2010, 2, 2132–2157. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, M.C.; Cohen, J.; Michelangeli, F. Role of Ca2+ in the Replication and Pathogenesis of Rotavirus and Other Viral Infections. Cell Calcium 2000, 28, 137–149. [Google Scholar] [CrossRef]
- Bertrand, B.; Wakabayashi, S.; Ikeda, T.; Pouysségur, J.; Shigekawa, M. The Na+/H+ Exchanger Isoform 1 (NHE1) Is a Novel Member of the Calmodulin-Binding Proteins. Identification and Characterization of Calmodulin-Binding Sites. J. Biol. Chem. 1994, 269, 13703–13709. [Google Scholar] [CrossRef]
- Linhares, R.E.; Wigg, M.D.; Lagrota, M.H.; Nozawa, C.M. The in Vitro Antiviral Activity of Isoprinosine on Simian Rotavirus (SA-11). Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Medicas Biol. 1989, 22, 1095–1103. [Google Scholar]
- Smee, D.F.; Sidwell, R.W.; Clark, S.M.; Barnett, B.B.; Spendlove, R.S. Inhibition of Rotaviruses by Selected Antiviral Substances: Mechanisms of Viral Inhibition and in Vivo Activity. Antimicrob. Agents Chemother. 1982, 21, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Clark, K.J.; Grant, P.G.; Sarr, A.B.; Belakere, J.R.; Swaggerty, C.L.; Phillips, T.D.; Woode, G.N. An in Vitro Study of Theaflavins Extracted from Black Tea to Neutralize Bovine Rotavirus and Bovine Coronavirus Infections. Vet. Microbiol. 1998, 63, 147–157. [Google Scholar] [CrossRef]
- Takahashi, K.; Matsuda, M.; Ohashi, K.; Taniguchi, K.; Nakagomi, O.; Abe, Y.; Mori, S.; Sato, N.; Okutani, K.; Shigeta, S. Analysis of Anti-Rotavirus Activity of Extract from Stevia Rebaudiana. Antivir. Res. 2001, 49, 15–24. [Google Scholar] [CrossRef]
- Cecílio, A.B.; de Faria, D.B.; de Carvalho Oliveira, P.; Caldas, S.; de Oliveira, D.A.; Sobral, M.E.G.; Duarte, M.G.R.; de Souza Moreira, C.P.; Silva, C.G.; de Almeida, V.L. Screening of Brazilian Medicinal Plants for Antiviral Activity against Rotavirus. J. Ethnopharmacol. 2012, 141, 975–981. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, J.L.S.; Lopes, R.C.; Oliveira, D.B.; Costa, S.S.; Miranda, M.; Romanos, M.T.V.; Santos, N.S.O.; Wigg, M.D. In Vitro Anti-Rotavirus Activity of Some Medicinal Plants Used in Brazil against Diarrhea. J. Ethnopharmacol. 2005, 99, 403–407. [Google Scholar] [CrossRef]
- Al-Salahi, R.; Taie, H.A.A.; Bakheit, A.H.; Marzouk, M.; Almehizia, A.A.; Herqash, R.; Abuelizz, H.A. Antioxidant Activities and Molecular Docking of 2-Thioxobenzo[g]Quinazoline Derivatives. Pharmacol. Rep. 2019, 71, 695–700. [Google Scholar] [CrossRef]
- Abuelizz, H.A.; El-Dib, R.A.; Marzouk, M.; Al-Salahi, R. In Vitro Evaluation of New 2-Phenoxy-Benzo[g][1,2,4]Triazolo[1,5-a]Quinazoline Derivatives as Antimicrobial Agents. Microb. Pathog. 2018, 117, 60–67. [Google Scholar] [CrossRef]
- Al-Salahi, R.; Moustapha, M.E.; Abuelizz, H.A.; Alharthi, A.I.; Alburikan, K.A.; Ibrahim, I.T.; Marzouk, M.; Motaleb, M.A. Radioiodination and Biodistribution of Newly Synthesized 3-Benzyl-2-([3-Methoxybenzyl] Thio) Benzo [g] Quinazolin-4-(3H)-One in Tumor Bearing Mice. Saudi Pharm. J. 2018, 26, 1120–1126. [Google Scholar] [CrossRef] [PubMed]
- Almehizia, A.A.; Abuelizz, H.A.; Taie, H.A.A.; ElHassane, A.; Marzouk, M.; Al-Salahi, R. Investigation the Antioxidant Activity of Benzo[g]Triazoloquinazolines Correlated with a DFT Study. Saudi Pharm. J. 2019, 27, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Abuelizz, H.A.; Marzouk, M.; Bakheit, A.H.; Awad, H.M.; Soltan, M.M.; Naglah, A.M.; Al-Salahi, R. Antiproliferative and Antiangiogenic Properties of New VEGFR-2-Targeting 2-Thioxobenzo [g] Quinazoline Derivatives (In Vitro). Molecules 2020, 25, 5944. [Google Scholar] [CrossRef] [PubMed]
- Al-Salahi, R.; El Dib, R.A.; Marzouk, M. Synthesis and in Vitro Cytotoxicity Evaluation of New 2-Thioxo-Benzo[g]Quinazolin-4(3h)-One Derivatives. Heterocycles 2015, 91, 1735–1751. [Google Scholar] [CrossRef]
- Al-Salahi, R.; Abuelizz, H.A.; El Dib, R.; Marzouk, M.; Alshammari, M.B. Antimicrobial Activity of New 2-Thioxo-Benzo[g]Quinazolin-4(3H)-One Derivatives. Med. Chem. 2017, 13, 82–95. [Google Scholar] [CrossRef]
- Al-Salahi, R.; Ahmad, R.; Anouar, E.; Azman, N.I.I.N.; Marzouk, M.; Abuelizz, H.A. 3-Benzyl(Phenethyl)-2-Thioxobenzo[g]Quinazolines as a New Class of Potent α-Glucosidase Inhibitors: Synthesis and Molecular Docking Study. Future Med. Chem. 2018, 10, 1889–1905. [Google Scholar] [CrossRef]
- Abuelizz, H.A.; Marzouk, M.; Bakhiet, A.; Abdel-Aziz, M.M.; Ezzeldin, E.; Rashid, H.; Al-Salahi, R. In Silico Study and Biological Screening of Benzoquinazolines as Potential Antimicrobial Agents against Methicillin-Resistant Staphylococcus Aureus, Carbapenem-Resistant Klebsiella Pneumoniae, and Fluconazole-Resistant Candida Albicans. Microb. Pathog. 2021, 160, 105157. [Google Scholar] [CrossRef]
- Al-Salahi, R.; A Abuelizz, H.; Wadi, M.; A El Dib, R.; A Alotaibi, M.; Marzouk, M. Antimicrobial Activity of Synthesized 2-Methylthiobenzo [g][1, 2, 4]-Triazolo [1, 5-a] Quinazoline Derivatives. Med. Chem. 2016, 12, 760–766. [Google Scholar] [CrossRef]
- Al-Salahi, R.; Abuelizz, H.A.; Ghabbour, H.A.; El-Dib, R.; Marzouk, M. Molecular Docking Study and Antiviral Evaluation of 2-Thioxo-Benzo[g]Quinazolin-4(3H)-One Derivatives. Chem. Cent. J. 2016, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Abuelizz, H.A.; Marzouk, M.; Bakheit, A.H.; Al-Salahi, R. Investigation of Some Benzoquinazoline and Quinazoline Derivatives as Novel Inhibitors of HCV-NS3/4A Protease: Biological, Molecular Docking and QSAR Studies. RSC Adv. 2020, 10, 35820–35830. [Google Scholar] [CrossRef]
- Al-Salahi, R.; Anouar, E.H.; Marzouk, M.; Abuelizz, H.A. Anti-HAV Evaluation and Molecular Docking of Newly Synthesized 3-Benzyl(Phenethyl)Benzo[g]Quinazolines. Bioorganic Med. Chem. Lett. 2019, 29, 1614–1619. [Google Scholar] [CrossRef]
- Simões, C.M.O.; Amoros, M.; Girre, L. Mechanism of Antiviral Activity of Triterpenoid Saponins. Phyther. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 1999, 13, 323–328. [Google Scholar] [CrossRef]
- Walum, E.; Stenberg, K.; Jenssen, D. Understanding Cell Toxicology; E. Horwood: Cambridge, UK, 1990. [Google Scholar]
- Schmidtke, M.; Knorre, C.; Blei, L.; Stelzner, A.; Birch-Hirschfeld, E. Penetration and Antiviral Activity of Coxsackievirus B3 (CVB3)-Specific Phosphorothioate Oligodeoxynucleotides (PS-ODN). Nucleosides Nucleotides 1998, 17, 1557–1566. [Google Scholar] [CrossRef]
- Eichhorn, T.; Kolbe, F.; Mišić, S.; Dimić, D.; Morgan, I.; Saoud, M.; Milenković, D.; Marković, Z.; Rüffer, T.; Dimitrić Marković, J. Synthesis, Crystallographic Structure, Theoretical Analysis, Molecular Docking Studies, and Biological Activity Evaluation of Binuclear Ru (II)-1-Naphthylhydrazine Complex. Int. J. Mol. Sci. 2022, 24, 689. [Google Scholar] [CrossRef]
- Xu, S.; Liu, Y.; Tan, M.; Zhong, W.; Zhao, D.; Jiang, X.; Kennedy, M.A. Molecular Basis of P [6] and P [8] Major Human Rotavirus VP8* Domain Interactions with Histo-Blood Group Antigens. bioRxiv 2019, 512301. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comp. | Non-Toxic Doses (µg/mL) | ||
|---|---|---|---|
| BGM | MA104 | Hep-2 | |
| 1 | 60 | 65 | 65 |
| 2 | 60 | 60 | 60 |
| 3 | 60 | 75 | 70 |
| 4 | 50 | 55 | 55 |
| 5 | 55 | 55 | 60 |
| 6 | 65 | 65 | 65 |
| 7 | 60 | 60 | 60 |
| 8 | 50 | 50 | 50 |
| 9 | 65 | 75 | 70 |
| 10 | 50 | 55 | 50 |
| 11 | 65 | 65 | 65 |
| 12 | 25 | 30 | 30 |
| 13 | 40 | 50 | 45 |
| 14 | 55 | 60 | 60 |
| 15 | 55 | 55 | 55 |
| 16 | 60 | 60 | 65 |
| Compound | % Reduction | Compound | % Reduction |
|---|---|---|---|
| 1 | 60% ± 3% | 2 | 50% ± 3% |
| 3 | 53.3% ± 3% | 4 | 10% ± 3% |
| 5 | 10% ± 4% | 6 | 20% ± 4% |
| 7 | 16.7% ± 3% | 8 | 30% ± 3% |
| 9 | 66.7% ± 1% | 10 | 20% ± 4% |
| 11 | 30% ± 2% | 12 | 16.7% ± 2% |
| 13 | 20% ± 3% | 14 | 30% ± 3% |
| 15 | 13.3% ± 3% | 16 | 50% ± 3% |
| Cp | Ligand | Receptor | Interaction | Distance | E (kcal/mol) | Score (kcal/mol) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | N | 9 | OE1 | GLU | 212 | (A) | H-donor | 3.61 | −0.3 | −3.953 |
| N | 9 | OE2 | GLU | 212 | (A) | H-donor | 3.11 | −5.2 | ||
| C | 25 | OE2 | GLU | 212 | (A) | H-donor | 3.38 | −0.3 | ||
| S | 27 | CD | ARG | 209 | (A) | H-acceptor | 3.71 | −0.8 | ||
| S | 27 | NH1 | ARG | 209 | (A) | H-acceptor | 3.57 | −0.8 | ||
| 6-ring | OG1 | THR | 185 | (A) | pi-H | 4.2 | −1.3 | |||
| 3 | N | 19 | OE2 | GLU | 212 | (A) | H-donor | 3.17 | −4.3 | −4.209 |
| C | 35 | OE2 | GLU | 212 | (A) | H-donor | 3.35 | −0.3 | ||
| S | 37 | CD | ARG | 209 | (A) | H-acceptor | 3.86 | −0.7 | ||
| S | 37 | NH1 | ARG | 209 | (A) | H-acceptor | 3.37 | −0.6 | ||
| 6-ring | OG1 | THR | 185 | (A) | pi-H | 4.1 | −1.3 | |||
| 9 | C | 10 | OE2 | GLU | 212 | (A) | H-donor | 2.96 | −0.7 | −4.826 |
| S | 9 | CZ3 | TRP | 174 | (A) | H-acceptor | 4.4 | −0.2 | ||
| N | 22 | NH1 | ARG | 172 | (A) | H-acceptor | 3.84 | −0.2 | ||
| C | 23 | 6-ring TRP | 174 | (A) | H-pi | 4.53 | −0.2 | |||
| 6-ring | CE2 | TYR | 169 | (A) | pi-H | 4.56 | −0.2 | |||
| 16 | C | 4 | OE2 | GLU | 212 | (A) | H-donor | 3.44 | −0.4 | −4.287 |
| N | 19 | OE2 | GLU | 212 | (A) | H-donor | 2.91 | −5 | ||
| N | 21 | OE1 | GLU | 212 | (A) | H-donor | 3.33 | −0.7 | ||
| 6-ring | CE2 | TYR | 169 | (A) | pi-H | 4.85 | −0.2 | |||
| 6-ring | NH2 | ARG | 172 | (A) | pi-cation | 4.4 | −0.9 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abuelizz, H.A.; Bakheit, A.H.; Marzouk, M.; El-Senousy, W.M.; Abdellatif, M.M.; Mostafa, G.A.E.; Al-Salahi, R. Evaluation of Some Benzo[g]Quinazoline Derivatives as Antiviral Agents against Human Rotavirus Wa Strain: Biological Screening and Docking Study. Curr. Issues Mol. Biol. 2023, 45, 2409-2421. https://doi.org/10.3390/cimb45030156
Abuelizz HA, Bakheit AH, Marzouk M, El-Senousy WM, Abdellatif MM, Mostafa GAE, Al-Salahi R. Evaluation of Some Benzo[g]Quinazoline Derivatives as Antiviral Agents against Human Rotavirus Wa Strain: Biological Screening and Docking Study. Current Issues in Molecular Biology. 2023; 45(3):2409-2421. https://doi.org/10.3390/cimb45030156
Chicago/Turabian StyleAbuelizz, Hatem A., Ahmed H. Bakheit, Mohamed Marzouk, Waled M. El-Senousy, Mohamed M. Abdellatif, Gamal A. E. Mostafa, and Rashad Al-Salahi. 2023. "Evaluation of Some Benzo[g]Quinazoline Derivatives as Antiviral Agents against Human Rotavirus Wa Strain: Biological Screening and Docking Study" Current Issues in Molecular Biology 45, no. 3: 2409-2421. https://doi.org/10.3390/cimb45030156