
Effect of Atorvastatin on Angiogenesis-Related Genes VEGF-A, HGF and IGF-1 and the Modulation of PI3K/AKT/mTOR Transcripts in Bone-Marrow-Derived Mesenchymal Stem Cells
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Morphology, Viability and Characterization of BM-MSCs Expanded In Vitro
2.3. Immunofluorescence Staining
2.4. Gene Expression Determination
2.5. Statistical Analysis
3. Results
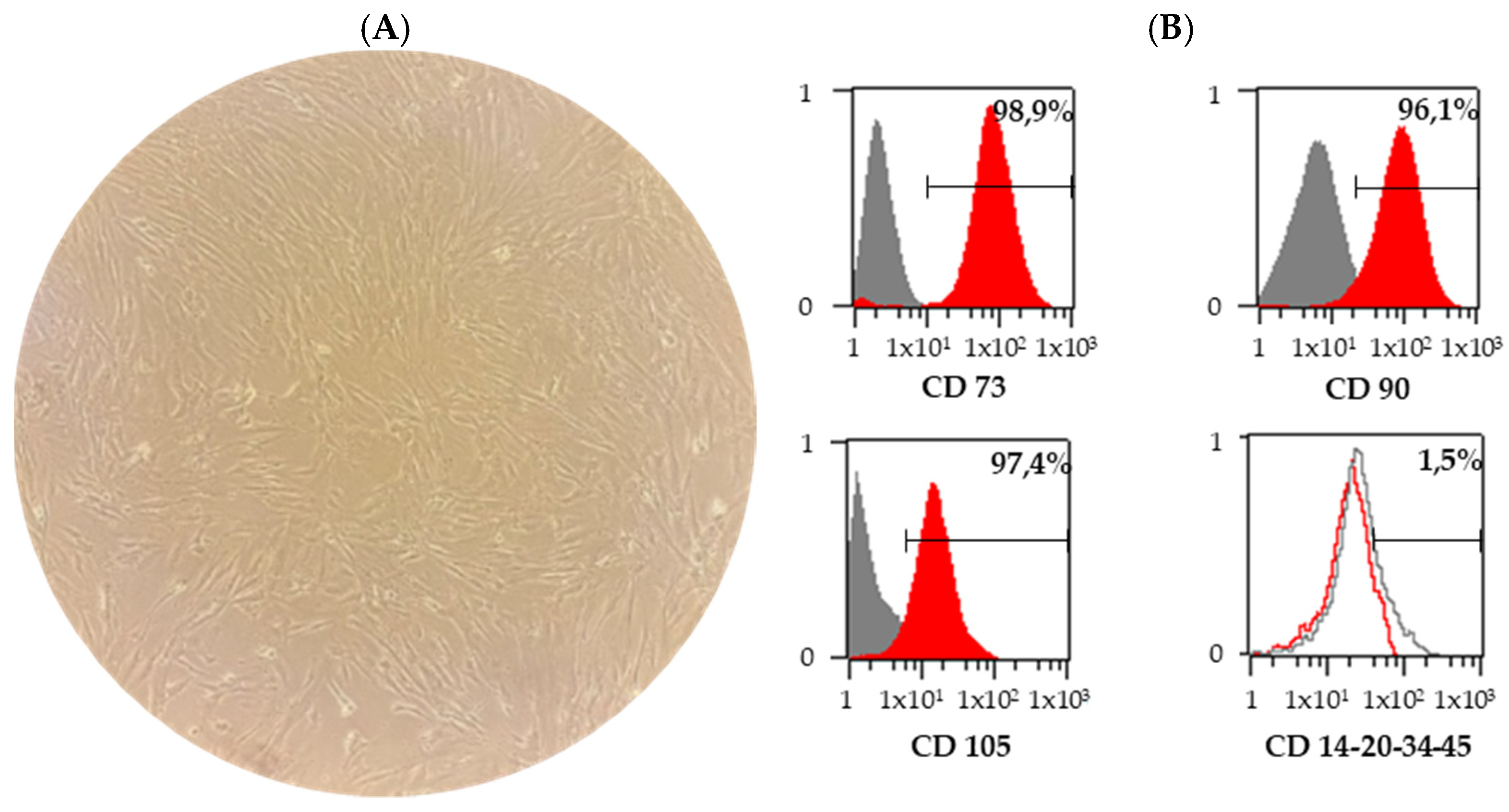
3.1. Characterization of BM-MSCs
3.2. Effect of Atorvastatin Pre-Treatment on Characteristics and Viability of BM-MSCs
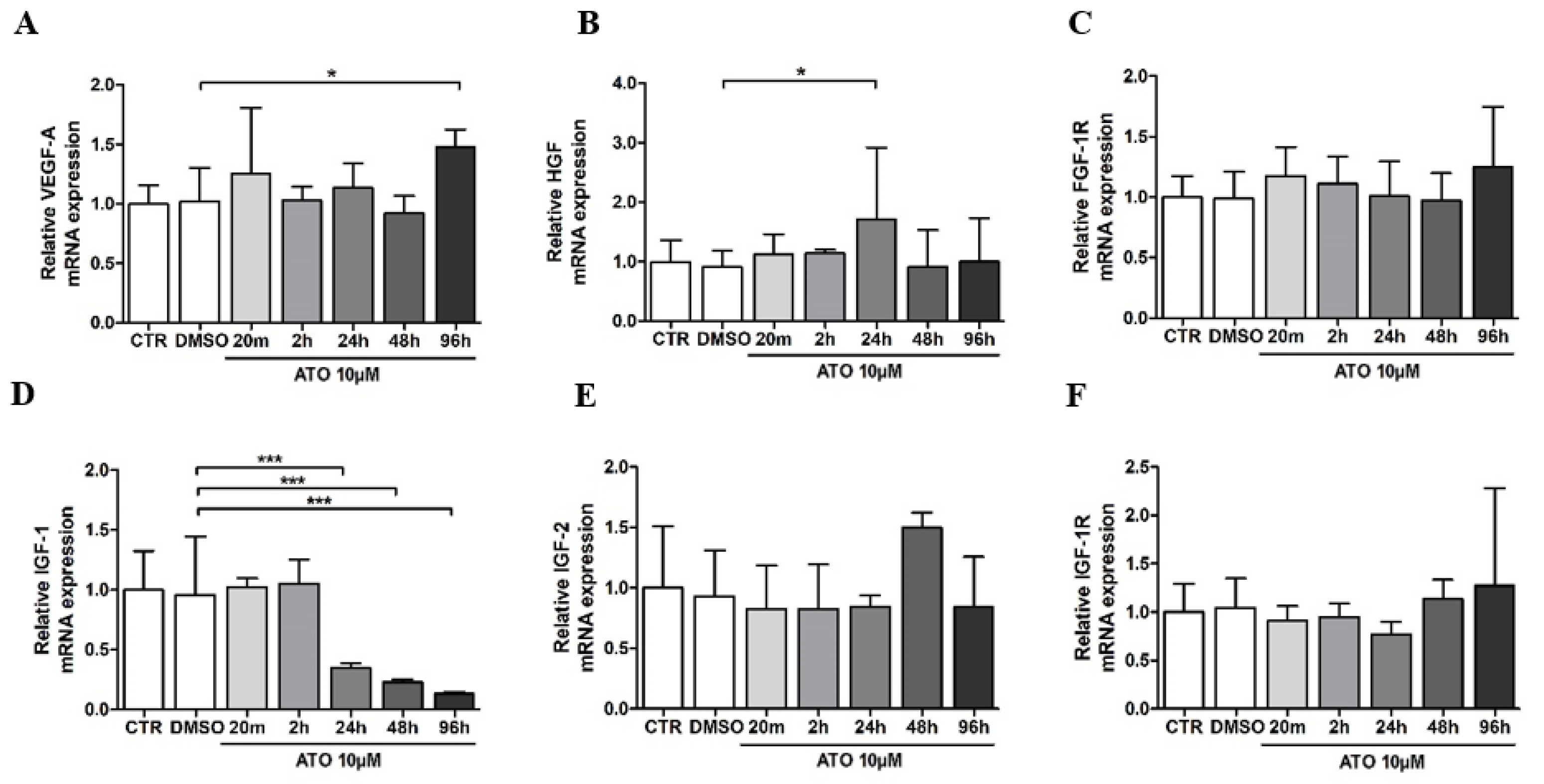
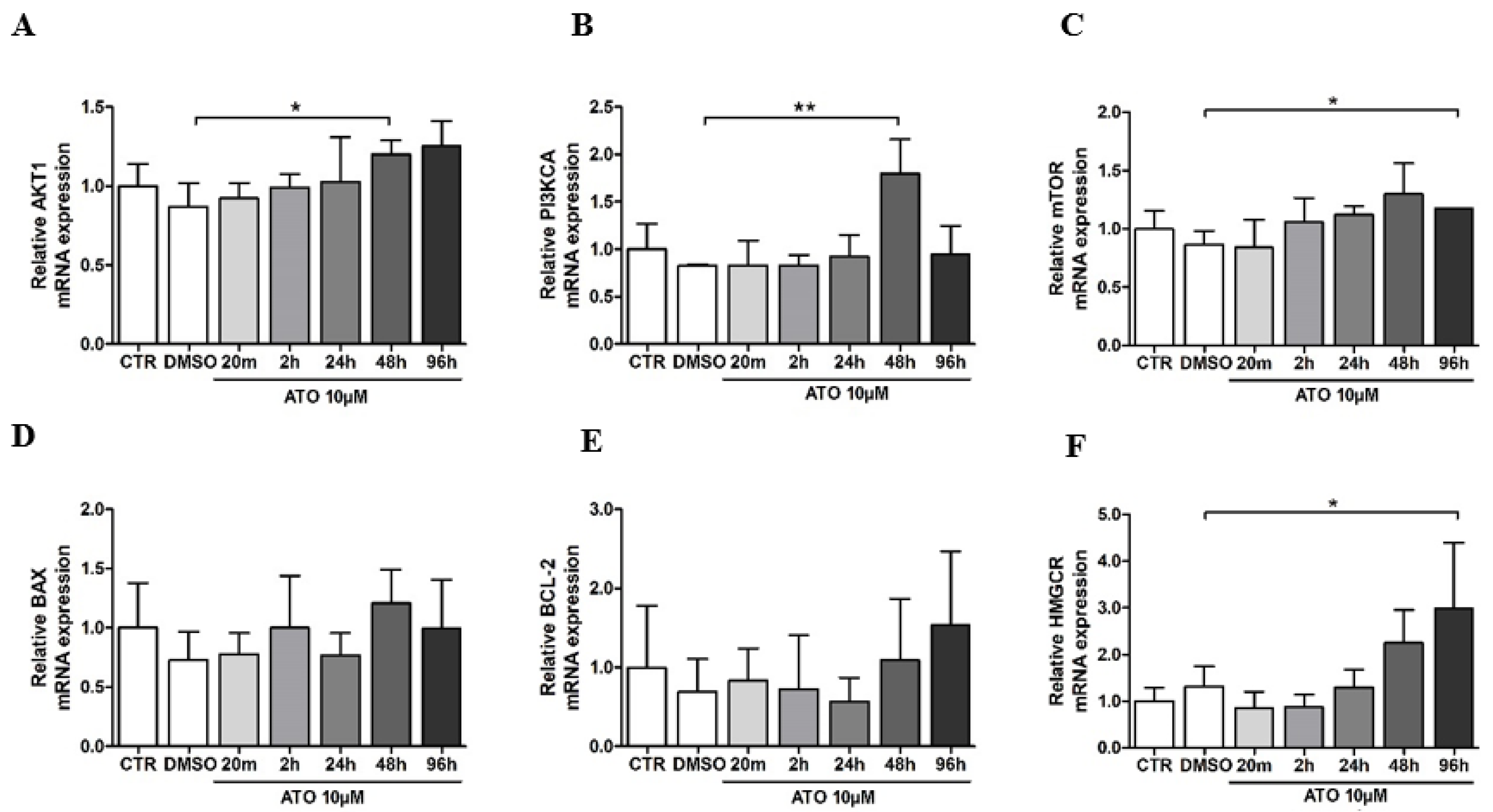
3.3. Atorvastatin’s Effect on Gene Expression in BM-MSCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wood, W.G.; Igbavboa, U.; Muller, W.E.; Eckert, G.P. Statins, Bcl-2, and Apoptosis: Cell Death or Cell Protection? Mol. Neurobiol. 2013, 48, 308–314. [Google Scholar] [CrossRef] [Green Version]
- Marcus, M.E.; Manne-Goehler, J.; Theilmann, M.; Farzadfar, F.; Moghaddam, S.S.; Keykhaei, M.; Hajebi, A.; Tschida, S.; Lemp, J.M.; Aryal, K.K.; et al. Use of Statins for the Prevention of Cardiovascular Disease in 41 Low-Income and Middle-Income Countries: A Cross-Sectional Study of Nationally Representative, Individual-Level Data. Lancet Glob. Health 2022, 10, e369–e379. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.; Deplazes, E.; Cranfield, C.G.; Garcia, A. The Role of Structure and Biophysical Properties in the Pleiotropic Effects of Statins. Int. J. Mol. Sci. 2020, 21, 8745. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, M.; Amiri, S.; Pecic, S.; Machaj, F.; Rosik, J.; Łos, M.J.; Alizadeh, J.; Mahdian, R.; da Silva Rosa, S.C.; Schaafsma, D.; et al. Pleiotropic Effects of Statins: A Focus on Cancer. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2020, 1866, 165968. [Google Scholar] [CrossRef]
- Cai, J.; Yu, X.; Zhang, B.; Zhang, H.; Fang, Y.; Liu, S.; Liu, T.; Ding, X. Atorvastatin Improves Survival of Implanted Stem Cells in a Rat Model of Renal Ischemia-Reperfusion Injury. Am. J. Nephrol. 2014, 39, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Chruściel, P.; Sahebkar, A.; Rembek-Wieliczko, M.; Serban, M.-C.; Ursoniu, S.; Mikhailidis, D.P.; Jones, S.R.; Mosteoru, S.; Blaha, M.J.; Martin, S.S.; et al. Impact of Statin Therapy on Plasma Adiponectin Concentrations: A Systematic Review and Meta-Analysis of 43 Randomized Controlled Trial Arms. Atherosclerosis 2016, 253, 194–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Yao, L.; Yang, J.; Wang, Z.; Du, G. PI3K/Akt and HIF-1 Signaling Pathway in Hypoxia-ischemia (Review). Mol. Med. Rep. 2018, 18, 3547–3554. [Google Scholar] [CrossRef] [Green Version]
- Gelosa, P.; Cimino, M.; Pignieri, A.; Tremoli, E.; Guerrini, U.; Sironi, L. The Role of HMG-CoA Reductase Inhibition in Endothelial Dysfunction and Inflammation. Vasc. Health Risk Manag. 2007, 3, 567–577. [Google Scholar]
- Xie, Y.; Shi, X.; Sheng, K.; Han, G.; Li, W.; Zhao, Q.; Jiang, B.; Feng, J.; Li, J.; Gu, Y. PI3K/Akt Signaling Transduction Pathway, Erythropoiesis and Glycolysis in Hypoxia (Review). Mol. Med. Rep. 2018, 19, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Adamickova, A.; Gazova, A.; Adamicka, M.; Chomanicova, N.; Valaskova, S.; Cervenak, Z.; Salingova, B.; Kyselovic, J. Molecular Basis of the Effect of Atorvastatin Pre-Treatment on Stem Cell Therapy in Chronic Ischemic Diseases—Critical Limb Ischemia. Physiol. Res. 2021, 70, S527–S533. [Google Scholar] [CrossRef]
- Dai, G.; Xu, Q.; Luo, R.; Gao, J.; Chen, H.; Deng, Y.; Li, Y.; Wang, Y.; Yuan, W.; Wu, X. Atorvastatin Treatment Improves Effects of Implanted Mesenchymal Stem Cells: Meta-Analysis of Animal Models with Acute Myocardial Infarction. BMC Cardiovasc. Disord. 2015, 15, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorabi, A.M.; Kiaie, N.; Pirro, M.; Bianconi, V.; Jamialahmadi, T.; Sahebkar, A. Effects of Statins on the Biological Features of Mesenchymal Stem Cells and Therapeutic Implications. Heart Fail. Rev. 2020, 26, 1259–1272. [Google Scholar] [CrossRef]
- Niu, J.; Ding, G.; Zhang, L. Effects of Simvastatin on the Osteogenic Differentiation and Immunomodulation of Bone Marrow Mesenchymal Stem Cells. Mol. Med. Rep. 2015, 12, 8237–8240. [Google Scholar] [CrossRef] [Green Version]
- Adams, S.P.; Tsang, M.; Wright, J.M. Atorvastatin for Lowering Lipids. Cochrane Database Syst. Rev. 2015, 2015, CD008226. [Google Scholar] [CrossRef] [PubMed]
- Klepanec, A.; Mistrik, M.; Altaner, C.; Valachovicova, M.; Olejarova, I.; Slysko, R.; Balazs, T.; Urlandova, T.; Hladikova, D.; Liska, B.; et al. No Difference in Intra-Arterial and Intramuscular Delivery of Autologous Bone Marrow Cells in Patients with Advanced Critical Limb Ischemia. Cell Transplant. 2012, 21, 1909–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björkhem-Bergman, L.; Lindh, J.D.; Bergman, P. What Is a Relevant Statin Concentration in Cell Experiments Claiming Pleiotropic Effects? Br. J. Clin. Pharmacol. 2011, 72, 164–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssef, S.; Radosevich, J.L.; Hur, E.M.; Bravo, M.; Mitchell, D.J.; Sobel, R.A.; Steinman, L.; Zamvil, S.S. The HMG-CoA Reductase Inhibitor, Atorvastatin, Promotes a Th2 Bias and Reverses Paralysis in Central Nervous System Autoimmune Disease. Nature 2002, 420, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [Green Version]
- Adamo, A.; Brandi, J.; Caligola, S.; Delfino, P.; Bazzoni, R.; Carusone, R.; Cecconi, D.; Giugno, R.; Manfredi, M.; Robotti, E.; et al. Extracellular Vesicles Mediate Mesenchymal Stromal Cell-Dependent Regulation of B Cell PI3K-AKT Signaling Pathway and Actin Cytoskeleton. Front. Immunol. 2019, 10, 446. [Google Scholar] [CrossRef]
- Wu, R.; Hu, X.; Wang, J. Concise Review: Optimized Strategies for Stem Cell-Based Therapy in Myocardial Repair: Clinical Translatability and Potential Limitation: Optimization of Stem Cells and Microenvironments. Stem Cells 2018, 36, 482–500. [Google Scholar] [CrossRef] [Green Version]
- Bybee, K.A.; Lee, J.H.; O’Keefe, J.H. Cumulative Clinical Trial Data on Atorvastatin for Reducing Cardiovascular Events: The Clinical Impact of Atorvastatin. Curr. Med. Res. Opin. 2008, 24, 1217–1229. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, H.S.; Goa, K.L. Atorvastatin: An Updated Review of Its Pharmacological Properties and Use in Dyslipidaemia. Drugs 2001, 61, 1835–1881. [Google Scholar] [CrossRef]
- Qu, Z.; Xu, H.; Tian, Y.; Jiang, X. Atorvastatin Improves Microenvironment to Enhance the Beneficial Effects of BMSCs Therapy in a Rabbit Model of Acute Myocardial Infarction. Cell. Physiol. Biochem. 2013, 32, 380–389. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, H.; Yang, Y.-J.; Dong, Q.-T.; Wang, T.-J.; Qian, H.-Y.; Li, N.; Wang, X.-M.; Jin, C. Atorvastatin Treatment Improves the Effects of Mesenchymal Stem Cell Transplantation on Acute Myocardial Infarction: The Role of the RhoA/ROCK/ERK Pathway. Int. J. Cardiol. 2014, 176, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, S.; Cui, M.; Gao, X.; Sun, D.; Qin, X.; Narsinh, K.; Li, C.; Jia, H.; Li, C.; et al. Rosuvastatin Enhances the Therapeutic Efficacy of Adipose-Derived Mesenchymal Stem Cells for Myocardial Infarction via PI3K/Akt and MEK/ERK Pathways. Basic Res. Cardiol. 2013, 108, 333. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Müller, A.L.; Ngo, M.A.; Sran, K.; Bellan, D.; Arora, R.C.; Kirshenbaum, L.A.; Freed, D.H. Statins Impair Survival of Primary Human Mesenchymal Progenitor Cells via Mevalonate Depletion, NF-ΚB Signaling, and Bnip3. J. Cardiovasc. Transl. Res. 2015, 8, 96–105. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Brasile, L.; Henry, N.; Orlando, G.; Stubenitsky, B. Potentiating Renal Regeneration Using Mesenchymal Stem Cells. Transplantation 2019, 103, 307–313. [Google Scholar] [CrossRef]
- Hodgkinson, C.P.; Bareja, A.; Gomez, J.A.; Dzau, V.J. Emerging Concepts in Paracrine Mechanisms in Regenerative Cardiovascular Medicine and Biology. Circ. Res. 2016, 118, 95–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Praveen Kumar, L.; Sangeetha, K.; Ranjita, M.; Vijayalakshmi, S.; Rajagopal, K.; Rama Shanker, V. The Mesenchymal Stem Cell Secretome: A New Paradigm towards Cell-Free Therapeutic Mode in Regenerative Medicine. Cytokine Growth Factor Rev. 2019, 46, 1–9. [Google Scholar] [CrossRef]
- Miranda, J.P.; Camões, S.P.; Gaspar, M.M.; Rodrigues, J.S.; Carvalheiro, M.; Bárcia, R.N.; Cruz, P.; Cruz, H.; Simões, S.; Santos, J.M. The Secretome Derived From 3D-Cultured Umbilical Cord Tissue MSCs Counteracts Manifestations Typifying Rheumatoid Arthritis. Front. Immunol. 2019, 10, 18. [Google Scholar] [CrossRef]
- Sun, H.; Pratt, R.E.; Hodgkinson, C.P.; Dzau, V.J. Sequential Paracrine Mechanisms Are Necessary for the Therapeutic Benefits of Stem Cell Therapy. Am. J. Physiol.-Cell Physiol. 2020, 319, C1141–C1150. [Google Scholar] [CrossRef]
- Cantoni, S.; Cavallini, C.; Bianchi, F.; Bonavita, F.; Vaccari, V.; Olivi, E.; Frascari, I.; Tassinari, R.; Valente, S.; Lionetti, V.; et al. Rosuvastatin Elicits KDR-Dependent Vasculogenic Response of Human Placental Stem Cells through PI3K/AKT Pathway. Pharmacol. Res. 2012, 65, 275–284. [Google Scholar] [CrossRef]
- Melincovici, C.S.; Bo, A.B.; Mihu, C.; Istrate, M.; Moldovan, I.-M.; Roman, A.L.; Mihu, C.M. Vascular Endothelial Growth Factor (VEGF)—Key Factor in Normal and Pathological Angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Johnson, K.E.; Wilgus, T.A. Vascular Endothelial Growth Factor and Angiogenesis in the Regulation of Cutaneous Wound Repair. Adv. Wound Care 2014, 3, 647–661. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Feng, X.; Wang, J.; Chen, T.; Bi, S.; Zhang, S. Simvastatin Regulates Endogenous Stem Cells to Reconstruct the Degenerative Intervertebral Disc. Chin. J. Tissue Eng. Res. 2017, 21, 809–814. [Google Scholar] [CrossRef]
- Qi, Y.; Zhao, T.; Yan, W.; Xu, K.; Shi, Z.; Wang, J. Mesenchymal Stem Cell Sheet Transplantation Combined with Locally Released Simvastatin Enhances Bone Formation in a Rat Tibia Osteotomy Model. Cytotherapy 2013, 15, 44–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, F.-Y.; Chen, B.-Y.; Chen, L.-L.; Zhang, F.-L.; Luo, Y.-K.; Jun, F. Improvement of the Survival and Therapeutic Effects of Implanted Mesenchymal Stem Cells in a Rat Model of Coronary Microembolization by Rosuvastatin Treatment. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2368–2381. [Google Scholar] [PubMed]
- Kawashiri, M.-A.; Nakanishi, C.; Tsubokawa, T.; Shimojima, M.; Yoshida, S.; Yoshimuta, T.; Konno, T.; Yamagishi, M.; Hayashi, K. Impact of Enhanced Production of Endogenous Heme Oxygenase-1 by Pitavastatin on Survival and Functional Activities of Bone Marrow-Derived Mesenchymal Stem Cells. J. Cardiovasc. Pharmacol. 2015, 65, 601–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mieno, S.; Ramlawi, B.; Boodhwani, M.; Clements, R.T.; Minamimura, K.; Maki, T.; Xu, S.-H.; Bianchi, C.; Li, J.; Sellke, F.W. Role of Stromal-Derived Factor-1alpha in the Induction of Circulating CD34+CXCR4+ Progenitor Cells after Cardiac Surgery. Circulation 2006, 114, 186–192. [Google Scholar] [CrossRef] [Green Version]
- Longobardi, L.; O’Rear, L.; Aakula, S.; Johnstone, B.; Shimer, K.; Chytil, A.; Horton, W.A.; Moses, H.L.; Spagnoli, A. Effect of IGF-I in the Chondrogenesis of Bone Marrow Mesenchymal Stem Cells in the Presence or Absence of TGF-β Signaling. J. Bone Miner. Res. 2005, 21, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.-F.; Jeng, L.-B.; Shyu, W.-C. Role of Insulin-like Growth Factor 1 Receptor Signaling in Stem Cell Stemness and Therapeutic Efficacy. Cell Transplant. 2018, 27, 1313–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Santis, M.C.; Gulluni, F.; Campa, C.C.; Martini, M.; Hirsch, E. Targeting PI3K Signaling in Cancer: Challenges and Advances. Biochim. Biophys. Acta (BBA)—Rev. Cancer 2019, 1871, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Luo, J.; Guo, J.; Yao, X.; Jing, X.; Guo, F. The PI3K/AKT/MTOR Signaling Pathway in Osteoarthritis: A Narrative Review. Osteoarthr. Cartil. 2020, 28, 400–409. [Google Scholar] [CrossRef]
- Guo, H.; Cui, H.; Peng, X.; Fang, J.; Zuo, Z.; Deng, J.; Wang, X.; Wu, B.; Chen, K.; Deng, J. Modulation of the PI3K/Akt Pathway and Bcl-2 Family Proteins Involved in Chicken’s Tubular Apoptosis Induced by Nickel Chloride (NiCl2). Int. J. Mol. Sci. 2015, 16, 22989–23011. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.-J.; Qian, H.-Y.; Huang, J.; Li, J.-J.; Gao, R.-L.; Dou, K.-F.; Yang, G.-S.; Willerson, J.T.; Geng, Y.-J. Combined Therapy With Simvastatin and Bone Marrow-Derived Mesenchymal Stem Cells Increases Benefits in Infarcted Swine Hearts. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 2076–2082. [Google Scholar] [CrossRef] [Green Version]
- Laffont, B.; Rayner, K.J. MicroRNAs in the Pathobiology and Therapy of Atherosclerosis. Can. J. Cardiol. 2017, 33, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Hitachi, K.; Tsuchida, K. Role of MicroRNAs in Skeletal Muscle Hypertrophy. Front. Physiol. 2014, 4, 408. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Guo, X.-Y.; Zhou, J.; Yan, X.-L.; Yu, F.-F. Atorvastatin Pretreatment Ameliorates Mesenchymal Stem Cell Migration through MiR-146a/CXCR4 Signaling. Tissue Eng. Regen. Med. 2021, 18, 863–873. [Google Scholar] [CrossRef]
- Bian, S.; Zhang, L.; Duan, L.; Wang, X.; Min, Y.; Yu, H. Extracellular Vesicles Derived from Human Bone Marrow Mesenchymal Stem Cells Promote Angiogenesis in a Rat Myocardial Infarction Model. J. Mol. Med. 2014, 92, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Wang, L.; Li, Q.; Tian, X.; Xu, J.; Xu, J.; Xiong, Y.; Chen, G.; Qian, H.; Jin, C.; et al. Atorvastatin Enhances the Therapeutic Efficacy of Mesenchymal Stem Cells-Derived Exosomes in Acute Myocardial Infarction via up-Regulating Long Non-Coding RNA H19. Cardiovasc. Res. 2020, 116, 353–367. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense: 5′ | Anti-Sense: 5′ | |
|---|---|---|---|
| VEGF-A | GGTCCCAGGCTGCACCCATG | ATTGCAGCAGCCCCCGCATC | |
| HGF | CTTCCATTCACTTGCAAGGCT | TGTTCCCTTGTAGCTGCGTC | |
| IGF-1 | TGGATGCTCTTCAGTTCGTG | ATCCACGATGCCTGTCTGA | |
| B2M | TCCGTGGCCTTAGCTGTGCT | TCCATTCTCTGCTGGATGACGTGAG | |
| GAPDH | TCCTGTTCGACAGTCAGCCGC | CATGGTGTCTGAGCGATGTGGC | |
| RPL13A | CTTTTCCAAGCGGCTGCCGAAGA | GGCCTCGACCATCAAGCACCAG | |
| Gene | Primer ID | Gene | Primer ID |
| FGF-1R | Hs00241111 | BAX | Hs00180269 |
| IGF-2 | Hs01005963 | BCL-2 | Hs00608023 |
| IGF-1R | Hs00609566 | HMGCR | Hs00168352 |
| PI3KCA | Hs00907957 | B2M | Hs99999907 |
| AKT1 | Hs00178289 | GAPDH | Hs99999905 |
| mTOR | Hs00234508 | RPL13A | Hs01578912 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adamičková, A.; Chomaničová, N.; Gažová, A.; Maďarič, J.; Červenák, Z.; Valášková, S.; Adamička, M.; Kyselovic, J. Effect of Atorvastatin on Angiogenesis-Related Genes VEGF-A, HGF and IGF-1 and the Modulation of PI3K/AKT/mTOR Transcripts in Bone-Marrow-Derived Mesenchymal Stem Cells. Curr. Issues Mol. Biol. 2023, 45, 2326-2337. https://doi.org/10.3390/cimb45030150
Adamičková A, Chomaničová N, Gažová A, Maďarič J, Červenák Z, Valášková S, Adamička M, Kyselovic J. Effect of Atorvastatin on Angiogenesis-Related Genes VEGF-A, HGF and IGF-1 and the Modulation of PI3K/AKT/mTOR Transcripts in Bone-Marrow-Derived Mesenchymal Stem Cells. Current Issues in Molecular Biology. 2023; 45(3):2326-2337. https://doi.org/10.3390/cimb45030150
Chicago/Turabian StyleAdamičková, Adriana, Nikola Chomaničová, Andrea Gažová, Juraj Maďarič, Zdenko Červenák, Simona Valášková, Matúš Adamička, and Jan Kyselovic. 2023. "Effect of Atorvastatin on Angiogenesis-Related Genes VEGF-A, HGF and IGF-1 and the Modulation of PI3K/AKT/mTOR Transcripts in Bone-Marrow-Derived Mesenchymal Stem Cells" Current Issues in Molecular Biology 45, no. 3: 2326-2337. https://doi.org/10.3390/cimb45030150