
Interpretable Classification of Tauopathies with a Convolutional Neural Network Pipeline Using Transfer Learning and Validation against Post-Mortem Clinical Cases of Alzheimer’s Disease and Progressive Supranuclear Palsy
 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
1.1. Immunodetection and Fluorescence Miscoscopy
1.2. Neurodegenerative Disease Classification Using Machine and Deep Learning
2. Materials and Methods
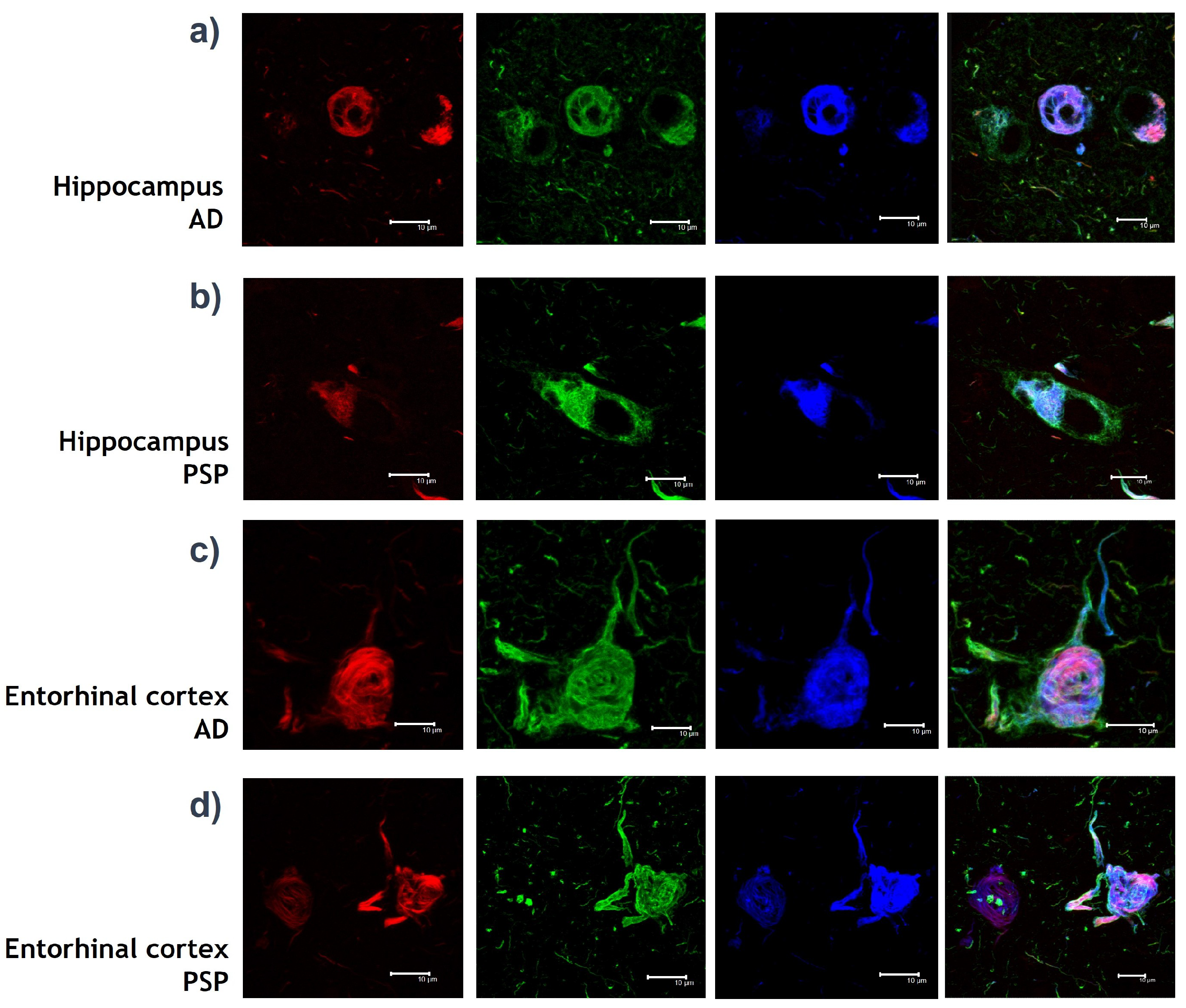
2.1. IPMB Images
- Four different brains were used.
- The brain areas used were Hippocampus CA1 and Entorhinal cortex.
- Two brains with diagnosed AD were used—one of a 90 year old female and another of an 81 year old male.
- Two male brains with diagnosed PSP were used—one of a 75 year old and another one of a 85 year old.
- The tissues of patients with AD used were of the Braak 5–6 stages.
- Green channel: Inmunodetection with AT8 mouse IgG antibody (MN1020, Invitrogene) against Tau protein. AT8 antibody detects phosphorylations of Tau protein in amino acids: Serine 202 and Threonine 305. The presence of phosphates translates into chemical changes that give protein Tau aberrant behaviour.
- Red channel: Inmunodetection with Thiazine red, which is a molecule that binds to fibrillar insoluble structures of protein polymers. This molecule specifically binds to protein Tau in its polymer conformation.
- Blue channel: Inmunodetection with pS396 rabbit IgG antibody against Tau protein [46]. The 396 antibody locates a phosphorylation in Serine 396 amino acid, which is known as a chemical change in protein Tau that associates with the formation of NFT.
- Merge channel: Visualization of the green, red and blue channel images together into one image.
2.2. Datasets
2.3. Model Development and Training
2.3.1. ResNet Models
2.3.2. Transfer Learning
- Homogeneous Transfer Learning: Source and target feature spaces are the same.
- Heterogeneous Transfer Learning: Source and target feature spaces are different.
- Transfer Learning: Initializing the weights of the model using a pre-trained architecture on the ImageNet dataset. Then, we preload these weights to train the entire architecture with our developed dataset.
- Transfer Learning and fine-tuning: Initializing the weights of the model using a pre-trained architecture on ImageNet dataset, but training only the last convolution block and the final fully connected layer with our developed dataset, for some epochs. Afterwards, we unfreeze the entire architecture and continue to train the model for additional epochs.
2.3.3. Classifier Development and Testing
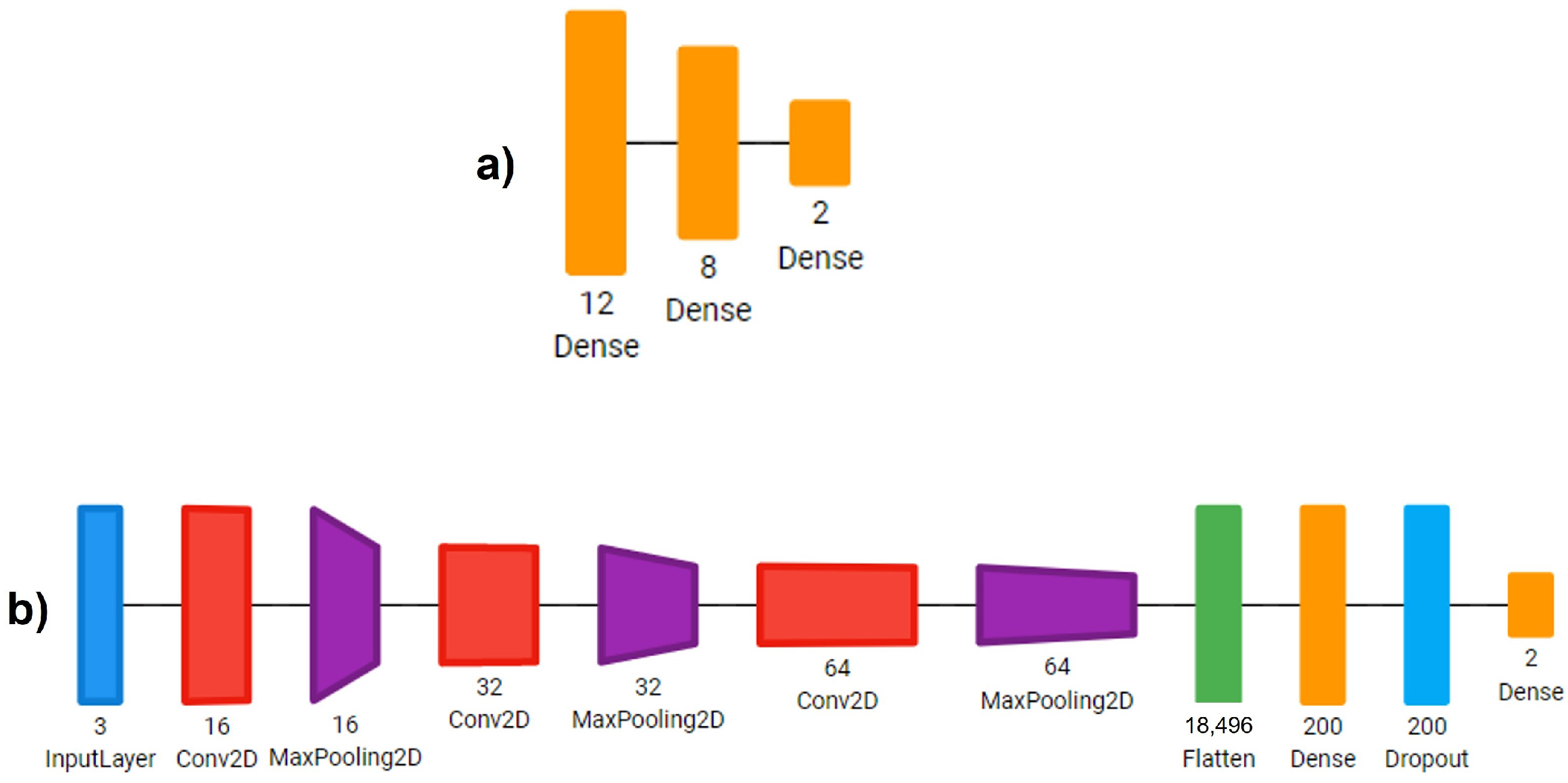
- Sequential CNN: A Multi-Layer Perceptron with three fully connected linear layers with 18, 8 and 2 neurons. L2 regularization was used for the last linear layer. The training was performed for 30 epochs.
- Simple CNN: A CNN with 3 convolution layers, starting with 16 filters, then 32 and lastly 64 filters of 3 × 3 kernels. Each convolution was followed up by a max pooling operation with a 2 × 2 window. Then, we reduced the images to a one-dimensional vector and used two fully connected layers with 200 and 2 neurons, respectively. We applied a dropout layer with a rate of 0.5 between these two fully connected layers. Each convolution and linear operation was performed with L2 regularization. The model was trained for 30 epochs.
- ResNet-IFTF: The architecture ResNet-50, as provided in Keras, was used. This architecture is pre-trained on the ImageNet dataset. Transfer Learning and fine-tuning were applied for the development of this model. We froze the entire model up to the last activation layer encountered and added a convolution layer with 512 filters with kernels of size 1 × 1, a batch normalization layer, an activation layer, a global average pooling layer and an output layer with 2 neurons and L2 regularization. The model was trained for 15 epochs with the frozen part of the architecture and then for 10 additional epochs unfreezing the entire model.
- ResNet-IFT: The architecture ResNet-50 from Keras was used. This architecture was pre-trained on the ImageNet dataset. For this model, we used Transfer Learning using the entire pre-trained RESNET without any further fine-tuning. The entire architecture was trained for 15 epochs. We added a global average pooling layer and an output layer with L2 regularization. In comparison to the original ResNet-50 architecture, we eliminated the flattening layer between the global average pooling layer and the dense layer.
2.4. Interpretation by XAI Algorithms
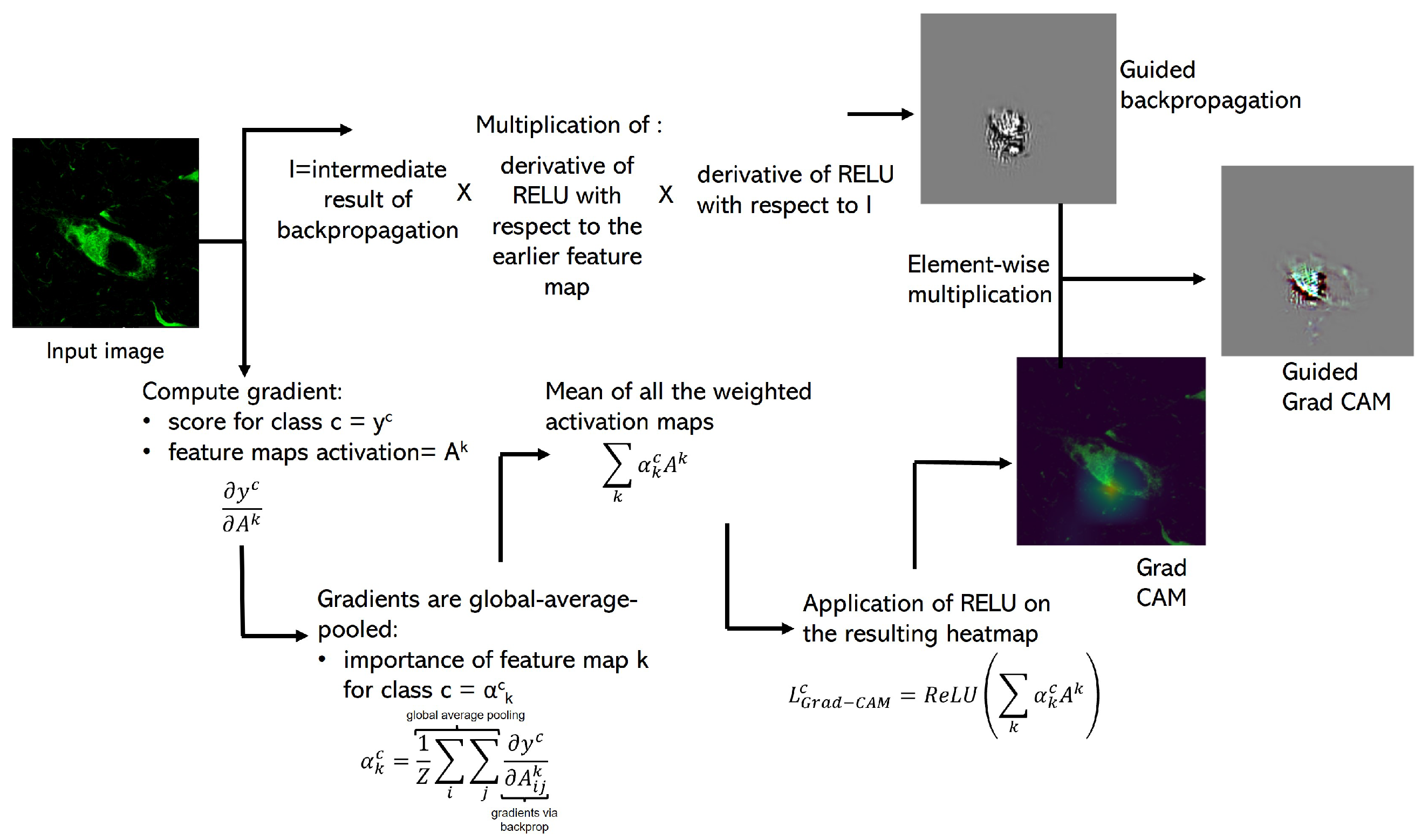
2.4.1. Guided Grad-CAM
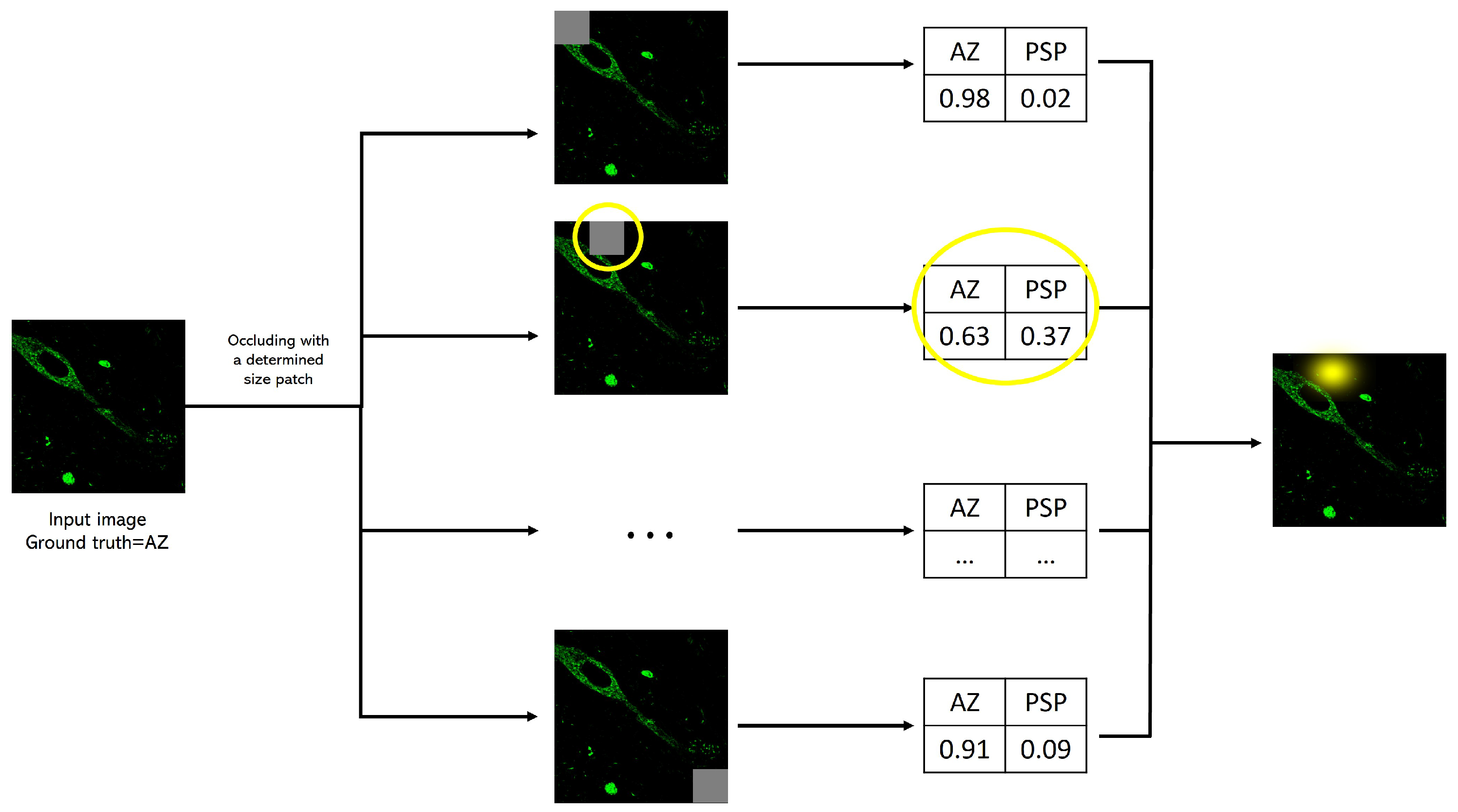
2.4.2. Occlusion Analysis
2.5. Evaluation Metrics
3. Results
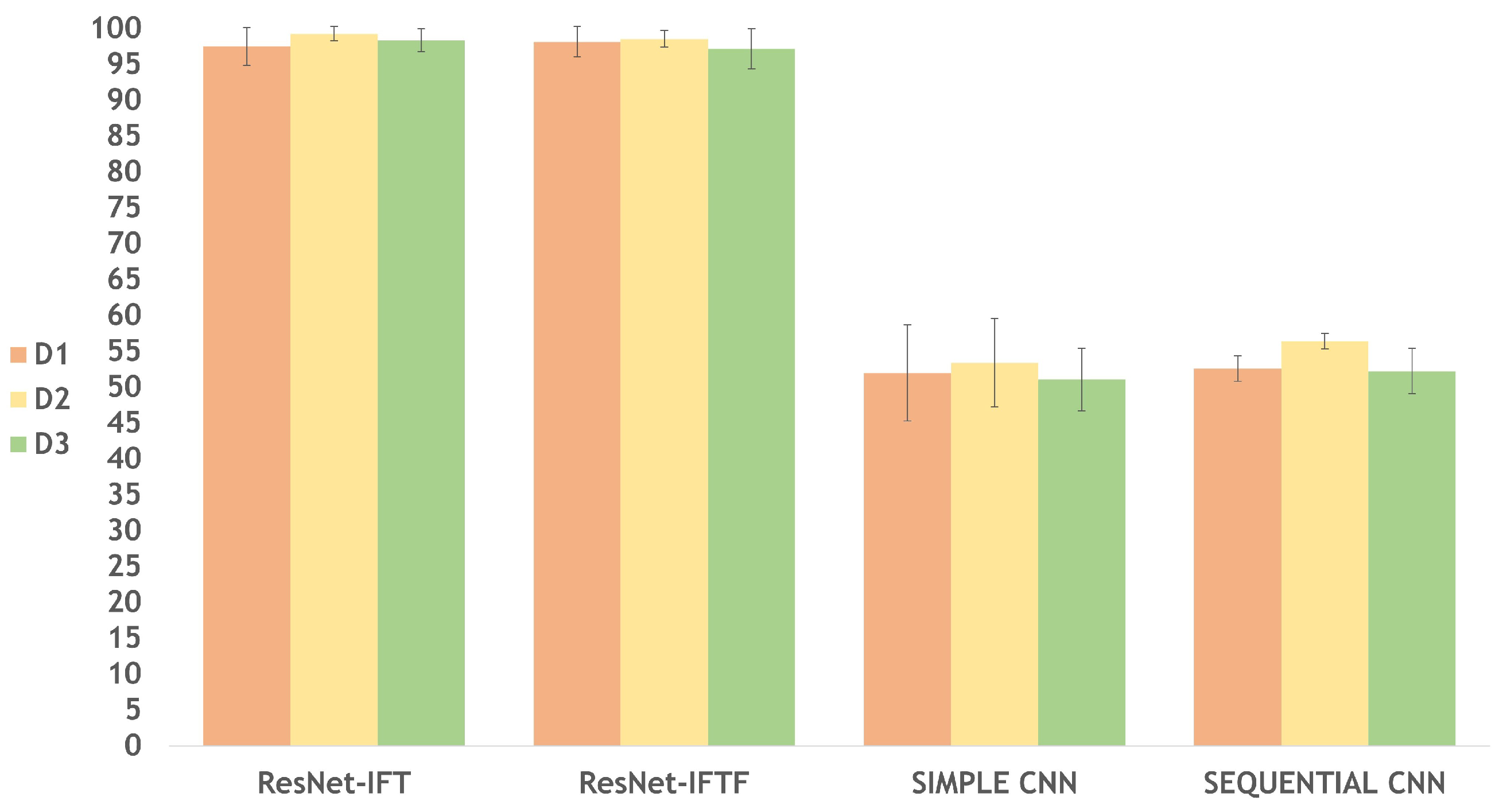
3.1. Classification Models per Dataset
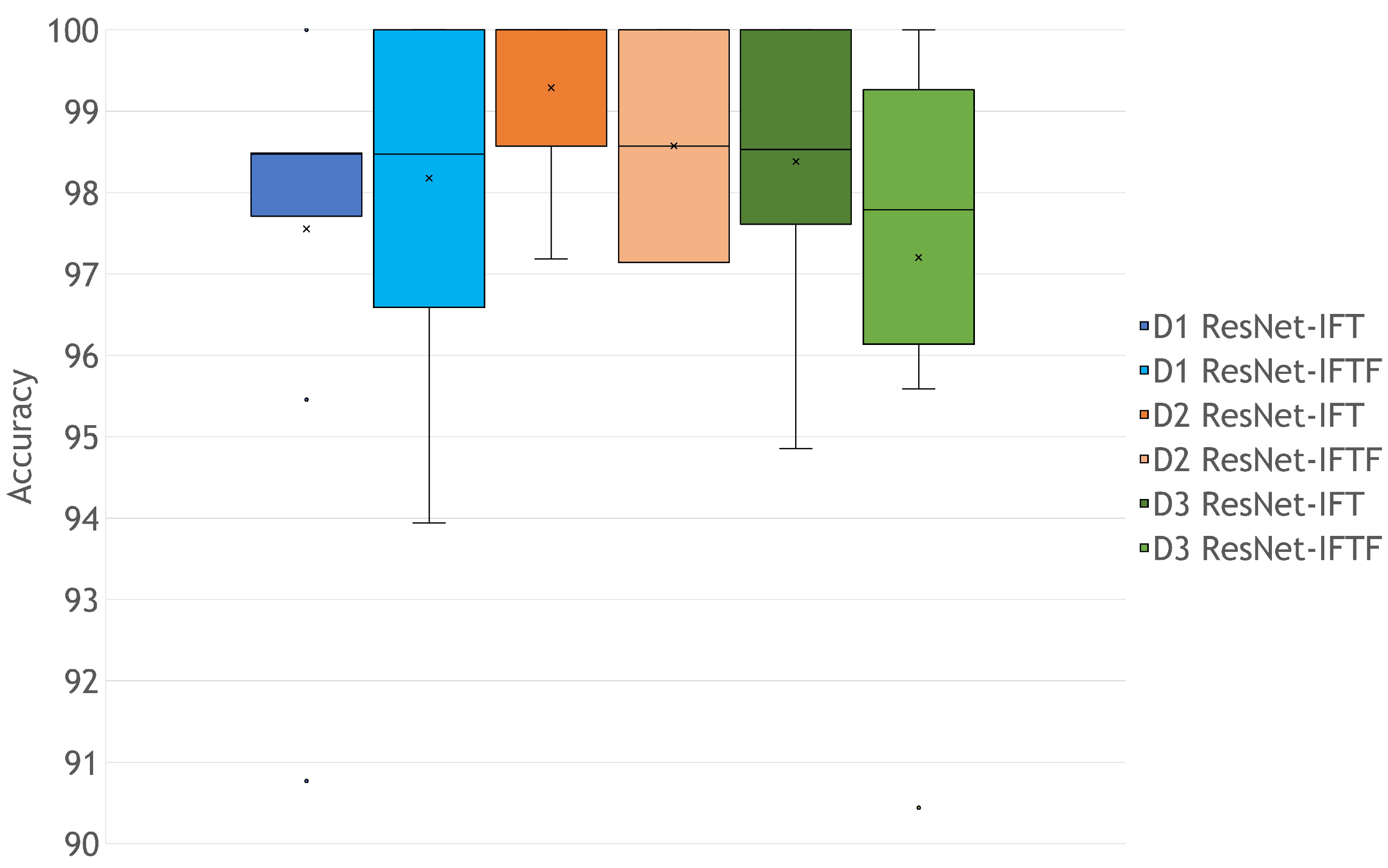
3.2. Rigor of the Classification
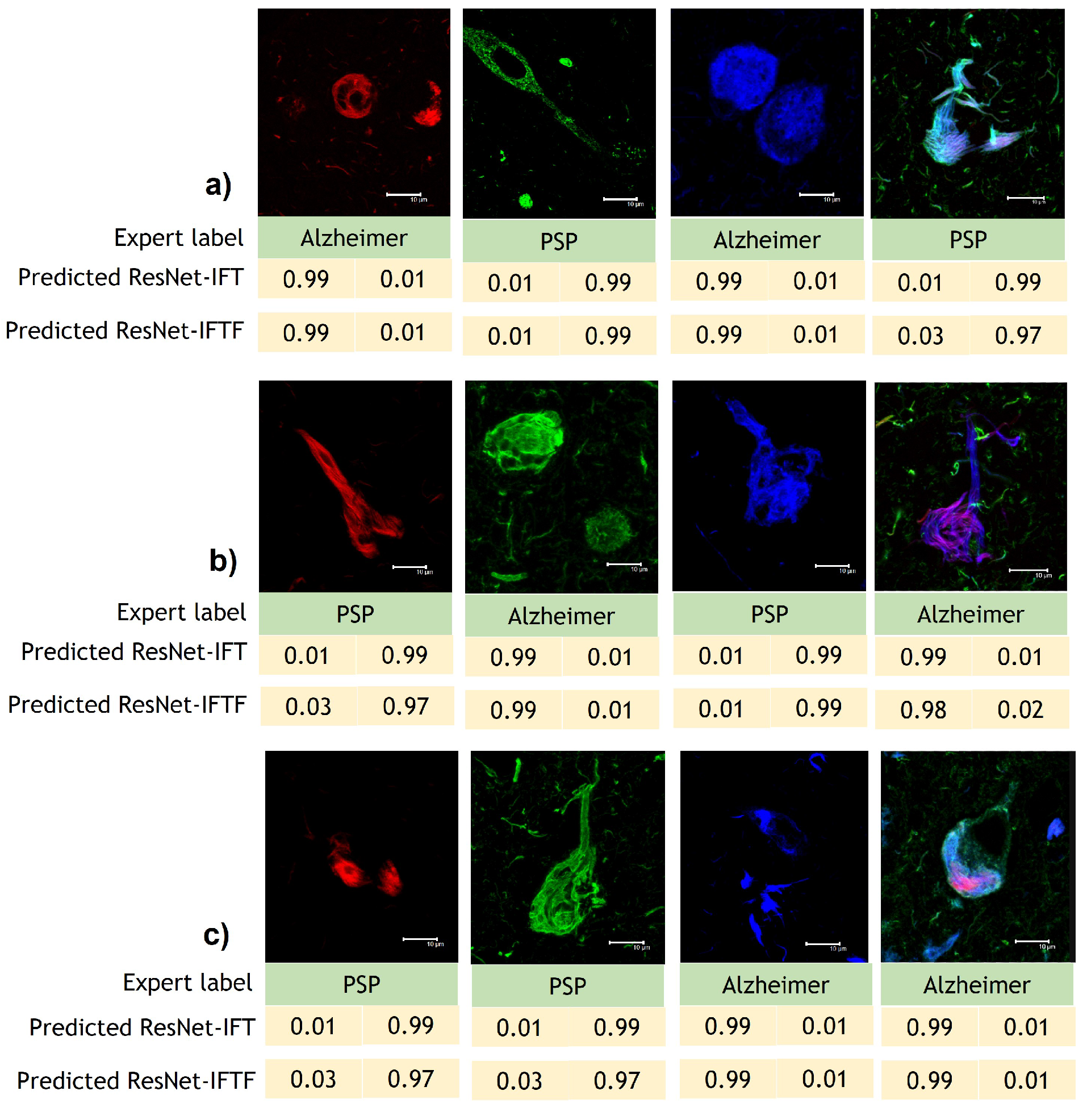
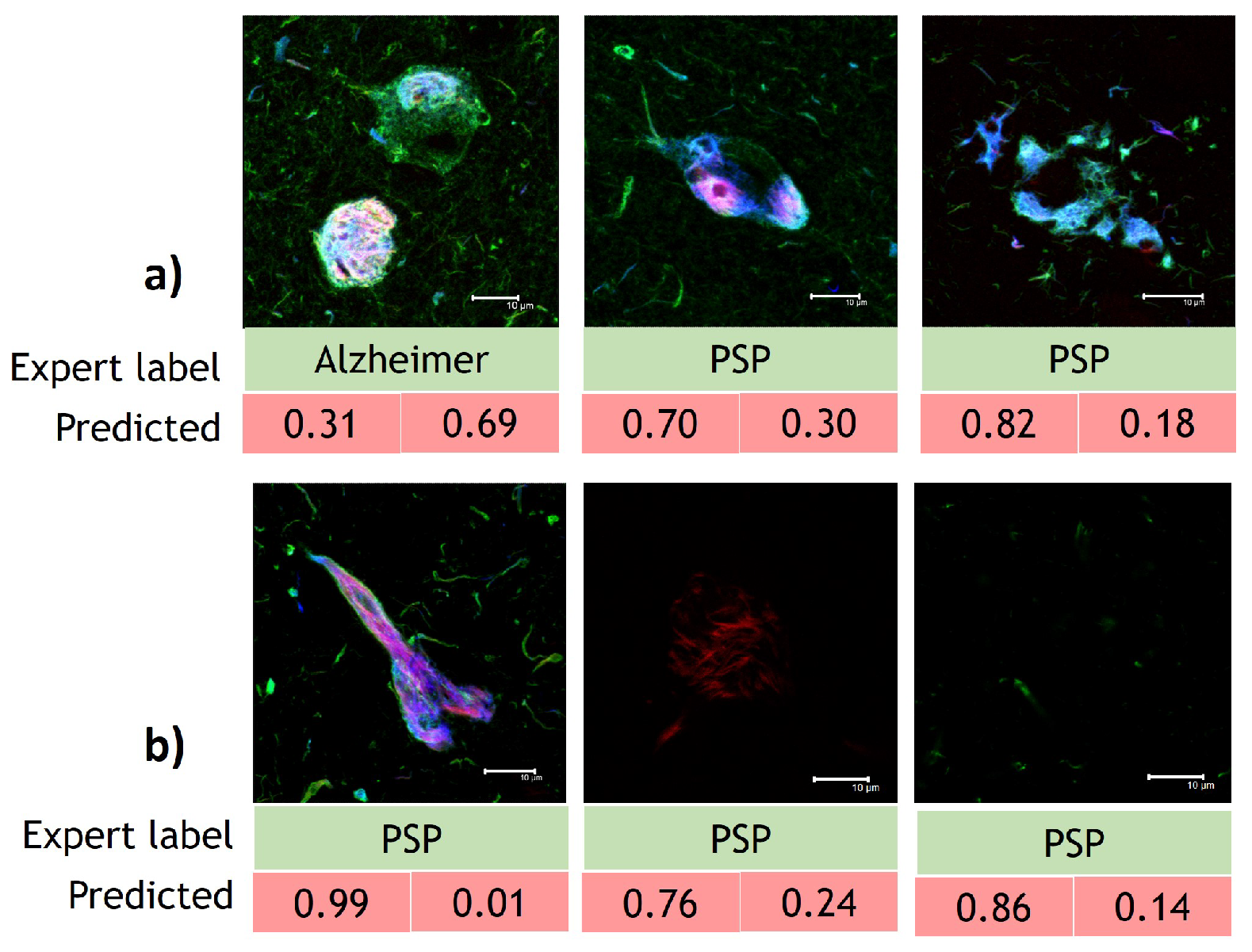
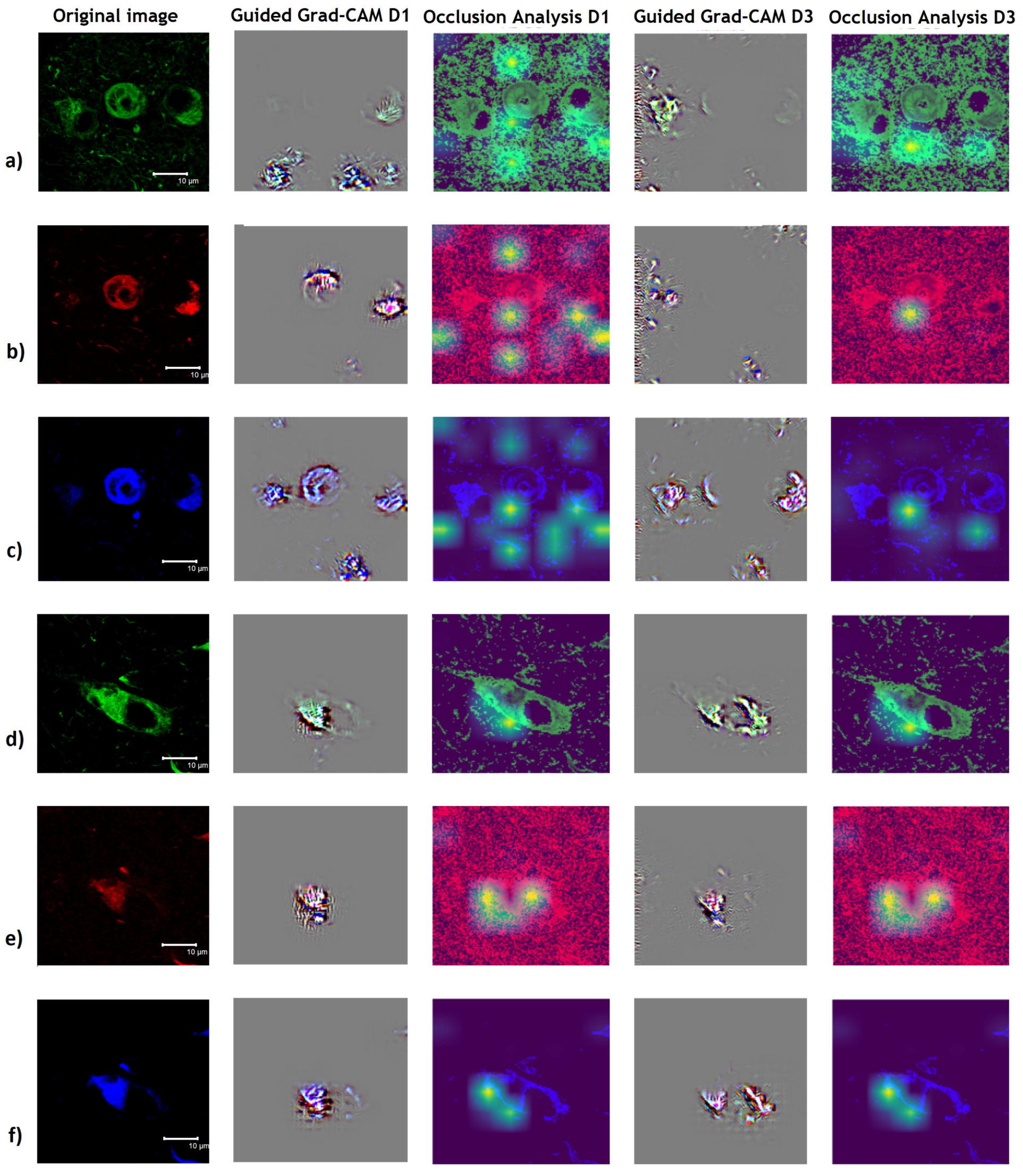
3.3. Interpretability Study
4. Discussion
4.1. Transfer Learning Model versus Fine-Tuning Model
4.2. Guided Grad-CAM and Occlusion Analysis Insights AD and PSP Classification
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NDs | Neurodegenerative diseases |
| AD | Alzheimer’s Disease |
| PSP | Progressive Supranuclear Palsy |
| CSF | Cerebrospinal Fluid |
| PTMs | Post-translational Modifications |
| CNN | Convolutional Neural Networks |
| MRI | Magnetic Resonance Imaging |
| CT | Computerized Tomography |
| PET | Positron Emission Tomography |
| IPMB | Immunofluorescence Post-Mortem Brain |
| XAI | Explainable Artificial Intelligence |
| UNAM | National Autonomous University of Mexico |
References
- Dugger, B.N.; Dickson, D.W. Pathology of neurodegenerative diseases. Cold Spring Harb. Perspect. Biol. 2017, 9, a028035. [Google Scholar] [CrossRef] [Green Version]
- Shakir, M.N.; Dugger, B.N. Advances in Deep Neuropathological Phenotyping of Alzheimer Disease: Past, Present and Future. J. Neuropathol. Exp. Neurol. 2022, 81, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Mena, R.; Edwards, P.; Pérez-Olvera, O.; Wischik, C.M. Monitoring pathological assembly of Tau and β-amyloid proteins in Alzheimer’s disease. Acta Neuropathol. 1995, 89, 50–56. [Google Scholar] [CrossRef]
- Silva, M.C.; Haggarty, S.J. Tauopathies: Deciphering disease mechanisms to develop effective therapies. Int. J. Mol. Sci. 2020, 21, 8948. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Liu, F.; Gong, C.X. Tau and neurodegenerative disease: The story so far. Nat. Rev. Neurol. 2016, 12, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Tarawneh, R.; Holtzman, D.M. The clinical problem of symptomatic Alzheimer disease and mild cognitive impairment. Cold Spring Harb. Perspect. Med. 2012, 2, a006148. [Google Scholar] [CrossRef]
- Martínez-Maldonado, A.; Ontiveros-Torres, M.Á.; Harrington, C.R.; Montiel-Sosa, J.F.; Prandiz, R.G.T.; Bocanegra-López, P.; Sorsby-Vargas, A.M.; Bravo-Muñoz, M.; Florán-Garduño, B.; Villanueva-Fierro, I.; et al. Molecular Processing of Tau Protein in Progressive Supranuclear Palsy: Neuronal and Glial Degeneration. J. Alzheimer’s Dis. 2021, 79, 1517–1531. [Google Scholar] [CrossRef]
- Santpere, G.; Ferrer, I. Delineation of early changes in cases with progressive supranuclear palsy-like pathology. Astrocytes in striatum are primary targets of tau phosphorylation and GFAP oxidation. Brain Pathol. 2009, 19, 177–187. [Google Scholar] [CrossRef]
- Kovacs, G.G.; Lukic, M.J.; Irwin, D.J.; Arzberger, T.; Respondek, G.; Lee, E.B.; Coughlin, D.; Giese, A.; Grossman, M.; Kurz, C.; et al. Distribution patterns of Tau pathology in progressive supranuclear palsy. Acta Neuropathol. 2020, 140, 99–119. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, G.G. Tauopathies. Handb. Clin. Neurol. 2018, 145, 355–368. [Google Scholar]
- Golbe, L.I. Progressive Supranuclear Palsy. In Proceedings of the Seminars in Neurology; Thieme Medical Publishers: New York, NY, USA, 2014; Volume 34, pp. 151–159. [Google Scholar]
- Prince, M.; Wimo, A.; Guerchet, M.; Ali, G.; Wu, Y.T.; Prina, M. World Alzheimer Report 2015—The Global Impact of Dementia: An Analysis of Prevalence, Incidence, Cost and Trends; Alzheimer’s Disease International: London, UK, 2015. [Google Scholar]
- Rohini, M.; Surendran, D. Classification of neurodegenerative disease stages using ensemble Machine Learning classifiers. Procedia Comput. Sci. 2019, 165, 66–73. [Google Scholar] [CrossRef]
- Simon, A. Neurodegenerative Diseases: Overview, Perspectives and Emerging Treatments; Neurology—Laboratory and Clinical Research Developments Series; Nova Science Publishers, Incorporated: Hauppauge, NY, USA, 2017. [Google Scholar]
- Luna-Muñoz, J.; García-Sierra, F.; Falcón, V.; Menéndez, I.; Chávez-Macías, L.; Mena, R. Regional conformational change involving phosphorylation of tau protein at the Thr 231, precedes the structural change detected by Alz-50 antibody in Alzheimer’s disease. J. Alzheimer’s Dis. 2005, 8, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Savastano, A.; Flores, D.; Kadavath, H.; Biernat, J.; Mandelkow, E.; Zweckstetter, M. Disease-associated Tau phosphorylation hinders tubulin assembly within Tau condensates. Angew. Chem. Int. Ed. 2021, 60, 726–730. [Google Scholar] [CrossRef]
- Luna-Viramontes, N.I.; Campa-Córdoba, B.B.; Ontiveros-Torres, M.Á.; Harrington, C.R.; Villanueva-Fierro, I.; Guadarrama-Ortíz, P.; Garcés-Ramírez, L.; de la Cruz, F.; Hernandes-Alejandro, M.; Martínez-Robles, S.; et al. PHF-core Tau as the potential initiating event for Tau pathology in Alzheimer’s disease. Front. Cell. Neurosci. 2020, 14, 247. [Google Scholar] [CrossRef] [PubMed]
- Rani, L.; Mallajosyula, S.S. Phosphorylation-induced structural reorganization in Tau-paired helical filaments. ACS Chem. Neurosci. 2021, 12, 1621–1631. [Google Scholar] [CrossRef] [PubMed]
- Ontiveros-Torres, M.Á.; Labra-Barrios, M.L.; Diaz-Cintra, S.; Aguilar-Vázquez, A.R.; Moreno-Campuzano, S.; Flores-Rodriguez, P.; Luna-Herrera, C.; Mena, R.; Perry, G.; Floran-Garduno, B.; et al. Fibrillar amyloid-β accumulation triggers an inflammatory mechanism leading to hyperphosphorylation of the carboxyl-terminal end of tau polypeptide in the hippocampal formation of the 3× Tg-AD transgenic mouse. J. Alzheimer’s Dis. 2016, 52, 243–269. [Google Scholar] [CrossRef] [PubMed]
- Morozova, A.; Zorkina, Y.; Abramova, O.; Pavlova, O.; Pavlov, K.; Soloveva, K.; Volkova, M.; Alekseeva, P.; Andryshchenko, A.; Kostyuk, G.; et al. Neurobiological Highlights of Cognitive Impairment in Psychiatric Disorders. Int. J. Mol. Sci. 2022, 23, 1217. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, Z.; Chen, P.; Cheng, P.; Zhao, K.; Zhang, H.; Shu, H.; Gu, L.; Gao, L.; Wang, Q.; et al. Episodic Memory-related Imaging Features as Valuable Biomarkers for the Diagnosis of Alzheimer’s Disease: A Multicenter Study Based on Machine Learning. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2020, 23, 1217. [Google Scholar] [CrossRef]
- Yu, H.; Yang, L.T.; Zhang, Q.; Armstrong, D.; Deen, M.J. Convolutional Neural Networks for medical image analysis: State-of-the-art, comparisons, improvement and perspectives. Neurocomputing 2021, 444, 92–110. [Google Scholar] [CrossRef]
- Lima, A.A.; Mridha, M.F.; Das, S.C.; Kabir, M.M.; Islam, M.R.; Watanobe, Y. A Comprehensive Survey on the Detection, Classification and Challenges of Neurological Disorders. Biology 2022, 11, 469. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ip, P.; Chakrabartty, A. Simple Elimination of Background Fluorescence in Formalin-Fixed Human Brain Tissue for Immunofluorescence Microscopy. J. Vis. Exp. 2017, 127, 56188. [Google Scholar] [CrossRef] [PubMed]
- Abdeladim, L.; Matho, K.S.; Clavreul, S.; Mahou, P.; Sintes, J.M.; Solinas, X.; Arganda-Carreras, I.; Turney, S.; Lichtman, J.; Chessel, A.; et al. Multicolor multiscale brain imaging with chromatic multiphoton serial microscopy. Nat. Commun. 2019, 10, 1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, G.; Samavedham, L.; Lim, E.C.H.; Alzheimer’s Disease Neuroimaging Initiative; Parkinson Progression Marker Initiative. Determination of imaging biomarkers to decipher disease trajectories and differential diagnosis of neurodegenerative diseases (DIsease TreND). J. Neurosci. Methods 2018, 305, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Sarvamangala, D.; Kulkarni, R.V. Convolutional Neural Networks in medical image understanding: A survey. Evol. Intell. 2021, 15, 1–22. [Google Scholar] [CrossRef]
- Tang, Z.; Chuang, K.V.; DeCarli, C.; Jin, L.W.; Beckett, L.; Keiser, M.J.; Dugger, B.N. Interpretable classification of Alzheimer’s disease pathologies with a Convolutional Neural Network pipeline. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Rajpurkar, P.; Irvin, J.; Zhu, K.; Yang, B.; Mehta, H.; Duan, T.; Ding, D.; Bagul, A.; Langlotz, C.; Shpanskaya, K.; et al. Chexnet: Radiologist-level pneumonia detection on chest x-rays with Deep Learning. arXiv 2017, arXiv:1711.05225. [Google Scholar]
- Phillips, N.A.; Rajpurkar, P.; Sabini, M.; Krishnan, R.; Zhou, S.; Pareek, A.; Phu, N.M.; Wang, C.; Ng, A.Y.; Lungren, M.P. Chexphoto: 10,000+ smartphone photos and synthetic photographic transformations of chest x-rays for benchmarking Deep Learning robustness. arXiv 2020, arXiv:2007.06199. [Google Scholar]
- Liu, X.; Wang, C.; Bai, J.; Liao, G. Fine-Tuning pre-trained Convolutional Neural Networks for gastric precancerous disease classification on magnification narrow-band imaging images. Neurocomputing 2020, 392, 253–267. [Google Scholar] [CrossRef]
- Zhuang, F.; Qi, Z.; Duan, K.; Xi, D.; Zhu, Y.; Zhu, H.; Xiong, H.; He, Q. A comprehensive survey on Transfer Learning. Proc. IEEE 2020, 109, 43–76. [Google Scholar] [CrossRef]
- Deng, J.; Dong, W.; Socher, R.; Li, L.J.; Li, K.; Fei-Fei, L. Imagenet: A large-scale hierarchical image database. In Proceedings of the 2009 IEEE Conference on Computer Vision and Pattern Recognition, Miami, FL, USA, 20–25 June 2009; IEEE: Piscataway, NJ, USA, 2009; pp. 248–255. [Google Scholar]
- Krizhevsky, A.; Sutskever, I.; Hinton, G.E. Imagenet classification with deep Convolutional Neural Networks. Adv. Neural Inf. Process. Syst. 2012, 25, 1106–1114. [Google Scholar] [CrossRef] [Green Version]
- Maqsood, M.; Nazir, F.; Khan, U.; Aadil, F.; Jamal, H.; Mehmood, I.; Song, O.y. Transfer Learning assisted classification and detection of Alzheimer’s disease stages using 3D MRI scans. Sensors 2019, 19, 2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, S.; Wang, L.; Sun, C.; Zhou, L. Deep Learning based HEp-2 image classification: A comprehensive review. Med. Image Anal. 2020, 65, 101764. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Han, T.; Zhou, F.; Yu, Z.; Qin, J.; Elazab, A.; Lei, B. A deeply supervised residual network for HEp-2 cell classification via cross-modal Transfer Learning. Pattern Recognit. 2018, 79, 290–302. [Google Scholar] [CrossRef]
- Rodrigues, L.F.; Naldi, M.C.; Mari, J.F. Comparing Convolutional Neural Networks and preprocessing techniques for HEp-2 cell classification in immunofluorescence images. Comput. Biol. Med. 2020, 116, 103542. [Google Scholar] [CrossRef]
- Yamashiro, K.; Liu, J.; Matsumoto, N.; Ikegaya, Y. Deep Learning-based classification of GAD67-positive neurons without the immunosignal. Front. Neuroanat. 2021, 15, 643067. [Google Scholar] [CrossRef]
- Ligabue, G.; Pollastri, F.; Fontana, F.; Leonelli, M.; Furci, L.; Giovanella, S.; Alfano, G.; Cappelli, G.; Testa, F.; Bolelli, F.; et al. Evaluation of the classification accuracy of the kidney biopsy direct immunofluorescence through Convolutional Neural Networks. Clin. J. Am. Soc. Nephrol. 2020, 15, 1445–1454. [Google Scholar] [CrossRef]
- Yetiş, S.Ç.; Çapar, A.; Ekinci, D.A.; Ayten, U.E.; Kerman, B.E.; Töreyin, B.U. Myelin detection in fluorescence microscopy images using machine learning. J. Neurosci. Methods 2020, 346, 108946. [Google Scholar] [CrossRef]
- Alegro, M.; Theofilas, P.; Nguy, A.; Castruita, P.A.; Seeley, W.; Heinsen, H.; Ushizima, D.M.; Grinberg, L.T. Automating cell detection and classification in human brain fluorescent microscopy images using dictionary learning and sparse coding. J. Neurosci. Methods 2017, 282, 20–33. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.H.; Chiu, S.I.; Chen, T.F.; Jang, J.S.R.; Chiu, M.J. Classifications of neurodegenerative disorders using a multiplex blood biomarkers-based Machine Learning model. Int. J. Mol. Sci. 2020, 21, 6914. [Google Scholar] [CrossRef]
- Gao, X.W.; Hui, R.; Tian, Z. Classification of CT brain images based on Deep Learning networks. Comput. Methods Programs Biomed. 2017, 138, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bramblett, G.T.; Goedert, M.; Jakes, R.; Merrick, S.E.; Trojanowski, J.Q.; Lee, V.M. Abnormal Tau phosphorylation at Ser396 in Alzheimer’s disease recapitulates development and contributes to reduced microtubule binding. Neuron 1993, 10, 1089–1099. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.J.; Yang, Q. A survey on Transfer Learning. IEEE Trans. Knowl. Data Eng. 2009, 22, 1345–1359. [Google Scholar] [CrossRef]
- Torrey, L.; Shavlik, J. Transfer Learning. In Handbook of Research on Machine Learning Applications and Trends: Algorithms, Methods and Techniques; IGI Global: Hershey, PA, USA, 2010; pp. 242–264. [Google Scholar]
- He, K.; Zhang, X.; Ren, S.; Sun, J. Deep residual learning for image recognition. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Las Vegas, NV, USA, 27–30 June 2016; pp. 770–778. [Google Scholar]
- Maeda-Gutierrez, V.; Galvan-Tejada, C.E.; Zanella-Calzada, L.A.; Celaya-Padilla, J.M.; Galván-Tejada, J.I.; Gamboa-Rosales, H.; Luna-Garcia, H.; Magallanes-Quintanar, R.; Guerrero Mendez, C.A.; Olvera-Olvera, C.A. Comparison of Convolutional Neural Network architectures for classification of tomato plant diseases. Appl. Sci. 2020, 10, 1245. [Google Scholar] [CrossRef] [Green Version]
- Talo, M. Convolutional Neural Networks for multi-class histopathology image classification. arXiv 2019, arXiv:1903.10035. [Google Scholar]
- Hussain, M.; Bird, J.J.; Faria, D.R. A study on cnn Transfer Learning for image classification. In Proceedings of the UK Workshop on Computational Intelligence, Nottingham, UK, 5–7 September 2018; Springer: Berlin/Heidelberg, Germany, 2018; pp. 191–202. [Google Scholar]
- Bäuerle, A.; van Onzenoodt, C.; Ropinski, T. Net2Vis—A Visual Grammar for Automatically Generating Publication-Tailored CNN Architecture Visualizations. IEEE Trans. Vis. Comput. Graph. 2021, 27, 2980–2991. [Google Scholar] [CrossRef]
- Selvaraju, R.R.; Cogswell, M.; Das, A.; Vedantam, R.; Parikh, D.; Batra, D. Grad-cam: Visual explanations from deep networks via gradient-based localization. In Proceedings of the IEEE International Conference on Computer Vision, Venice, Italy, 22–29 October 2017; pp. 618–626. [Google Scholar]
- Khandelwal, R. How to Visually Explain Any CNN Based Models? 2020. Available online: https://towardsdatascience.com/how-to-visually-explain-any-cnn-based-models-80e0975ce57 (accessed on 1 January 2022).
- Zeiler, M.D.; Fergus, R. Visualizing and understanding convolutional networks. In Proceedings of the European Conference on Computer Vision, Zurich, Switzerland, 6–12 September 2014; Springer: Berlin/Heidelberg, Germany, 2014; pp. 818–833. [Google Scholar]
- Meudec, R. tf-explain. 2021. Available online: https://doi.org/10.5281/zenodo.5711704 (accessed on 1 January 2022).
- Remy, P. Keract: A Library for Visualizing Activations and Gradients. 2020. Available online: https://github.com/philipperemy/keract (accessed on 1 March 2022).
- Guo, Y.; Shi, H.; Kumar, A.; Grauman, K.; Rosing, T.; Feris, R. Spottune: Transfer Learning through adaptive Fine-Tuning. In Proceedings of the IEEE/CVF Conference on Computer Vision and Pattern Recognition, Long Beach, CA, USA, 15–20 June 2019; pp. 4805–4814. [Google Scholar]
- Veit, A.; Wilber, M.J.; Belongie, S. Residual networks behave like ensembles of relatively shallow networks. Adv. Neural Inf. Process. Syst. 2016, 29, 550–558. [Google Scholar]
- Peng, P.; Wang, J. How to fine-tune deep neural networks in few-shot learning? arXiv 2020, arXiv:2012.00204. [Google Scholar]
- Koson, P.; Zilka, N.; Kovac, A.; Kovacech, B.; Korenova, M.; Filipcik, P.; Novak, M. Truncated Tau expression levels determine life span of a rat model of tauopathy without causing neuronal loss or correlating with terminal neurofibrillary tangle load. Eur. J. Neurosci. 2008, 28, 239–246. [Google Scholar] [CrossRef]
- Guillozet-Bongaarts, A.L.; Garcia-Sierra, F.; Reynolds, M.R.; Horowitz, P.M.; Fu, Y.; Wang, T.; Cahill, M.E.; Bigio, E.H.; Berry, R.W.; Binder, L.I. Tau truncation during neurofibrillary tangle evolution in Alzheimer’s disease. Neurobiol. Aging 2005, 26, 1015–1022. [Google Scholar] [CrossRef]
- Kellogg, E.H.; Hejab, N.M.; Poepsel, S.; Downing, K.H.; DiMaio, F.; Nogales, E. Near-atomic model of microtubule-tau interactions. Science 2018, 360, 1242–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantrelle, F.X.; Loyens, A.; Trivelli, X.; Reimann, O.; Despres, C.; Gandhi, N.S.; Hackenberger, C.P.; Landrieu, I.; Smet-Nocca, C. Phosphorylation and O-GlcNAcylation of the PHF-1 Epitope of Tau Protein Induce Local Conformational Changes of the C-terminus and modulate Tau self-assembly into fibrillar aggregates. Front. Mol. Neurosci. 2021, 14, 661368. [Google Scholar] [CrossRef] [PubMed]
- Rosenqvist, N.; Asuni, A.A.; Andersson, C.R.; Christensen, S.; Daechsel, J.A.; Egebjerg, J.; Falsig, J.; Helboe, L.; Jul, P.; Kartberg, F.; et al. Highly specific and selective anti-pS396-tau antibody C10. 2 targets seeding-competent tau. Alzheimer’s Dementia Transl. Res. Clin. Interv. 2018, 4, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Fadul, M.M.; Garwood, C.J.; Waller, R.; Garrett, N.; Heath, P.R.; Matthews, F.E.; Brayne, C.; Wharton, S.B.; Simpson, J.E. NDRG2 Expression Correlates with Neurofibrillary Tangles and Microglial Pathology in the Ageing Brain. Int. J. Mol. Sci. 2020, 21, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloret, A.; Esteve, D.; Lloret, M.A.; Cervera-Ferri, A.; Lopez, B.; Nepomuceno, M.; Monllor, P. When does Alzheimer’s disease really start? The role of biomarkers. Int. J. Mol. Sci. 2019, 20, 5536. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset Label | Brain Regions Included | Classes | Number of Images per Class | Total Images |
|---|---|---|---|---|
| D1 | Hippocampus | Alzheimer | 346 | 656 |
| PSP | 310 | |||
| D2 | Entorhinal cortex | Alzheimer | 393 | 702 |
| PSP | 309 | |||
| D3 | Hippocampus and Entorhinal cortex | Alzheimer | 739 | 1358 |
| PSP | 619 |
| Architecture Label | Structure | Pretrained | Transfer/Fine Tuning |
|---|---|---|---|
| ResNet-IFT | 48-convolution layers | Yes, ImageNet | Transfer |
| 1 Global Average Pooling | |||
| 1 Dense Layer | |||
| ResNet-IFTF | 49-convolution layers | Yes, ImageNet | Transfer and Fine Tuning |
| 1 Global Average Pooling | |||
| 1 Dense Layer | |||
| Simple CNN | 3 blocks of convolution layer and max pooling | No | No |
| 2 Dense Layer | |||
| Sequential CNN | 3 Dense layers | No | No |
| Architecture Label | Dataset | Accuracy |
|---|---|---|
| ResNet-IFT | D1 | 97.55 ± 2.63 |
| D2 | 99.29 ± 1.00 | |
| D3 | 98.38 ± 1.58 | |
| ResNet-IFTF | D1 | 98.18 ± 2.12 |
| D2 | 98.57 ± 1.17 | |
| D3 | 97.20 ± 2.83 | |
| Simple CNN | D1 | 52.00 ± 6.71 |
| D2 | 53.42 ± 6.14 | |
| D3 | 51.11 ± 4.35 | |
| Sequential CNN | D1 | 52.60 ± 1.76 |
| D2 | 56.41 ± 1.07 | |
| D3 | 52.36 ± 3.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diaz-Gomez, L.; Gutierrez-Rodriguez, A.E.; Martinez-Maldonado, A.; Luna-Muñoz, J.; Cantoral-Ceballos, J.A.; Ontiveros-Torres, M.A. Interpretable Classification of Tauopathies with a Convolutional Neural Network Pipeline Using Transfer Learning and Validation against Post-Mortem Clinical Cases of Alzheimer’s Disease and Progressive Supranuclear Palsy. Curr. Issues Mol. Biol. 2022, 44, 5963-5985. https://doi.org/10.3390/cimb44120406
Diaz-Gomez L, Gutierrez-Rodriguez AE, Martinez-Maldonado A, Luna-Muñoz J, Cantoral-Ceballos JA, Ontiveros-Torres MA. Interpretable Classification of Tauopathies with a Convolutional Neural Network Pipeline Using Transfer Learning and Validation against Post-Mortem Clinical Cases of Alzheimer’s Disease and Progressive Supranuclear Palsy. Current Issues in Molecular Biology. 2022; 44(12):5963-5985. https://doi.org/10.3390/cimb44120406
Chicago/Turabian StyleDiaz-Gomez, Liliana, Andres E. Gutierrez-Rodriguez, Alejandra Martinez-Maldonado, Jose Luna-Muñoz, Jose A. Cantoral-Ceballos, and Miguel A. Ontiveros-Torres. 2022. "Interpretable Classification of Tauopathies with a Convolutional Neural Network Pipeline Using Transfer Learning and Validation against Post-Mortem Clinical Cases of Alzheimer’s Disease and Progressive Supranuclear Palsy" Current Issues in Molecular Biology 44, no. 12: 5963-5985. https://doi.org/10.3390/cimb44120406