
A Combination of Deep-Sea Water and Fucoidan Alleviates T2DM through Modulation of Gut Microbiota and Metabolic Pathways
 ,
,
Abstract
:1. Introduction
2. Results
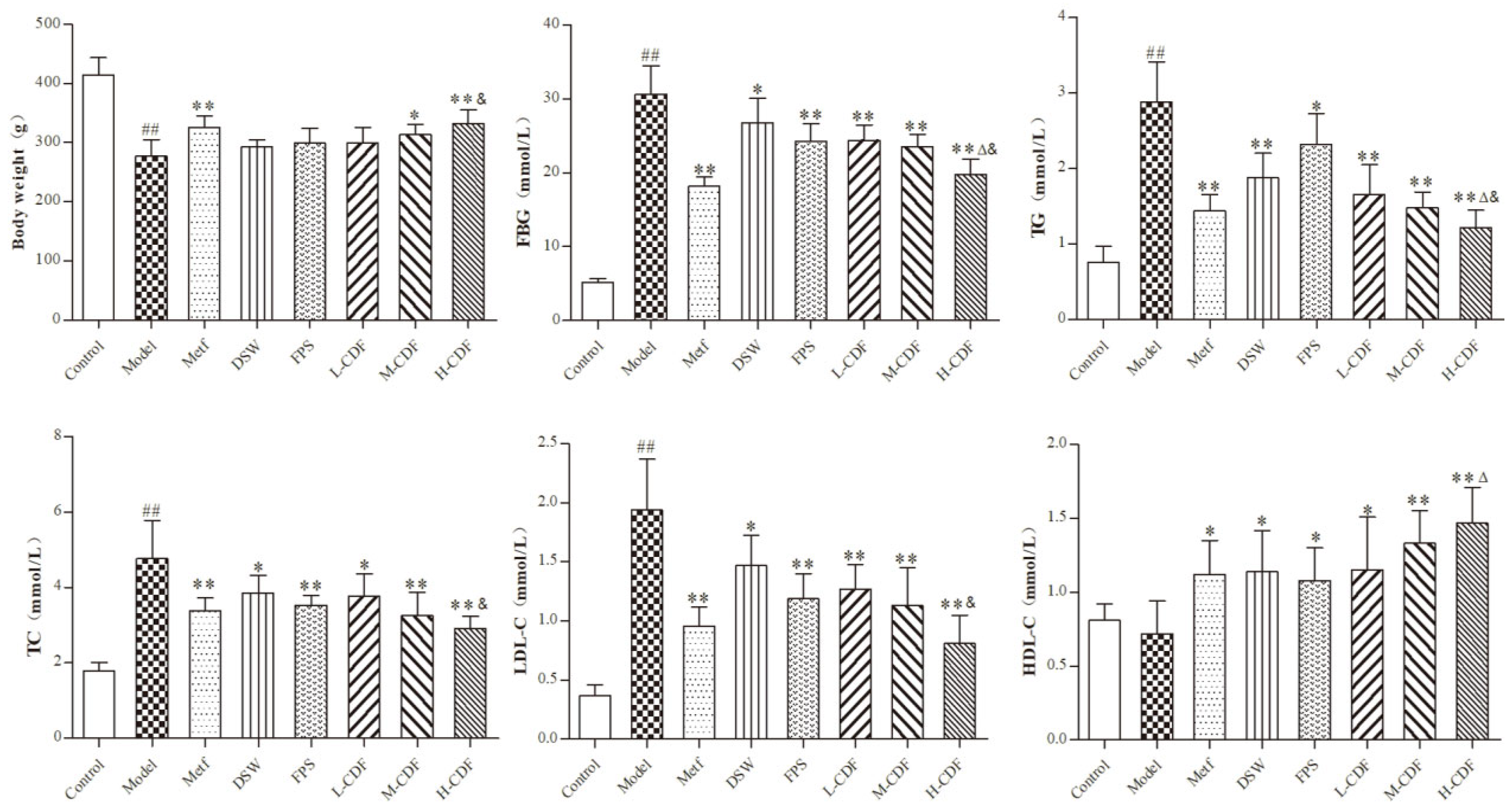
2.1. H-CDF Results in Better Improvement in Blood Glucolipid-Related Indices in T2DM Rats

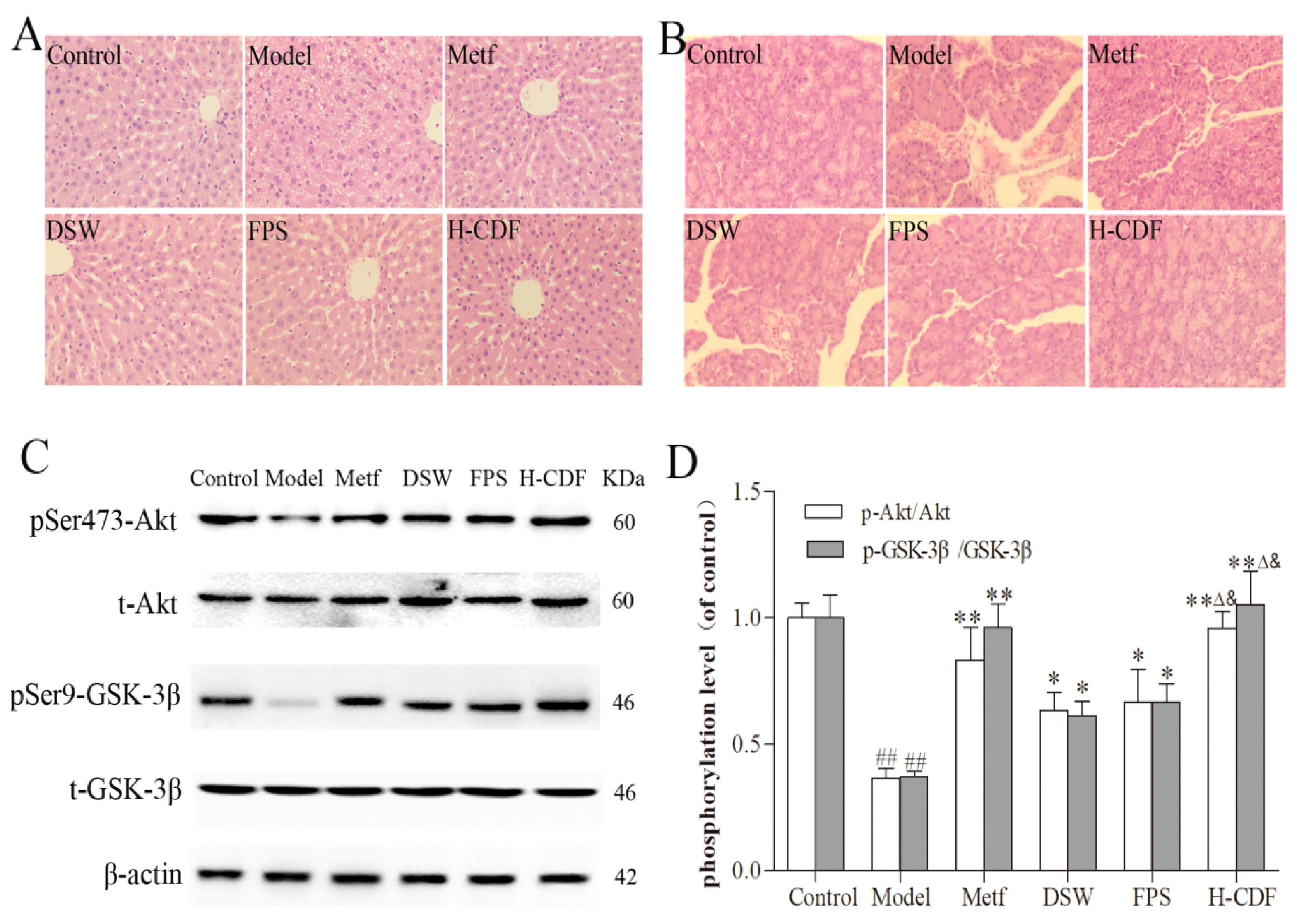
2.2. H-CDF Preferably Improves Histopathological Changes and Akt/GSK3β Signaling in T2DM Rats
2.3. H-CDF Improves Metabolic Disorders in T2DM Rats
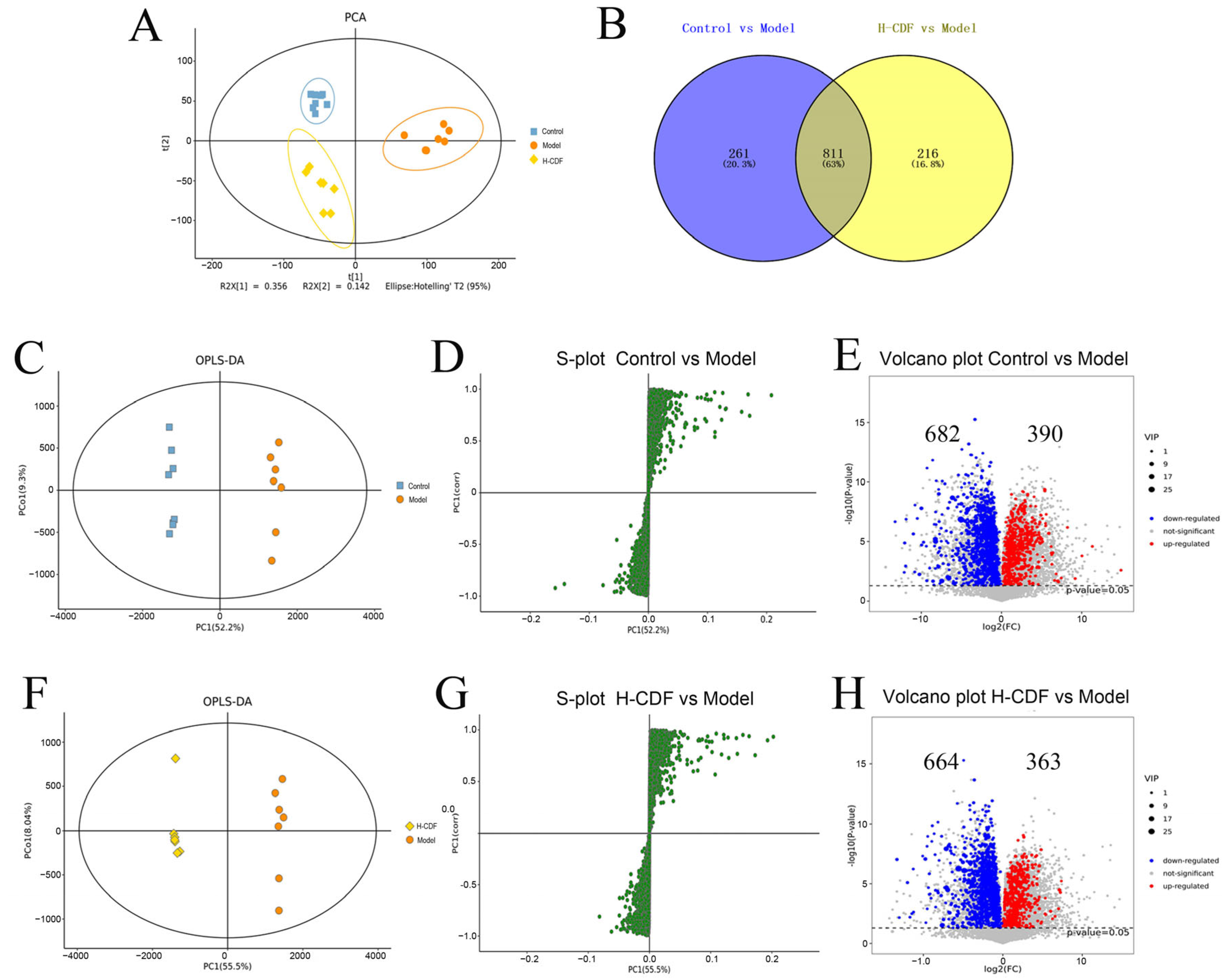
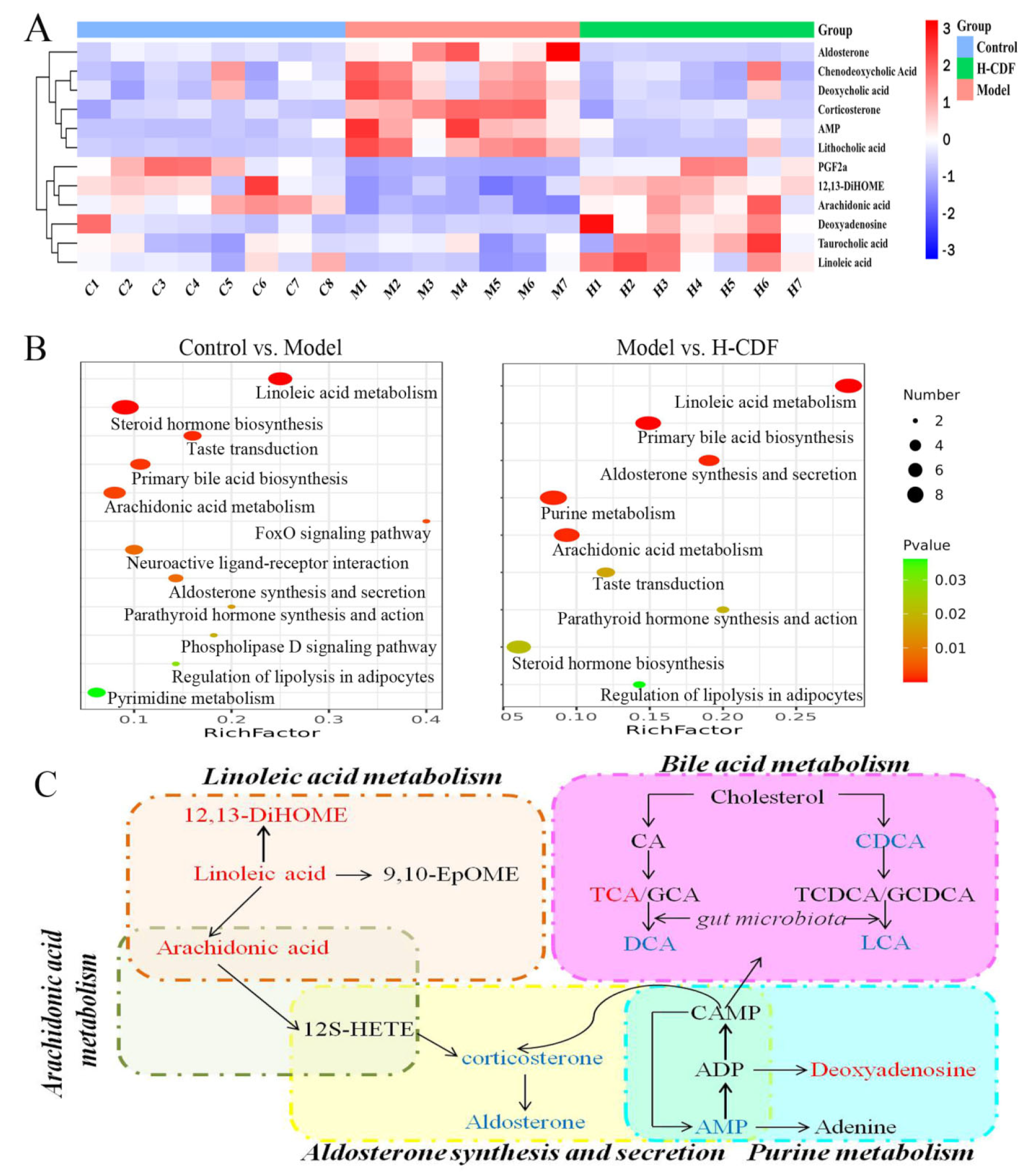
2.4. Identification of Potential Biomarkers and Analysis of Pathway Enrichment for Effect of H-CDF in Improving Insulin Resistance
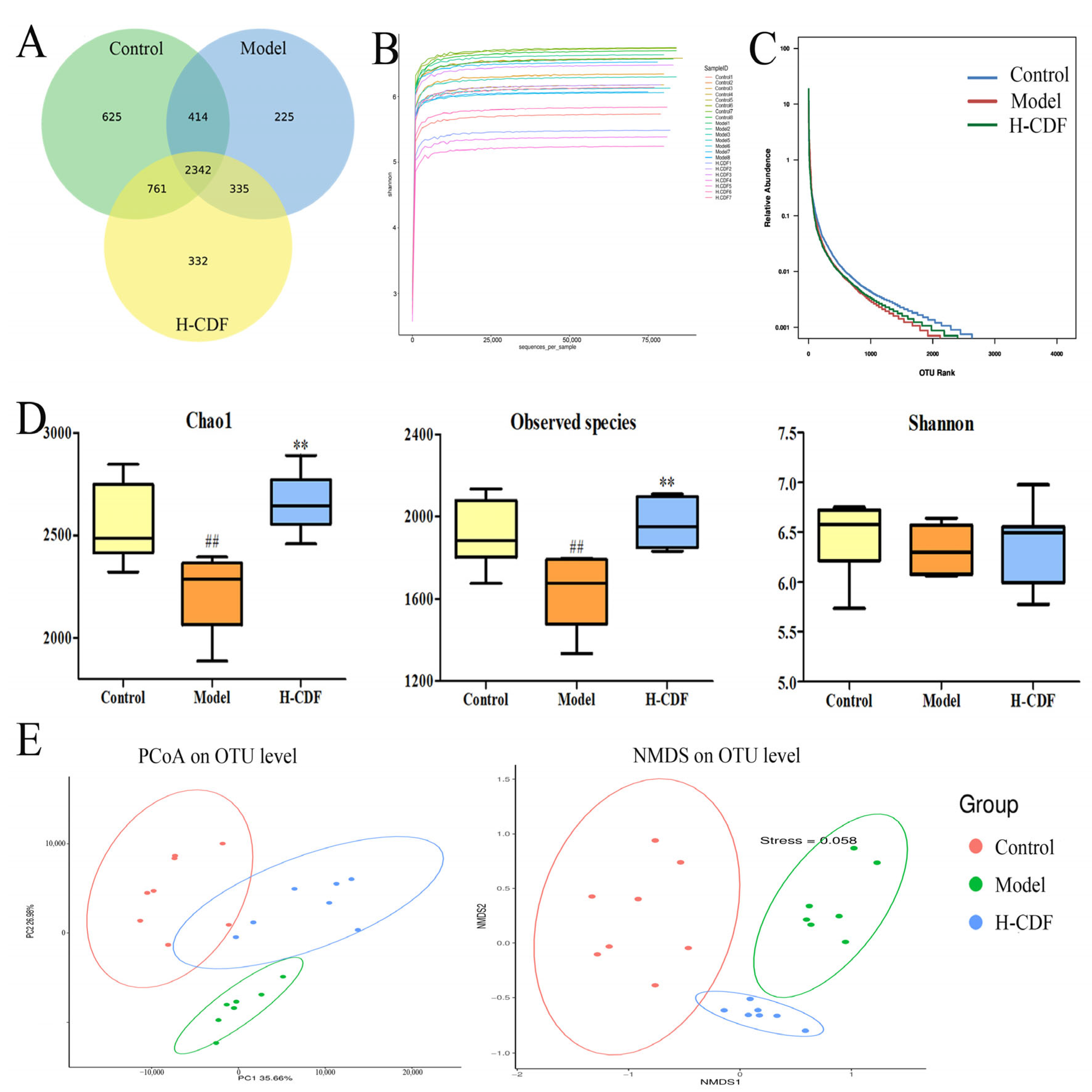
2.5. H-CDF Improves Bacterial Diversity in T2DM Rats
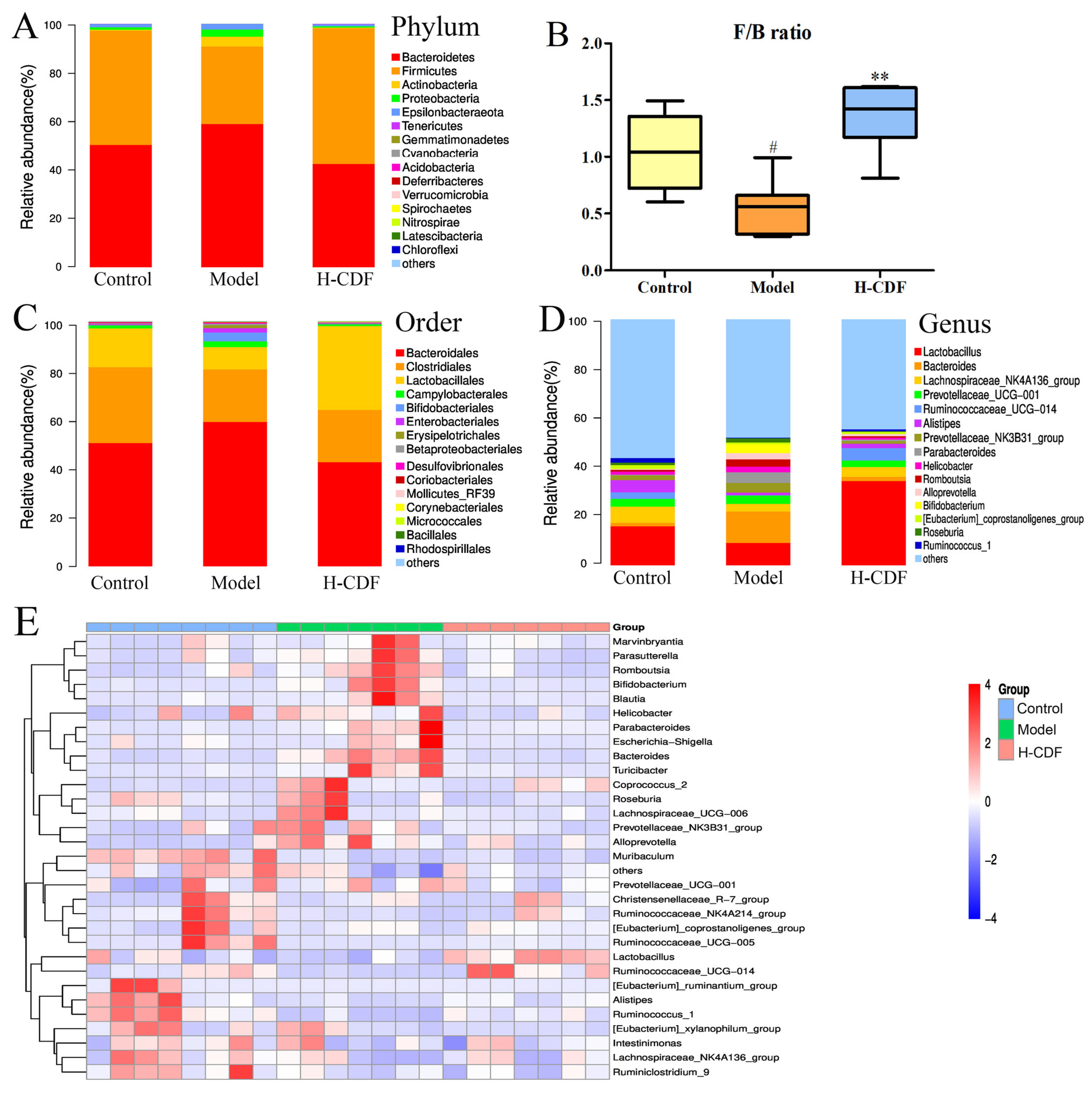
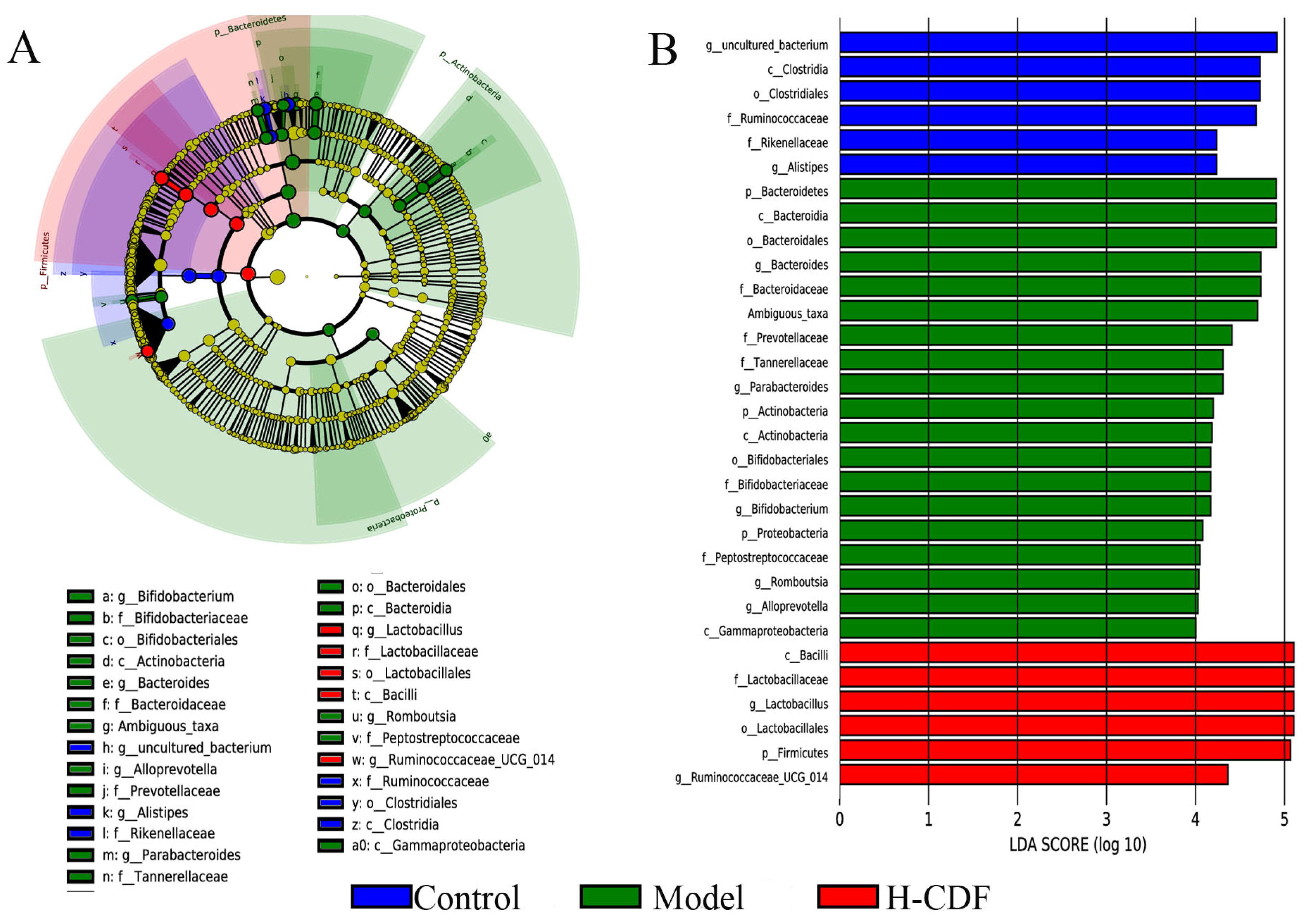
2.6. H-CDF Regulates the Structure of the Intestinal Flora at Multiple Taxonomic Levels
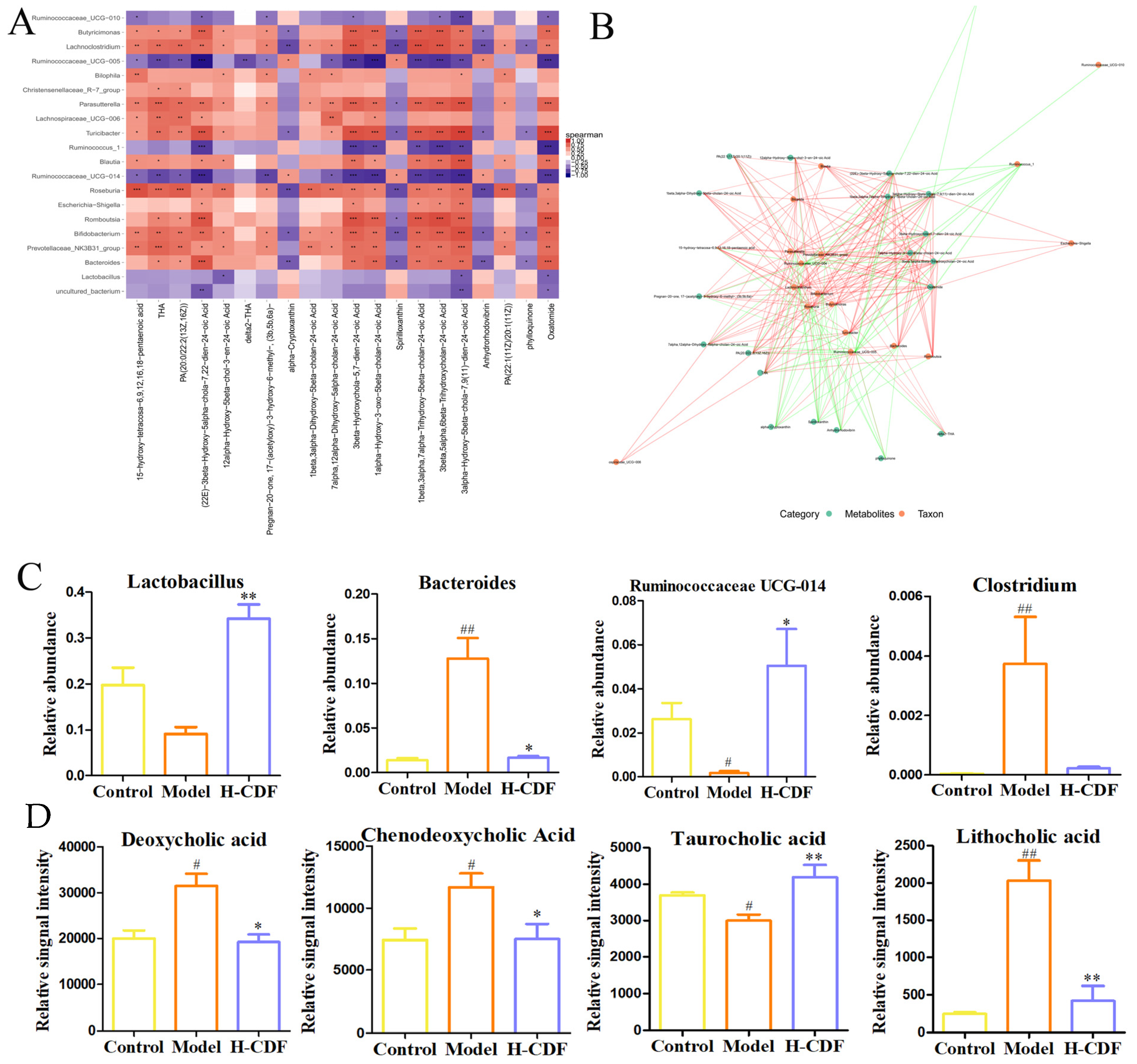
2.7. Spearman Correlation Analysis between Differential Flora and Metabolites in H-CFD Group
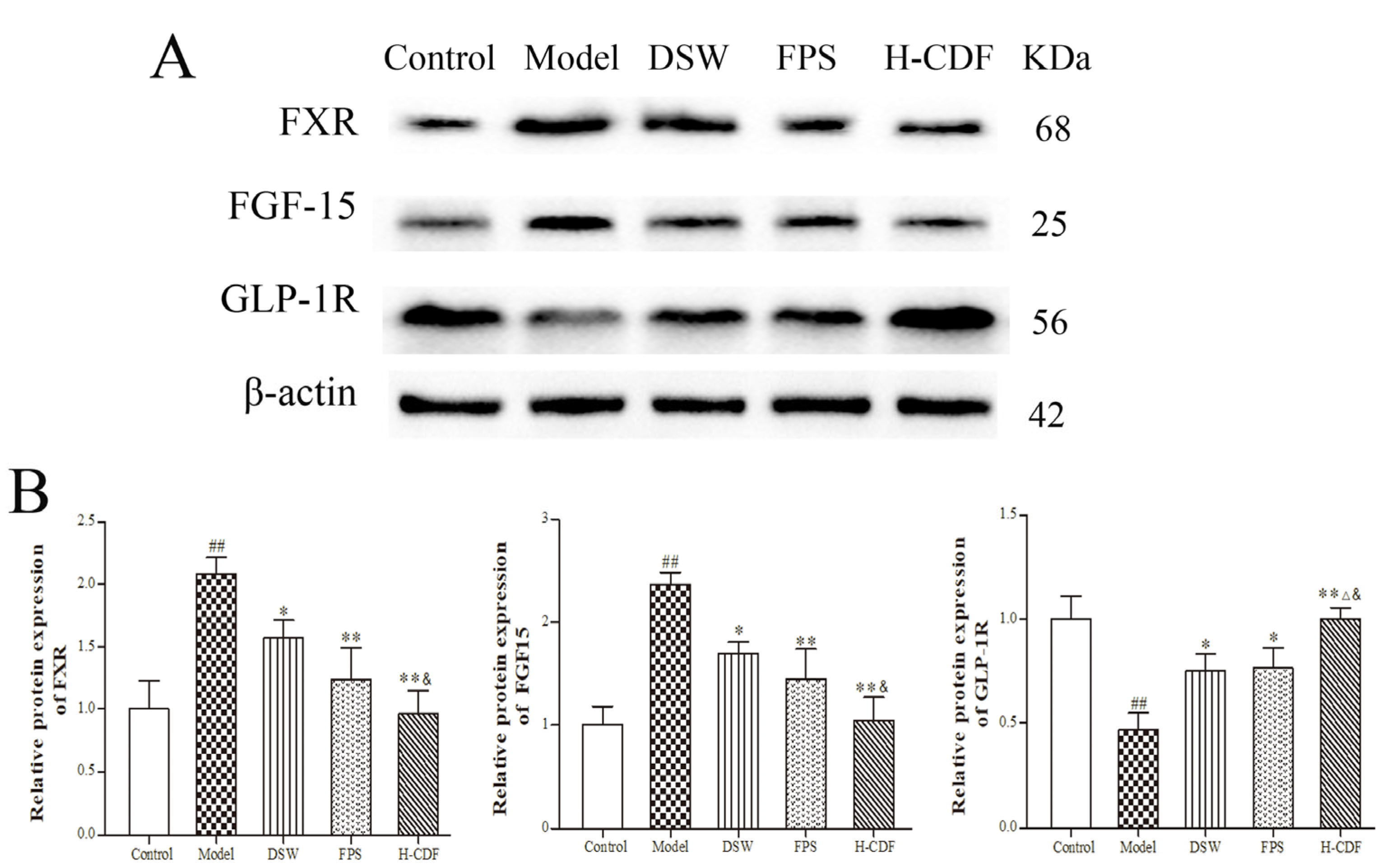
2.8. H-CDF Inhibits the FXR–FGF15 Pathway in Ileum Cells
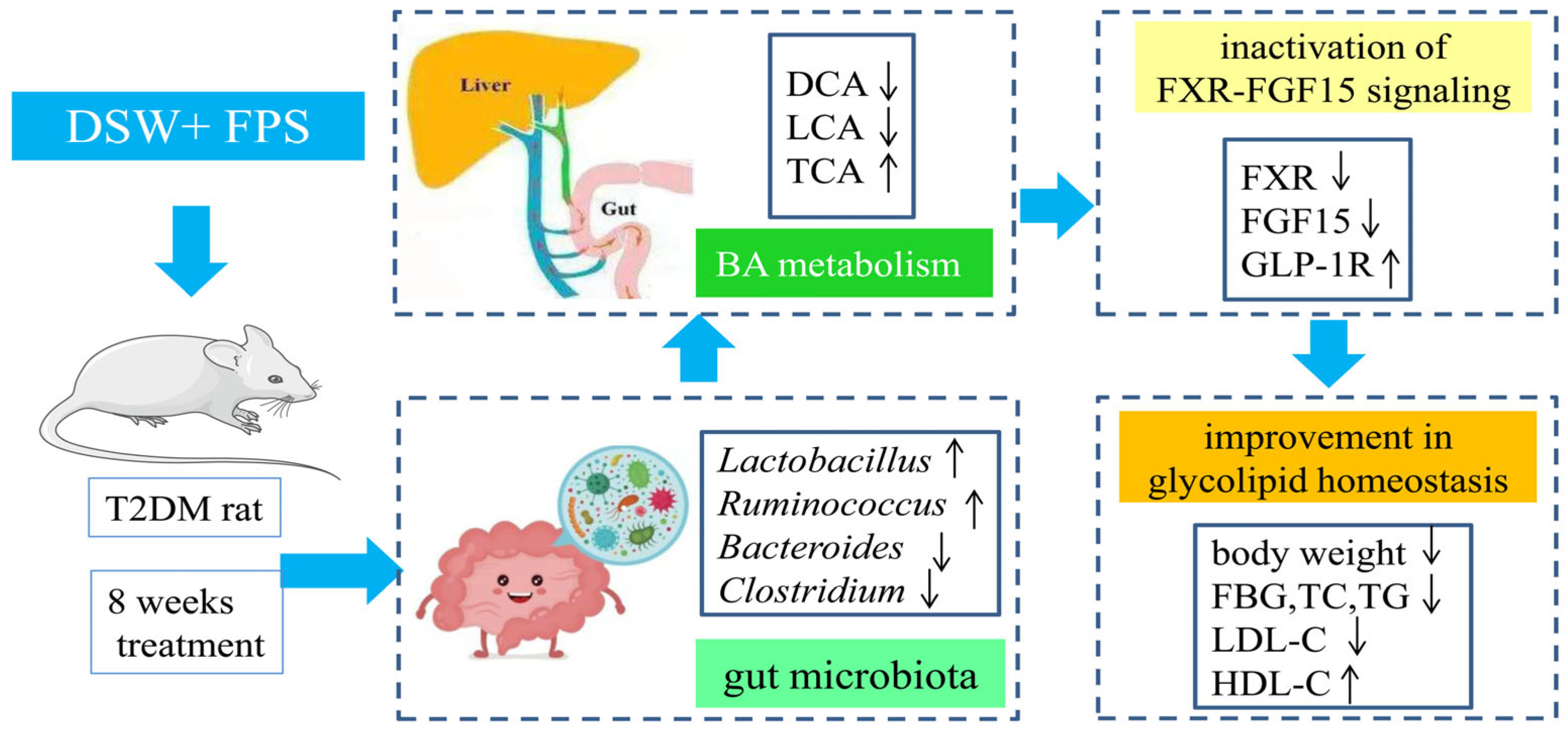
3. Discussion
4. Materials and Methods
4.1. Preparation and Elementary Analysis of DSW
4.2. Analysis of Fucoidan Physicochemical Properties
4.3. T2DM Model Induction and Drug Administration in Rats
4.4. Biomedical Analysis
4.5. Histological Analysis
4.6. Untargeted Metabolomics Analyses
4.7. Identification of Potential Biomarkers and Metabolic Pathways
4.8. 16S rDNA Gene Sequencing
4.9. Spearman Correlation Analysis
4.10. WB Detection
4.11. Statistical Analysis
5. Conclusions
6. Strengths and Limitations of This Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [Green Version]
- Cunha, L.; Grenha, A. Sulfated Seaweed Polysaccharides as Multifunctional Materials in Drug Delivery Applications. Mar. Drugs 2016, 14, 42. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Xu, X.; Wei, Q.; Yang, Q.; Zhao, J.; Wang, Y.; Li, X.; Ji, K.; Song, S. Application of fucoidan as treatment for cardio-vascular and cerebrovascular diseases. Ther. Adv. Chronic Dis. 2022, 13, 20406223221076891. [Google Scholar] [CrossRef]
- Zhang, X.; You, Y.; Wang, L.; Ai, C.; Huang, L.; Wang, S.; Wang, Z.; Song, S.; Zhu, B. Anti-obesity effects of Laminaria japonica fucoidan in high-fat diet-fed mice vary with the gut microbiota structure. Food Funct. 2022, 13, 6259–6270. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, S.; Hamada, A.; Cui, T.; Yokota, J.; Yamamoto, S.; Kusunose, M.; Miyamura, M.; Kyotani, S.; Kaneda, R.; Tsutsui, Y.; et al. Pharmacological Activity of Deep-Sea Water: Examination of Hyperlipemia Prevention and Medical Treatment Effect. Biol. Pharm. Bull. 2003, 26, 1552–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, S.; Liu, H.B.; Yang, X.; Li, C.X.; Guan, H.S. What depth should deep-sea water be pumped up from in the South China Sea for medicinal research? J. Ocean Univ. China 2013, 12, 134–138. [Google Scholar] [CrossRef]
- Shi, M.; Hu, B.; Hong, Y.; Wang, M.; Yao, Y. Deep Sea Water Inhibited Pancreatic β-Cell Apoptosis and Regulated Glucose Homeostasis by Affecting Lipid Metabolism in Db/Db Mice. Diabetes Metab. Syndr. Obes. 2023, 16, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Ham, J.Y.; Shon, Y.H. Natural Magnesium-Enriched Deep-Sea Water Improves Insulin Resistance and the Lipid Profile of Prediabetic Adults: A Randomized, Double-Blinded Crossover Trial. Nutrients 2020, 12, 515. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.J.; Joo, E.J. Effect of the supply of natural water from deep sea rock on the immune response and antioxidant activity in rats. J. Anim. Sci. 2006, 48, 211–215. [Google Scholar]
- Lee, K.-S.; Lee, M.-G.; Woo, Y.-J.; Nam, K.-S. The preventive effect of deep sea water on the development of cancerous skin cells through the induction of autophagic cell death in UVB-damaged HaCaT keratinocyte. Biomed. Pharmacother. 2019, 111, 282–291. [Google Scholar] [CrossRef]
- Chen, P.C.; Lee, Y.C.; Jao, H.Y.; Wang, C.P.; Jacobs, A.; Hu, K.; Chen, J.; Lo, C.S.; Lee, H.J. Supplementation of nanofiltrated deep ocean water ameliorate the progression of osteoporosis in ovariectomized rat via regulating osteoblast differentiation. J. Food Biochem. 2020, 44, e13236. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Yoshikane, Y.; Takenaka, H.; Kimura, A.; Islam, J.M.; Matsuda, R.; Okamoto, A.; Hashimoto, Y.; Yano, R.; Yamaguchi, K.; et al. Health Effects of Drinking Water Produced from Deep Sea Water: A Randomized Double-Blind Controlled Trial. Nutrients 2022, 14, 581. [Google Scholar] [CrossRef]
- Ha, B.G.; Jung, S.S.; Jang, Y.K.; Jeon, B.Y.; Shon, Y.H. Mineral-Enriched Deep-Sea Water Modulates Lactate Metabolism via PGC-1α-Mediated Metabolic Reprogramming. Mar. Drugs 2019, 17, 611. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Peng, W.-B.; Zhou, H.-L. Combination Treatment of Deep Sea Water and Fucoidan Attenuates High Glucose-Induced Insulin-Resistance in HepG2 Hepatocytes. Mar. Drugs 2018, 16, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, G.; Li, M.; Li, Y.; Mao, Y. Metabolomics: A New Tool to Reveal the Nature of Diabetic Kidney Disease. Lab. Med. 2022, 53, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Brunkwall, L.; Orho-Melander, M. The gut microbiome as a target for prevention and treatment of hyperglycaemia in type 2 diabetes: From current human evidence to future possibilities. Diabetologia 2017, 60, 943–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iatcu, C.O.; Steen, A.; Covasa, M. Gut Microbiota and Complications of Type-2 Diabetes. Nutrients 2021, 14, 166. [Google Scholar] [CrossRef]
- Zeng, S.-L.; Li, S.-Z.; Xiao, P.-T.; Cai, Y.-Y.; Chu, C.; Chen, B.-Z.; Li, P.; Li, J.; Liu, E.-H. Citrus polymethoxyflavones attenuate metabolic syndrome by regulating gut microbiome and amino acid metabolism. Sci. Adv. 2020, 6, eaax6208. [Google Scholar] [CrossRef] [Green Version]
- Bala, A.; Roy, S.; Das, D.; Marturi, V.; Mondal, C.; Patra, S.; Haldar, P.K.; Samajdar, G. Role of Glycogen synthase kinase-3 in the etiology of Type 2 Diabetes Mellitus: A review. Curr. Diabetes Rev. 2021, 18, e300721195147. [Google Scholar] [CrossRef]
- Xie, C.; Huang, W.; Young, R.; Jones, K.; Horowitz, M.; Rayner, C.; Wu, T. Role of Bile Acids in the Regulation of Food Intake, and Their Dysregulation in Metabolic Disease. Nutrients 2021, 13, 1104. [Google Scholar] [CrossRef]
- Winston, J.A.; Theriot, C.M. Diversification of host bile acids by members of the gut microbiota. Gut Microbes 2020, 11, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Wahlström, A.; Sayin, S.I.; Marschall, H.-U.; Bäckhed, F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab. 2016, 24, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Radjabzadeh, D.; Chen, L.; Kurilshikov, A.; Kavousi, M.; Ahmadizar, F.; Ikram, M.A.; Uitterlinden, A.G.; Zhernakova, A.; Fu, J.; et al. Association of Insulin Resistance and Type 2 Diabetes With Gut Microbial Diversity: A Microbiome-Wide Analysis From Population Studies. JAMA Netw. Open 2021, 4, e2118811. [Google Scholar] [CrossRef]
- Perino, A.; Schoonjans, K. Metabolic Messengers: Bile acids. Nat. Metab. 2022, 4, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zheng, X.; Ma, X.; Jiang, R.; Zhou, W.; Zhou, S.; Zhang, Y.; Lei, S.; Wang, S.; Kuang, J.; et al. Theabrownin from Pu-erh tea attenuates hypercholesterolemia via modulation of gut microbiota and bile acid metabolism. Nat. Commun. 2019, 10, 4971. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, J.S.; Klett, E.L. Linoleic acid and the regulation of glucose homeostasis: A review of the evidence. Prostaglandins Leukot. Essent. Fat. Acids 2021, 175, 102366. [Google Scholar] [CrossRef]
- Mousavi, S.M.; Jalilpiran, Y.; Karimi, E.; Aune, D.; Larijani, B.; Mozaffarian, D.; Willett, W.C.; Esmaillzadeh, A. Dietary Intake of Linoleic Acid, Its Concentrations, and the Risk of Type 2 Diabetes: A Systematic Review and Dose-Response Meta-analysis of Prospective Cohort Studies. Diabetes Care 2021, 44, 2173–2181. [Google Scholar] [CrossRef] [PubMed]
- Vasan, S.K.; Noordam, R.; Gowri, M.S.; Neville, M.J.; Karpe, F.; Christodoulides, C. The proposed systemic thermogenic me-tabolites succinate and 12;13-diHOME are inversely associated with adiposity and related metabolic traits: Evidence from a large human cross-sectional study. Diabetologia 2019, 62, 2079–2087. [Google Scholar] [CrossRef] [Green Version]
- McGlone, E.R.; Bloom, S.R. Bile acids and the metabolic syndrome. Ann. Clin. Biochem. 2019, 56, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Zaborska, K.E.; Lee, S.A.; Garribay, D.; Cha, E.; Cummings, B.P. Deoxycholic acid supplementation impairs glucose homeo-stasis in mice. PLoS ONE 2018, 13, e0200908. [Google Scholar] [CrossRef] [Green Version]
- Bothou, C.; Beuschlein, F.; Spyroglou, A. Links between aldosterone excess and metabolic complications: A comprehensive review. Diabetes Metab. 2020, 46, 1–7. [Google Scholar] [CrossRef]
- Sonnweber, T.; Pizzini, A.; Nairz, M.; Weiss, G.; Tancevski, I. Arachidonic Acid Metabolites in Cardiovascular and Metabolic Diseases. Int. J. Mol. Sci. 2018, 19, 3285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Sun, B.; Yu, D.; Zhu, C. Gut Microbiota: An Important Player in Type 2 Diabetes Mellitus. Front. Cell. Infect. Microbiol. 2022, 12, 834485. [Google Scholar] [CrossRef]
- Larsen, N.; Vogensen, F.K.; Van Den Berg, F.W.J.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut Microbiota in Human Adults with Type 2 Diabetes Differs from Non-Diabetic Adults. PLoS ONE 2010, 5, e9085. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, Q.-L.; Shen, J.-H.; Wang, K.; Liu, J. Magnesium lithospermate B improves the gut microbiome and bile acid metabolic profiles in a mouse model of diabetic nephropathy. Acta Pharmacol. Sin. 2019, 40, 507–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Lu, J.; Wang, Y.; Gu, W.; Yang, X.; Yu, J. Intake of total saponins and polysaccharides from Polygonatum kingianum affects the gut microbiota in diabetic rats. Phytomedicine 2017, 26, 45–54. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [Green Version]
- Kanazawa, A.; Aida, M.; Yoshida, Y.; Kaga, H.; Katahira, T.; Suzuki, L.; Tamaki, S.; Sato, J.; Goto, H.; Azuma, K.; et al. Effects of Synbiotic Supplementation on Chronic Inflammation and the Gut Microbiota in Obese Patients with Type 2 Diabetes Mellitus: A Randomized Controlled Study. Nutrients 2021, 13, 558. [Google Scholar] [CrossRef] [PubMed]
- Aydin, Ö.; Nieuwdorp, M.; Gerdes, V. The Gut Microbiome as a Target for the Treatment of Type 2 Diabetes. Curr. Diabetes Rep. 2018, 18, 55. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Li, N.; Shi, J.; Li, H.; Yue, Y.; Jiao, W.; Wang, N.; Song, Y.; Huo, G.; Li, B. Lactobacillus acidophilus alleviates type 2 diabetes by regulating hepatic glucose, lipid metabolism and gut microbiota in mice. Food Funct. 2019, 10, 5804–5815. [Google Scholar] [CrossRef]
- Falony, G.; Joossens, M.; Vieira-Silva, S.; Wang, J.; Darzi, Y.; Faust, K.; Kurilshikov, A.; Bonder, M.J.; Valles-Colomer, M.; Vandeputte, D.; et al. Population-level analysis of gut microbiome variation. Science 2016, 352, 560–564. [Google Scholar] [CrossRef]
- Zarrinpar, A.; Loomba, R. Review article: The emerging interplay among the gastrointestinal tract, bile acids and incretins in the pathogenesis of diabetes and non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2012, 36, 909–921. [Google Scholar] [CrossRef] [Green Version]
- Jia, W.; Xie, G.; Jia, W. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Chen, T.; Jiang, R.; Zhao, A.; Wu, Q.; Kuang, J.; Sun, D.; Ren, Z.; Li, M.; Zhao, M.; et al. Hyocholic acid species improve glucose homeostasis through a distinct TGR5 and FXR signaling mechanism. Cell Metab. 2021, 33, 791–803.e7. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Wang, X.; Li, J.; Zhang, Y.; Zhong, H.; Liu, R.; Zhang, D.; Feng, Q.; Xie, X.; Hong, J.; et al. Analyses of gut microbiota and plasma bile acids enable stratification of patients for antidiabetic treatment. Nat. Commun. 2017, 8, 1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.-Y.; Lim, S. Fucoidans and Bowel Health. Mar. Drugs 2021, 19, 436. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Xue, M.; Yang, J.; Pei, Z.; Zhang, N.; Qin, K.; Liang, H. Metabolic regulation mechanism of fucoidan via intestinal microecology in diseases. J. Sci. Food Agric. 2021, 101, 4456–4463. [Google Scholar] [CrossRef]
- Zhang, Y.; Zuo, J.; Yan, L.; Cheng, Y.; Li, Q.; Wu, S.; Chen, L.; Thring, R.W.; Yang, Y.; Gao, Y.; et al. Sargassum fusiforme Fucoidan Alleviates High-Fat Diet-Induced Obesity and Insulin Resistance Associated with the Improvement of Hepatic Oxidative Stress and Gut Microbiota Profile. J. Agric. Food Chem. 2020, 68, 10626–10638. [Google Scholar] [CrossRef]
- Wu, Q.; Wu, S.; Cheng, Y.; Zhang, Z.; Mao, G.; Li, S.; Yang, Y.; Zhang, X.; Wu, M.; Tong, H. Sargassum fusiforme fucoidan modifies gut microbiota and intestinal metabolites during alleviation of hyperglycemia in type 2 diabetic mice. Food Funct. 2021, 12, 3572–3585. [Google Scholar] [CrossRef]
- Lin, C.-H.; Chen, Y.-H.; Tsai, T.-Y.; Pan, T.-M. Effects of deep sea water and Lactobacillus paracasei subsp. paracasei NTU 101 on hypercholesterolemia hamsters gut microbiota. Appl. Microbiol. Biotechnol. 2017, 101, 321–329. [Google Scholar] [CrossRef]
- Takeuchi, H.; Higuchi, K.; Yoshikane, Y.; Takagi, R.; Tokuhiro, S.; Takenaka, K.; Oboshi, W.; Kimura, A.; Islam, J.; Kaneko, A.; et al. Drinking Refined Deep-Sea Water Improves the Gut Ecosystem with Beneficial Effects on Intestinal Health in Humans: A Randomized Double-Blind Controlled Trial. Nutrients 2020, 12, 2646. [Google Scholar] [CrossRef] [PubMed]
- Petrič, D.; Mravčáková, D.; Kucková, K.; Kišidayová, S.; Cieslak, A.; Szumacher-Strabel, M.; Huang, H.; Kolodziejski, P.; Lu-komska, A.; Slusarczyk, S.; et al. Impact of Zinc and/or Herbal Mixture on Ruminal Fermentation, Microbiota, and Histopathology in Lambs. Front. Vet. Sci. 2021, 8, 630971. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.L.U.; Sena-Evangelista, K.C.M.; de Azevedo, E.P.; Pinheiro, F.I.; Cobucci, R.N.; Pedrosa, L.F.C. Selenium in Human Health and Gut Microflora: Bioavailability of Selenocompounds and Relationship with Diseases. Front. Nutr. 2021, 8, 685317. [Google Scholar] [CrossRef]
- He, S.; Peng, W.-B.; Fu, X.-J.; Zhou, H.-L.; Wang, Z.-G. Deep Sea Water Alleviates Tau Phosphorylation and Cognitive Impairment via PI3K/Akt/GSK-3β Pathway. Mar. Biotechnol. 2022, 24, 68–81. [Google Scholar] [CrossRef]
- Nie, X.-R.; Li, H.-Y.; Du, G.; Lin, S.; Hu, R.; Li, H.-Y.; Zhao, L.; Zhang, Q.; Chen, H.; Wu, D.-T.; et al. Structural characteristics, rheological properties, and biological activities of polysaccharides from different cultivars of okra (Abelmoschus esculentus) collected in China. Int. J. Biol. Macromol. 2019, 139, 459–467. [Google Scholar] [CrossRef]
- Dai, B.; Wu, Q.; Zeng, C.; Zhang, J.; Cao, L.; Xiao, Z.; Yang, M. The effect of Liuwei Dihuang decoction on PI3K/Akt signaling pathway in liver of type 2 diabetes mellitus (T2DM) rats with insulin resistance. J. Ethnopharmacol. 2016, 192, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Qian, D.-W.; Jiang, S.; Shang, E.-X.; Zhu, Z.-H.; Duan, J.-A. Scutellariae Radix and Coptidis Rhizoma Improve Glucose and Lipid Metabolism in T2DM Rats via Regulation of the Metabolic Profiling and MAPK/PI3K/Akt Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 3634. [Google Scholar] [CrossRef] [Green Version]
- Liao, X.; Song, L.; Zeng, B.; Liu, B.; Qiu, Y.; Qu, H.; Zheng, Y.; Long, M.; Zhou, H.; Wang, Y.; et al. Alteration of gut microbiota induced by DPP-4i treatment improves glucose homeostasis. Ebiomedicine 2019, 44, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-M.; Chen, M.-J.; Zou, J.-F.; Jiang, S.; Shang, E.-X.; Qian, D.-W.; Duan, J.-A. UPLC-Q-TOF/MS based fecal metabolomics reveals the potential anti-diabetic effect of Xiexin Decoction on T2DM rats. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2021, 1173, 122683. [Google Scholar] [CrossRef]
- Chen, M.; Liao, Z.; Lu, B.; Wang, M.; Lin, L.; Zhang, S.; Li, Y.; Liu, D.; Liao, Q.; Xie, Z. Huang-Lian-Jie-Du-Decoction Ameliorates Hyperglycemia and Insulin Resistant in Association With Gut Microbiota Modulation. Front. Microbiol. 2018, 9, 2380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Li, Y.; Sun, M.; Xin, L.; Wang, T.; Wei, L.; Yu, C.; Liu, M.; Ni, Y.; Lu, R.; et al. The Chinese Herbal Formula Shenzhu Tiaopi Granule Results in Metabolic Improvement in Type 2 Diabetic Rats by Modulating the Gut Microbiota. Evid. Based Complement. Altern. Med. 2019, 2019, 6976394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Li, P.; Liu, S.; Liu, Q.; Li, Y.; Zhang, Z.; Yang, C.; Hu, M.; Sun, Y.; He, C.; et al. The chemopreventive effects of Huangqin-tea against AOM-induced preneoplastic colonic aberrant crypt foci in rats and omics analysis. Food Funct. 2020, 11, 9634–9650. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Drinking Water | Intragastric (ig) Administration | Dose/mg·kg−1 (ig) |
|---|---|---|---|
| Control | distilled water | distilled water | — |
| Model | distilled water | distilled water | — |
| Metf | distilled water | Metf | 150 |
| FPS | distilled water | fucoidan | 70 |
| DSW | DSW | distilled water | — |
| L-CDF | DSW | fucoidan | 17.5 |
| M-CDF | DSW | fucoidan | 35 |
| H-CDF | DSW | fucoidan | 70 |
| Metabolites | m/z | Rt (min) | VIP | Ion Mode | Formula | Change Trend | Fold-Change H/M | |
|---|---|---|---|---|---|---|---|---|
| M/C | H/M | |||||||
| PGF2a | 377.227 | 4.7717 | 1.68222 | pos | C20H34O5 | down | up | 4.87625 |
| Deoxyadenosine | 252.109 | 2.20938 | 2.36237 | pos | C10H13N5O3 | down | up | 3.72648 |
| Linoleic acid | 281.247 | 11.3704 | 5.63973 | pos | C18H32O2 | down | up | 2.65520 |
| Arachidonic acid | 327.227 | 7.29902 | 2.01829 | pos | C20H32O2 | down | up | 2.32247 |
| 12,13-DiHOME | 313.238 | 10.3238 | 2.95125 | neg | C18H34O4 | down | up | 1.903507 |
| Taurocholic acid | 516.301 | 7.08687 | 1.51054 | pos | C26H45NO7S | down | up | 1.27752 |
| Chenodeoxycholic Acid | 375.289 | 10.0277 | 3.27826 | pos | C24H40O4 | up | down | 0.64045 |
| Deoxycholic acid | 410.325 | 10.6713 | 6.28420 | pos | C24H40O4 | up | down | 0.61063 |
| Corticosterone | 385.175 | 4.55788 | 2.84140 | pos | C21H30O4 | up | down | 0.30670 |
| AMP | 348.07 | 1.19063 | 2.49771 | pos | C10H14N5O7P | up | down | 0.25528 |
| Lithocholic acid | 375.246 | 9.58685 | 2.37074 | neg | C24H40O3 | up | down | 0.20711 |
| Aldosterone | 359.189 | 8.7599 | 4.07483 | neg | C21H28O5 | up | down | 0.11069 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, S.; Peng, W.-B.; Zhou, H.-L.; Fu, X.-J.; Sun, Y.-H.; Wang, Z.-G. A Combination of Deep-Sea Water and Fucoidan Alleviates T2DM through Modulation of Gut Microbiota and Metabolic Pathways. Pharmaceuticals 2023, 16, 462. https://doi.org/10.3390/ph16030462
He S, Peng W-B, Zhou H-L, Fu X-J, Sun Y-H, Wang Z-G. A Combination of Deep-Sea Water and Fucoidan Alleviates T2DM through Modulation of Gut Microbiota and Metabolic Pathways. Pharmaceuticals. 2023; 16(3):462. https://doi.org/10.3390/ph16030462
Chicago/Turabian StyleHe, Shan, Wei-Bing Peng, Hong-Lei Zhou, Xian-Jun Fu, Yan-Hua Sun, and Zhen-Guo Wang. 2023. "A Combination of Deep-Sea Water and Fucoidan Alleviates T2DM through Modulation of Gut Microbiota and Metabolic Pathways" Pharmaceuticals 16, no. 3: 462. https://doi.org/10.3390/ph16030462