
Targeting the Endoplasmic Reticulum Stress-Linked PERK/GRP78/CHOP Pathway with Magnesium Sulfate Attenuates Chronic-Restraint-Stress-Induced Depression-like Neuropathology in Rats
 , , and
, , and 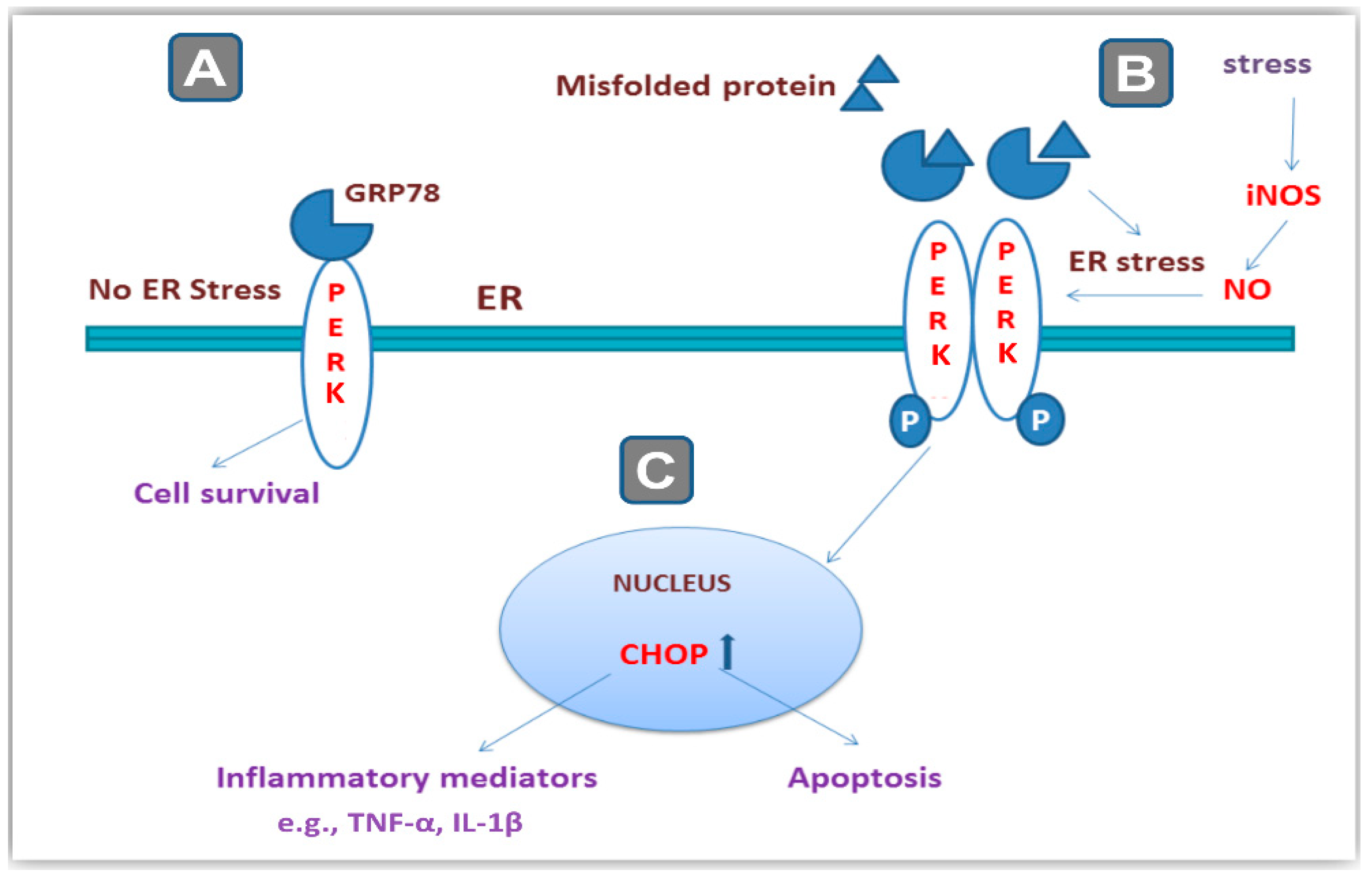{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
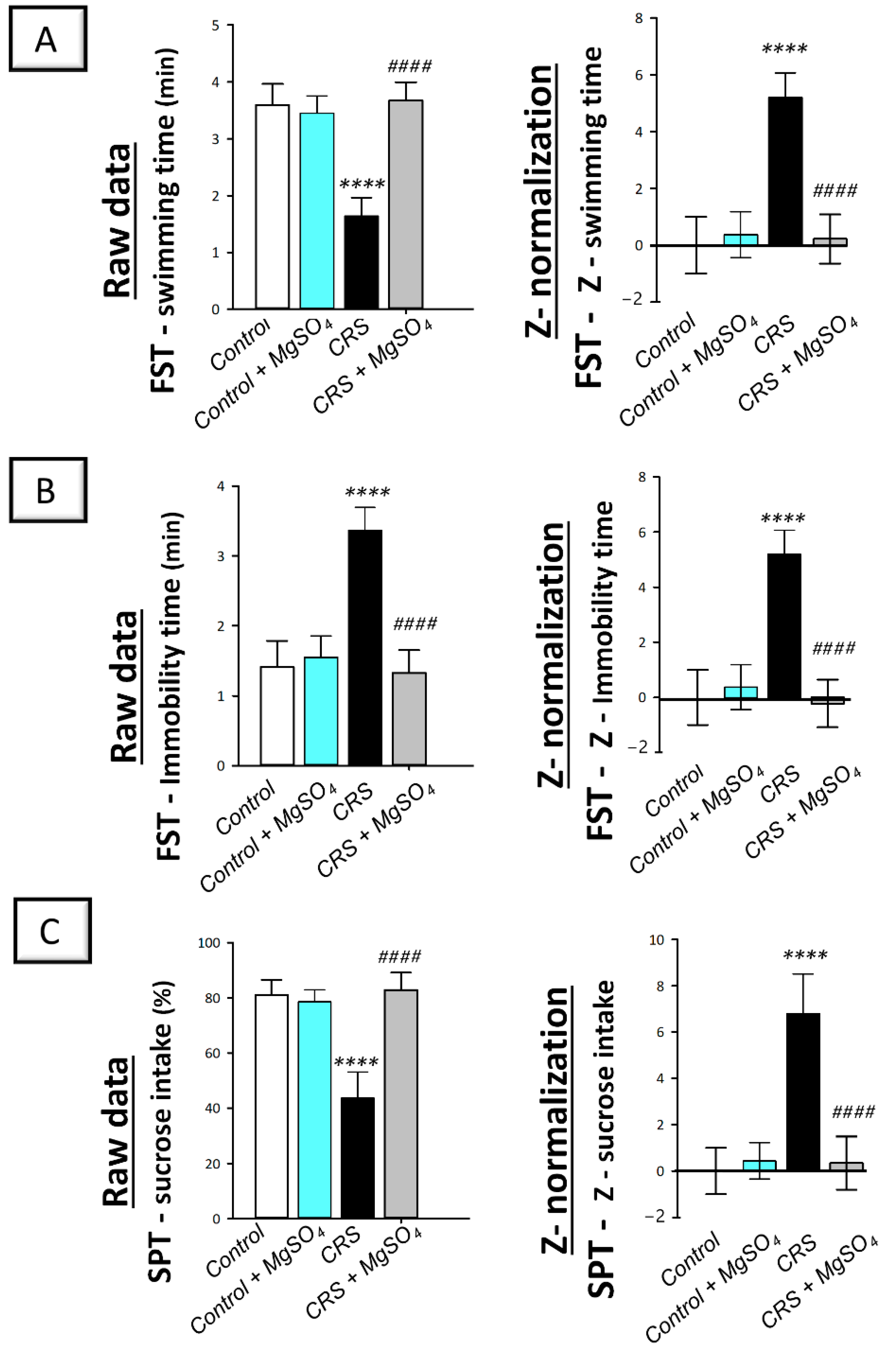
2.1. Chronic-Restraint-Stress (CRS)-Induced Depression-like Behavioral Deficits Were Mitigated by Magnesium Sulfate
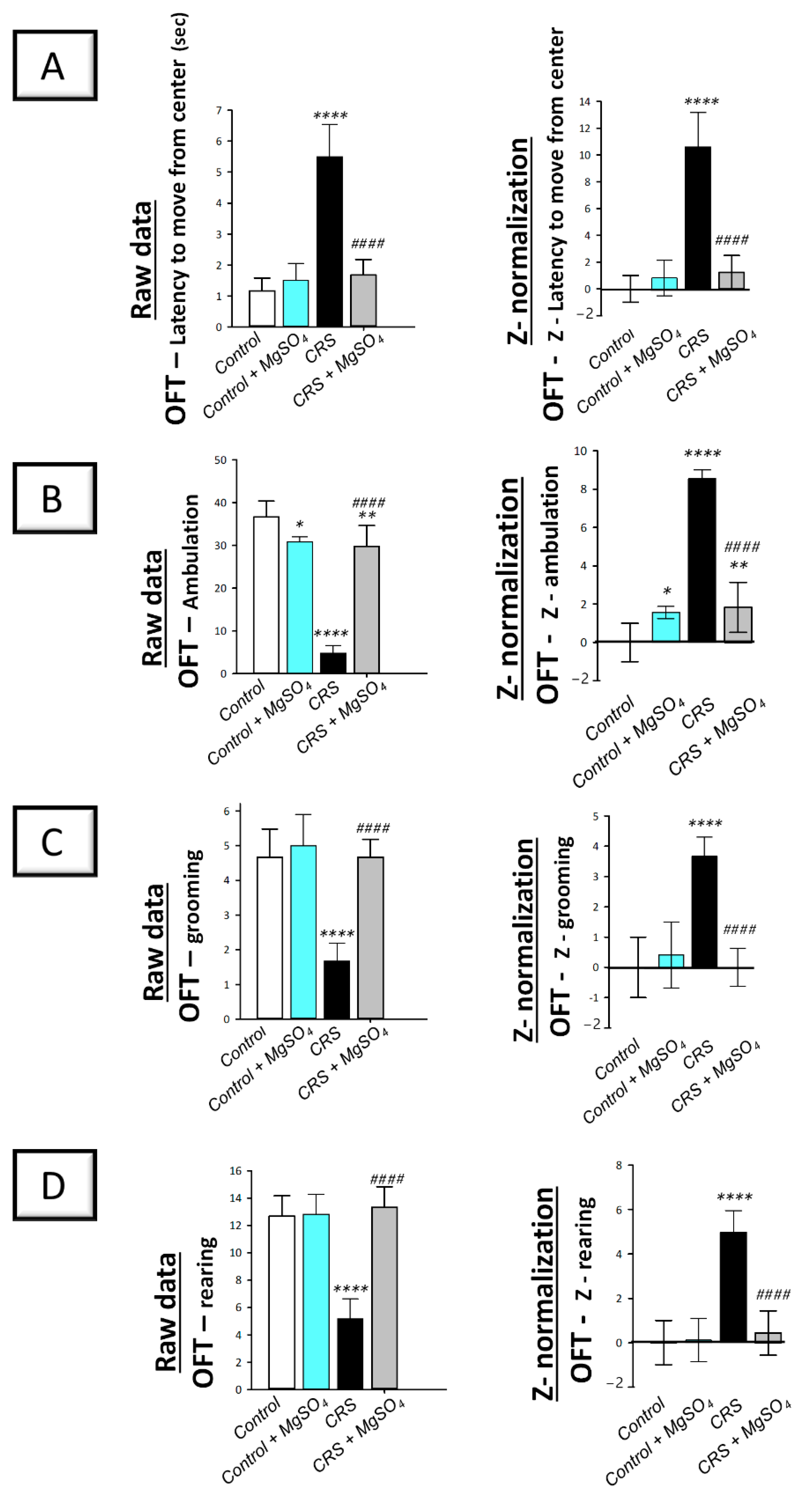
2.2. CRS-Induced Decline in the Locomotor Activity Was Counteracted by Magnesium Sulfate
2.3. CRS-Induced Spike in Serum Corticosterone Was Attenuated by Magnesium Sulfate
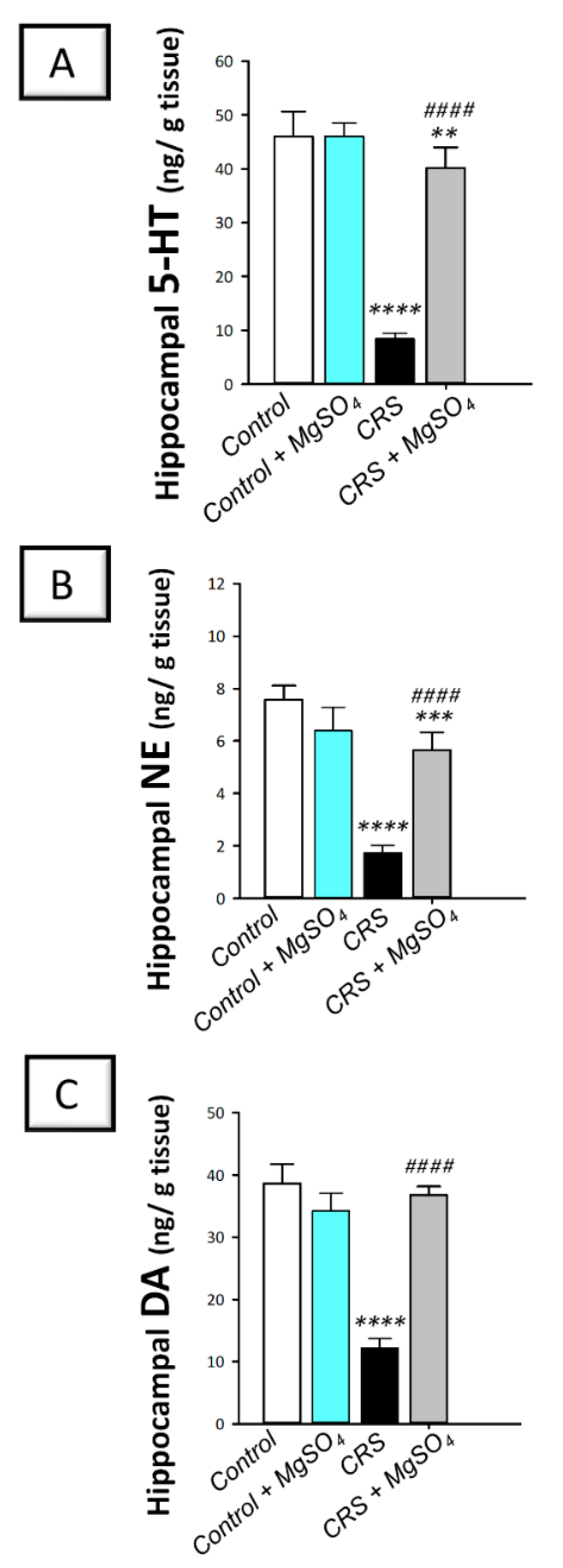
2.4. CRS-Induced Decline in Hippocampal Monoamine Neurotransmitters Was Counteracted by Magnesium Sulfate
2.5. CRS-Induced Increase in Hippocampal Pro-Inflammatory Cytokines Was Curbed by Magnesium Sulfate
2.6. CRS-Induced Increase in the Hippocampal Oxidative/Nitrosative Stress Markers Was Attenuated by Magnesium Sulfate
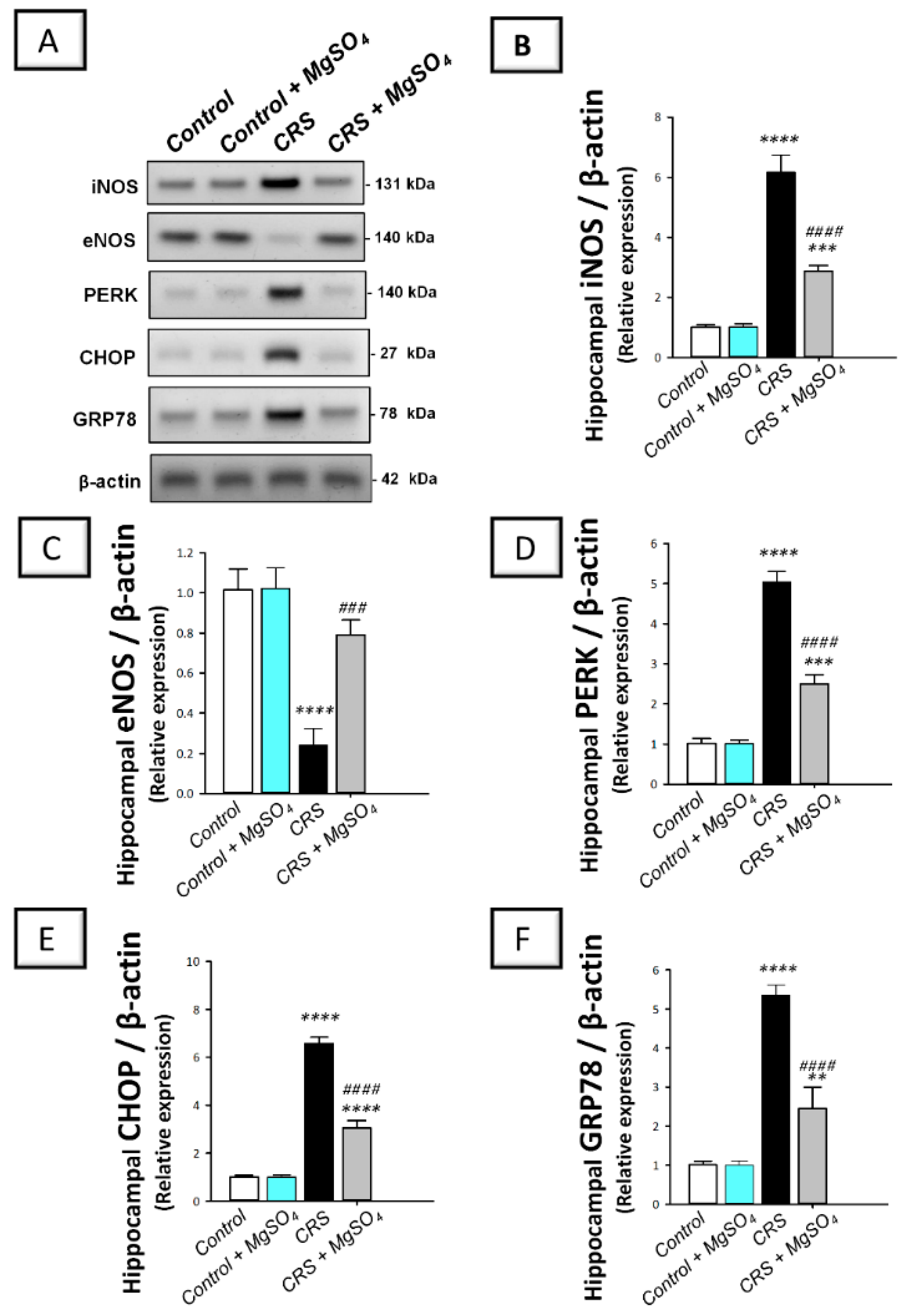
2.7. CRS-Induced Hippocampal Activation of PERK/GRP78/CHOP Pathway Was Curtailed by Magnesium Sulfate
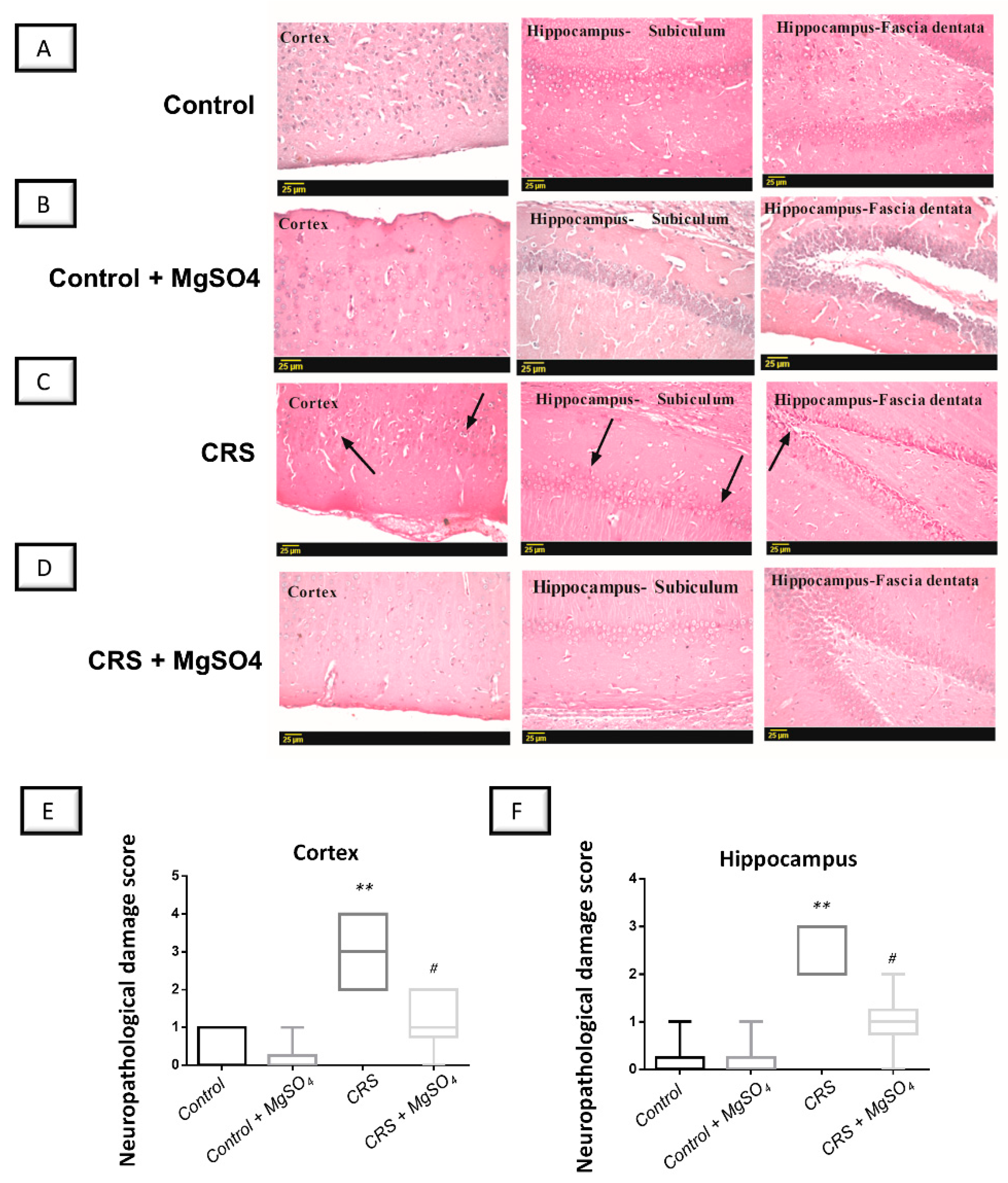
2.8. CRS-Induced Histopathological Changes in Brain Regions Were Attenuated by Magnesium Sulfate
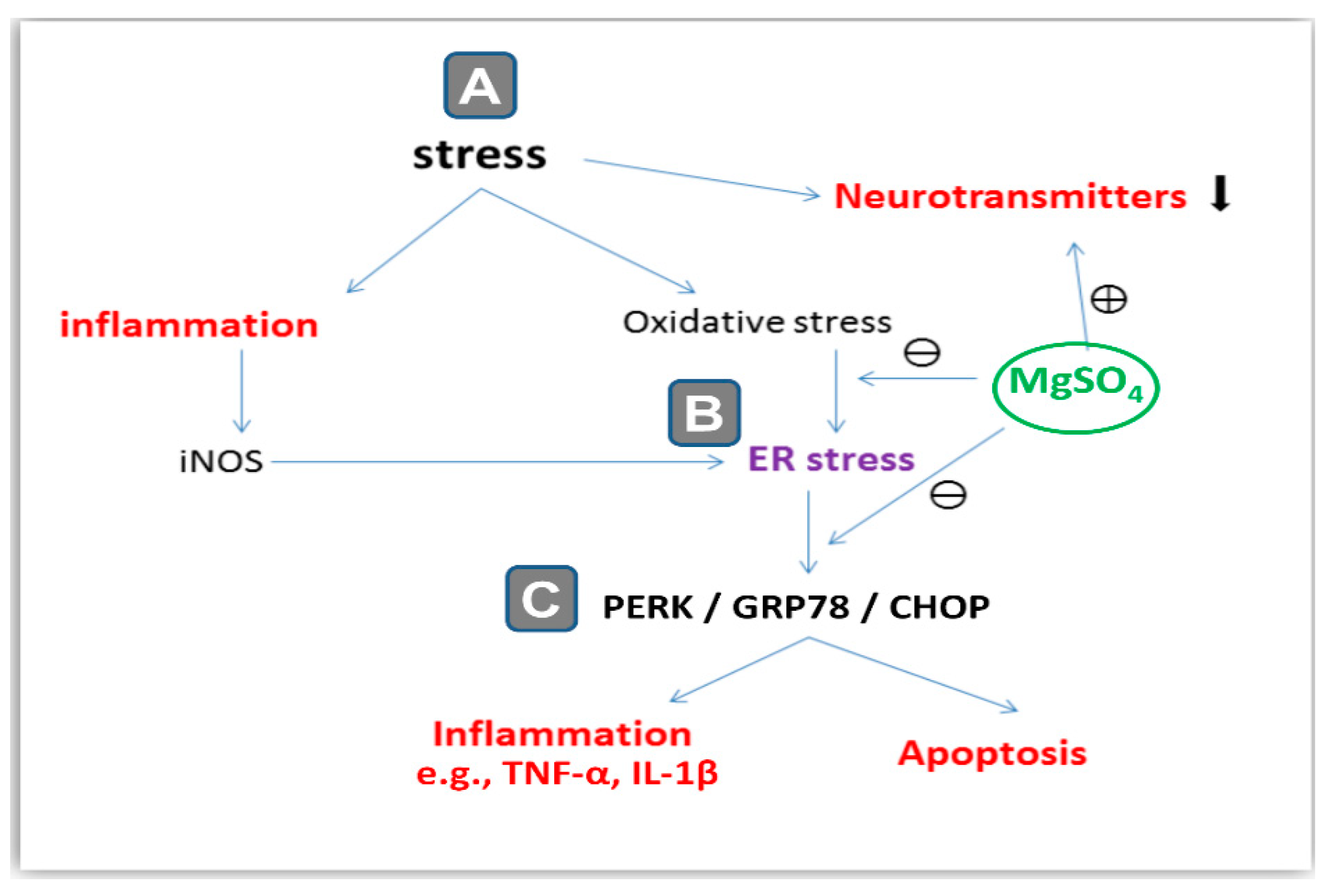
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals
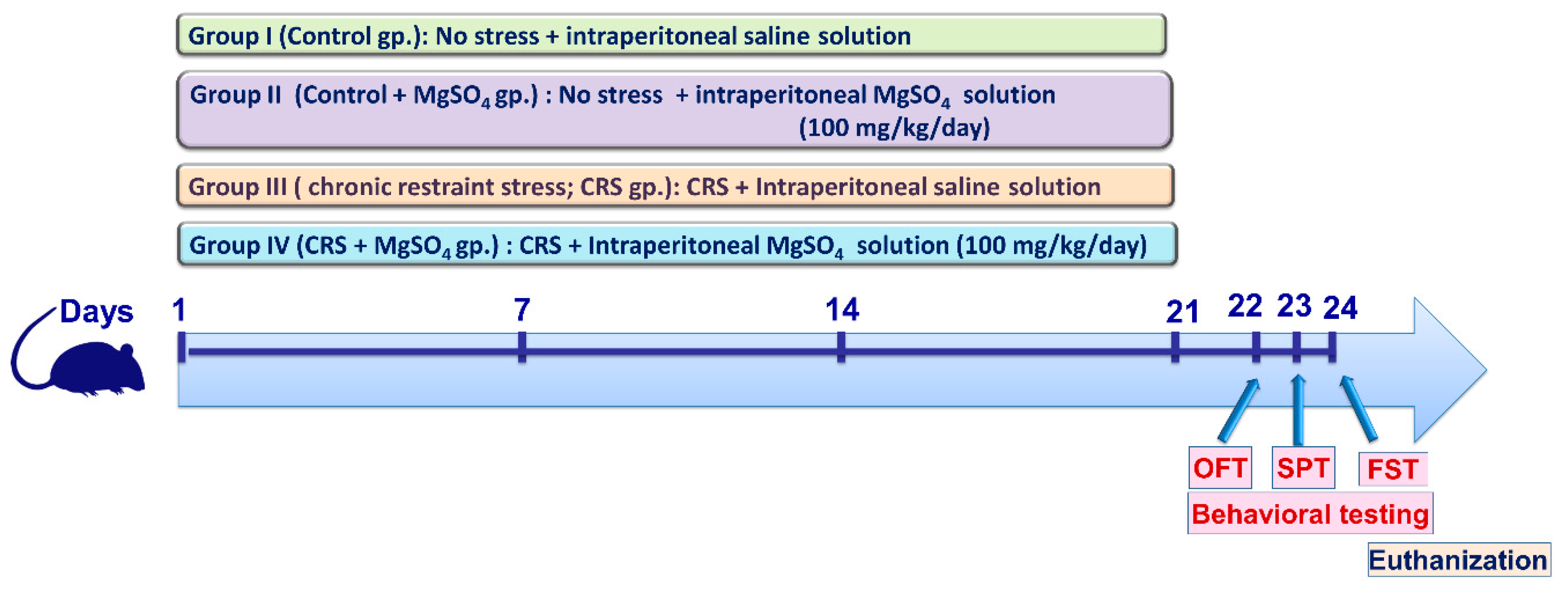
4.3. Animal Experiments
4.4. CRS Procedures
4.5. Behavioral Assessment
4.5.1. Open-Field Test
4.5.2. Sucrose Preference Test
4.5.3. Forced Swimming Test
4.5.4. Z-Score Normalization of Behavioral Data
4.6. Measurement of Hippocampal Neurotransmitters
4.7. Measurement of Serum Corticosterone
4.8. Measurement of Inflammatory Markers
4.9. Measurement of 8-OHdG
4.10. Western Blot Analysis of iNOS/ PERK/GRP78/CHOP Axis
4.11. Histopathological Examination and Neuropathological Damage Scoring
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benazzi, F. Various forms of depression. Dialogues Clin. Neurosci. 2022, 8, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.R.; Yoo, J.S.; Kim, Y.; Kang, H.; Lee, H.; Lm, S.J.; Choi, E.J.; Jung, M.A.; Bae, D.; Oh, K.N.; et al. Vaccinium bracteatum Leaf Extract Reverses Chronic Restraint Stress-Induced Depression-Like Behavior in Mice: Regulation of Hypothalamic-Pituitary-Adrenal Axis, Serotonin Turnover Systems, and ERK/Akt Phosphorylation. Front. Pharmacol. 2018, 9, 604. [Google Scholar] [CrossRef] [Green Version]
- Jangra, A.; Sriram, C.S.; Dwivedi, S.; Gurjar, S.S.; Hussain, M.I.; Borah, P.; Lahkar, M. Sodium Phenylbutyrate and Edaravone Abrogate Chronic Restraint Stress-Induced Behavioral Deficits: Implication of Oxido-Nitrosative, Endoplasmic Reticulum Stress Cascade, and Neuroinflammation. Cell. Mol. Neurobiol. 2017, 37, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Jangra, A.; Dwivedi, S.; Sriram, C.S.; Gurjar, S.S.; Kwatra, M.; Sulakhiya, K.; Baruah, C.C.; Lahkar, M. Honokiol abrogates chronic restraint stress-induced cognitive impairment and depressive-like behaviour by blocking endoplasmic reticulum stress in the hippocampus of mice. Eur. J. Pharmacol. 2016, 770, 25–32. [Google Scholar] [CrossRef]
- Wang, Y.-l.; Wu, H.-r.; Zhang, S.-s.; Xiao, H.-l.; Yu, J.; Ma, Y.-y.; Zhang, Y.-d.; Liu, Q. Catalpol ameliorates depressive-like behaviors in CUMS mice via oxidative stress-mediated NLRP3 inflammasome and neuroinflammation. Transl. Psychiatry 2021, 11, 353. [Google Scholar] [CrossRef]
- Saad, M.A.; Al-Shorbagy, M.Y.; Arab, H.H. Targeting the TLR4/NF-kappaBeta Axis and NLRP1/3 Inflammasomes by Rosuvastatin: A Role in Impeding Ovariectomy-Induced Cognitive Decline Neuropathology in Rats. Mol. Neurobiol. 2022, 59, 4562–4577. [Google Scholar] [CrossRef] [PubMed]
- Hamon, M.; Blier, P. Monoamine neurocircuitry in depression and strategies for new treatments. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 45, 54–63. [Google Scholar] [CrossRef]
- Plattner, F.; Hayashi, K.; Hernandez, A.; Benavides, D.R.; Tassin, T.C.; Tan, C.; Day, J.; Fina, M.W.; Yuen, E.Y.; Yan, Z.; et al. The role of ventral striatal cAMP signaling in stress-induced behaviors. Nat. Neurosci. 2015, 18, 1094–1100. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wey, S.; Zhang, Y.; Ye, R.; Lee, A.S. Role of the unfolded protein response regulator GRP78/BiP in development, cancer, and neurological disorders. Antioxid. Redox Signal. 2009, 11, 2307–2316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.; Li, L.; Chen, T.; Chen, X.; Tao, L.; Lin, X.; Tao, J.; Huang, X.; Jiang, J.; Liu, H.; et al. β-Arrestin-1 protects against endoplasmic reticulum stress/p53-upregulated modulator of apoptosis-mediated apoptosis via repressing p-p65/inducible nitric oxide synthase in portal hypertensive gastropathy. Free Radic. Biol. Med. 2015, 87, 69–83. [Google Scholar] [CrossRef]
- Schonthal, A.H. Endoplasmic reticulum stress: Its role in disease and novel prospects for therapy. Scientifica 2012, 2012, 857516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marciniak, S.J.; Yun, C.Y.; Oyadomari, S.; Novoa, I.; Zhang, Y.; Jungreis, R.; Nagata, K.; Harding, H.P.; Ron, D. CHOP induces death by promoting protein synthesis and oxidation in the stressed endoplasmic reticulum. Genes Dev. 2004, 18, 3066–3077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, K.; Posternak, M.; Jonathan, E.A. Toward achieving optimal response: Understanding and managing antidepressant side effects. Dialogues Clin. Neurosci. 2022, 10, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Hengartner, M.P. How effective are antidepressants for depression over the long term? A critical review of relapse prevention trials and the issue of withdrawal confounding. Ther. Adv. Psychopharmacol. 2020, 10, 2045125320921694. [Google Scholar] [CrossRef] [PubMed]
- Sravani, P.; Katasani, M.R.; Sharada, K. Comparative study of serum magnesium levels between low dose [mgso. sub. 4] and Pritchard regimen in treatment of eclampsia. Eur. J. Mol. Clin. Med. 2022, 9, 736–742. [Google Scholar]
- Li, X.; Han, X.; Yang, J.; Bao, J.; Di, X.; Zhang, G.; Liu, H. Magnesium Sulfate Provides Neuroprotection in Eclampsia-Like Seizure Model by Ameliorating Neuroinflammation and Brain Edema. Mol. Neurobiol. 2017, 54, 7938–7948. [Google Scholar] [CrossRef]
- Saver, J.L.; Starkman, S.; Eckstein, M.; Stratton, S.J.; Pratt, F.D.; Hamilton, S.; Conwit, R.; Liebeskind, D.S.; Sung, G.; Kramer, I. Prehospital use of magnesium sulfate as neuroprotection in acute stroke. N. Engl. J. Med. 2015, 372, 528–536. [Google Scholar] [CrossRef] [Green Version]
- Chollat, C.; Sentilhes, L.; Marret, S. Fetal Neuroprotection by Magnesium Sulfate: From Translational Research to Clinical Application. Front. Neurol. 2018, 9, 247. [Google Scholar] [CrossRef] [Green Version]
- Soleimanpour, H.; Imani, F.; Dolati, S.; Soleimanpour, M.; Shahsavarinia, K. Management of pain using magnesium sulphate: A narrative review. Postgrad. Med. 2022, 134, 260–266. [Google Scholar] [CrossRef]
- Guilloux, J.P.; Seney, M.; Edgar, N.; Sibille, E. Integrated behavioral z-scoring increases the sensitivity and reliability of behavioral phenotyping in mice: Relevance to emotionality and sex. J. Neurosci. Methods 2011, 197, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Gallo-Payet, N.; Battista, M.C. Steroidogenesis-adrenal cell signal transduction. Compr. Physiol. 2014, 4, 889–964. [Google Scholar] [CrossRef]
- Thoresen, M.; Bagenholm, R.; Loberg, E.M.; Apricena, F.; Kjellmer, I. Posthypoxic cooling of neonatal rats provides protection against brain injury. Arch. Dis. Child. Fetal Neonatal Ed. 1996, 74, F3–F9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhammad, R.N.; Ahmed, L.A.; Abdul Salam, R.M.; Ahmed, K.A.; Attia, A.S. Crosstalk Among NLRP3 Inflammasome, ET(B)R Signaling, and miRNAs in Stress-Induced Depression-Like Behavior: A Modulatory Role for SGLT2 Inhibitors. Neurotherapeutics 2021, 18, 2664–2681. [Google Scholar] [CrossRef]
- Gałecki, P.; Talarowska, M. Inflammatory theory of depression. Psychiatr. Pol. 2018, 52, 437–447. [Google Scholar] [CrossRef]
- Ismail, Z.; Gatchel, J.; Bateman, D.R.; Barcelos-Ferreira, R.; Cantillon, M.; Jaeger, J.; Donovan, N.J.; Mortby, M.E. Affective and emotional dysregulation as pre-dementia risk markers: Exploring the mild behavioral impairment symptoms of depression, anxiety, irritability, and euphoria. Int. Psychogeriatr. 2018, 30, 185–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Ce, Q.; Jin, L.; Zheng, J.; Sun, M.; Tang, X.; Li, D.; Sun, J. Deoiled sunflower seeds ameliorate depression by promoting the production of monoamine neurotransmitters and inhibiting oxidative stress. Food Funct. 2021, 12, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Marathe, S.V.; D’almeida, P.L.; Virmani, G.; Bathini, P.; Alberi, L. Effects of monoamines and antidepressants on astrocyte physiology: Implications for monoamine hypothesis of depression. J. Exp. Neurosci. 2018, 12, 1179069518789149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Jiang, M.; Gu, S.; Wang, F.; Yuan, B. Early Life Stress Induced DNA Methylation of Monoamine Oxidases Leads to Depressive-Like Behavior. Front. Cell Dev. Biol. 2020, 8, 582247. [Google Scholar] [CrossRef] [PubMed]
- Kvetnanský, R.; Fukuhara, K.; Pacak, K.; Cizza, G.; Goldstein, D.a.; Kopin, I. Endogenous glucocorticoids restrain catecholamine synthesis and release at rest and during immobilization stress in rats. Endocrinology 1993, 133, 1411–1419. [Google Scholar] [CrossRef] [PubMed]
- Brvar, M.; Chan, M.Y.; Dawson, A.H.; Ribchester, R.R.; Eddleston, M. Magnesium sulfate and calcium channel blocking drugs as antidotes for acute organophosphorus insecticide poisoning–a systematic review and meta-analysis. Clin. Toxicol. 2018, 56, 725–736. [Google Scholar] [CrossRef] [Green Version]
- Teng, T.; Shively, C.A.; Li, X.; Jiang, X.; Neigh, G.N.; Yin, B.; Zhang, Y.; Fan, L.; Xiang, Y.; Wang, M. Chronic unpredictable mild stress produces depressive-like behavior, hypercortisolemia, and metabolic dysfunction in adolescent cynomolgus monkeys. Transl. Psychiatry 2021, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-M.; Pei, L.-X.; Zhang, Y.-Y.; Cheng, Y.-X.; Niu, C.-L.; Cui, Y.; Feng, W.-S.; Wang, G.-F. Ethanol extract of Rehmannia glutinosa exerts antidepressant-like effects on a rat chronic unpredictable mild stress model by involving monoamines and BDNF. Metab. Brain Dis. 2018, 33, 885–892. [Google Scholar] [CrossRef]
- Kim, I.-B.; Lee, J.-H.; Park, S.-C. The relationship between stress, inflammation, and depression. Biomedicines 2022, 10, 1929. [Google Scholar] [CrossRef]
- Wang, P.; Yu, X.; Guan, P.P.; Guo, J.W.; Wang, Y.; Zhang, Y.; Zhao, H.; Wang, Z.Y. Magnesium ion influx reduces neuroinflammation in Abeta precursor protein/Presenilin 1 transgenic mice by suppressing the expression of interleukin-1beta. Cell. Mol. Immunol. 2017, 14, 451–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burd, I.; Breen, K.; Friedman, A.; Chai, J.; Elovitz, M.A. Magnesium sulfate reduces inflammation-associated brain injury in fetal mice. Am. J. Obstet. Gynecol. 2010, 202, 292.e1–292.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daher, I.; Le Dieu-Lugon, B.; Lecointre, M.; Dupre, N.; Voisin, C.; Leroux, P.; Dourmap, N.; Gonzalez, B.J.; Marret, S.; Leroux-Nicollet, I.; et al. Time- and sex-dependent efficacy of magnesium sulfate to prevent behavioral impairments and cerebral damage in a mouse model of cerebral palsy. Neurobiol. Dis. 2018, 120, 151–164. [Google Scholar] [CrossRef]
- Koning, G.; Leverin, A.L.; Nair, S.; Schwendimann, L.; Ek, J.; Carlsson, Y.; Gressens, P.; Thornton, C.; Wang, X.; Mallard, C.; et al. Magnesium induces preconditioning of the neonatal brain via profound mitochondrial protection. J. Cereb. Blood Flow Metab. 2019, 39, 1038–1055. [Google Scholar] [CrossRef] [Green Version]
- Khalilzadeh, M.; Abdollahi, A.; Abdolahi, F.; Abdolghaffari, A.H.; Dehpour, A.R.; Jazaeri, F. Protective effects of magnesium sulfate against doxorubicin induced cardiotoxicity in rats. Life Sci. 2018, 207, 436–441. [Google Scholar] [CrossRef]
- Nishitoh, H. CHOP is a multifunctional transcription factor in the ER stress response. J. Biochem. 2012, 151, 217–219. [Google Scholar] [CrossRef] [Green Version]
- Nosyreva, E.; Kavalali, E.T. Activity-dependent augmentation of spontaneous neurotransmission during endoplasmic reticulum stress. J. Neurosci. 2010, 30, 7358–7368. [Google Scholar] [CrossRef] [Green Version]
- Aalling, N.; Hageman, I.; Miskowiak, K.; Orlowski, D.; Wegener, G.; Wortwein, G. Erythropoietin prevents the effect of chronic restraint stress on the number of hippocampal CA3c dendritic terminals-relation to expression of genes involved in synaptic plasticity, angiogenesis, inflammation, and oxidative stress in male rats. J. Neurosci. Res. 2018, 96, 103–116. [Google Scholar] [CrossRef]
- Hageman, I.; Nielsen, M.; Wortwein, G.; Diemer, N.H.; Jorgensen, M.B. Electroconvulsive stimulations prevent stress-induced morphological changes in the hippocampus. Stress 2008, 11, 282–289. [Google Scholar] [CrossRef]
- Kao, M.-C.; Jan, W.-C.; Tsai, P.-S.; Wang, T.-Y.; Huang, C.-J. Magnesium sulfate mitigates lung injury induced by bilateral lower limb ischemia-reperfusion in rats. J. Surg. Res. 2011, 171, e97–e106. [Google Scholar] [CrossRef] [PubMed]
- Pryde, P.G.; Mittendorf, R. Contemporary usage of obstetric magnesium sulfate: Indication, contraindication, and relevance of dose. Obstet. Gynecol. 2009, 114, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Arab, H.H.; Abd El Aal, H.A.; Alsufyani, S.E.; El-Sheikh, A.A.K.; Arafa, E.A.; Ashour, A.M.; Kabel, A.M.; Eid, A.H. Topiramate Reprofiling for the Attenuation of Cadmium-Induced Testicular Impairment in Rats: Role of NLRP3 Inflammasome and AMPK/mTOR-Linked Autophagy. Pharmaceuticals 2022, 15, 1402. [Google Scholar] [CrossRef] [PubMed]
- Safar, M.M.; Arab, H.H.; Rizk, S.M.; El-Maraghy, S.A. Bone Marrow-Derived Endothelial Progenitor Cells Protect Against Scopolamine-Induced Alzheimer-Like Pathological Aberrations. Mol. Neurobiol. 2016, 53, 1403–1418. [Google Scholar] [CrossRef] [PubMed]
- Arab, H.H.; Safar, M.M.; Shahin, N.N. Targeting ROS-Dependent AKT/GSK-3beta/NF-kappaB and DJ-1/Nrf2 Pathways by Dapagliflozin Attenuates Neuronal Injury and Motor Dysfunction in Rotenone-Induced Parkinson’s Disease Rat Model. ACS Chem. Neurosci. 2021, 12, 689–703. [Google Scholar] [CrossRef] [PubMed]
- Scheggi, S.; De Montis, M.G.; Gambarana, C. Making Sense of Rodent Models of Anhedonia. Int. J. Neuropsychopharmacol. 2018, 21, 1049–1065. [Google Scholar] [CrossRef]
- Lu, X.; Yang, R.R.; Zhang, J.L.; Wang, P.; Gong, Y.; Hu, W.F.; Wu, Y.; Gao, M.H.; Huang, C. Tauroursodeoxycholic acid produces antidepressant-like effects in a chronic unpredictable stress model of depression via attenuation of neuroinflammation, oxido-nitrosative stress, and endoplasmic reticulum stress. Fundam. Clin. Pharmacol. 2018, 32, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Porsolt, R.D.; Bertin, A.; Jalfre, M. Behavioral despair in mice: A primary screening test for antidepressants. Arch. Int. Pharmacodyn. Ther. 1977, 229, 327–336. [Google Scholar]
- Belovicova, K.; Bogi, E.; Csatlosova, K.; Dubovicky, M. Animal tests for anxiety-like and depression-like behavior in rats. Interdiscip. Toxicol. 2017, 10, 40–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nassar, N.N.; Al-Shorbagy, M.Y.; Arab, H.H.; Abdallah, D.M. Saxagliptin: A novel antiparkinsonian approach. Neuropharmacology 2015, 89, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Arab, H.H.; Abd El-Aal, S.A.; Ashour, A.M.; El-Sheikh, A.A.K.; Al Khabbaz, H.J.; Arafa, E.A.; Mahmoud, A.M.; Kabel, A.M. Targeting inflammation and redox perturbations by lisinopril mitigates Freund’s adjuvant-induced arthritis in rats: Role of JAK-2/STAT-3/RANKL axis, MMPs, and VEGF. Inflammopharmacology 2022, 30, 1909–1926. [Google Scholar] [CrossRef]
- Kamel, K.M.; Gad, A.M.; Mansour, S.M.; Safar, M.M.; Fawzy, H.M. Venlafaxine alleviates complete Freund’s adjuvant-induced arthritis in rats: Modulation of STAT-3/IL-17/RANKL axis. Life Sci. 2019, 226, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Arab, H.H.; Eid, A.H.; El-Sheikh, A.A.K.; Arafa, E.A.; Ashour, A.M. Irbesartan reprofiling for the amelioration of ethanol-induced gastric mucosal injury in rats: Role of inflammation, apoptosis, and autophagy. Life Sci. 2022, 308, 120939. [Google Scholar] [CrossRef] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arab, H.H.; Khames, A.; Alsufyani, S.E.; El-Sheikh, A.A.K.; Gad, A.M. Targeting the Endoplasmic Reticulum Stress-Linked PERK/GRP78/CHOP Pathway with Magnesium Sulfate Attenuates Chronic-Restraint-Stress-Induced Depression-like Neuropathology in Rats. Pharmaceuticals 2023, 16, 300. https://doi.org/10.3390/ph16020300
Arab HH, Khames A, Alsufyani SE, El-Sheikh AAK, Gad AM. Targeting the Endoplasmic Reticulum Stress-Linked PERK/GRP78/CHOP Pathway with Magnesium Sulfate Attenuates Chronic-Restraint-Stress-Induced Depression-like Neuropathology in Rats. Pharmaceuticals. 2023; 16(2):300. https://doi.org/10.3390/ph16020300
Chicago/Turabian StyleArab, Hany H., Ali Khames, Shuruq E. Alsufyani, Azza A. K. El-Sheikh, and Amany M. Gad. 2023. "Targeting the Endoplasmic Reticulum Stress-Linked PERK/GRP78/CHOP Pathway with Magnesium Sulfate Attenuates Chronic-Restraint-Stress-Induced Depression-like Neuropathology in Rats" Pharmaceuticals 16, no. 2: 300. https://doi.org/10.3390/ph16020300