
The Antidepressant-like Activity and Cognitive Enhancing Effects of the Combined Administration of (R)-Ketamine and LY341495 in the CUMS Model of Depression in Mice Are Related to the Modulation of Excitatory Synaptic Transmission and LTP in the PFC

Abstract
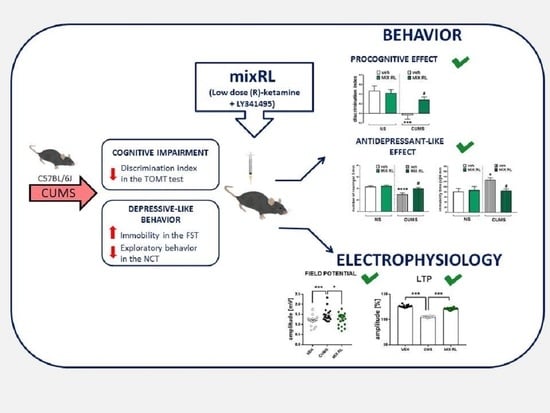
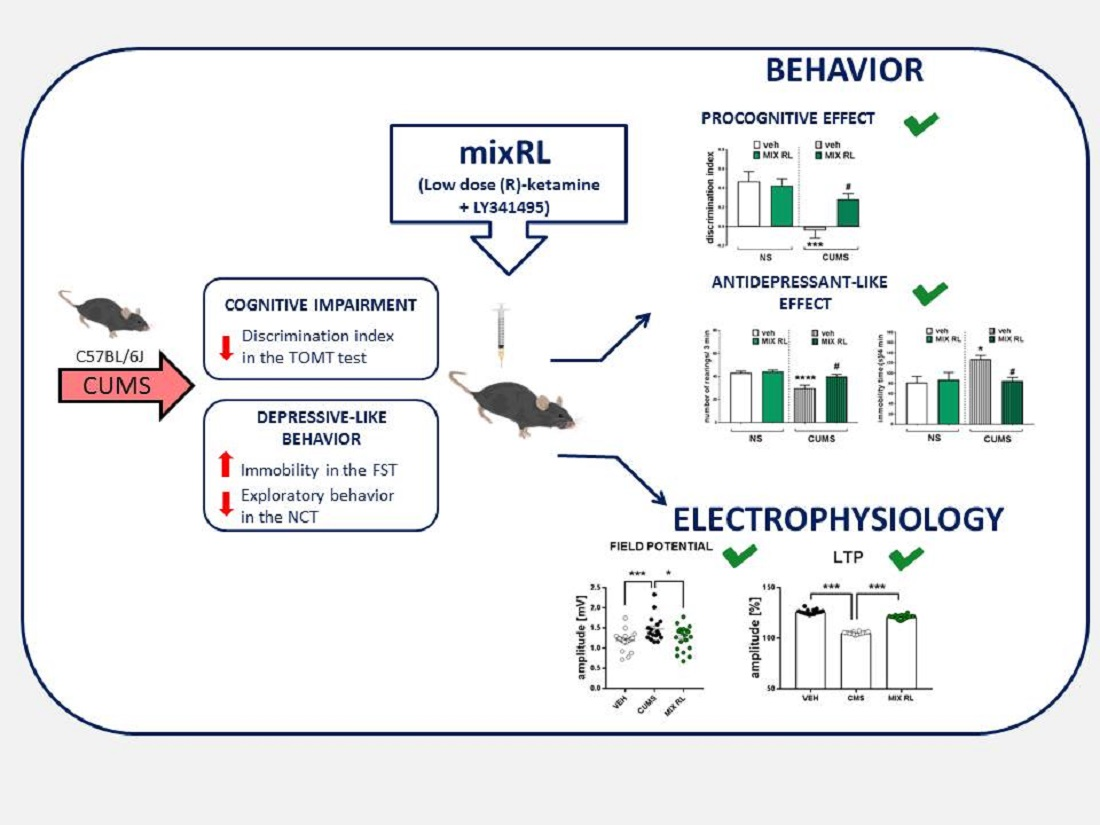
:
1. Introduction
2. Results
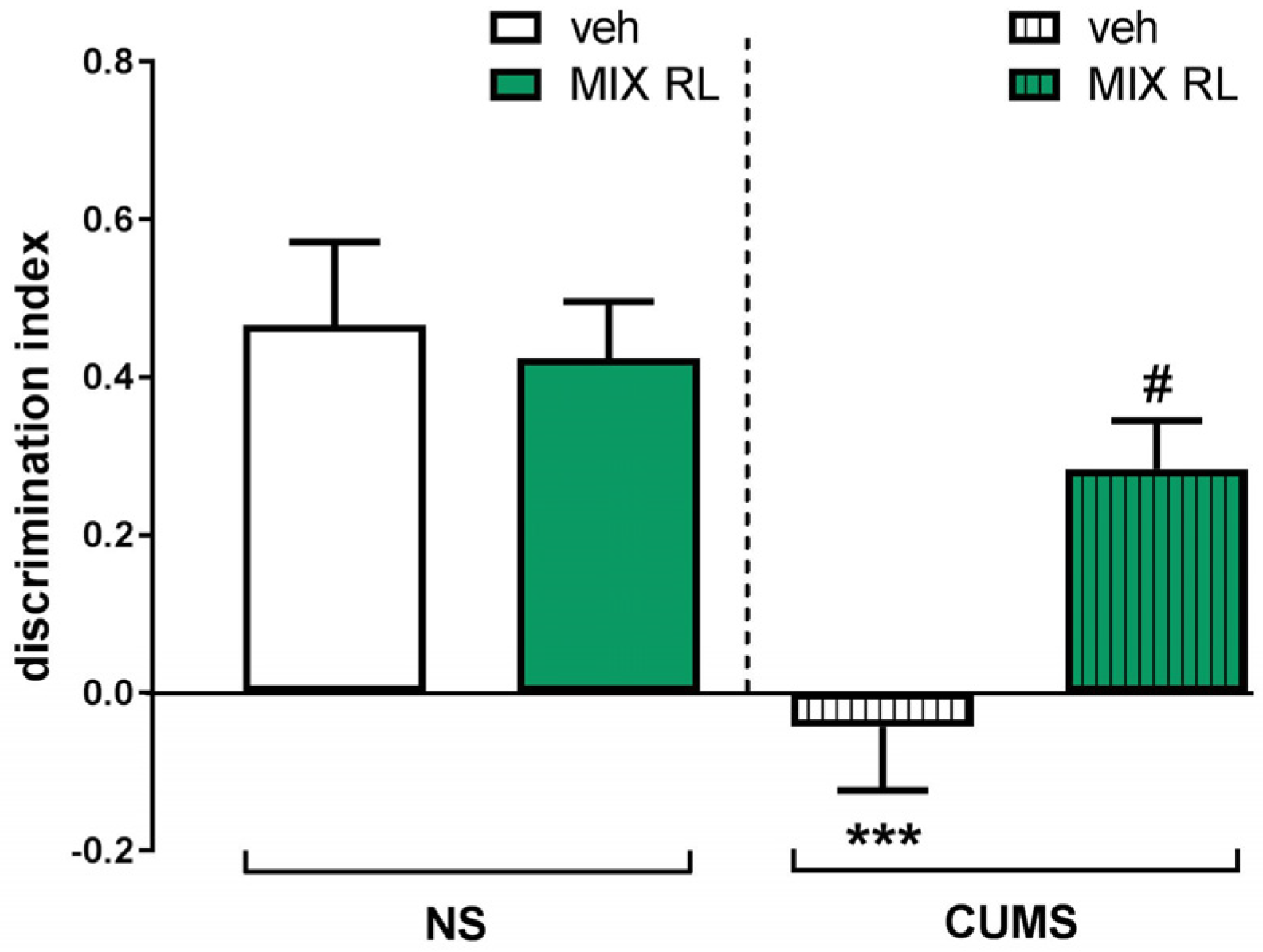
2.1. Effect of (R)-Ketamine Co-Administered with LY341495 (mixRL) in the Temporal Order Memory Task in the CUMS Model of Depression in Mice
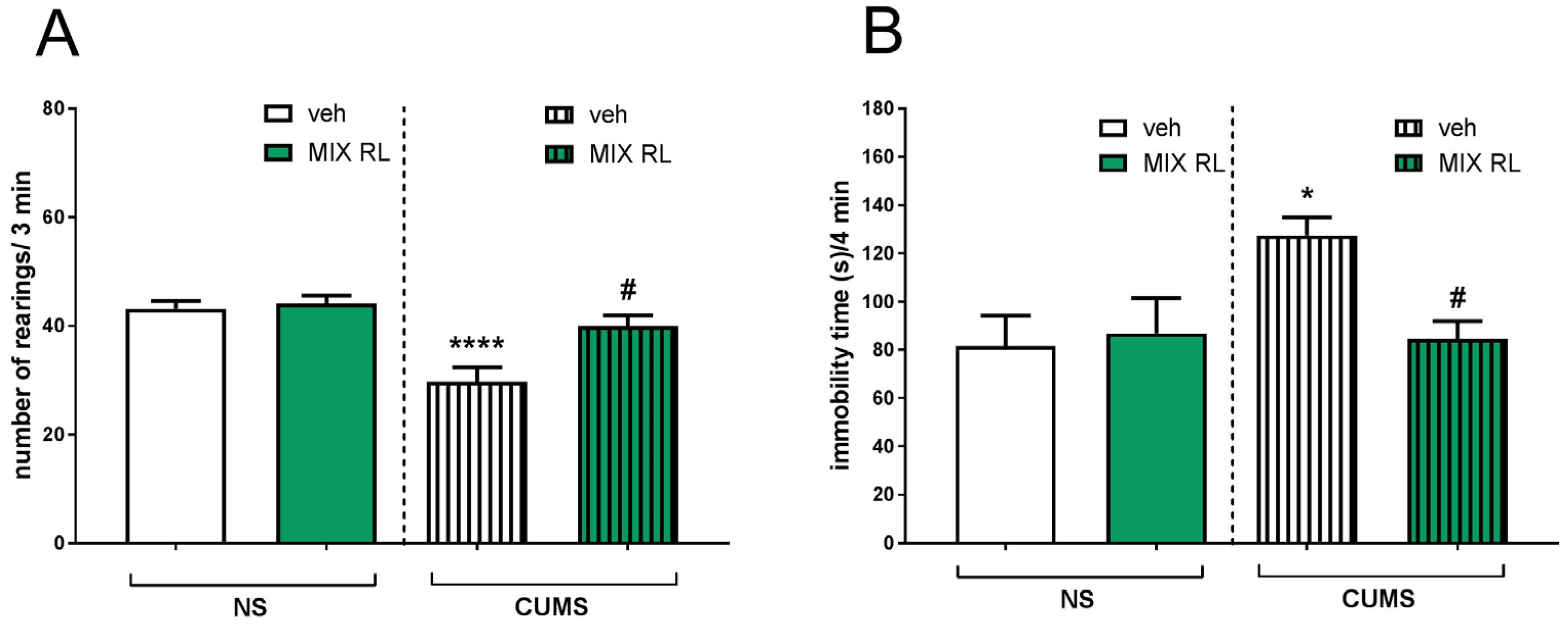
2.2. Antidepressant-like Effects of (R)-Ketamine Co-Administered with LY341495 (mixRL) in the CUMS Model of Depression in Mice
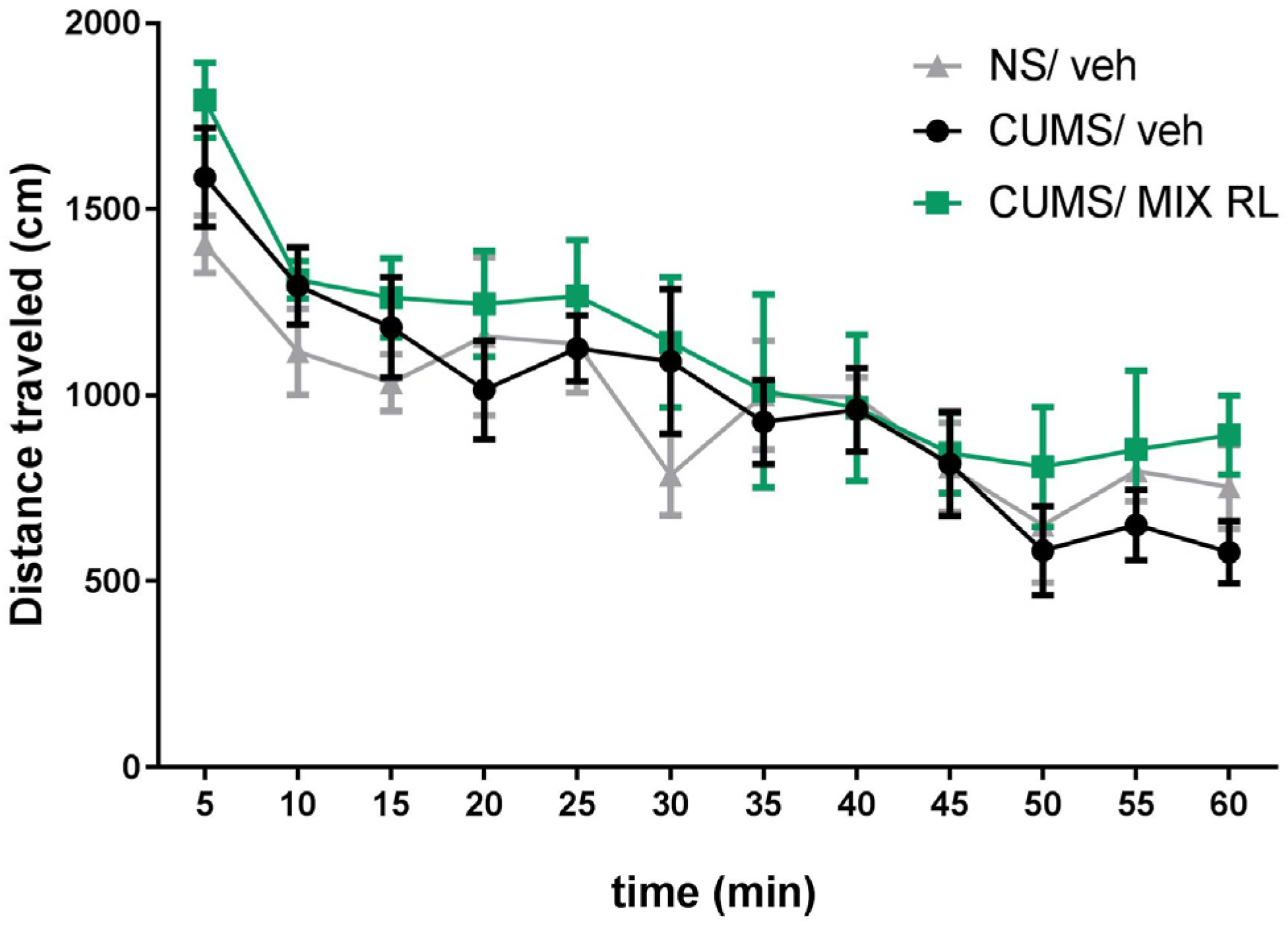
2.3. Effects of (R)-Ketamine Co-Administered with LY341495 (mixRL) on the Locomotor Activity of Mice in the CUMS Model of Depression
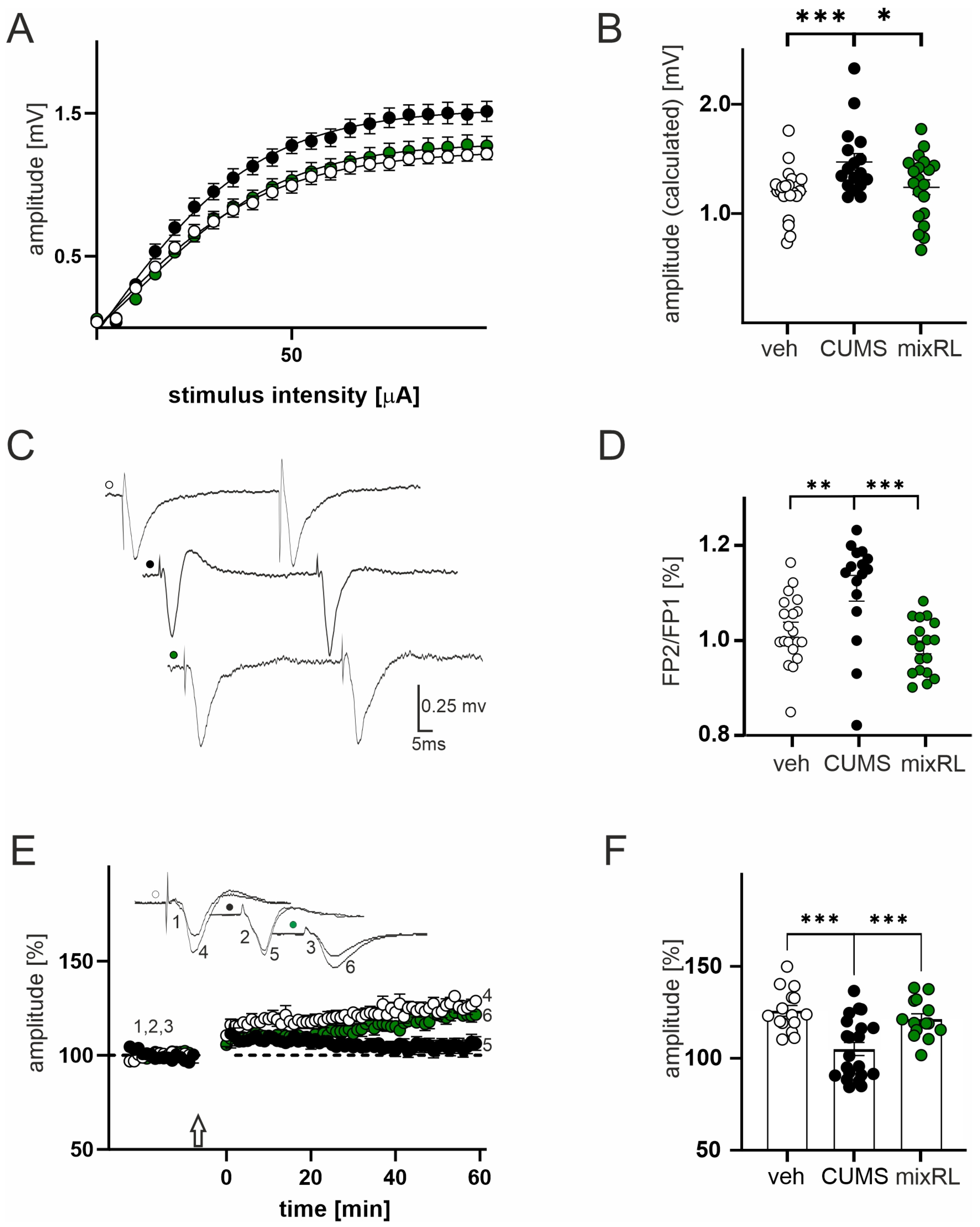
2.4. Effects of (R)-Ketamine Co-Administered with LY341495 (mixRL) in the CUMS Model of Depression on Field Potential (FP) Recording and Paired-Pulse Stimulation
2.5. Effects of (R)-Ketamine Co-Administered with LY341495 (mixRL) in the CUMS Model of Depression on LTP Induction
3. Discussion
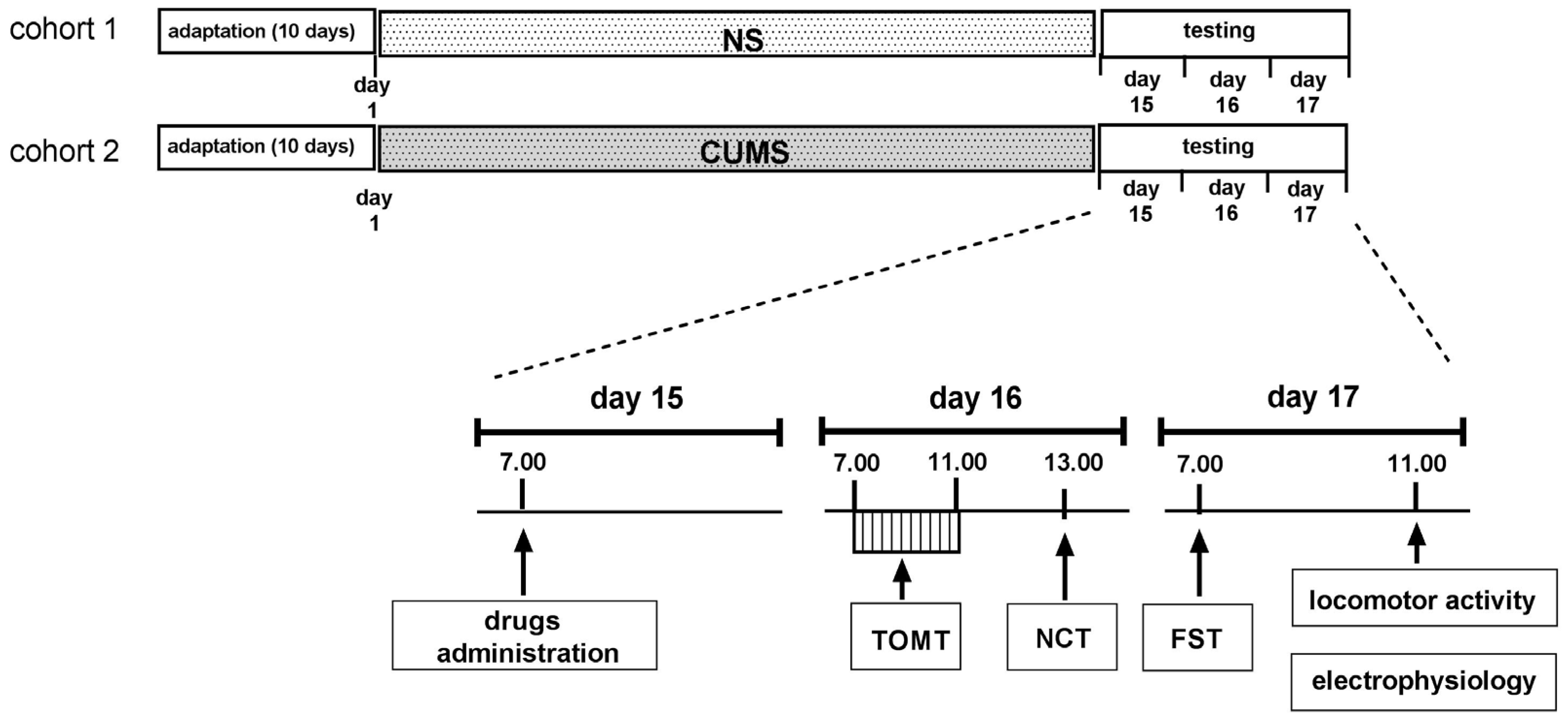
4. Materials and Methods
4.1. Animals and Housing
4.2. Compounds
4.3. CUMS Procedure
4.4. Behavioral Studies
4.4.1. Temporal Order Memory Task (TOMT)
4.4.2. New Cage Test (NCT)
4.4.3. Forced Swim Test (FST)
4.4.4. Locomotor Activity Test
4.5. Electrophysiology
4.5.1. Slice Preparation
4.5.2. Field Potential (FP) Recording, Paired-Pulse Stimulation and LTP Induction
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thase, M.E.; Schwartz, T.L. Using mechanism of action to choose medications for treatment-resistant depression. J. Clin. Psychiatry 2015, 76, e1147. [Google Scholar] [CrossRef] [PubMed]
- Berman, R.M.; Cappiello, A.; Anand, A.; Oren, D.A.; Heninger, G.R.; Charney, D.S.; Krystal, J.H. Antidepressant effects of ketamine in depressed patients. Biol. Psychiatry 2000, 47, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Mahase, E. Esketamine is approved in Europe for treating resistant major depressive disorder. BMJ 2019, 367, l7069. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.C.; Yao, W.; Hashimoto, K. Arketamine, a new rapid-acting antidepressant: A historical review and future directions. Neuropharmacology 2022, 218, 109219. [Google Scholar] [CrossRef]
- Leal, G.C.; Bandeira, I.D.; Correia-Melo, F.S.; Telles, M.; Mello, R.P.; Vieira, F.; Lima, C.S.; Jesus-Nunes, A.P.; Guerreiro-Costa, L.N.F.; Marback, R.F.; et al. Intravenous arketamine for treatment-resistant depression: Open-label pilot study. Eur. Arch. Psychiatry Clin. Neurosci. 2021, 271, 577–582. [Google Scholar] [CrossRef]
- Pałucha-Poniewiera, A. The role of mGlu2/3 receptor antagonists in the enhancement of the antidepressant-like effect of ketamine. Pharmacol. Biochem. Behav. 2022, 220, 173454. [Google Scholar] [CrossRef]
- Zanos, P.; Highland, J.N.; Stewart, B.W.; Georgiou, P.; Jenne, C.E.; Lovett, J.; Morris, P.J.; Thomas, C.J.; Moaddel, R.; Zarate, C.A., Jr.; et al. (2R,6R)-hydroxynorketamine exerts mGlu2 receptor-dependent antidepressant actions. Proc. Natl. Acad. Sci. USA 2019, 116, 6441–6450. [Google Scholar] [CrossRef]
- Pałucha-Poniewiera, A.; Podkowa, K.; Rafało-Ulińska, A. The group II mGlu receptor antagonist LY341495 induces a rapid antidepressant-like effect and enhances the effect of ketamine in the chronic unpredictable mild stress model of depression in C57BL/6J mice. Prog. Neuro. Psychopharmacol. Biol. Psychiatry 2021, 109, 110239. [Google Scholar] [CrossRef]
- Podkowa, K.; Pochwat, B.; Brański, P.; Pilc, A.; Pałucha-Poniewiera, A. Group II mGlu receptor antagonist LY341495 enhances the antidepressant-like effects of ketamine in the forced swim test in rats. Psychopharmacology 2016, 233, 2901–2914. [Google Scholar] [CrossRef]
- Rafało-Ulińska, A.; Brański, P.; Pałucha-Poniewiera, A. Combined administration of (R)-Ketamine and the mGlu2/3 receptor antagonist LY341495 induces rapid and sustained effects in the CUMS model of depression via a TrkB/BDNF-dependent mechanism. Pharmaceuticals 2022, 15, 125. [Google Scholar] [CrossRef]
- Radley, J.J.; Sisti, H.M.; Hao, J.; Rocher, A.B.; McCall, T.; Hof, P.R.; McEwen, B.S.; Morrison, J.H. Chronic behavioral stress induces apical dendritic reorganization in pyramidal neurons of the medial prefrontal cortex. Neuroscience 2004, 125, 1–6. [Google Scholar] [CrossRef]
- Li, N.; Lee, B.; Liu, R.J.; Banasr, M.; Dwyer, J.M.; Iwata, M.; Li, X.Y.; Aghajanian, G.; Duman, R.S. mTOR-dependent synapse formation underlies the rapid antidepressant effects of NMDA antagonists. Science 2010, 329, 959–964. [Google Scholar] [CrossRef]
- Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2017, 6, 78–93. [Google Scholar] [CrossRef]
- Pałucha-Poniewiera, A.; Podkowa, K.; Rafało-Ulińska, A.; Brański, P.; Burnat, G. The influence of the duration of chronic unpredictable mild stress on the behavioural responses of C57BL/6J mice. Behav. Pharmacol. 2020, 31, 574–582. [Google Scholar] [CrossRef]
- Strekalova, T.; Spanagel, R.; Bartsch, D.; Henn, F.A.; Gass, P. Stress-induced anhedonia in mice is associated with deficits in forced swimming and exploration. Neuropsychopharmacology 2004, 29, 2007–2017. [Google Scholar] [CrossRef]
- Bobula, B.; Sowa, J.; Hess, G. Anti-interleukin-1β antibody prevents the occurrence of repeated restraint stress-induced alterations in synaptic transmission and long-term potentiation in the rat frontal cortex. Pharmacol. Rep. 2015, 67, 123–128. [Google Scholar] [CrossRef]
- Czéh, B.; Vardya, I.; Varga, Z.; Febbraro, F.; Csabai, D.; Martis, L.S.; Højgaard, K.; Henningsen, K.; Bouzinova, E.V.; Miseta, A.; et al. Long-term stress disrupts the structural and functional integrity of GABAergic neuronal networks in the medial prefrontal cortex of rats. Front. Cell. Neurosci. 2018, 12, 148. [Google Scholar] [CrossRef]
- Delgado y Palacios, R.; Campo, A.; Henningsen, K.; Verhoye, M.; Poot, D.; Dijkstra, J.; Van Audekerke, J.; Benveniste, H.; Sijbers, J.; Wiborg, O.; et al. Magnetic resonance imaging and spectroscopy reveal differential hippocampal changes in anhedonic and resilient subtypes of the chronic mild stress rat model. Biol. Psychiatry 2011, 70, 449–457. [Google Scholar] [CrossRef]
- Popoli, M.; Yan, Z.; McEwen, B.S.; Sanacora, G. The stressed synapse: The impact of stress and glucocorticoids on glutamate transmission. Nat. Rev. Neurosci. 2011, 13, 22–37. [Google Scholar] [CrossRef]
- Sowa, J.; Bobula, B.; Głombik, K.; Ślusarczyk, J.; Basta-Kaim, A.; Hess, G. Prenatal stress enhances excitatory synaptic transmission and impairs long-term potentiation in the frontal cortex of adult offspring rats. PLoS ONE 2015, 10, e0119407. [Google Scholar] [CrossRef] [Green Version]
- Musazzi, L.; Treccani, G.; Popoli, M. Glutamate hypothesis of depression and its consequences for antidepressant treatments. Expert Rev. Neurother. 2012, 12, 1169–1172. [Google Scholar] [CrossRef] [PubMed]
- Pałucha-Poniewiera, A.; Pilc, A. Glutamate-based drug discovery for novel antidepressants. Expert Opin. Drug Discov. 2016, 11, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Bobula, B.; Tokarski, K.; Hess, G. Repeated administration of antidepressants decreases field potentials in rat frontal cortex. Neuroscience 2003, 120, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Michael-Titus, A.T.; Bains, S.; Jeetle, J.; Whelpton, R. Imipramine and phenelzine decrease glutamate overflow in the prefrontal cortex: A possible mechanism of neuroprotection in major depression? Neuroscience 2000, 100, 681–684. [Google Scholar] [CrossRef]
- Gołembiowska, K.; Dziubina, A. Effect of acute and chronic administration of citalopram on glutamate and aspartate release in the rat prefrontal cortex. Pol. J. Pharmacol. 2000, 52, 441–448. [Google Scholar]
- Moghaddam, B.; Adams, B.; Verma, A.; Daly, D. Activation of glutamatergic neurotransmission by ketamine: A novel step in the pathway from NMDA receptor blockade to dopaminergic and cognitive disruptions associated with the prefrontal cortex. J. Neurosci. 1997, 17, 2921–2927. [Google Scholar] [CrossRef]
- Sala, N.; Paoli, C.; Bonifacino, T.; Mingardi, J.; Schiavon, E.; La Via, L.; Milanese, M.; Tornese, P.; Datusalia, A.K.; Rosa, J.; et al. Acute ketamine facilitates fear memory extinction in a rat model of PTSD along with restoring glutamatergic alterations and dendritic atrophy in the prefrontal cortex. Front. Pharmacol. 2022, 13, 759626. [Google Scholar] [CrossRef]
- McEwen, B.S.; Bowles, N.P.; Gray, J.D.; Hill, M.N.; Hunter, R.G.; Karatsoreos, I.N.; Nasca, C. Mechanisms of stress in the brain. Nat. Neurosci. 2015, 18, 1353–1363. [Google Scholar] [CrossRef]
- McEwen, B.S.; Nasca, C.; Gray, J.D. Stress effects on neuronal structure: Hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacology 2016, 41, 3–23. [Google Scholar] [CrossRef]
- Bobula, B.; Wabno, J.; Hess, G. Imipramine counteracts corticosterone-induced enhancement of glutamatergic transmission and impairment of long-term potentiation in the rat frontal cortex. Pharmacol. Rep. 2011, 63, 1404–1412. [Google Scholar]
- Zelek-Molik, A.; Bobula, B.; Gądek-Michalska, A.; Chorążka, K.; Bielawski, A.; Kuśmierczyk, J.; Siwiec, M.; Wilczkowski, M.; Hess, G.; Nalepa, I. Psychosocial crowding stress-induced changes in synaptic transmission and glutamate receptor expression in the rat frontal cortex. Biomolecules 2021, 11, 294. [Google Scholar] [CrossRef]
- Burgdorf, J.; Kroes, R.A.; Zhang, X.L.; Gross, A.L.; Schmidt, M.; Weiss, C.; Disterhoft, J.F.; Burch, R.M.; Stanton, P.K.; Moskal, J.R. Rapastinel (GLYX-13) has therapeutic potential for the treatment of post-traumatic stress disorder: Characterization of a NMDA receptor-mediated metaplasticity process in the medial prefrontal cortex of rats. Behav. Brain Res. 2015, 294, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Zhang, F.; You, Y.; Wang, H.; Yuan, S.; Wu, B.; Zhu, R.; Liu, D.; Yan, F.; Wang, Z. S-ketamine exerts antidepressant effects by regulating Rac1 GTPase mediated synaptic plasticity in the hippocampus of stressed rats. Cell Mol. Neurobiol. 2022, 43, 299–314. [Google Scholar] [CrossRef]
- Aleksandrova, L.R.; Wang, Y.T.; Phillips, A.G. Ketamine and its metabolite, (2R,6R)-HNK, restore hippocampal LTP and long-term spatial memory in the Wistar-Kyoto rat model of depression. Mol. Brain 2020, 13, 92. [Google Scholar] [CrossRef]
- Yang, Y.; Ju, W.; Zhang, H.; Sun, L. Effect of ketamine on LTP and NMDAR EPSC in hippocampus of the chronic social defeat stress mice model of depression. Front. Behav. Neurosci. 2018, 12, 229. [Google Scholar] [CrossRef]
- Lynch, M.A. Long-term potentiation and memory. Physiol. Rev. 2004, 84, 87–136. [Google Scholar] [CrossRef]
- Dillon, D.G.; Pizzagalli, D.A. Mechanisms of memory disruption in depression. Trends Neurosci. 2018, 41, 137–149. [Google Scholar] [CrossRef]
- Liu, S.C.; Hu, W.Y.; Zhang, W.Y.; Yang, L.; Li, Y.; Xiao, Z.C.; Zhang, M.; He, Z.Y. Paeoniflorin attenuates impairment of spatial learning and hippocampal long-term potentiation in mice subjected to chronic unpredictable mild stress. Psychopharmacology 2019, 236, 2823–2834. [Google Scholar] [CrossRef]
- Barker, G.R.; Bird, F.; Alexander, V.; Warburton, E.C. Recognition memory for objects, place, and temporal order: A disconnection analysis of the role of the medial prefrontal cortex and perirhinal cortex. J. Neurosci. 2007, 27, 2948–2957. [Google Scholar] [CrossRef] [Green Version]
- Ting, J.T.; Lee, B.R.; Chong, P.; Soler-Llavina, G.; Cobbs, C.; Koch, C.; Zeng, H.; Lein, E. Preparation of acute brain slices using an optimized N-methyl-D-glucamine protective recovery method. J. Vis. Exp. 2018, 132, 53825. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vmax (mV) | uh (µA) | S | N | |
|---|---|---|---|---|
| veh | 1.18 ± 0.04 | 27.26 ± 1.7 | 11.39 ± 0.9 | 24 |
| CUMS | 1.47 ± 0.07 ***,# | 24.59 ± 2.33 | 9.82 ± 1.6 | 17 |
| MixRL | 1.24 ± 0.06 | 27.59 ± 1.33 | 11.65 ± 1.43 | 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pałucha-Poniewiera, A.; Bobula, B.; Rafało-Ulińska, A. The Antidepressant-like Activity and Cognitive Enhancing Effects of the Combined Administration of (R)-Ketamine and LY341495 in the CUMS Model of Depression in Mice Are Related to the Modulation of Excitatory Synaptic Transmission and LTP in the PFC. Pharmaceuticals 2023, 16, 288. https://doi.org/10.3390/ph16020288
Pałucha-Poniewiera A, Bobula B, Rafało-Ulińska A. The Antidepressant-like Activity and Cognitive Enhancing Effects of the Combined Administration of (R)-Ketamine and LY341495 in the CUMS Model of Depression in Mice Are Related to the Modulation of Excitatory Synaptic Transmission and LTP in the PFC. Pharmaceuticals. 2023; 16(2):288. https://doi.org/10.3390/ph16020288
Chicago/Turabian StylePałucha-Poniewiera, Agnieszka, Bartosz Bobula, and Anna Rafało-Ulińska. 2023. "The Antidepressant-like Activity and Cognitive Enhancing Effects of the Combined Administration of (R)-Ketamine and LY341495 in the CUMS Model of Depression in Mice Are Related to the Modulation of Excitatory Synaptic Transmission and LTP in the PFC" Pharmaceuticals 16, no. 2: 288. https://doi.org/10.3390/ph16020288