
Radiotracers for the Central Serotoninergic System


1
Centre d’Etudes et de Recherche sur le Médicament de Normandie (CERMN), UNICAEN, Normandie Univ., 14000 Caen, France
2
Institut Blood and Brain @ Caen Normandie (BB@C), Boulevard Henri Becquerel, 14000 Caen, France
3
UNICAEN, IMOGERE, Normandie Univ., 14000 Caen, France
4
CHU Côte de Nacre, Department of Nuclear Medicine, 14000 Caen, France
*
Authors to whom correspondence should be addressed.
Pharmaceuticals 2022, 15(5), 571; https://doi.org/10.3390/ph15050571
Submission received: 25 March 2022
/
Revised: 27 April 2022
/
Accepted: 28 April 2022
/
Published: 3 May 2022
(This article belongs to the Special Issue Alliance of PET/SPECT Imaging and Drug Design/Discovery for the Development of Novel Diagnostic or Therapeutic Tools)
Abstract
:This review lists the most important radiotracers described so far for imaging the central serotoninergic system. Single-photon emission computed tomography and positron emission tomography radiotracers are reviewed and critically discussed for each receptor.
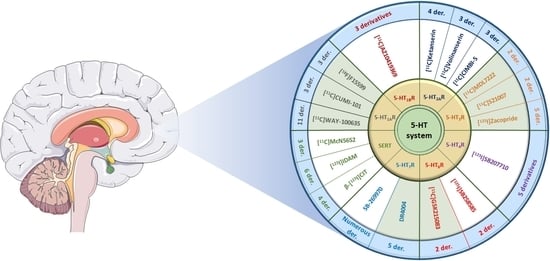
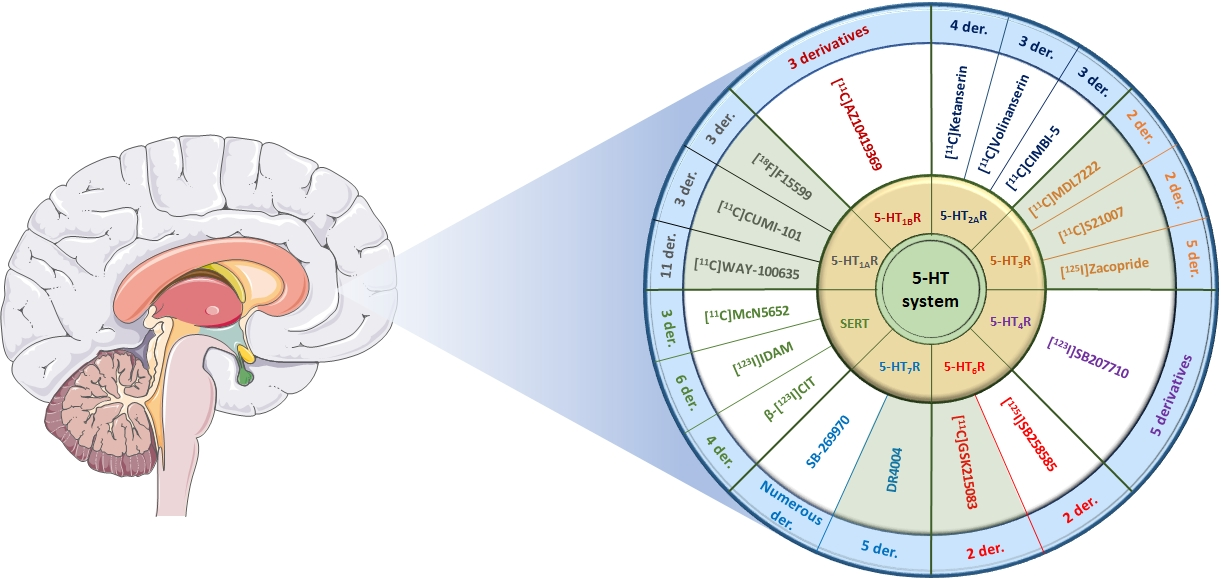{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
In recent decades, the serotoninergic system constituting serotonin (5-HT) and its receptors (5-HTR) has become a promising target in neuroscience. The many studies carried out have shown its involvement in many brain functions and neuropsychiatric disorders such as Alzheimer’s disease, Parkinson’s disease, schizophrenia, or depression [1].
Among the G protein-coupled receptors (GPCR), 5-HTRs appear to be the largest family of receptors. Indeed, the physiological action of serotonin is carried by at least 14 subtypes of receptors: 5-HT1A,B,D,E,F, 5-HT2A-C, 5-HT3, 5-HT4, 5-HT5A, 5-HT6, and 5-HT7 [2,3]. This classification (IUPHAR) is based on their signal transduction pathways, pharmacological effects, and molecular structures.
The significant variability of these receptors has led to the development of new tools, allowing the selective in vivo exploration of each of them. This exploration is based in particular on the use of medical imaging techniques such as single-photon emission computed tomography (SPECT) and positron emission tomography (PET). These non-invasive imaging techniques are commonly used by researchers and clinicians to perform the mapping of neurotransmitter systems in the living brain in healthy and sick patients. Thanks to the development and use of new radiolabeled molecules (radiotracers or radiopharmaceuticals), the precise quantification of the density of these receptors within different brain areas is possible. Radiotracers, which are molecular scaffold containing a radioactive isotope, must fulfill several criteria to be effective brain receptor imaging agents: a high selectivity and high affinity (nM range) for the target receptor, a low or moderate lipophilicity (logD = 1–3) for blood–brain barrier penetration and to avoid excessive nonspecific binding to brain tissue, a slow metabolism (or non-interfering radioactive metabolites), and a high specific activity to visualize weakly abundant biological targets.
In this review, for each 5-HTR subtype, we will focus on the central radiotracers of interest mentioned in the literature, sorted by chemical families. Their interesting properties will be discussed, and their uses in clinical or preclinical trials will be specified. Moreover, the radiotracers used for the detection and quantification of 5-HT reuptake transporters (SERTs) will be described as well. The SERT does not belong to the 5-HTR family, but its involvement in the regulation of the serotonin concentration in the inter-synaptic space makes it an important target in neuroscience and therapeutics [4].
2. 5-HT1 Receptors
5-HT1R receptors are involved in many biological and physiological processes and constitute a large family of receptors. Five different isoforms (5-HT1A, 5-HT1B, 5HT1D, 5-HT1E, and 5-HT1F) exist, all coupled to a Gαi/o protein which inhibits cAMP synthesis and therefore potentially inhibits signal transduction. In the current classification, we can note the absence of 5-HT1C because the latter has been reclassified as 5-HT2c following numerous discoveries for GPCRs and their sequence homology with the 5-HT2R family [5]. To date, the description of efficient radiotracers of the 5-HT1R family have been restricted to the 5-HT1AR and 5-HT1BR subtypes.
2.1. 5-HT1A Receptors
Among the 5-HTRs, 5-HT1AR is the first to have been cloned and characterized. In 1987, its gene (G-21) was cloned and identified as a GPCR by structural homology with the β2-adrenergic receptor [6]. However, this is the work of Fargin et al., which determined that G-21 coded for 5-HT1AR [7]. In the mammalian brain, 5-HT1ARs are divided into two distinct populations based on their location, including (1) presynaptic 5-HT1AR (autoreceptors) located in the soma and dendrites of serotoninergic neurons in the raphe nucleus [8,9], where their activation leads to a reduction in nerve impulses (Gαi/o), resulting in a decrease in the release of 5-HT in the inter-synaptic space [10,11], and (2) postsynaptic 5-HT1AR (heteroreceptors) localized in non-serotonergic neurons, mainly in limbic areas such as on the dendrites and soma of glutamatergic neurons, the terminal axons of GABAergic neurons, or on cholinergic neurons [12]. Their action is inhibitory and extends to the many areas of the brain innervated by the serotoninergic system. It has to be noted that 5-HT1AR heteroreceptors are expressed in a high density in the hippocampus (zone CA1, CA2, and serrated gyrus), the septum pellucidum, layers II and VI of the frontal cortex, the lateral and medial septum, and more moderately in the amygdala, the inferior colliculus, and in the thalamic and hypothalamic nuclei [8]. The wide distribution of 5-HT1AR suggests that they have a great variety of functions in the brain. Indeed, they are distinguished by their influence in the phenomena of neurogenesis, neuroprotection, neuronal plasticity, memorization, and learning, but also by their involvement in various behavioral disorders such as anxiety and depression [13,14,15]. They are currently a target of choice in medical therapy, research, and medical imaging via the development of antagonists, agonists, and radiotracers specific to this target.
Many 5-HT1A receptor (5-HT1AR) radiotracers are described in the literature, but very few of them are currently used in human studies [16]. Indeed, there is a significant bias between the in vitro results and the different models (rodent, non-rodent, monkey, etc.) used in pre-clinical trials due to a difference in the distribution of 5-HT1AR in the cerebral areas of the species studied, the intensity of the signal collected compared with the background noise, and the metabolization or distribution phenomena limiting the contact between the radiotracer and its target.
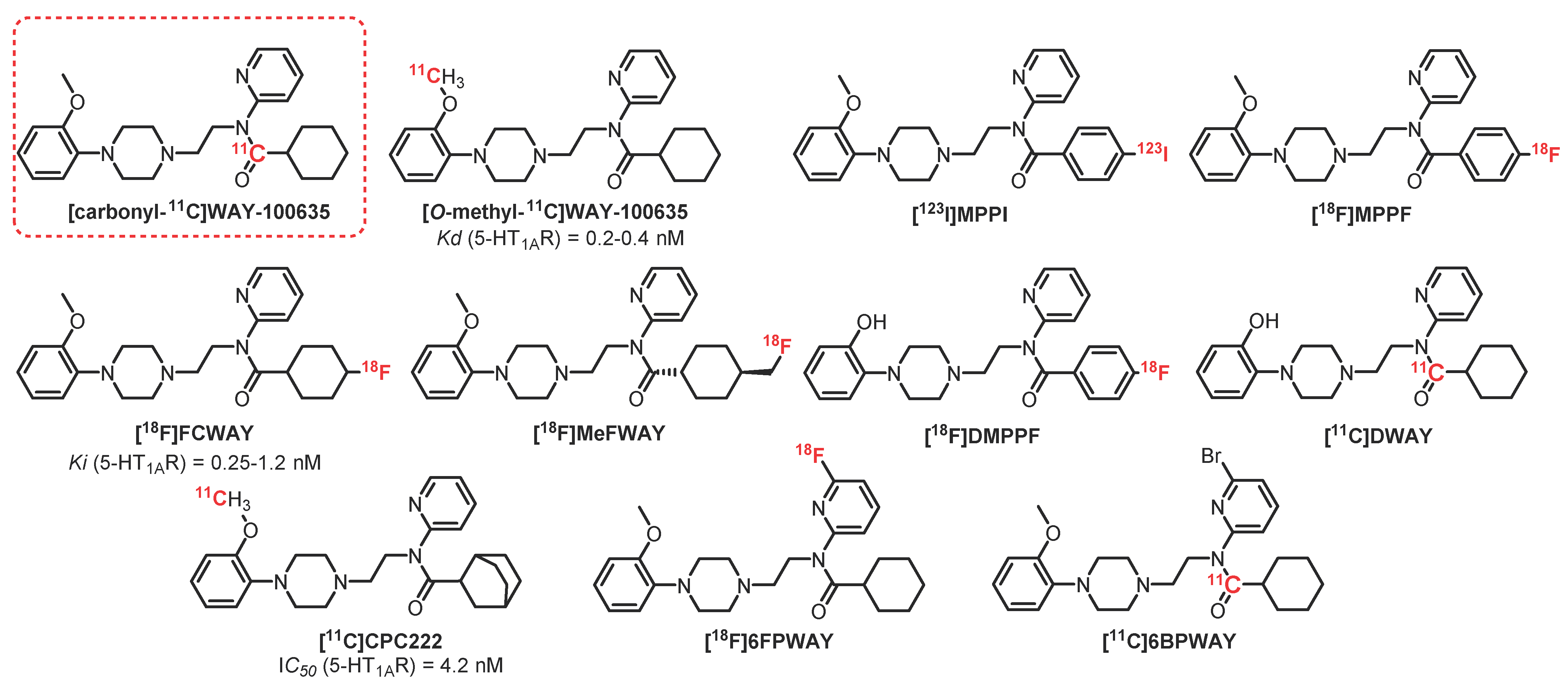
2.2. N-Acetamide Pyridine Series: WAY-100635 Derivatives
[11C]WAY-100635 (Figure 1) is the first radioligand described as a potent antagonist, presenting good affinity for 5-HT1AR (KD = 0.2–0.4 nM) [17]. This radioligand has shown a partial selectivity, albeit it is also known as a dopamine D4 receptor agonist (KD = 2.4 nM). This lack of selectivity is not really a problem because the 5-HT1AR receptor density in the brain is significantly greater than the population of D4 receptors. Studies carried out on this radiotracer have shown good correlation between the distribution of receptors compared to that obtained previously using agonists and antagonists on human tissues (postmortem) [18]. The [O-methyl-11C]WAY-100635 analogous has been the first radioligand to be synthesized for PET purposes, which led to 5-HT1AR images in rodents, monkeys, and humans [19,20]. However, a high level of nonspecific radioactivity has been detected in the brain due to the easy crossing of BBB by radiometabolites. Then, [carbonyl-11C]WAY-100635 was synthesized, which was degraded into polar radioactive metabolites that did not cross the BBB [21,22]. Since its first synthesis, [carbonyl-11C]WAY-100635 has become a reference radioligand, allowing several studies to be performed first on healthy patients and then on patients presenting several psychiatric conditions such as depression, anxiety, or anorexia [22,23,24,25]. Nevertheless, several pharmacomodulations have been performed in order to mainly reduce its metabolization, leading to the preparation of carbon-11 and fluorine-18 radioligands.
[123I]MPPI, an iodinated analogous of [11C]WAY-100635, was prepared in 1994 by Kung [26]. Nevertheless, due to the instability of the C-I bond in vivo, this compound was radiolabeled with fluorine-18, as the C-F bond is known to be less metabolized. Then, in 1997, the same group prepared [18F]MPPF, which has been proven to be a valuable 5-HT1AR radiotracer in monkeys [27]. [18F]MPPF is a selective antagonist of 5-HT1AR used to measure the evolution of serotonin levels in vivo [28]. Studies realized in rodents, cats, monkeys, and dogs led to a selective labeling of 5-HT1AR-rich cerebral regions (hippocampus, entorhinal cortex, and raphe nuclei) [29,30]. [18F]MPPF has also been tested on healthy patients [31] as well as patients presenting Alzheimer’s disease [32], depression, and temporal lobe epilepsy [33,34].
[18F]FCWAY is a fluorinated analogue of WAY-100635 developed by Lang [35]. Its trans isomer, [4-trans-18F]FCWAY, exhibits a 0.25 nM affinity toward 5-HT1AR, and its cis isomer, [4-cis-18F]FCWAY, has a slightly weaker affinity of 1.2 nM. However, [4-trans-18F]FCWAY showed the best pharmacological and pharmacokinetic properties, leading to the measurement of the 5-HT1AR density in the hippocampus and cerebellum. [18F]FCWAY has also been used to quantify 5-HT1AR in patients with behavioral disorders such as epilepsy, post-traumatic stress, or panic attacks [36,37,38]. However, the accumulation of radiofluorine in bones due to the metabolism of the radiotracer decreases the resolution of brain imaging. The use of disulfiram, a cytochrome P450 isozyme 2E1 inhibitor which decreases defluorination, limited this phenomenon and improved the visualization of these receptors, but it was not sufficient to justify its use or its marketing [39].
[18F]MeFWAY is a derivative of [18F]FCWAY, possessing a fluoromethyl group at cyclohexane’s position 4, thus limiting metabolization issues. In vivo assays in rodents and primates showed that the [trans-18F]MeFWAY isomer presented good affinity and specificity for 5-HT1AR and was suitable for in vivo imaging [40,41]. In 2014, the first assays on six healthy volunteers allowed obtaining preliminary evidences of its potential as a radiopharmaceutical [42]. In 2015, Choi et al. confirmed this conclusion by using a comparative study with [18F]FCWAY associated with disulfirame [43]. Nevertheless, to date, there is no publication demonstrating its usefulness in sick patients.
[18F]DMPPF is the demethylated analogue of [18F]MPPF. This pharmacomodulation was performed in order to increase the PET signal by improving the cerebral uptake of the compound. [18F]DMPPF was crossing the BBB and leading to an efficient imaging contrast with a lower clearance in rats than [18F]MPPF [44]. Despite this encouraging first result, this radiotracer has not been used, probably due to the great interest of the community in [11C]WAY-100635.
[11C]DWAY is a putative metabolite of [11C]WAY-100635 which presents similar characteristics but provides a twofold higher brain uptake than [11C]WAY-100635 in the human brain [45]. However, despite these interesting results, this radiotracer has not been investigated so far.
[11C]CPC-222 is a WAY-100635 analogue with good 5-HT1AR affinity (IC50 = 4.2 nM) which crosses the BBB [46]. However, this molecule has not been used because of its too weak signal-to-noise ratio.
[18F]6FPWAY and [11C]6BPWAY are halogenated (fluorinated and brominated, respectively) analogues of WAY-100635 [47]. Halogen atoms have been introduced on this scaffold in order to decrease the metabolism of the compound while allowing further radiolabeling with fluorine-18 or bromine-76. [18F]6FPWAY was initially synthesized and tested as [11C]6FPWAY in monkeys and then was prepared while incorporating fluorine-18 [48]. Assays have shown a moderate interest in these ligands due to faster metabolization of the compound compared with [O-methyl-11C]WAY-100635. This phenomenon is probably due to faster degradation of the amide bond of the 6-fluoropyridine group [49]. [18F]6FPWAY and [11C]6BPWAY have not been further studied so far.
2.3. 1,2,4-Triazine-3,5-Dione Series: CUMI-101 Derivatives
[11C]CUMI-101 and its derivative [11C]FECUMI-101 are 5-HT1AR agonists (Figure 2). Agonists are good candidates for estimating the level of 5-HT receptors present in the synaptic cleft, the receptor’s occupancy, and the determination of the internalization of autoreceptors after administration of the serotonin transporter’s (SERT) inhibitors, which are used to treat depressive states. [11C]CUMI-101, also called [11C]MMP, is a 2-methoxyphenyl analogous of a previous radiotracer (MPT) described by the same authors in 2006. This former radiotracer has shown a promising agonist profile with good affinity for 5-HT1AR (Ki = 1.36 nM) in baboons [50], but slow washout kinetics prompted the authors to perform structure–activity relationship studies, leading to the preparation of [11C]CUMI-101. This compound has been demonstrated to be a potent (Ki = 0.15 nM) and selective 5-HT1AR agonist ligand, with a high uptake in 5-HT1AR-rich areas in baboons [51]. Used in healthy volunteers, [11C]CUMI-101 led to determination of the distribution and quantification of central and peripheral 5-HT1AR [52,53]. Notably, this radiotracer shows interesting properties, as its metabolites are not crossing the BBB. In 2011, Hendry et al. investigated the pharmalogical profile determination of [11C]CUMI-101 in rat brain tissue and demonstrated that, while acting as an 5-HT1AR agonist in humans, it behaves as an antagonist in the rat cortex and rat hippocampal tissue [54]. Thus, this compound may act as a partial agonist, leading to interpretation difficulties from one model to another. In 2020, [11C]CUMI was also used to assess the 5-HT1AR occupancies of brexpiprazole in adult subjects with schizophrenia [55].
[18F]FECUMI-101 is the ethylfluorinated analogue of [11C] CUMI-101 synthesized in 2013 by Kumar [56]. Studies carried out in baboons showed that this radiotracer crosses the BBB with good distribution of the radiotracer in the 5-HT1AR-rich brain regions, with the exception of the thalamus. In 2016, autoradiographic studies carried out on human brain sections confirmed this specific binding, along with some binding with α1 receptors [57]. A possible explanation is the difference in the concentration of α1 receptors in different species in this brain region or a specific binding on an unknown target. In addition, competition experiments in the presence of WAY-100635 have shown a binding blockade of [18F]FECUMI-101 on the receptors present in the cerebellum, limiting its use as a reference for in vivo PET applications.
[18F]FEMPT is a radiotracer synthesized in 2017 by Collier et al. via a continuous flow microfluidic method [58]. The advantage of this technique lies mainly in the saving of time and the reagent, allowing one to increase and optimize the parameters and the yield of reactions for radiolabeling with fluorine-18. The first in vitro assays carried out with this compound showed a strong affinity of [18F]FEMPT toward 5-HT1AR (Ki = 0.2 nM). In addition, the affinity for α1 adrenergic receptors (Ki = 180 nM) was much lower than that observed for [11C]CUMI-101. These initial results are very encouraging, and preclinical studies are currently being carried out on this new radiotracer.
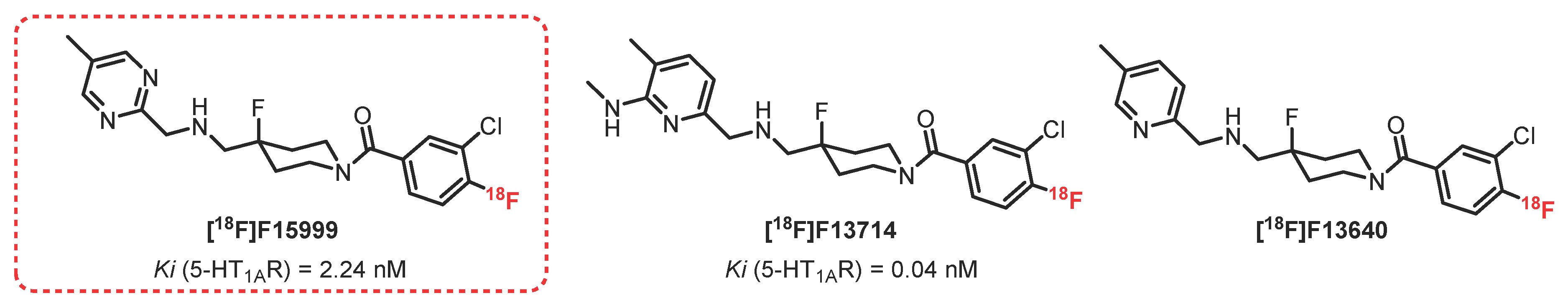
2.4. 2-Pyridinemethylamine Series: F15599 Derivatives
F15599 (Figure 3) is the first 5-HT1AR ligand described in the 2-pyrimidinemethylamine series, presenting a strong affinity and very high specificity for 5-HT1AR. In vitro and in vivo studies led to assessing its particular affinity for the postsynaptic 5-HT1AR present in the cortical areas of the brain [44]. In vivo imaging with [18F]F15599 in rats and cats demonstrated a rapid accumulation of this radioligand in the brain, suggesting good BBB passage as well as good distribution in brain areas such as the raphe nucleus, the hippocampus, and the side bridge [45]. However, the labeling intensity was considered insufficient for further investigations as a radiopharmaceutical, probably due to its relatively low affinity (Ki = 2.24 nM) [16]. Efforts have been therefore focused on pharmacomodulations of [18F]F15599.
[18F]F13714, a 2-pyridinemethylamine analogue of [18F]F15999, presented a particularly strong affinity for recombinant human 5-HT1AR (Ki = 0.04 nM) [59,60]. In vitro autoradiography carried out on rats revealed significant radioactivity in the targeted brain areas [61]. In addition, [18F]F13714 was shown to have very good resistance to metabolism. Specific binding of this radiotracer in 5-HT1AR-dense areas was observed in in vivo PET imaging (cats), with a particular tropism for the cortex compared with the hippocampus. In 2016, Yokoyama et al. published a comparative study between [18F]F13714 and [18F]MPPF in awake and anesthetized marmosets, demonstrating the strong influence of anesthesia on functional 5-HT1AR and therefore suggesting that caution is necessary when interpreting results from agonists in PET imaging [62]. Although the results were encouraging, [18F]F13714 was not optimal for in vivo imaging because of its irreversible binding.
F13640 (also known as befiradol or NLX-112) is a 5-HT1AR ligand that has recently been investigated from the same series, presenting selectivity and efficacy for 5-HT1AR along with strong analgesic properties [63]. Demonstrated on a large panel of animal models presenting physiological or pathological acute pain, and in comparison with a large intake of opioids, its analgesic action led to the development of a reverse tolerance [64]. In 2018, Zimmer et al. carried out the first in vivo imaging assays in rats, cats, and primates [65]. [18F]F13640 has been proven to cross the BBB (without formation of radioactive metabolites), and its distribution has been observed in 5-HT1AR-rich areas, with notable differences in comparison with the distribution observed using antagonist radiotracers so far. In addition, [18F]F13640 has been used to determine the evolution of the number of these receptors in the hippocampus in the prodromal stage of Alzheimer’s disease (post mortem) [66]. In 2019, [18F]F13640 was used in vivo in healthy humans, affording promising results since the visualized distribution correlated with that of functional 5-HT1AR receptors [67]. [18F]F13640 is the first radiopharmaceutical that enables in vivo investigation of functional 5-HT1AR that is likely to be altered in pathological conditions such as neuro-degenerative diseases or psychiatric disorders. Indeed, in 2021, [18F]F13640 was proven to be a valuable tool to explore neurological or neuropsychiatric pathologies involving fluctuations in extracellular serotonin. Indeed, Zimmer et al. have shown its sensitivity to detecting in vivo 5-HT concentration fluctuations [68].
2.5. 5-HT1B Receptors
Like 5-HT1AR, 5-HT1BR exists at the pre- and postsynaptic level. However, their locations are different; presynaptic 5-HT1AR is localized mainly in the cell body and the dendrites of the serotoninergic neurons, while 5-HT1BR is expressed at the axonal endings. The 5-HT1BR has an important influence in the regulation of the concentration of 5-HT in the intersynaptic space and induces a negative feedback limiting 5-HT’s release (homoreceptors). All postsynaptic 5-HT1BR (heteroreceptors) receptors are located on the terminal axons of non-serotonergic neurons (GABAergic, muscarinic, or dopaminergic) and will influence the release of their neurotransmitters. There is a great homology between the receptors present in humans and rats, with the only difference being a replacement of a threonine by an asparagine on the seventh transmembrane domain. However, this slight difference is responsible for significant variations during pharmacological studies. The 5-HT1BR receptors are present in a high density in different brain locations, such as the globus pallidus, substance nigra, ventral pallidum, or dorsal subiculum and more moderately in the cortex, the hippocampus, and the putamen [69]. The 5-HT1BR receptors are involved in learning and memorization phenomena. Thus, the injection of antagonists into knockout mice allowed the observation of an improvement in their cognitive performance, with a probable involvement of the cholinergic system [70]. In addition, pharmacological studies have demonstrated the interest of 5-HT1BR in aggression phenomena, with a decrease of the latter in the presence of agonists [71].
2.6. Chromen-4-One Series: AZ10419369 Derivatives
[11C]AZ10419369 (Figure 4) is a partial 5-HT1BR agonist (KD = 0.37 nM). This compound is considered a reference radiotracer for these receptors and has been used on numerous occasions to assess the influence of 5-HT1BR in healthy volunteers as well as in neuropsychiatric disorders. In 2008, Pierson et al. carried out the first studies in macaques and observed a specific radiolabeling of 5-HT1BR-rich areas through a competitive method in the presence of a reference antagonist: AR-A000002 [72]. In addition, the interesting potential of this compound has been confirmed thanks to the intensity of the collected signal and the absence of detection of any radiometabolite. In this same study, [11C]AZ10419369 was tested in two human subjects, and good correlation in the receptor’s distribution in both macaques and human brains was observed, as well as those obtained previously by autoradiogram in guinea pig brains [73]. From 2010 to 2020, many studies used [11C]AZ10419369 as a reference in PET imaging [74,75]. Used in patients with depressive disorders and Parkinson’s disease, a decrease in 5-HT1BR in the targeted areas of the brain was observed [76,77]. Very recently, pharmacomodulations of [11C]AZ10419369 have been performed by Lindberg et al. in order to replace carbon-11 with the longer half-life fluorine-18, allowing them to increase the acquisition time in imaging [78,79]. Among the described radiotracers, [18F]AZ10419096, a radiotracer published in 2019 by Lindberg et al. [80], revealed interesting properties. This compound is an antagonist of 5-HT1BR presenting very good affinity (Ki = 0.13 nM). The first in vivo tests in monkeys demonstrated a specific binding of this compound to 5-HT1BR-rich brain areas (e.g., the occipital cortex or globus pallidus). Approximately 80% of this binding is inhibited in the presence of a reference antagonist (pretreatment with AR-A000002). In addition, the main metabolites of [18F]AZ10419096 seemed to be too polar to cross the BBB. The only drawback of this compound lies in its synthesis, since the yield of radiolabeling from the boronic precursor is very low (<5%) and needs to be optimized.
[11C]P943, obtained through a structural simplification of [11C]AZ10419369, is a 5-HT1BR antagonist presenting a very strong affinity and specificity for these receptors (Ki = 0.77 nM). A complete pharmacokinetic study was performed in baboons (Papio anubis) in order to determine the BBB crossing, the presence of metabolites, and its affinity for 5-HT1BR. The specificity of [11C]P943 for 5-HT1BR was revealed by blocking the receptors with a specific 5-HT1BR antagonist (SB-616234-S), while no significant change was observed using a 5-HT1DR antagonist (SB-714786) [81]. Thanks to this radiotracer, the implication of 5-HT1BR was evaluated in patients with various pathologies (e.g., depression or post-traumatic stress) or drug addiction (cocaine), where a decrease in the binding of [11C]P943 to 5-HT1BR was observed compared with the healthy patients [82,83]. This phenomenon is reversed in people with alcohol addiction, where an increase in binding was observed, especially in the ventral striatum [84].
3. 5-HT2 Receptors
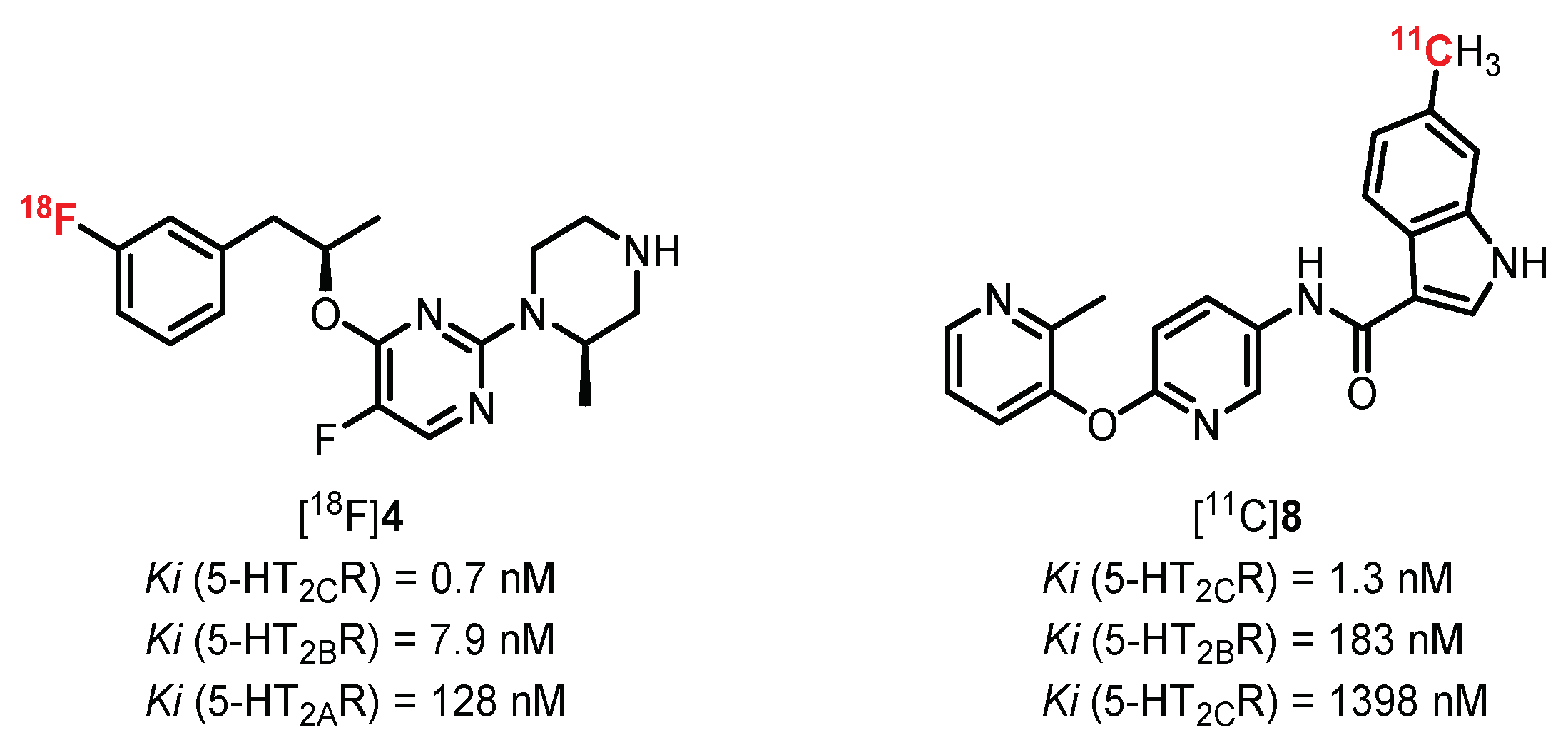
5-HT2R receptors are among the first serotoninergic receptors to be identified with 5-HT1AR. Their identification was possible thanks to genetic cloning, allowing for distinguishing three different subtypes (5-HT2A, 5-HT2B, and 5-HT2C). Structural variations in these 3 isomers with 471 amino acids for 5-HT2AR, 481 for 5-HT2BR, and 458 for 5-HT2CR (in humans) can be noted. This variation in structure is not surprising, since each of them is encoded by different genes carried on three distinct chromosomes (chromosomes 13, 2, and X, respectively). These receptors possess a wide area of localization and are mainly involved in cell development and migration processes [85]. All these receptors are coupled to a Gq/11 protein, and their activation regulates an enzymatic cascade involving the phospholipase C. This enzyme leads to the hydrolysis of membrane phosphatidylinositol biphosphate into two intracellular secondary messengers: diacylglycerol and inositol-1,4,5-triphosphate (IP3). IP3 plays an important role in the release of calcium ions by binding to the IP3 receptors on the membrane of the endoplasmic reticulum. This increase in Ca2+ ions in the cytosol is completed by the action of ion pumps (allowing the passage of extracellular Ca2+ ions) which are activated by protein kinases C, themselves activated by the diacylglycerols synthesized beforehand [86]. Currently, there are no potent and selective radioligands commonly used in clinic or research facilities to quantify and characterize 5-HT2BR and 5-HT2CR. It is nevertheless interesting to note the recent development of promising new molecules that are affine and selective for 5-HT2CR, specifically marking the choroid plexus (cerebral area rich in 5-HT2CR) (Figure 5) [87,88]. These molecules could be the subject of future clinical studies and could enhance the interest shown in these receptors via PET and SPECT imaging.
Conversely, numerous studies involving 5-HT2AR radioligands have been described in the literature. In humans, 5-HT2AR was identified and cloned for the first time by Branchek et al. in 1990 [89]. It quickly aroused the particular interest of the scientific community because of its role in certain behavioral disorders and its psychomimetic effects when taking psychotropic substances such as lysergic acid diethylamide (LSD), mescaline, or psilocybin [90]. Human 5-HT2AR radioligands are located mainly in the cortex and in a lower density in the central brain structures such as the hippocampus or the caudate nucleus. In the cortex, they are found mainly in layers II–III but also in lesser quantities in layers V and VI [91]. At the cellular level, they are mainly postsynaptic receptors and are expressed in non-serotonergic (cholinergic or GABAergic) neurons with the result of increasing cellular hyperexcitability and promoting nerve impulses [92]. 5-HT2AR has been at the center of many studies on the one hand for its involvement in many neuropsychiatric disorders such as depression, anxiety, schizophrenia, and psychosis and on the other hand because it is the target of second-generation antipsychotics (D1/D2 and 5-HT2A antagonists) such as Clozapine, Olanzapine, or Risperidone [93]. To date, many efficient 5-HT2AR radiotracers have been developed in several chemical series both for SPECT and PET.
3.1. Quinazoline-2,4-Dione and Thiazolo [3,2-a]Pyrimidin-5-One Series: Ketanserin Derivatives
The first evaluations of 5-HT2R radiotracers were carried out in 1985 using a very affine and selective antagonist (in vitro and in vivo in rats): [11C]Ketanserin [94]. In humans, a too rapid metabolization along with nonspecific binding rapidly stopped the development of this molecule as a 5-HT2R radiotracer. However, these first trials were the starting point for more specific studies of 5-HT2AR in PET imaging through the development of ketanserin analogues (Figure 6).
[18F]Setoperone is one of the reference radioligands for 5-HT2AR. It is a very potent antagonist of these receptors (Ki = 0.2 nM) along with relative specificity since in baboons, it also interacts with the dopaminergic D2 receptors present in the striatum [95]. Despite this specificity issue, it has been used regularly as a radiotracer for 5-HT2AR in PET imaging, especially in healthy subjects or subjects with various pathologies such as depression, migraines, or Alzheimer’s disease [96,97,98,99,100]. [18F]Setoperone is not used anymore in human studies due to the discovery of [18F]Altanserin, which has demonstrated a higher potential in animal and human studies.
As a 5-HT2AR antagonist (Ki = 0.3 nM), [18F]Altanserin was first evaluated in rats in 1991 by Lemaire et al., and an important signal in the frontal cortex along with a more moderate one in the striatum were detected [101]. The specificity of the signal was determined by prior blocking of the receptors in the presence of 5-HT2AR ligands (ketanserin, pipampone, and methylsergide) and D2 receptor ligands (sulpiride and halopemide). Subsequently, [18F]Altanserin was tested in healthy volunteers, and good distribution of the radioligand was found despite the presence of nonspecific binding, particularly in the cerebellum [102]. This off-target labeling was later attributed to the in vivo production of radiometabolites [103,104]. A more recent study on 52 healthy volunteers demonstrated the absence of a significant difference in [18F]Altanserin binding between the male and female genders and a significant decrease in 5-HT2AR in the elderly and in patients with high body mass indexes [105]. [18F]Altanserin has also been used to determine the density of 5-HT2AR in subjects with various CNS-related disorders such as anorexia, schizophrenia, obsessive-compulsive disorder, depression, and Alzheimer’s disease [105,106,107,108,109]. In light of all these characteristics, numerous pharmacomodulations of [18F]Altanserin were considered in order to increase the resolution of PET imaging by limiting nonspecific binding and by decreasing the production of lipophilic radioactive metabolites.
In 2001, in order to reduce metabolite production, a deuterated analogue was considered: [18F]Deuteroaltanserin. The main objective was to maintain the pharmacological and pharmacokinetic properties while delaying the metabolization by inserting an isotope of hydrogen: deuterium. Initially, a study was carried out in baboons and did not allow observing any significant difference between the two radioligands (neither for radiolabeling nor metabolization) [110]. Subsequently, a study on a healthy man was conducted and allowed the observation of good distribution of the radiotracer as well as a modest decrease in the production of radiometabolites in the plasma [111,112]. In 2009, Santhosh et al. carried out a study with [18F]Deuteroaltanserin on nine subjects affected by Alzheimer’s disease and concluded that there was a significant reduction in the binding of the radiotracer to 5-HT2AR, mainly in the anterior, frontal, medial, parietal, and temporal cingulate cortex [113]. Despite these promising results, no study has referred to [18F]Deuteroaltanserin since then.
3.2. Piperidin-4-Ylmethanol Series: Volinanserin Derivatives
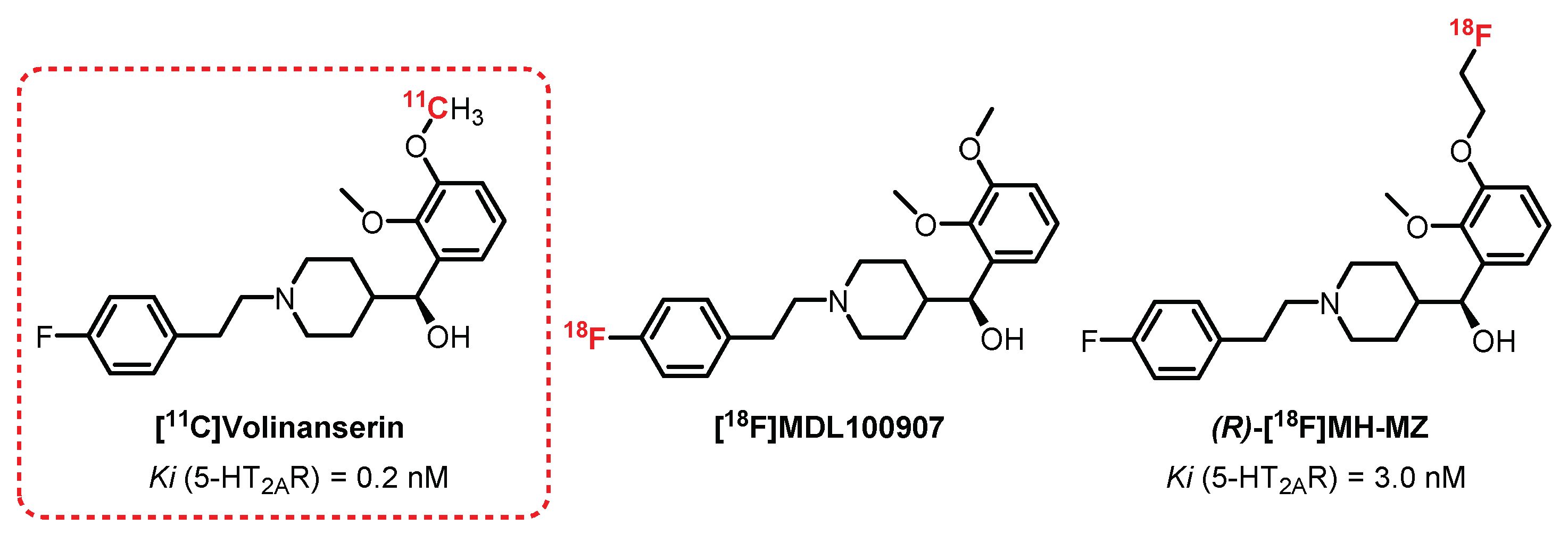
[11C]Volinanserin, or [11C]MDL100907 or [11C]MDL100,907 (Figure 7), is a very selective and potent 5-HT2AR antagonist (Ki = 0.2 nM) [114]. Its interesting pharmacokinetic properties, such as its moderate lipophilicity (logP = 2.7), allowed [11C]Volinanserin to become a reference radiotracer in the study of the distribution of 5-HT2AR. First, its tritiated analogue [3H]MDL10090 has been used in animals (rats and monkeys) to assess its strong specificity, allowing the visualization, quantification, and direct characterization of these receptors by autoradiography [115,116]. The monkey studies were enlarged to the use of [11C]Volinanserin in PET imaging by Lundkvist et al. [117] and permitted the observation of specific labeling of the frontal, temporal, and visual cortex through competition with ketanserin. In 1998, Ito et al. confirmed these results in healthy humans (three volunteers), where a specific labeling of the neocortex (occipital, frontal, and temporal cortex) with moderate nonspecific labeling was observed [114]. In 2012, a comparative study on a large cohort of healthy volunteers consolidated the interest in [11C]Volinanserin since, in addition to obtaining good distribution of labeling of 5-HT2AR receptors, a decline in this population with age for 5-HT2AR expression was evidenced. In addition, the analyses carried out demonstrated a slow metabolization of the radiotracer limiting the presence of radiometabolites, the main drawback encountered with ketanserin derivatives [118]. [11C]Volinanserin represents a reference 11C-radiotracer for the study of 5-HT2AR receptors and has been used in numerous studies on humans and animals in order to observe the influence of 5-HT2AR in various pathologies, such as depression, Huntington’s disease, Asperger’s syndrome, as well as in obsessive-compulsive disorders [119,120,121]. Interesting fluorinated analogues of Volinanserin have also been evaluated recently. [18F]MDL100907 is a Volinanserin analogue labeled with fluorine-18 instead of carbon-11, and it was considered mainly for the benefit represented by the longer half-life of fluorine-18 compared with carbon-11. Few publications refer to this radiotracer, mainly because of the difficulty in achieving an efficient radiofluorination of Volinanserin [122]. In 2014, a study carried out by Ren et al. identified a new method for obtaining [18F]MDL100907 using a nickel-mediated fluorination strategy. In nonhuman primates, a comparison of binding results between [18F]MDL100907 and [11C]Volinanserin concluded that [18F]MDL100907 was able to provide a better 5HT2AR distribution and density through PET imaging [123]. (R)-[18F]MH-MZ is an ethylfluorinated analogue of [11C]Volinanserin which exhibits very good affinity and specificity for 5-HT2AR (Ki = 0.72 nM), along with easier radiosynthetic access compared with [18F]MDL100907. In rats, the (R)-[18F]MH-MZ enantiomer showed the best radiolabeling results, as the affinity of the racemic mixture [18F]MH-MZ was quite lower (Ki = 3.0 nM) [124,125]. The comparative study between [18F]MH-MZ and [18F]Altanserin demonstrated a more specific labeling of areas rich in 5-HT2AR by [18F]MH-MZ. In addition, it revealed a low level of radiometabolites in the cortex extracts compared with the plasma levels, thus confirming that the latter cannot pass the BBB [126]. Despite these encouraging results, to date, no publication refers to the use of this radiotracer in humans.
3.3. N-Benzylphenethylamine Series: CIMBI-5 Derivatives
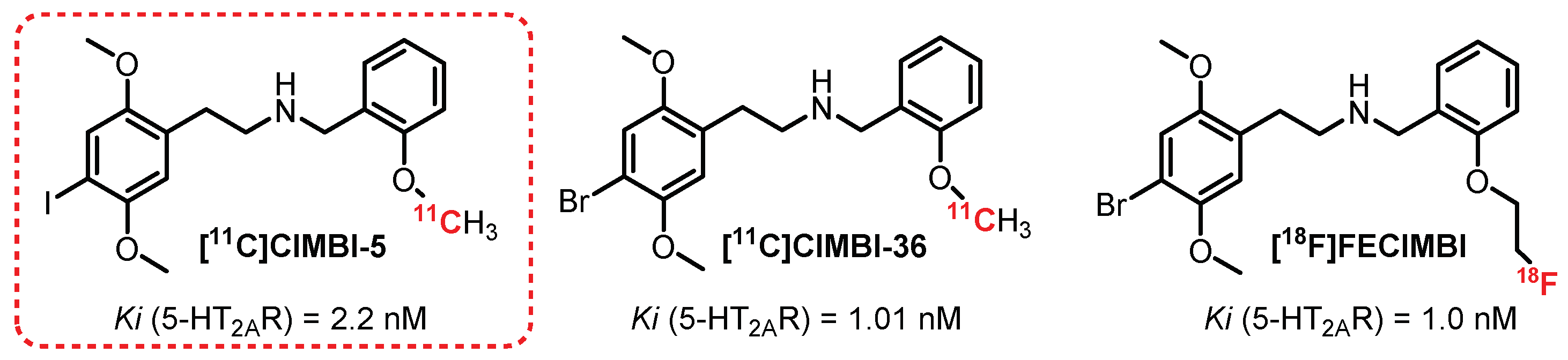
The vast majority of radiotracers synthesized to radiolabel 5-HT2AR are antagonists of these receptors. However, the development of a series of agonist compounds constitutes a major interest, allowing for distinguishing the functional receptors involved in various brain pathologies. It is in this context that [11C]CIMBI-5 was synthesized (Figure 8). In 2010, Ettrup et al. carried out the first studies on this compound, which revealed that it was indeed a potent 5-HT2AR agonist with moderate specificity for these receptors (Ki (5-HT2AR) = 2.2 nM; Ki (5-HT2BR) = 2,3 nM; Ki (5-HT2CR) = 7.0 nM) [127]. The ex vivo study carried out on rat brains showed absorption of the radiotracer mainly in the frontal cortex equivalent to that obtained with [18F]Altanserin. This binding is specific to 5-HT2AR, as proven by a significant displacement of the radiotracer in the presence of ketanserin. In vivo (pigs) imaging revealed significant cortical absorption as well as more moderate absorption in the thalamic and striatal regions, thus evidencing the interest in evaluating [11C]CIMBI-5 in humans. In 2019, additional tests carried out on nonhuman primates (two baboons and two vervet monkeys) allowed observing a radiolabeling in correlation with those obtained previously but with a lower intensity compared with [11C]Volinanserin [128].
Many derivatives of [11C]CIMBI-5 were synthesized and tested, with widely varying radiolabeling results [129]. Among them, [11C]CIMBI-36, the brominated analogue of [11C]CIMBI-5, demonstrated the most interesting results. [11C]CIMBI-36 possesses a strong affinity and specificity toward 5-HT2AR (Ki = 1.01 nM) and afforded an intense radiolabeling of the cerebral cortex in pigs [129]. In 2014, [11C]CIMBI-36 was evaluated against a cohort of 29 healthy volunteers, and good distribution of the radiotracer was demonstrated, particularly in the neocortex and the sensory cortex. The study suggested a weak plasmatic stability of [11C]CIMBI-36, with a 50% decrease in plasma 10 min after injection. This metabolism issue was not evidenced in the pig brain tissue but was the subject of additional research [130,131]. Finally, in the presence of ketanserin, radiolabelling was significantly reduced in the neocortex, confirming the binding selectivity toward 5-HT2AR. Additional studies were carried out on humans and animals with the aim of observing the changes induced by the administration of substances that interact with the serotonergic system [132,133,134]. The study on pigs and monkeys revealed a decrease in the uptake of [11C]CIMBI-36 (46%) in the presence of a significant increase in 5-HT. In humans, [11C]CIMBI-36 allowed assessing a decrease in radioemission in areas rich in 5-HT2AR in the presence of D-amphetamine, thus evidencing the interest in this radiotracer as a potential new tool for the evaluation of neuropathologies such as depression [135]. Finally, in 2016, a study by Ettrup et al. revealed a probable binding of [11C]CIMBI-36 to 5-HT2CR detected by radiolabeling of the hippocampus and choroid plexus greater than that in the presence of [18F]Altanserin [136].
[18F]FECIMBI, an ethylfluorinated analogue of [11C]CIMBI-36, exhibited interesting binding properties toward 5-HT2Rs (Ki (5-HT2AR) = 1 nM; Ki (5-HT2CR) = 1.7 nM) and an agonist profile. [18F]FECIMBI was synthesized by Prabhakaran et al. in 2015 and was able to radiolabel the temporal cortex, the hippocampus, and the choroidal plexus (an area rich in 5-HT2CR) by autoradiography [137]. In 2017, a complementary study in monkeys showed insufficient cerebral absorption and an insufficient volume of distribution for the development of an in vivo quantification tool [138].
3.4. Miscellaneous Derivatives as SPECT Imaging Tracers
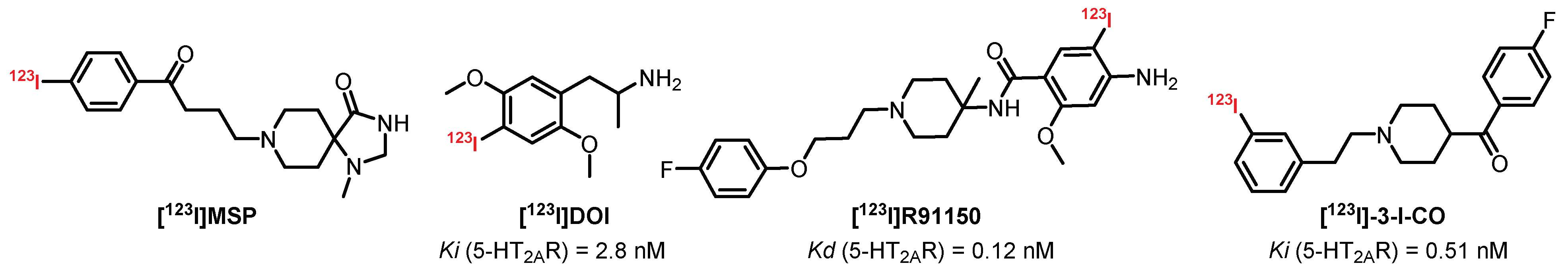
Over the past few decades, many SPECT radiotracers with an affinity for 5-HT2AR have been synthesized (Figure 9). However, due to moderate selectivity, the presence of radiometabolites interfering with imaging and signals that are too weak, very few of them are currently used to quantify 5-HT2AR in humans. Discovered in the 1970s, [123I]DOI is a mixed agonist of 5-HT2AR and 5-HT2CR which allowed the first scintigraphic explorations of the brain even before the emergence of SPECT and PET imaging techniques [139]. [123I]DOI was previously used in its racemic form, in particular in rats (Ki = 2.8 nM) [140]. In 1989, Nazarali et al. published a comparative study highlighting the interest of (R)-[125I]DOI (Kd = 1.2 nM) in its enantiomer (S)-[125I]DOI (Kd = 2.1 nM) [141]. Finally, the results obtained for baboons did not conclude favorably with regard to its use as a specific radiotracer for 5-HT2AR and 5-HT2CR [142].
[123I]R91150 or [123I]R93274 are the most popular radiotracers used to quantify and characterize 5-HT2AR by SPECT imaging. [123I]R91150 is a potent (Kd = 0.12 nM) and selective antagonist of 5-HT2AR which demonstrated its effectiveness through pronounced radiolabeling of the frontal cortex (FC) compared with the cerebellum (CER, nonspecific), with an FC/CER ratio greater than 10 in rats (in vivo) [143]. In 1997, Abi-Dargham et al. published the first complete preclinical study carried out on baboons and observed a specific radiolabeling of the occipital and temporal cortex (reversible in the presence of ketanserin) with a lower FC/CER ratio (1.5). The plasma study revealed a rapid metabolization of [123I]R91150 (75% metabolized 3 h after the injection) with the presence of a less lipophilic metabolic fraction which did not cross the BBB [144]. These observations were confirmed by studies carried out on healthy volunteers who did not question the usefulness of this radiotracer for the quantification and characterization of cerebral 5-HT2AR [145,146]. In 1998, [123I]R91150 was tested on a larger cohort of healthy volunteers (13 women and 13 men) and confirmed the results obtained with [18F]Altanserin (i.e., the absence of a significant difference between genders and the significant decrease in 5-HT2AR with aging) [147]. During the last two decades, [123I]R91150 was used on numerous occasions to study various neuropathologies such as Alzheimer’s disease, Parkinson’s disease, or schizophrenia [148,149,150]. [123I]R91150 became a reference for evaluating the distribution and the quantification of 5-HT2AR in subjects (humans and animals) presenting neuropsychiatric disorders (with or without treatment) such as depression, anxiety, anorexia, or having suicidal behaviors [151,152,153,154,155,156,157]. In 2009, a fluorinated analogue usable in PET imaging was synthesized: [18F]R91150. Despite encouraging results, the difficulty of obtaining the radiotracer did not encourage its development for further studies [158].
[123I]MSP is an iodinated derivative of spiperone, an atypical antipsychotic drug belonging to the butyrophenone family used as a treatment for schizophrenia. [123I]MSP was designed to maintain the affinity for 5-HT2AR of spiperone while decreasing its affinity toward dopaminergic D2 and D3 receptors. The study carried out with [123I]MSP evidenced a strong affinity and selectivity for 5-HT2AR, with a radiolabeling distribution in correlation with those obtained with the reference radiotracers (intense labeling of the mouse frontal cortex) [159]. Despite promising results, [123I]MSP has not been the subject of recent studies.
[123I]-3-I-CO is a potent (Ki = 0.51 nM) and selective antagonist of 5-HT2AR which was developed by Fu et al. in 2002. In 2008, a study on rodents showed good distribution of the radiotracer with a moderate intensity and poorly specific signal. In addition, [123I]-3-I-CO was the target of efflux mechanisms at the BBB level, demonstrated by a significant increase in the intensity of the radiosignal after prior administration of a P-glycoprotein inhibitor (cyclosporine A) [160]. The potential of this radioligand was therefore limited for clinical development and was simply abandoned.
4. 5-HT3 Receptors
Among the 14 serotonin receptor subtypes, 5-HT3R is the only ionotropic receptor which belongs to the pentameric ligand-gated ion channel (LGIC) superfamily [161]. In humans, there are five 5-HT3R subtypes, from 5-HT3A to 5-HT3E. The subunits are encoded by the HTR3 genes located on chromosome 11q23 (HTR3A and HTR3B) and 3q27 (HTR3C, HTR3D, and HTR3E) [162]. 5-HT3R is found in both the CNS and peripheral nervous system. It is mainly located in the dorsal vagal complex of the brainstem, regions involved in the vomiting reflex, and in low levels in many forebrain areas, including the hippocampus, amygdala, nucleus accumbens, putamen, and caudate nucleus [163,164]. The 5-HT3Rs are involved in many physiological processes and neuropsychiatric disorders such as schizophrenia, depression, bulimia, anxiety, pain, autism, bipolarity, and learning and memory disorders [165,166,167]. A class of high-affinity 5-HT3R antagonists named “setrons” has been established as antiemetic drugs, resulting from chemo- and radiotherapy and general anesthesia [166,168]. Moreover, many selective agonists and antagonists are described, thereby facilitating the development of promising radioprobes. However, to date, no radiotracers have demonstrated useful applications in humans.
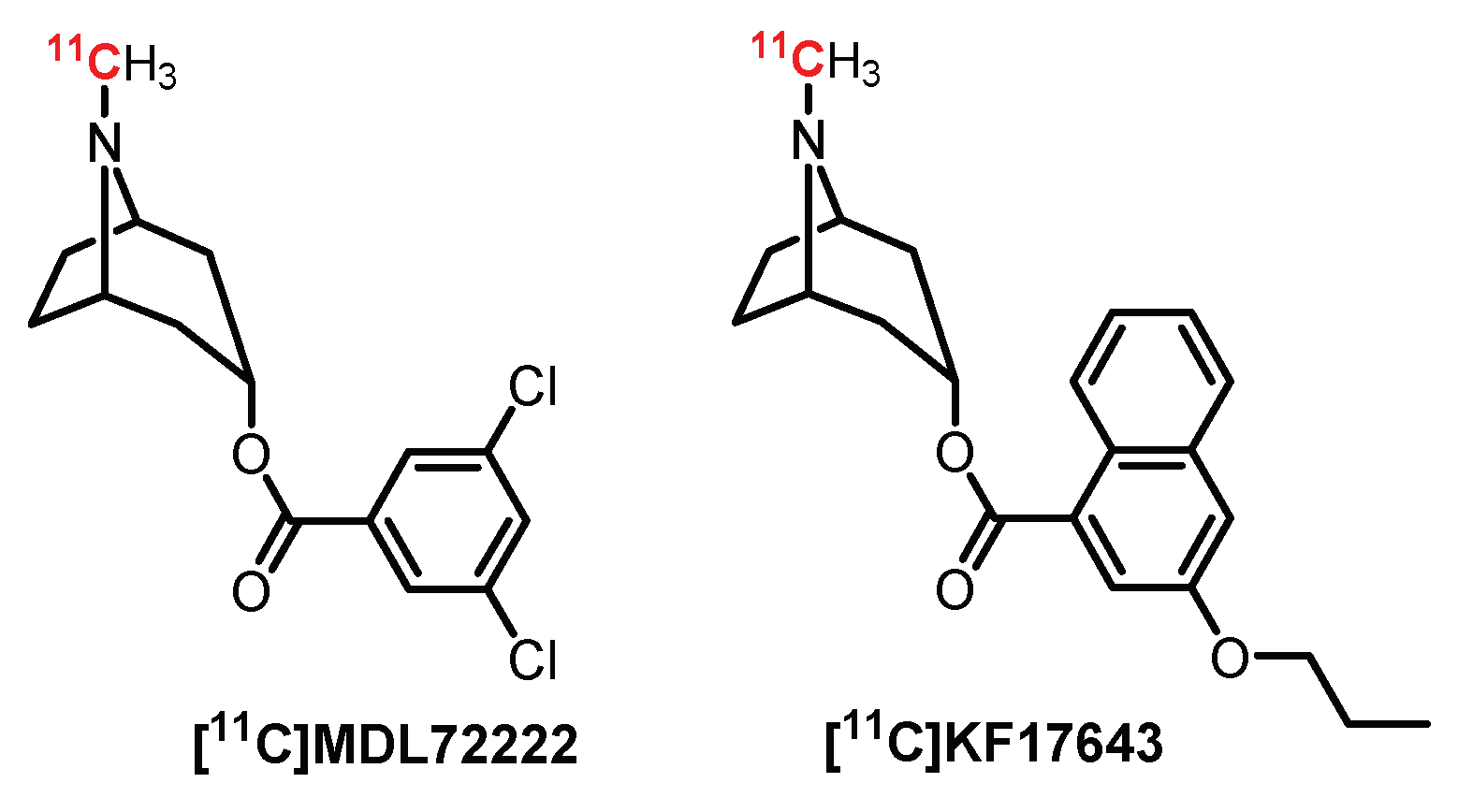
4.1. Tropane Series: MDL7222 Derivatives
First, several antagonists labeled with carbon-11 (Figure 10) were designed as potential PET radiotracers [169]. Thus, MDL72222, a selective 5-HT3R antagonist, was radiolabeled with carbon-11 and further evaluated in rats and baboons. Despite [11C]MDL72222 quickly crossing the BBB in preliminary studies, a heterogeneous diffusion in the brain was observed, probably due to its high lipophilicity and low specificity [170]. A closely related analogue of MDL72222 was radiolabeled by Ishiwata et al. for PET imaging: [11C]KF17643. Nevertheless, [11C]KF17643 displayed nonspecific binding [171].
4.2. Pyrrolo[1,2-a]Pyrazine Series: S21007 Derivatives
S21007 is a 5-HT3R partial agonist (Figure 11) which has also been evaluated as a potential PET ligand in rats and baboons because of its strong affinity (Ki = 1.4 nM), selectivity, and high brain uptake. However, [11C]S21007 was unable to detect the 5-HT3R via a specific binding signal, as [11C]S21007 did not displace unlabeled S21007 in the brain areas with 5-HT3 receptors, suggesting a lack of specific binding [172]. This series was further investigated with the synthesis of [18F]MR18445, another pyrrolo[1,2-a]pyrazine derivative radiolabeled with fluorine-18. Autoradiography and PET studies showed rapid uptake in both rat and baboon brains, but no specific binding in 5-HT3R-rich brain regions was detected with [18F]MR18445, probably due to its high lipophilicity [173]. All these compounds were not successfully used for selective binding to 5-HT3R in vivo.
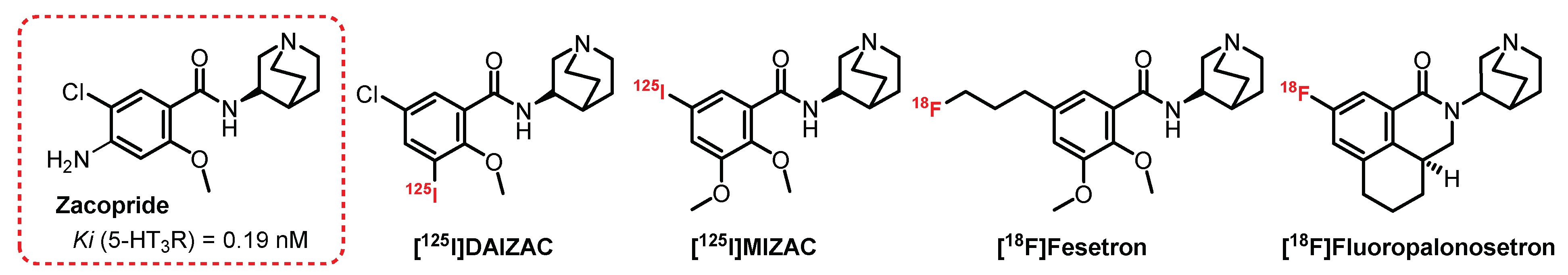
4.3. (S)-Quinuclidin-3-Amine Series: Zacopride Derivatives
Zacopride, a 3-aminoquinuclidinyl derivative, is a well-known mixed 5-HT3R antagonist and 5-HT4R agonist [174]. A series of Zacopride derivatives was developed by Ebert et al. (Figure 12) [175,176]. [125I]DAIZAC exhibited high affinity (Ki = 0.19 nM) and enhanced selectivity for the 5-HT3R. Autoradiographic studies also demonstrated the usefulness of this ligand, but to date, despite these promising characteristics, the ligand has not yet been further developed for SPECT imaging [176]. Highly selective and potent antagonists, such as granisetron and palonosetron, belong to a family of drugs (the “setrons”) that is well established for clinical use, especially because of their effects on Chemotherapy-Induced Nausea and Vomiting (CINV). Among them, [18F]Fesetron was designed based on the model of [125I]MIZAC, a potent and selective radioligand of 5-HT3R [175]. [18F]Fesetron was shown to display a binding profile to rat brain regions known to contain a high density of 5-HT3R. However, the in vivo rat brain uptake of [18F]Fesetron was low, and a significant amount of activity was unable to cross the BBB. This lack of brain permeability, which could be explained by low lipophilicity, limits the usefulness of [18F]Fesetron as a PET radiotracer in this animal model [177].
The second “setron” drug developed in 2016 was a fluorine analogue of the clinically used palonosetron which also contained the 3-aminoquinuclidinyl moiety. In vitro autoradiography showed specific labeling in rodent brain slices, but in vivo PET studies in rats with [18F]Fluoropalonosetron gave low and homogeneous radiotracer signals [178]. This could be related to the low expression of 5-HT3R in the brain combined with the low lipophilicity of the radioligand and could explain why [18F]Fluoropalonosetron is not suitable for in vivo PET imaging of 5-HT3 receptors.
4.4. Miscellaneous Derivatives as 5-HT3R Radiotracers
In 1995, [11C]YM060 was labeled with carbon-11 through N-methylation of indole and revealed poor brain uptake in mice (Figure 13) [179]. N-methylquipazine (NMQ, (2-[1-(4-methyl)-piperazinyl]quinoline) is an agonist with good affinity and selectivity for 5-HT3R (IC50 = 4.7 nM). Its biodistribution has been evaluated in vivo in rats and nonhuman primates using PET imaging [180]. The imaging results indicate that its uptake was observed in structures known to contain 5-HT3R, although the radiotracer also interacted with non-5-HT3R sites. In 2008, Gao et al. designed a new carbon-11-labeled benzisoxazole derivative for PET imaging of 5-HT3 receptors [181]. These partial agonists exhibited high binding affinity and could be promising for future in vivo biological evaluation.
5. 5-HT4 Receptors
5-HT4R receptors were first described by Dumuis et al. in 1988 [182]. They are GPCRs positively coupled to adenylyl cyclase and induce cyclic adenosine monophosphate (cAMP) production [183]. 5-HT4Rs are located in the peripheral nervous system (PNS), mainly being found in the heart, gastrointestinal tract, and enteric nervous system [184], as well as in the CNS, where they are highly expressed in different brain regions like the hippocampus, amygdala, and cerebral cortex, suggesting the involvement of the receptor in brain physiological functions [185] such as learning and memory [186,187,188], eating disorders [189], and mood behavior [190]. In order to further specify in vivo the 5-HT4R physiological or pathological role, selective radioligands able to target 5-HT4R have been developed for SPECT and PET applications.
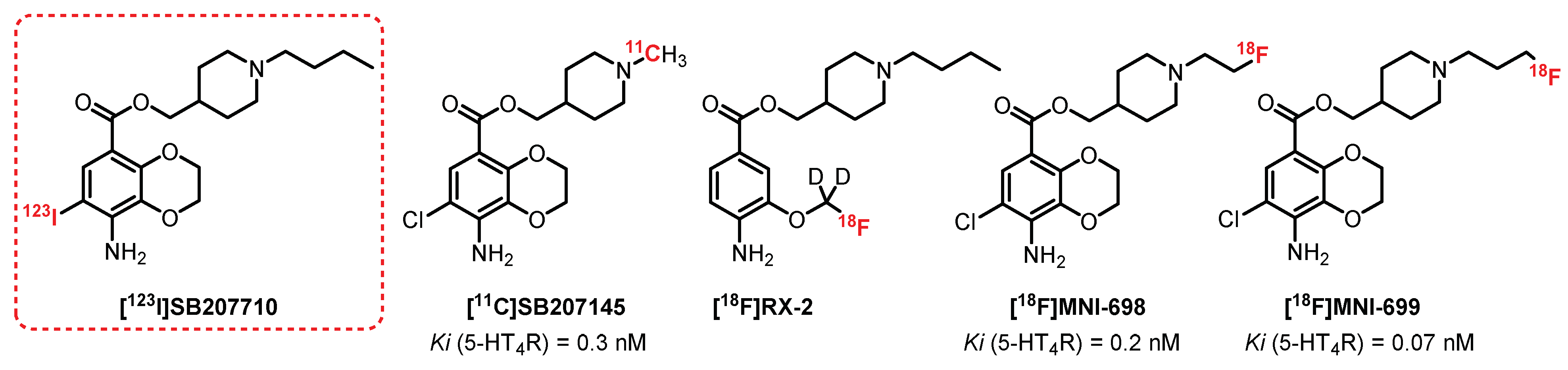
5.1. Benzodioxane Series: SB207710 Derivatives
Kaumann et al. [191] reported on SB207710, a selective high-affinity benzodioxane antagonist for 5-HT4R (Figure 14). Autoradiographic experiments on humans [192] and on rat brains with its radiolabeled analogue, [125I]SB207710, confirmed its potency and selectivity [193] and led Pike et al. to characterize its efficacy in vivo as a SPECT radioligand [194]. A good brain uptake in rats was observed for [125I]SB207710, and the study revealed specific radiolabeling of regions rich in 5-HT4R (striatum, frontal, and temporal cortex). [123I]SB207710 was then examined by SPECT imaging in nonhuman primates. The radiotracer readily accumulated in 5-HT4R-rich areas. The specificity of this radiolabeling was underlined through pretreatment with a selective 5-HT4R antagonist, SB204070, which reduced the radioactivity in all brain regions. Even if [123I]SB207710 entered the brain, its signal quickly disappeared in SPECT, thus proving rapid clearance. Since then, no further investigations for SPECT studies in humans have been reported. Nevertheless, the use of [123I]SB207710 in SPECT imaging provided the first demonstration of 5-HT4R imaging in primate brains in vivo [194]. The antagonist SB207145 (Ki = 0.3 nM), structurally related to SB207710, was radiolabeled with carbon-11 by Gee et al. in 2008 [195], and its potential as a PET radioligand was evaluated for 5-HT4R imaging. The preliminary data in a few healthy subjects indicated that [11C]SB207145 was distributed in the brain according to the known 5-HT4R distribution. It was then successfully used for in vivo studies in animals to evaluate the metabolism and binding kinetics in a minipig brain [196] and human subjects [195,197,198,199]. To date, [11C]SB207145 remains the only 5-HT4R radiotracer that has been evaluated in human studies [200,201,202,203]. However, the short half-life of the radioisotope (t1/2 = 20.4 min) significantly restricted its use for more advanced clinical trials.
In 2010, Xu et al. reported the first fluorine-18-radiolabeled 5-HT4R radiotracer [18F]RX-2 [204] in which deuterium was incorporated to provide greater in vivo resistance to defluorination [205]. PET studies with [18F]RX-2 were performed on monkeys in 2014 and showed a high brain uptake, quantifiable distribution, and insignificant radiometabolite detection [206]. The mild but stable 5-HT4R-specific signal validated the potential use of [18F]RX-2 for mapping 5-HT4R, but no transposition to human PET studies were published to date.
More recently, [18F]MNI-698 and [18F]MNI-699, two fluorinated analogues of SB207145 containing fluorine-18 in the terminal piperidine alkyl chain, were described [207]. These radioligands have been shown to be affine with 5-HT4R (Ki = 0.2 nM and 0.07 nM, respectively). In vivo PET studies on adult rhesus monkeys established that both radioligands displayed suitable brain penetration and brain distribution consistent with the known 5HT4R densities. The whole-body biodistribution and dosimetry of [18F]MNI-698 were then investigated and supported that this radioligand could be used for investigation in humans [208,209]. All these ligands belong to the same family of compounds and contain an ester function which can generate short metabolic stability. Other chemical series deprived of ester functions have been explored recently, such as azaphenanthrene derivatives [210].
5.2. 5-HT5 Receptors
Two subtypes of 5-HT5R are known—5-HT5AR and 5-HT5BR—but only 5-HT5AR is expressed in the human brain. Because of the lack of a selective and potent ligand described for this receptor to date, no potent radiotracer or even radioligand has been described.
5.3. 5-HT6 Receptors
The 5-HT6 receptor is one of the most recently identified members of the 5-HT receptor family. First cloned in a rat, the 5-HT6R gene encodes a 438-aminoacid protein [211,212] which is well-conserved in humans [213], and 5-HT6R is a Gαs-coupled receptor (GPCR) that is almost exclusively expressed in the CNS and is abundant in the brain regions involved in cognitive functions, such as the prefrontal cortex, hippocampus, striatum, and nucleus accubens [214,215,216]. Similar distribution patterns between rats, nonhuman primates, and humans have been established [217,218]. In addition to the adenylyl cyclase signaling pathway [219], 5-HT6R has been linked to cellular signaling cascades involved in cognitive processes and neurogenesis, such as the mammalian target of the rapamycin (mTOR) pathway [220]. Thus, 5-HT6R has emerged as a promising target for cognitive enhancement in neurodegenerative or in psychiatric diseases and for antiobesity drugs [187,221,222,223,224,225].
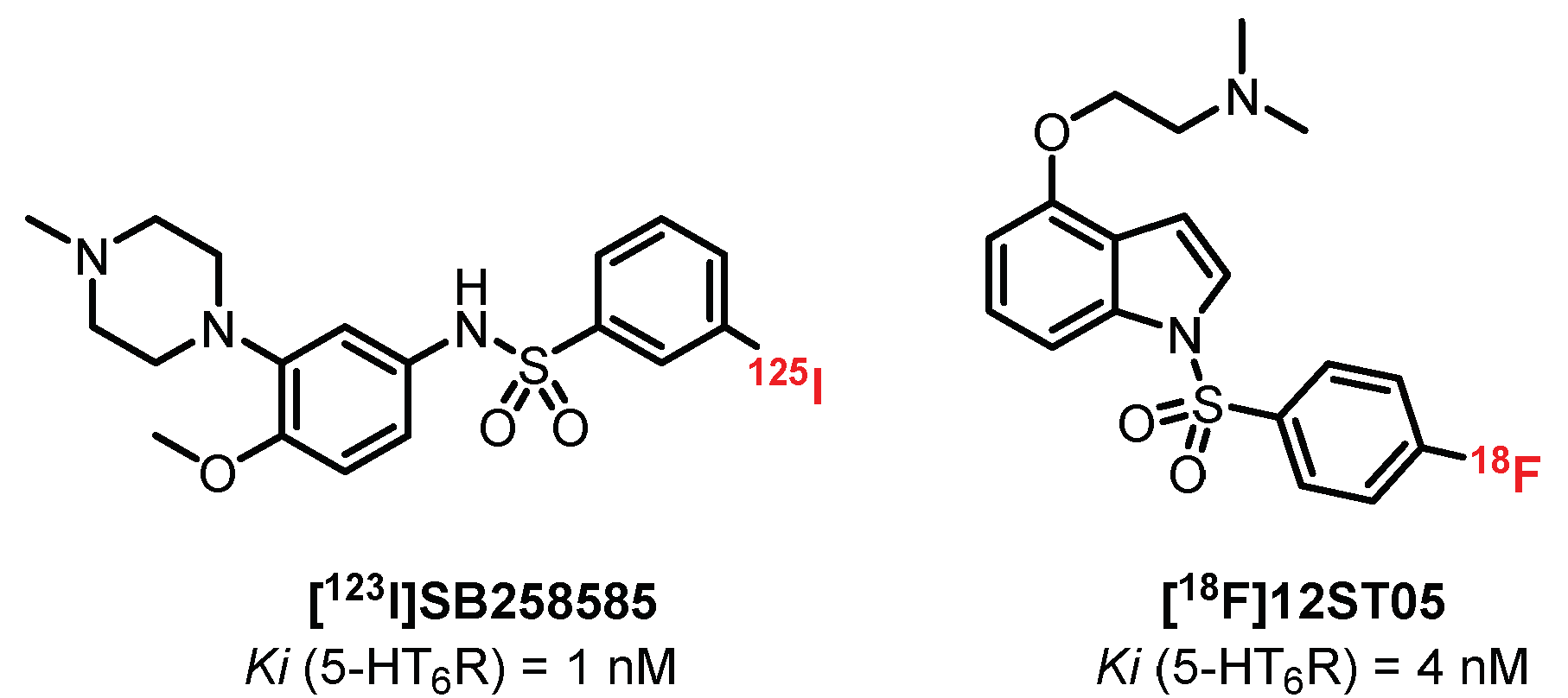
5.4. Benzene Sulfonamide Derivatives: SB258585 Derivatives
Selective radioligands for in vitro autoradiographic studies are available, such as [125I]SB258585 (Ki = 1 nM), but their development as in vivo imaging tools was restricted due to poor brain uptake (Figure 15) [226]. Tang et al. [227] developed a series of 4-(2-aminoethoxy)-N-(phenylsulfonyl)indoles based on the selective 5-HT6R antagonist SB271046 [228]. This study led to the design of [18F]12ST05, presenting good affinity for 5-HT6R (Ki = 4 nM). Ex vivo autoradiographic studies in rat brain slices established high labeling in regions known to contain 5-HT6R. In vivo studies in cats failed to show the specific binding of [18F]12ST05 to 5-HT6R in the cerebral regions, and hence, further investigations were not undertaken for [18F]12ST05 as a PET radioligand.
5.5. 5-(Piperazin-1-yl)Quinolone Derivatives: GSK215083 Derivatives
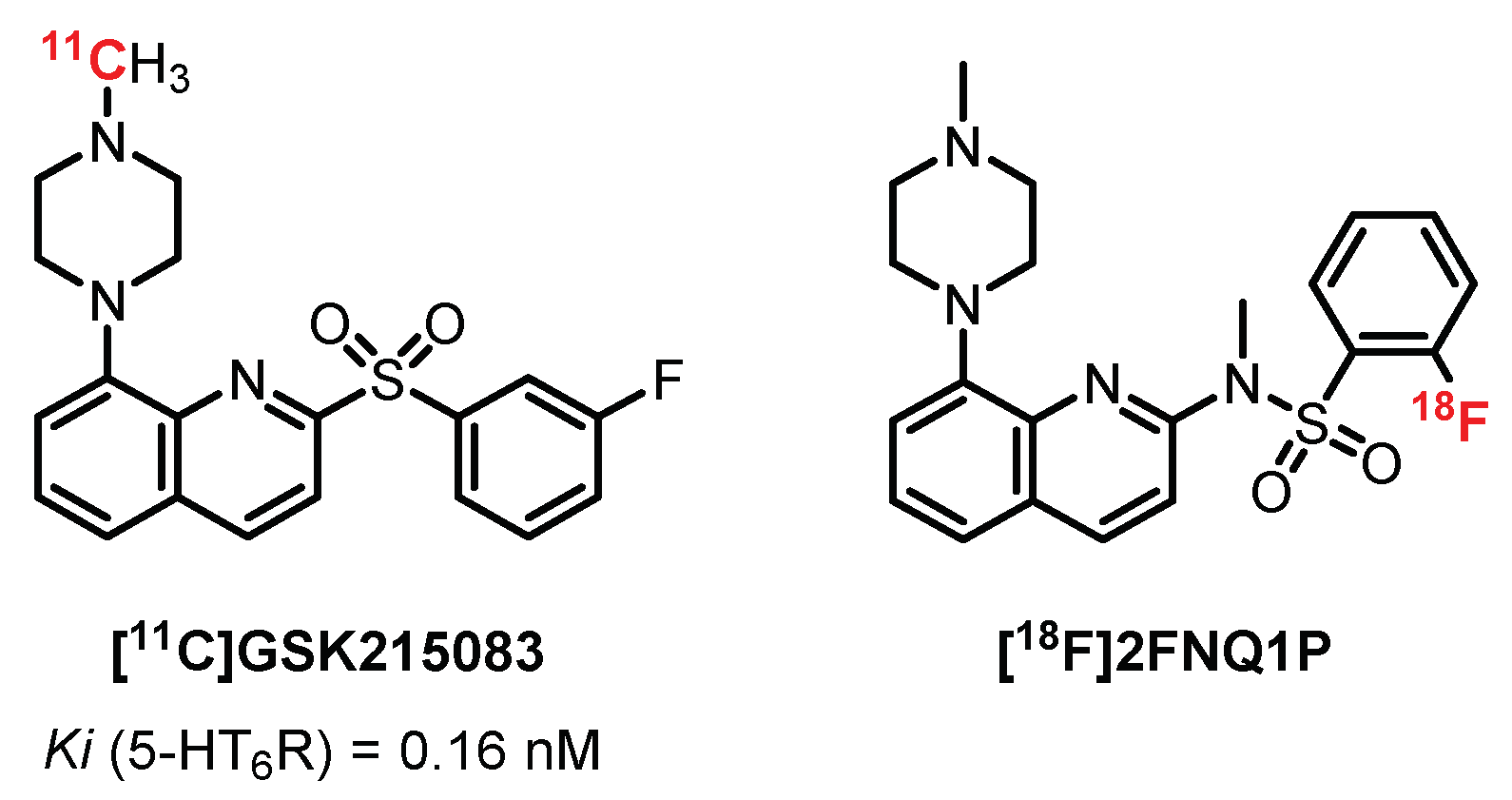
In 2012, Parker et al. published an interesting study presenting the in vivo evaluation of [11C]GSK215083 (Ki (5-HT6R) = 0.16 nM and Ki (5-HT2AR) = 0.79 nM) in pigs, nonhuman primates, and humans (4 healty volunteers) (Figure 16). They revealed a significant binding of the radiotracer on the striatum and frontal cortex in correlation with the autoradiograms carried out previously on cats. Two additional studies confirmed the interest shown in this radiotracer and revealed a decrease in 5-HT6R with age. Moreover, its mixed action (5-HT6R and 5-HT2AR) was effective in the study of neuropsychiatric diseases such as AD and schizophrenia [229,230].
In 2014, Rosse designed radiolabeled quinolone derivatives structurally related to GSK215083 as 5-HT6R PET ligands [231]. In 2014, Colomb et al. developed a new chemical series of compounds inspired by GSK215083 in order to maintain a strong affinity toward 5-HT6R and to optimize its specificity. The bioisosteric replacement of the sulfonyl part by sulfonamide led to a new ligand, 2FNQ1P, with high affinity and selectivity for 5-HT6 receptors [232]. The distribution of [18F]2FNQ1P in 5-HT6R-dense regions (i.e., the cortex and striatum) was assessed by semiquantitative autoradiography in rat brains. Furthermore, the addition of SB2258585 (a specific 5-HT6R antagonist) displaced the binding of [18F]2FNQ1P in a concentration-dependent manner, confirming its excellent specificity for 5-HT6R [233]. The cerebral distribution of [18F]2FNQ1P was then studied in vivo in various animal models, from rodents to nonhuman primates. The in vivo evaluation in rats using microPET exhibited reduced brain uptake of [18F]2FNQ1P. By contrast, in vivo PET imaging with [18F]2FNQ1P in feline and primate models showed high uptake and distribution in the striatal regions particularly rich in 5-HT6R, suggesting that [18F]2FNQ1P could be a suitable 5-HT6R PET tracer for quantification of the brain 5-HT6R in primates. A recent in vitro study demonstrates a decrease in the caudate nucleus 5-HT6R density with [18F]2FNQ1P in human brain tissue from patients with different stages of Alzheimer’s disease [233]. Future in vivo exploration should be carried out to establish the relevance of [18F]2FNQ1P to confirm an early decline in 5-HT6R expression during the progression of the disease.
5.6. 5-HT7 Receptors
In 1993, 5-HT7R was independently cloned and discovered by several scientific teams in different species including mice, rats, and humans [212,234,235]. In human, the protein is encoded by the HTR7 gene located on the long arm of chromosome 10 (10q23.31), comprising 3 introns involved in alternative splicing phenomena. Splicing of intron 2 and 3 is responsible for the existence of 3 distinct isoforms—5-HT7A, 5-HT7B and 5-HT7D—showing no pharmacological difference. The 5-HT7R receptors are GPCRs (coupled to a Gs protein) that will diffuse nerve impulses through the activation of adenylate cyclase and the production of the secondary messenger: cAMP. In human CNS, 5-HT7Rs are localized mainly in the anterior thalamus and in the dentate gyrus. The hypothalamus, anterior cingulate gyrus, hippocampus, and amygdala constitute other human brain regions containing substantial amounts of 5-HT7Rs [236].
The 5-HT7R receptors are involved in the phenomena of learning and memorization as well as in the regulation of temperature and circadian rhythm. In addition, they play a decisive role in the development of certain pathologies such as schizophrenia, migraines, and Alzheimer’s disease [237]. Among the 14 known serotonergic receptors, 5-HT7R is the most recently discovered and explored one. Its implication in the genesis of certain pathologies such as depression, anxiety, and epilepsy make it a target of choice for the development of curative and diagnostic tools [238]. The development of radiotracers allowing their visualization and quantification has become a major axis of research in medicinal chemistry.
5.7. Oxindoles Series: DR4004 Derivatives
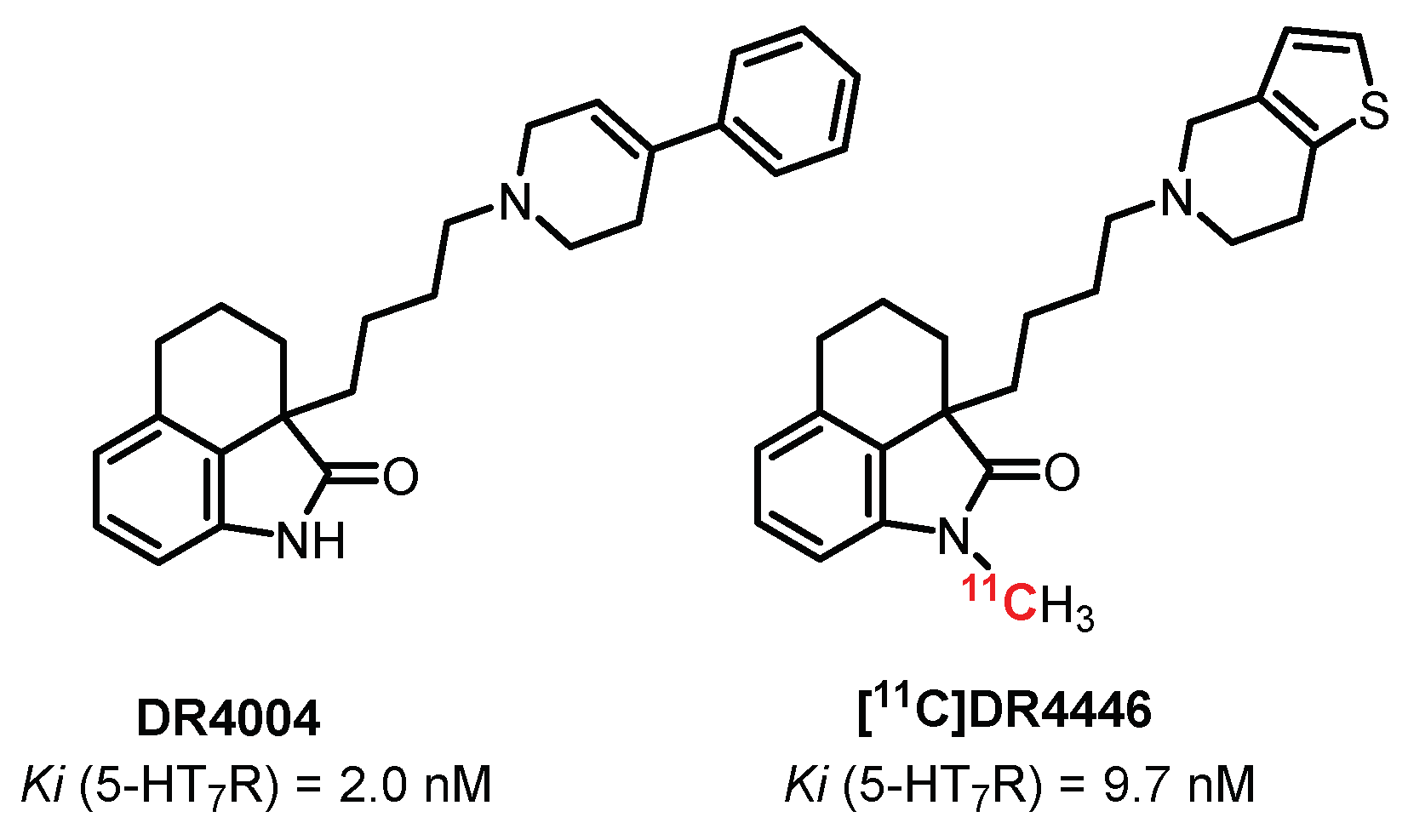
DR4004 (Figure 17) is a selective and potent antagonist of 5-HT7 receptors (Ki = 2 nM) discovered in 1999 by Kikuchi et al. [239]. In 2002, the same group proposed a N-methylated analogue, DR4446, while maintaining the affinity and selectivity toward 5-HT7R (Ki = 9.7 nM) [240]. Radiolabeling through [11C]N-methylation, performed by Zhang et al., allowed obtaining the first imaging study with the 5-HT7R antagonist radiotracer [241]. [11C]DR4446 showed good diffusion through the BBB in vivo (rhesus monkeys), with a high brain uptake especially in the 5-HT7R-rich region: the thalamus. However, this study did not allow a favorable transfer to human experiments due to the impossibility to observe a blocking effect in a self-block study, probably due to off-target labeling of other CNS receptors.
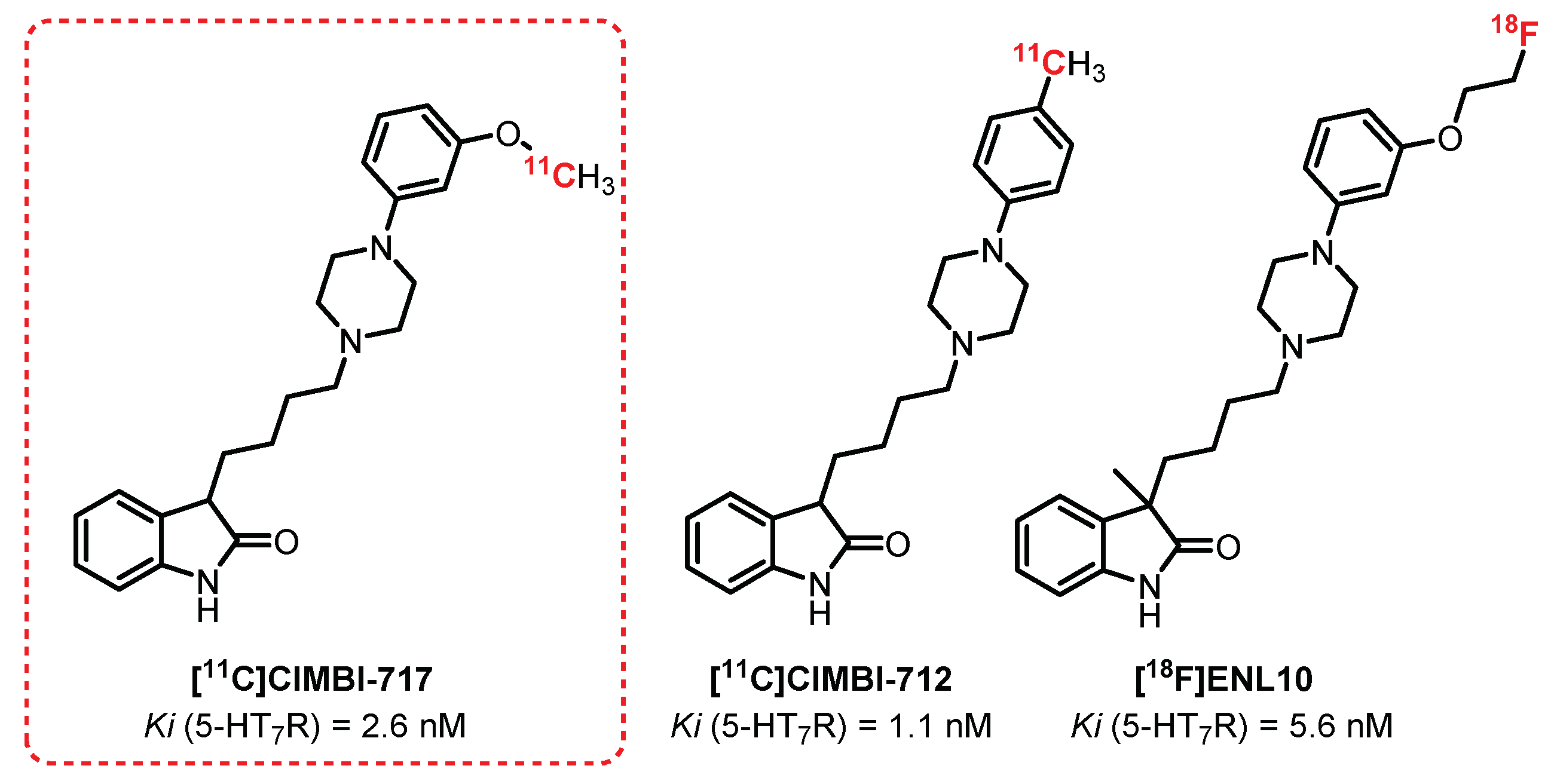
CIMBI-717 and CIMBI-712 (Figure 18) are potent 5-HT7R antagonists (Ki = 2.6 nM and 1.1 nM, respectively) discovered in 2012 by Herth et al. [242]. In 2014, both of these ligands, after radiolabeling with carbon-11, were evaluated in vivo (pigs) as 5-HT7R tracers by Knudsen et al. A high brain uptake was observed for both [11C]CIMBI-712 and [11C]CIMBI-717. Their specificity was demonstrated through blocking studies after the injection of SB-269970, a 5-HT7R antagonist [243]. For [11C]CIMBI-717, a radiolabeling of the thalamus, the striatum, the hippocampus, and the cortex in correlation with the in vitro autoradiographs determined beforehand with [3H]SB-269970 was also observed. Despite these encouraging results, no recent publication mentions its use in humans. However, in the same series, Herth et al. proposed in 2019 a closely related analogue of CIMBI-717 and CIMBI-712: ENL10 [244] while overcoming the problematic short half-life of carbon-11 through fluorine-18 labeling. [18F]ENL10, a potent and selective 5-HT7R antagonist (Ki = 5.6 nM) demonstrated in vivo (rats) a very modest passage across the BBB due to the efflux phenomena induced by a significant expression of the transporter P-gp in rats. In the presence of Elacridar (P-gp inhibitor) and SB-269970, the study revealed specific radiolabeling of 5-HT7R-rich areas, making this compound a serious candidate for possible application of these tests to other species (e.g., pigs or nonhuman primates). This suggestion is motivated mainly by recent observations, indicating a lower expression of P-gp in non-rodents and making the use of Elacridar probably obsolete for these species.
5.8. N-Sulfopyrrolidine Series: SB-269970 Derivatives
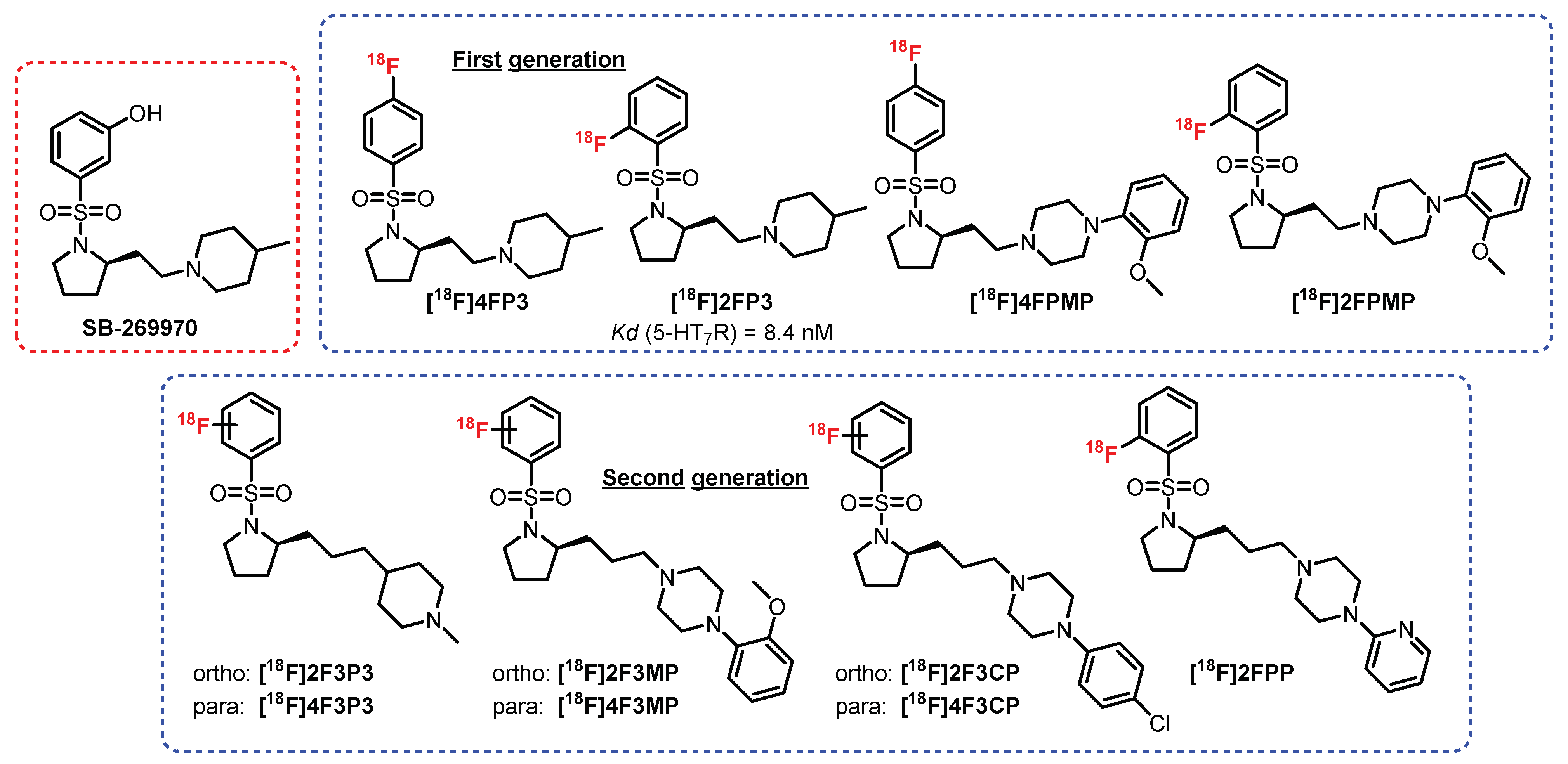
In 2011, Zimmer et al. developed a new chemical series of compounds inspired by the N-sulfopyrrolidine structure from a 5-HT7R antagonist: SB-269970 [243] (Figure 19). Among this first generation of radioligands, [18F]2FP3 proved to be the most promising. Indeed, [18F]4FP3, [18F]4FPMP, and [18F]2FPMP exhibited nonspecific radiolabeling in the presence of a reference antagonist (SB269970) in rats (ex vivo and in vivo). [18F]2FP3 is an affine and selective 5-HT7R antagonist (KD = 8.4 nM) which was previously tested in rats (ex vivo autoradiography) and in cats (in vivo) [243]. The first results were very promising, since selective radiolabeling of the hippocampus, cingulate cortex, and thalamus was observed. In addition, 95% of the collected signal was assigned to [18F]2FP3 and not from a radiometabolite. In 2019, a complementary study on pigs and nonhuman primates was carried out [245]. Partial specificity of [18F]2FP3 for the cerebellum (in monkeys) was revealed, clearly reflecting the difficulties in translating the results from one species to another. In addition, [18F]2FP3 was found to be rapidly metabolized in pigs (50% metabolized 10 min after injection) and more moderately in primates (50% in 60 min). This study did not provide a favorable conclusion for its use in humans. This first approach was further completed by the evaluation of a second generation of radioligands, where additional carbon between the pyrrole and the piperidine or the piperazine was inserted [246]. These radioligands were optimized in terms of radiolabeling specificity for 5-HT7R, but additional pharmacomodulations will be necessary in order to improve the biodistribution and passage of the BBB.
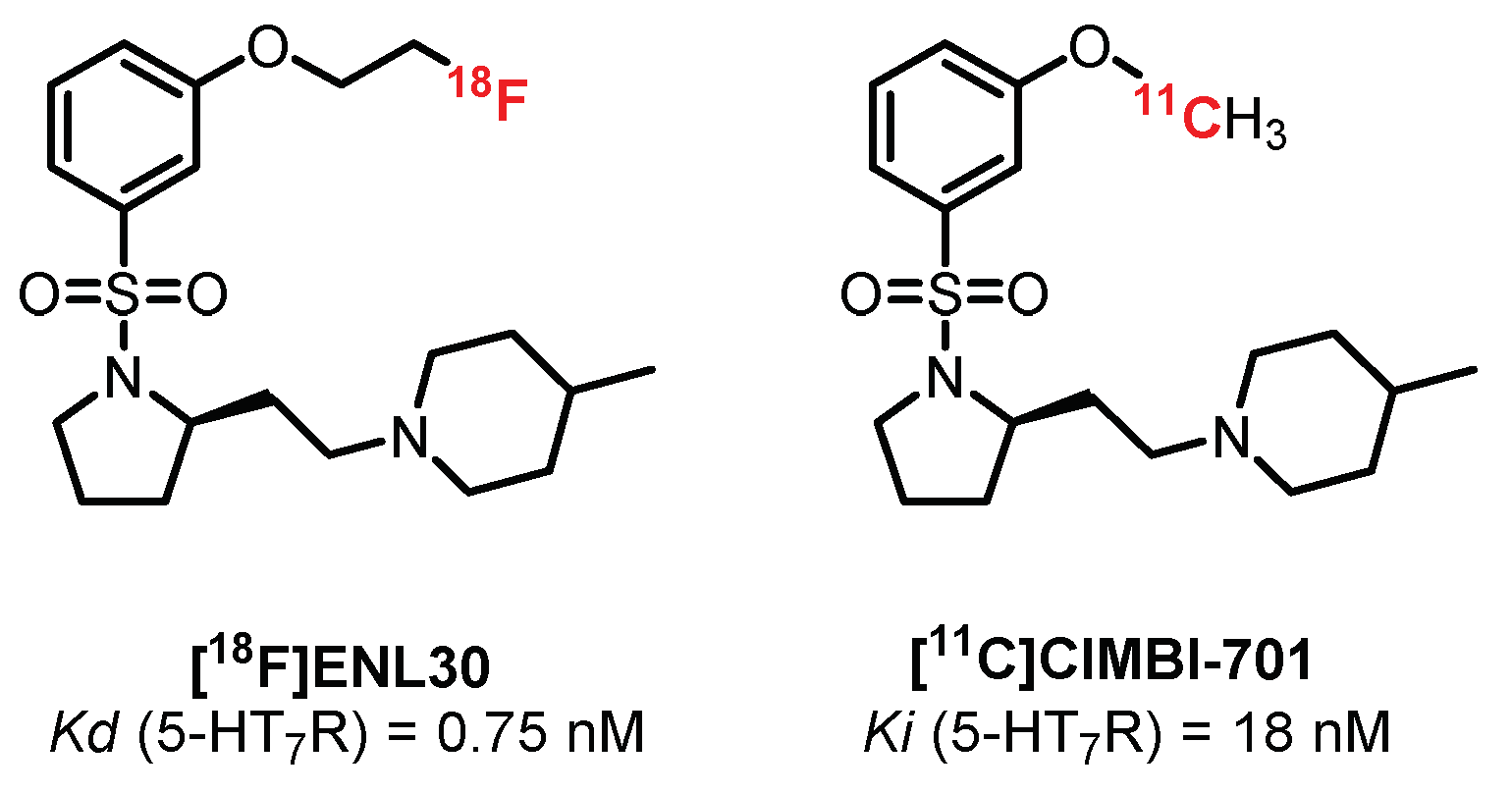
Still in the N-sulfopyrrolidine series, two radioligands were synthesized and tested by Herth et al.: [18F]ENL30 [247] and [11C]CIMBI-701 [248] (Figure 20). These compounds exhibited mixed labeling of the 5-HT7R and σ-receptors and added to a moderate signal in the brain (probably linked to the mechanisms of efflux (P-gp substrate)), thus limiting their use in humans.
6. Serotonin Transporter
A serotonin transporter (SERT) is not a serotonergic receptor per se. However, it is heavily involved in the regulation of 5-HT. The development of radioligands that are affine and selective for these transporters constitutes an interesting axis of development in clinical research.
SERTs are proteins consisting of 12 transmembrane helices whose main role is to regulate the concentration of 5-HT in the inter-synaptic space. The reuptake of 5-HT results from a complex catalytic reaction incorporating both symporter and antiporter transport involving three distinct ions: Na+, Cl−, and K+. This transport takes place in four successive stages [249]. (1) A conformation change of SERTs, resulting in exposure of the binding sites to the intracellular medium, takes place due to the extracellular stoichiometric binding of 5-HT, Na+ ions, and Cl− ions. (2) The 5-HT, Na+, and Cl- ions are released in the intracellular medium. (3) Intracellular binding of K+ ions leads to a conformational modification of the SERT, which regains its original conformation. (4) K+ ions are released into the extracellular medium. SERT expression is tightly regulated by the concentration of 5-HT in the intersynaptic space. In the presence of an increased amount of 5-HT, there is a decrease in the internalization of the transporter, resulting in an increase in its density on the presynaptic membrane. On the contrary, SERTs are downregulated in the presence of a small amount of 5-HT. In this context, a large panel of compounds has been developed with the main objective of blocking the reuptake of serotonin by inhibiting these transporters. The serotonin reuptake inhibitors (SSRI drug class), including fluoxetine (Prozac®), sertraline (Zoloft®), and citalopram (Celexa®), are mainly prescribed to treat major depressive disorders as well as obsessive-compulsive disorders.
During the last three decades, the development of new potent and selective radiotracers of SERTs has become a major axis of development in clinical and preclinical research. Indeed, their expression fluctuations, dependent on the endogenous 5-HT level, are an interesting marker in various pathologies such as depression, bipolarity, and eating disorders. In addition, these studies could allow assessing the impact of regular administration of SSRIs on the serotonergic system, thus allowing a better understanding of their mode of action.
6.1. Isoquinoline Series: McN5652 Derivatives
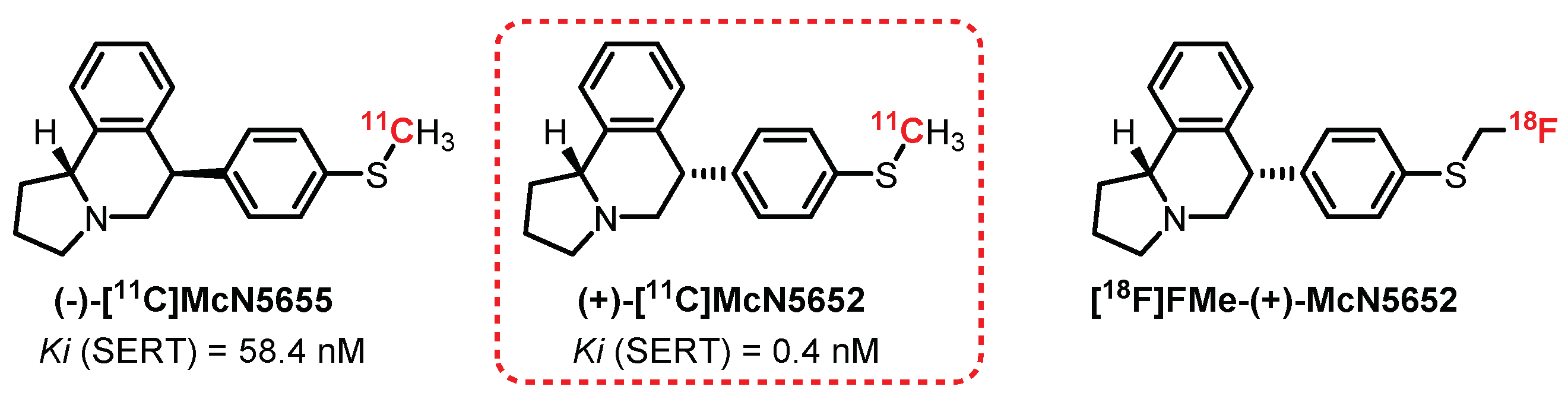
McN5652 is the first SERT ligand evaluated as a radiotracer. The two evaluated diastereomers of McN5652—(+)-[11C]McN5652 and (−)-[11C]McN5655—present significant differences in terms of affinity toward SERTs at Ki of 0.4 nM and 58.4 nM, respectively (Figure 21) [250]. Preliminary studies carried out on mice allowed appreciating the difference in radiolabeling between the two enantiomers, with clearly more interesting results for the (+) enantiomer. In 1995, a study on baboon brains revealed significant absorption of the radiotracer in the midbrain, hypothalamus, thalamus, and striatum. This distribution correlates with the results obtained for brain section autoradiography with [3H]paroxetine [251]. In 2000, Parsey et al. carried out a study on six healthy male volunteers and confirmed the interest in (+)-[11C]McN5652 as a radiotracer, allowing the quantification of SERTs in the limbic, striatal, and thalamic regions. However, the study revealed major limitations to its use, in particular with the presence of a non-selective labeling in the neocortex area and a specific or nonspecific binding ratio (satisfactory after 115 min) not being in adequacy with the use of carbon-11 (t1/2 = 20.4 min) [251,252]. Nevertheless, (+)-[11C]McN5652 has been used on numerous occasions to assess SERT biodistribution in various pathological conditions such as depression and obsessive-compulsive and mood disorders [253,254,255].
In order to improve its properties, a methylfluorinated analogue of (+)-[11C]McN5652 was synthesized: [18F]FMe-(+)-McN5652. Preliminary evaluations in a rat (ex vivo) revealed intense and specific radiolabeling (competitive tests in the presence of fluoxetine, nisoxetine, and GBR12909) of the amygdala, hypothalamus, raphe nuclei, thalamus, and locus coeruleus compared with the cerebellum (nonspecific binding) [256]. A complementary study on a pig (in vivo) allowed concluding favorably regarding the use of this radiotracer as an alternative to (+)-[11C]McN5652 [257]. However, as its affinity for SERTs was found to be more moderate (Ki = 2.3 nM) than (+)-[11C]McN5652, its binding parameters were estimated to be less reliable [258]. Despite promising results, no publication to date mentions the use of this tracer in humans.
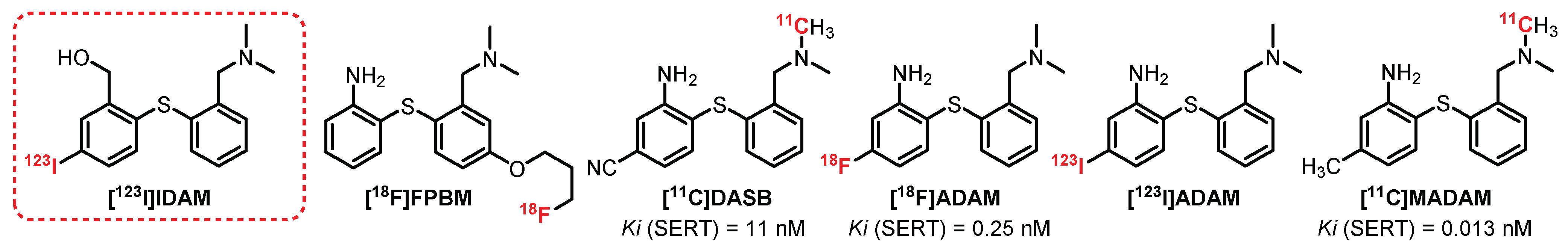
6.2. Diarylthioether Series: [123I]IDAM Derivatives
[11C]DASB is probably the most used radiotracer to explore the distribution of SERTs at the CNS level. This radiotracer was synthesized in the early 2000s as an analogue of [123I]IDAM, the first radiotracer to be synthesized and evaluated in the diarylthioether series [259], but quickly abandoned in favor of its analogues (Figure 22). [11C]DASB is very affine and selective toward SERT (Ki = 1.1 nM) [260]. The first human study (nine healthy volunteers) was performed by Wilson et al. and allowed observing an intense assimilation of [11C]DASB in the midbrain, the thalamus, the hypothalamus, and the striatum [261]. The specificity was confirmed by a prior oral intake of citalopram, decreasing the radioactivity of these cerebral areas to equivalent values to those obtained in the cerebellum (nonspecific labeling). In addition, comparative tests with (+)-[11C]McN5652 indicated a much more interesting specific or nonspecific binding ratio for [11C]DASB [262,263]. Between 2002 and 2004, several studies were carried out on animals (rats, cats, and baboons), allowing for evaluating the general distribution of [11C]DASB, and concluded favorably regarding its use to determine the influence of SERTs in various psychiatric and neurological disorders [264,265,266]. Thus, [11C]DASB was used on numerous occasions in patients presenting various pathologies such as depression, schizophrenia, obsessive-compulsive disorder, Parkinson’s disease, alcoholism, and bipolarity [267,268,269,270,271,272,273]. To date, [11C]DASB constitutes a reference in the quantification and characterization of SERTs at the cerebral level.
In 2001, [11C]MADAM, a methylated analogue of [11C]DASB exhibiting a very strong affinity and specificity for SERTs (Ki = 0.013 nM), was described [274]. The first baboon studies indicated consistent radiolabeling of SERT-rich areas of the brain [275]. In 2005, imaging studies on healthy humans (nine volunteers) confirmed the previous observations, with a major absorption of [11C]MADAM in the raphe nuclei, the putamen, the hippocampus, and more moderately in the frontal and cingulate cortex [276,277]. Finally, [11C]MADAM has been used more recently to study the density of brain SERTs in people with eating disorders and gambling addiction [278].
In 2000, the discovery of [123I]ADAM offered the possibility to perform human imaging exploration of SERTs using SPECT imaging [279]. [123I]ADAM was first tested in mice, rats, rabbits, and nonhuman primates and allowed observing intense and specific radiolabeling of the midbrain [280,281,282,283]. In 2003 and 2004, two preliminary studies evaluated the biodistribution of [123I]ADAM in humans (7 and 11 healthy volunteers) and allowed observing the specific labeling of the midbrain [284,285]. These results were confirmed by several additional studies which demonstrated a specific and reproducible labeling in the midbrain, the thalamus, and the striatum, particularly in people with an SERT polymorphism [286,287,288]. Finally, a double-blind trial was carried out (12 healthy men volunteers) in order to visualize the differences in radiolabeling with an administration of paroxetine versus a placebo [289]. [123I]ADAM allows exploration of the variations of SERTs, particularly in the presence of various pathologies such as depression, Parkinson’s disease, and eating disorders [289,290,291,292,293,294]. In 2003, [18F]ADAM, a fluorinated analogue of [123I]ADAM with good affinity and selectivity toward SERTs (Ki = 4.8 nM), was described [295,296]. [18F]ADAM was first tested on monkeys and allowed observing an intense radiolabeling of the striatum, thalamus, and midbrain. The selectivity was confirmed in the presence of increasing doses of fluoxetine, allowing visualization of the displacement of the radioligand [297]. [18F]ADAM was then further studied in humans with major depressive disorders and allowed the observation of a significant decrease in radiolabeling in the midbrain and the striatum in those who had attempted suicide recently. These contradictory results are probably linked to a recent administration of an SSRI, which can induce a decrease in the central SERTs [298,299]. The results obtained with [18F]ADAM are very encouraging and place it as a future radiotracer for exploring and quantifying the central SERTs using PET. A closely related analogue of [18F]ADAM was described in 2009 by Wang et al., exhibiting high affinity and selectivity toward SERTs (Ki = 0.25 nM): [18F]FPBM [300]. The first studies in animals (rats) revealed specific labeling of the midbrain, cortex, striatum, and thalamus. [18F]FPBM evaluation was recently supplemented by tests on monkeys, which confirmed the interest of this compound as a new PET tracer of central SERTs [301].
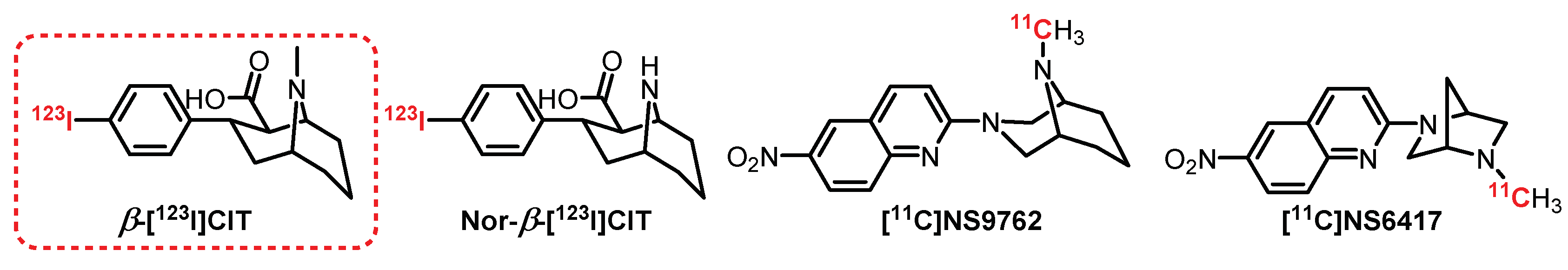
6.3. Tropane Series: β-[123I]CIT Derivatives
β-[123I]CIT (Figure 23) is one of the first radiotracers to be synthesized and tested for SPECT imaging in humans [302,303,304]. β-[123I]CIT has a mixed affinity for serotoninergic and dopaminergic transporters, which is not a major drawback since there is a different distribution within the brain for these two targets. However, its demethylated counterpart Nor-β-[123I]CIT has an affinity 10 times higher for SERTs and has allowed the observation in humans of a very strong accumulation of radioactivity in the midbrain and the striatum [305]. Since then, β-[123I]CIT and Nor-β-[123I]CIT have been used repeatedly to determine the evolution of the SERT density in the presence of pathologies such as depression and autism and in people with obsessive-compulsive disorders [306,307,308]. Recently, compounds exhibiting similar structures have been synthesized, particularly [11C]NS9762 and [11C]NS6417, which could provide promising alternatives in the coming years [309].
7. Conclusions
As described in this review, several potent and selective radiotracers for 5-HTR and SERTs have been developed in recent decades. Three radiotracers with good affinity to 5-HT1AR are commonly used in clinical trials: [11C]WAY100635, [18F]MPPF, and [18F]FCWAY. The characterization and distribution of 5-HT2AR is frequently performed by [123I]5-I-R91150 in SPECT imaging, with that of [18F]Setoperone, [18F]Altanserine, and [11C]MDL100907 performed in PET imaging. Moreover, the utilization of [123I]ADAM, [11C]DASB, and [11C]MADAM has improved the exploration of SERTs. These radiotracers have greatly contributed to facilitating the characterization, localization, and biological implication of the brain serotonin system in normal and pathological conditions. Indeed, in vivo PET and SPECT studies carried out on humans and animals allowed the observation of brain areas rich in 5-HTR by precise detection of radioactive emission. The signal specificities were confirmed via radiotracer displacement methods in the presence of reference ligands. Furthermore, these studies have shown the importance of multiparametric approaches to designing a new radiotracer, since a lack of selectivity, insufficient passage through the BBB, and fast metabolization can significantly decrease its effectiveness. In addition, these investigations also revealed the difficult transposition of the results from one species to another due to the interspecies variation of 5-HTR distribution, which could lead to misinterpretation. Overall, despite interesting results during clinical trials, none of these radiotracers are approved as diagnostic tools for neuropsychiatric disorders in clinics. New radiolabel-capable, potent, and selective ligands of the 5-HTRs allowing exploration of brain functions are thus still needed to fully understand the role of these receptors in either healthy or sick individuals.
Author Contributions
All the aspects of the writing of this review were achieved collectively by the co-authors (R.M., E.D., T.C. and V.C.). All authors have read and agreed to the published version of the manuscript.
Funding
The authors are grateful for financial support provided by the Regional Council of Normandy, FEDER, the University of Caen, and Carnot I2C.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Švob Štrac, D.; Pivac, N.; Mück-Šeler, D. The serotonergic system and cognitive function. Transl. Neurosci. 2016, 7, 35–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, D.E.; Nichols, C.D. Serotonin receptors. Chem. Rev. 2008, 108, 1614–1641. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, D. Serotonin receptors nomenclature. In The Serotonin System; Elsevier: Amsterdam, The Netherlands, 2019; pp. 63–93. ISBN 9780128133231. [Google Scholar]
- Homberg, J.R.; Schubert, D.; Gaspar, P. New perspectives on the neurodevelopmental effects of SSRIs. Trends Pharmacol. Sci. 2010, 31, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, P.P.A.; Hartig, P.; Hoyer, D. A proposed new nomenclature for 5-HT receptors. Trends Pharmacol. Sci. 1993, 14, 233–236. [Google Scholar] [CrossRef]
- Kobilka, B.K.; Frielle, T.; Collins, S.; Yang-Feng, T.; Kobilka, T.S.; Francke, U.; Lefkowitz, R.J.; Caron, M.G. An intronless gene encoding a potential member of the family of receptors coupled to guanine nucleotide regulatory proteins. Nature 1987, 329, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Fargin, A.; Raymond, J.R.; Lohse, M.J.; Kobilka, B.K.; Caron, M.G.; Lefkowitz, R.J. The genomic clone G-21 which resembles a β-adrenergic receptor sequence encodes the 5-HT1A receptor. Nature 1988, 335, 358–360. [Google Scholar] [CrossRef]
- Zhou, F.C.; Patel, T.D.; Swartz, D.; Xu, Y.; Kelley, M.R. Production and characterization of an anti-serotonin 1A receptor antibody which detects functional 5-HT(1A) binding sites. Mol. Brain Res. 1999, 69, 186–201. [Google Scholar] [CrossRef]
- Pompeiano, M.; Palacios, J.M.; Mengod, G. Distribution and cellular localization of mRNA coding for 5-HT(1A) receptor in the rat brain: Correlation with receptor binding. J. Neurosci. 1992, 12, 440–453. [Google Scholar] [CrossRef]
- Richardson-Jones, J.W.; Craige, C.P.; Guiard, B.P.; Stephen, A.; Metzger, K.L.; Kung, H.F.; Gardier, A.M.; Dranovsky, A.; David, D.J.; Beck, S.G.; et al. 5-HT1A Autoreceptor Levels Determine Vulnerability to Stress and Response to Antidepressants. Neuron 2010, 65, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Sprouse, J.S.; Aghajanian, G.K. Electrophysiological responses of serotoninergic dorsal raphe neurons to 5-HT1A and 5-HT1B agonists. Synapse 1987, 1, 3–9. [Google Scholar] [CrossRef]
- Riad, M.; Garcia, S.; Watkins, K.C.; Jodoin, N.; Doucet, E.; Langlois, X.; El Mestikawy, S.; Hamon, M.; Descarries, L. Somatodendritic localization of 5-HT1A and preterminal axonal localization of 5-HT1B serotonin receptors in adult rat brain. J. Comp. Neurol. 2000, 417, 181–194. [Google Scholar] [CrossRef]
- Maier, S.F.; Grahn, R.E.; Watkins, L.R. 8-OH-DPAT microinjected in the region of the dorsal raphe nucleus blocks and reverses the enhancement of fear conditioning and interference with escape produced by exposure to inescapable shock. Behav. Neurosci. 1995, 109, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Mendelson, S.D.; Quartermain, D.; Francisco, T.; Shemer, A. 5-HT1A receptor agonists induce anterograde amnesia in mice through a postsynaptic mechanism. Eur. J. Pharmacol. 1993, 236, 177–182. [Google Scholar] [CrossRef]
- Zhuang, X.; Gross, C.; Santarelli, L.; Compan, V.; Trillat, A.C.; Hen, R. Altered emotional states in knockout mice lacking 5-HT1A or 5-HT1B receptors. Neuropsychopharmacology 1999, 21, 52S–60S. [Google Scholar] [CrossRef] [Green Version]
- Billard, T.; Bars, D.; Zimmer, L. PET Radiotracers for Molecular Imaging of Serotonin 5-HT1A Receptors. Curr. Med. Chem. 2014, 21, 70–81. [Google Scholar] [CrossRef]
- Forster, E.A.; Cliffe, I.A.; Bill, D.J.; Dover, G.M.; Jones, D.; Reilly, Y.; Fletcher, A. A pharmacological profile of the selective silent 5-HT1A receptor antagonist, WAY-100635. Eur. J. Pharmacol. 1995, 281, 81–88. [Google Scholar] [CrossRef]
- Hall, H.; Lundkvist, C.; Halldin, C.; Farde, L.; Pike, V.W.; McCarron, J.A.; Fletcher, A.; Cliffe, I.A.; Barf, T.; Wikström, H.; et al. Autoradiographic localization of 5-HT(1A) receptors in the post-mortem human brain using [3H]WAY-100635. Brain Res. 1997, 745, 96–108. [Google Scholar] [CrossRef]
- Mathis, C.A.; Simpson, N.R.; Mahmood, K.; Kinahan, P.E.; Mintun, M.A. [11C]WAY 100635: A radioligand for imaging 5-HT1A receptors with positron emission tomography. Life Sci. 1994, 55, PL403–PL407. [Google Scholar] [CrossRef]
- Pike, V.W.; McCarron, J.A.; Lammerstma, A.A.; Hume, S.P.; Poole, K.; Grasby, P.M.; Malizia, A.; Cliffe, I.A.; Fletcher, A.; Bench, C.J. First delineation of 5-HT1A receptors in human brain with PET and [11C]WAY-100635. Eur. J. Pharmacol. 1995, 283, 3–5. [Google Scholar] [CrossRef]
- Hwang, D.R.; Simpson, N.R.; Montoya, J.; Mann, J.J.; Laruelle, M. An improved one-pot procedure for the preparation of [11C-carbonyl]-WAY100635. Nucl. Med. Biol. 1999, 26, 815–819. [Google Scholar] [CrossRef]
- Rabiner, E.A.; Messa, C.; Sargent, P.A.; Husted-Kjaer, K.; Montgomery, A.; Lawrence, A.D.; Bench, C.J.; Gunn, R.N.; Cowen, P.; Grasby, P.M. A database of [11C]WAY-100635 binding to 5-HT1A receptors in normal male volunteers: Normative data and relationship to methodological, demographic, physiological, and behavioral variables. Neuroimage 2002, 15, 620–632. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, G.M.; Ogden, R.T.; Oquendo, M.A.; Kumar, J.S.D.; Simpson, N.; Huang, Y.-y.; Mann, J.J.; Parsey, R.V. Positron Emission Tomography Quantification of Serotonin-1A Receptor Binding in Medication-Free Bipolar Depression. Biol. Psychiatry 2009, 66, 223–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penttilä, J.; Hirvonen, J.; Tuominen, L.; Lumme, V.; Ilonen, T.; Någren, K.; Hietala, J. Verbal memory and 5-HT1A receptors in healthy volunteers—A PET study with [carbonyl-11C]WAY-100635. Eur. Neuropsychopharmacol. 2016, 26, 570–577. [Google Scholar] [CrossRef]
- Metts, A.V.; Rubin-Falcone, H.; Ogden, R.T.; Lin, X.; Wilner, D.E.; Burke, A.K.; Sublette, M.E.; Oquendo, M.A.; Miller, J.M.; Mann, J.J. Antidepressant medication exposure and 5-HT 1A autoreceptor binding in major depressive disorder. Synapse 2019, 73, e22089. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Kung, M.-P.; Kung, H.F. Synthesis And Evaluation of 4-(2′-Methoxyphenyl)-1-[2′-[N-(2″-pyridinyl)-p-iodobenzamido]ethyl]piperazine (p-MPPI): A New Iodinated 5-HT1A Ligand. J. Med. Chem. 1994, 37, 1406–1407. [Google Scholar] [CrossRef] [PubMed]
- Shiue, C.-Y.; Shiue, G.G.; Mozley, P.D.; Kung, M.-P.; Zhuang, Z.-P.; Kim, H.-J.; Kung, H.F. p-[18F]-MPPF: A potential radioligand for PET studies of 5-HT1A receptors in humans. Synapse 1997, 25, 147–154. [Google Scholar] [CrossRef]
- Aznavour, N.; Zimmer, L. [18F]MPPF as a tool for the in vivo imaging of 5-HT1A receptors in animal and human brain. Neuropharmacology 2007, 52, 695–707. [Google Scholar] [CrossRef] [PubMed]
- Pauwelyn, G.; Vlerick, L.; Dockx, R.; Verhoeven, J.; Dobbeleir, A.; Peremans, K.; Goethals, I.; Bosmans, T.; Vanhove, C.; De Vos, F.; et al. PET quantification of [18F]MPPF in the canine brain using blood input and reference tissue modelling. PLoS ONE 2019, 14, e0218237. [Google Scholar] [CrossRef]
- Plenevaux, A.; Lemaire, C.; Aerts, J.; Lacan, G.; Rubins, D.; Melega, W.; Brihaye, C.; Degueldre, C.; Fuchs, S.; Salmon, E.; et al. [18F]p-MPPF: A Radiolabeled Antagonist for the Study of 5-HT1A Receptors with PET. Nucl. Med. Biol. 2000, 27, 467–471. [Google Scholar] [CrossRef]
- Costes, N.; Merlet, I.; Ostrowsky, K.; Faillenot, I.; Lavenne, F.; Zimmer, L.; Ryvlin, P.; Le Bars, D. A 18F-MPPF PET normative database of 5-HT1A receptor binding in men and women over aging. J. Nucl. Med. 2005, 46, 1980–1989. [Google Scholar]
- Kepe, V.; Barrio, J.R.; Huang, S.-C.; Ercoli, L.; Siddarth, P.; Shoghi-Jadid, K.; Cole, G.M.; Satyamurthy, N.; Cummings, J.L.; Small, G.W.; et al. Serotonin 1A receptors in the living brain of Alzheimer’s disease patients. Proc. Natl. Acad. Sci. USA 2006, 103, 702–707. [Google Scholar] [CrossRef] [Green Version]
- Merlet, I.; Ostrowsky, K.; Costes, N.; Ryvlin, P.; Isnard, J.; Faillenot, I.; Lavenne, F.; Dufournel, D.; Le Bars, D.; Mauguière, F. 5-HT1A receptor binding and intracerebral activity in temporal lobe epilepsy: An [18F]MPPF-PET study. Brain 2004, 127, 900–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lothe, A.; Didelot, A.; Hammers, A.; Costes, N.; Saoud, M.; Gilliam, F.; Ryvlin, P. Comorbidity between temporal lobe epilepsy and depression: A [18 F]MPPF PET study. Brain 2008, 131, 2765–2782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, L.; Jagoda, E.; Schmall, B.; Vuong, B.; Adams, H.R.; Nelson, D.L.; Carson, R.E.; Eckelman, W.C. Development of Fluorine-18-Labeled 5-HT 1A Antagonists. J. Med. Chem. 1999, 42, 1576–1586. [Google Scholar] [CrossRef]
- Toczek, M.T.; Carson, R.E.; Lang, L.; Ma, Y.; Spanaki, M.V.; Der, M.G.; Fazilat, S.; Kopylev, L.; Herscovitch, P.; Eckelman, W.C.; et al. PET imaging of 5-HT1A receptor binding in patients with temporal lobe epilepsy. Neurology 2003, 60, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Neumeister, A.; Bain, E.; Nugent, A.C.; Carson, R.E.; Bonne, O.; Luckenbaugh, D.A.; Eckelman, W.; Herscovitch, P.; Charney, D.S.; Drevets, W.C. Reduced Serotonin Type 1A Receptor Binding in Panic Disorder. J. Neurosci. 2004, 24, 589–591. [Google Scholar] [CrossRef] [Green Version]
- Bonne, O.; Bain, E.; Neumeister, A.; Nugent, A.C.; Vythilingam, M.; Carson, R.E.; Luckenbaugh, D.A.; Eckelman, W.; Herscovitch, P.; Drevets, W.C.; et al. No Change in Serotonin Type 1A Receptor Binding in Patients with Posttraumatic Stress Disorder. Am. J. Psychiatry 2005, 162, 383–385. [Google Scholar] [CrossRef]
- Ryu, Y.H.; Liow, J.-S.; Zoghbi, S.; Fujita, M.; Collins, J.; Tipre, D.; Sangare, J.; Hong, J.; Pike, V.W.; Innis, R.B. Disulfiram Inhibits Defluorination of 18F-FCWAY, Reduces Bone Radioactivity, and Enhances Visualization of Radioligand Binding to Serotonin 5-HT1A Receptors in Human Brain. J. Nucl. Med. 2007, 48, 1154–1161. [Google Scholar] [CrossRef] [Green Version]
- Saigal, N.; Bajwa, A.K.; Faheem, S.S.; Coleman, R.A.; Pandey, S.K.; Constantinescu, C.C.; Fong, V.; Mukherjee, J. Evaluation of serotonin 5-HT 1A receptors in rodent models using [18F]mefway PET. Synapse 2013, 67, 596–608. [Google Scholar] [CrossRef] [Green Version]
- Wooten, D.W.; Hillmer, A.T.; Moirano, J.M.; Ahlers, E.O.; Slesarev, M.; Barnhart, T.E.; Mukherjee, J.; Schneider, M.L.; Christian, B.T. Measurement of 5-HT 1A receptor density and in-vivo binding parameters of 18Fmefway in the nonhuman primate. J. Cereb. Blood Flow Metab. 2012, 32, 1546–1558. [Google Scholar] [CrossRef] [Green Version]
- Hillmer, A.T.; Wooten, D.W.; Bajwa, A.K.; Higgins, A.T.; Lao, P.J.; Betthauser, T.J.; Barnhart, T.E.; Rowley, H.A.; Stone, C.K.; Johnson, S.C.; et al. First-in-Human Evaluation of 18F-Mefway, a PET Radioligand Specific to Serotonin-1A Receptors. J. Nucl. Med. 2014, 55, 1973–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.Y.; Lyoo, C.H.; Kim, J.S.; Kim, K.M.; Kang, J.H.; Choi, S.-H.; Kim, J.-J.; Ryu, Y.H. 18F-Mefway PET Imaging of Serotonin 1A Receptors in Humans: A Comparison with 18F-FCWAY. PLoS ONE 2015, 10, e0121342. [Google Scholar] [CrossRef]
- Defraiteur, C.; Lemaire, C.; Luxen, A.; Plenevaux, A. Radiochemical synthesis and tissue distribution of p-[18F]DMPPF, a new 5-HT1A ligand for PET, in rats. Nucl. Med. Biol. 2006, 33, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Andrée, B.; Halldin, C.; Pike, V.W.; Gunn, R.N.; Olsson, H.; Farde, L. The PET radioligand [carbonyl-(11)C]desmethyl-WAY-100635 binds to 5-HT(1A) receptors and provides a higher radioactive signal than [carbonyl-(11)C]WAY-100635 in the human brain. J. Nucl. Med. 2002, 43, 292–303. [Google Scholar]
- Houle, S.; Wilson, A.A.; Inaba, T.; Fisher, N.; Dasilva, J.N. Imaging 5-ht1a receptors with positron emission tomography initial human studies with [x11c]cpc-222. Nucl. Med. Commun. 1997, 18, 1130–1134. [Google Scholar] [CrossRef]
- Sandell, J.; Halldin, C.; Pike, V.W.; Chou, Y.H.; Varnäs, K.; Hall, H.; Marchais, S.; Nowick, B.; Wikström, H.V.; Swahn, C.G.; et al. New halogenated [11C]WAY analogues, [11C]6FPWAY and [11C]6BPWAY-Radiosynthesis and assessment as radioligands for the study of brain 5-HT1A receptors in living monkey. Nucl. Med. Biol. 2001, 28, 177–185. [Google Scholar] [CrossRef]
- Karramkam, M.; Hinnen, F.; Berrehouma, M.; Hlavacek, C.; Vaufrey, F.; Halldin, C.; McCarron, J.A.; Pike, V.W.; Dollé, F. Synthesis of a [6-Pyridinyl-18F]-labelled fluoro derivative of WAY-100635 as a candidate radioligand for brain 5-HT 1A receptor imaging with PET. Bioorg. Med. Chem. 2003, 11, 2769–2782. [Google Scholar] [CrossRef]
- McCarron, J. The Pyridinyl-6 Position of WAY-100635 as a site for radiofluorination—Effect on 5-HT1A receptor radioligand behavior in vivo. Mol. Imaging Biol. 2004, 6, 17–26. [Google Scholar] [CrossRef]
- Kumar, J.S.D.; Majo, V.J.; Hsiung, S.; Millak, M.S.; Liu, K.; Tamir, H.; Prabhakaran, J.; Simpson, N.R.; Van Heertum, R.L.; Mann, J.J.; et al. Synthesis and in Vivo Validation of [O-Methyl-11C]2-{4-[4-(7-methoxynaphthalen-1-yl)piperazin-1-yl]butyl}-4-methyl-2H-[1,2,4]triazine-3,5-dione: A Novel 5-HT 1A Receptor Agonist Positron Emission Tomography Ligand. J. Med. Chem. 2006, 49, 125–134. [Google Scholar] [CrossRef]
- Kumar, J.S.D.; Prabhakaran, J.; Majo, V.J.; Milak, M.S.; Hsiung, S.-C.; Tamir, H.; Simpson, N.R.; Van Heertum, R.L.; Mann, J.J.; Parsey, R.V. Synthesis and in vivo evaluation of a novel 5-HT1A receptor agonist radioligand [O-methyl-11C]2-(4-(4-(2-methoxyphenyl)piperazin-1-yl)butyl)-4-methyl-1,2,4-triazine-3,5(2H,4H)dione in nonhuman primates. Eur. J. Nucl. Med. Mol. Imaging 2007, 34, 1050–1060. [Google Scholar] [CrossRef]
- Milak, M.S.; DeLorenzo, C.; Zanderigo, F.; Prabhakaran, J.; Kumar, J.S.D.; Majo, V.J.; Mann, J.J.; Parsey, R.V. In Vivo Quantification of Human Serotonin 1A Receptor Using 11 C-CUMI-101, an Agonist PET Radiotracer. J. Nucl. Med. 2010, 51, 1892–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hines, C.S.; Liow, J.-S.; Zanotti-Fregonara, P.; Hirvonen, J.; Morse, C.; Pike, V.W.; Innis, R.B. Human Biodistribution and Dosimetry of 11C-CUMI-101, an Agonist Radioligand for Serotonin-1A Receptors in Brain. PLoS ONE 2011, 6, e25309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendry, N.; Christie, I.; Rabiner, E.A.; Laruelle, M.; Watson, J. In vitro assessment of the agonist properties of the novel 5-HT1A receptor ligand, CUMI-101 (MMP), in rat brain tissue. Nucl. Med. Biol. 2011, 38, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Girgis, R.R.; Forbes, A.; Abi-Dargham, A.; Slifstein, M. A positron emission tomography occupancy study of brexpiprazole at dopamine D2 and D3 and serotonin 5-HT1A and 5-HT2A receptors, and serotonin reuptake transporters in subjects with schizophrenia. Neuropsychopharmacology 2020, 45, 786–792. [Google Scholar] [CrossRef]
- Majo, V.J.; Milak, M.S.; Prabhakaran, J.; Mali, P.; Savenkova, L.; Simpson, N.R.; Mann, J.J.; Parsey, R.V.; Kumar, J.S.D. Synthesis and in vivo evaluation of [18F]2-(4-(4-(2-(2-fluoroethoxy)phenyl)piperazin-1-yl)butyl)-4-methyl-1,2,4-triazine-3,5(2H,4H)-dione ([18F]FECUMI-101) as an imaging probe for 5-HT1A receptor agonist in nonhuman primates. Bioorg. Med. Chem. 2013, 21, 5598–5604. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.S.D.; Underwood, M.D.; Simpson, N.R.; Kassir, S.A.; Prabhakaran, J.; Majo, V.J.; Bakalian, M.J.; Parsey, R.V.; Mann, J.J.; Arango, V. Autoradiographic Evaluation of [18F]FECUMI-101, a High Affinity 5-HT1AR Ligand in Human Brain. ACS Med. Chem. Lett. 2016, 7, 482–486. [Google Scholar] [CrossRef] [Green Version]
- Collier, T.L.; Liang, S.H.; Mann, J.J.; Vasdev, N.; Kumar, J.S.D. Microfluidic radiosynthesis of [ 18 F]FEMPT, a high affinity PET radiotracer for imaging serotonin receptors. Beilstein J. Org. Chem. 2017, 13, 2922–2927. [Google Scholar] [CrossRef] [Green Version]
- Vacher, B.; Bonnaud, B.; Funes, P.; Jubault, N.; Koek, W.; Assié, M.B.; Cosi, C.; Kleven, M. Novel derivatives of 2-pyridinemethylamine as selective, potent, and orally active agonists at 5-HT(1A) receptors. J. Med. Chem. 1999, 42, 1648–1660. [Google Scholar] [CrossRef] [Green Version]
- Koek, W.; Vacher, B.; Cosi, C.; Assié, M.B.; Patoiseau, J.F.; Pauwels, P.J.; Colpaert, F.C. 5-HT1A receptor activation and antidepressant-like effects: F 13714 has high efficacy and marked antidepressant potential. Eur. J. Pharmacol. 2001, 420, 103–112. [Google Scholar] [CrossRef]
- Lemoine, L.; Becker, G.; Vacher, B.; Billard, T.; Lancelot, S.; Newman-Tancredi, A.; Zimmer, L. Radiosynthesis and Preclinical Evaluation of 18 F-F13714 as a Fluorinated 5-HT 1A Receptor Agonist Radioligand for PET Neuroimaging. J. Nucl. Med. 2012, 53, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, C.; Mawatari, A.; Kawasaki, A.; Takeda, C.; Onoe, K.; Doi, H.; Newman-Tancredi, A.; Zimmer, L.; Onoe, H. Marmoset Serotonin 5-HT 1A Receptor Mapping with a Biased Agonist PET Probe 18 F-F13714: Comparison with an Antagonist Tracer 18 F-MPPF in Awake and Anesthetized States. Int. J. Neuropsychopharmacol. 2016, 19, pyw079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heusler, P.; Palmier, C.; Tardif, S.; Bernois, S.; Colpaert, F.C.; Cussac, D. [3H]-F13640, a novel, selective and high-efficacy serotonin 5-HT1A receptor agonist radioligand. Naunyn Schmiedebergs Arch. Pharmacol. 2010, 382, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.-J.; Colpaert, F.; Wiesenfeld-Hallin, Z. Opioid hyperalgesia and tolerance versus 5-HT1A receptor-mediated inverse tolerance. Trends Pharmacol. Sci. 2003, 24, 634–639. [Google Scholar] [CrossRef]
- Vidal, B.; Fieux, S.; Colom, M.; Billard, T.; Bouillot, C.; Barret, O.; Constantinescu, C.; Tamagnan, G.; Newman-Tancredi, A.; Zimmer, L. 18F-F13640 preclinical evaluation in rodent, cat and primate as a 5-HT1A receptor agonist for PET neuroimaging. Brain Struct. Funct. 2018, 223, 2973–2988. [Google Scholar] [CrossRef] [PubMed]
- Vidal, B.; Sebti, J.; Verdurand, M.; Fieux, S.; Billard, T.; Streichenberger, N.; Troakes, C.; Newman-Tancredi, A.; Zimmer, L. Agonist and antagonist bind differently to 5-HT1A receptors during Alzheimer’s disease: A post-mortem study with PET radiopharmaceuticals. Neuropharmacology 2016, 109, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Colom, M.; Costes, N.; Redouté, J.; Dailler, F.; Gobert, F.; Le Bars, D.; Billard, T.; Newman-Tancredi, A.; Zimmer, L. 18F-F13640 PET imaging of functional receptors in humans. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 220–221. [Google Scholar] [CrossRef]
- Colom, M.; Vidal, B.; Fieux, S.; Redoute, J.; Costes, N.; Lavenne, F.; Mérida, I.; Irace, Z.; Iecker, T.; Bouillot, C.; et al. [18F]F13640, a 5-HT1A Receptor Radiopharmaceutical Sensitive to Brain Serotonin Fluctuations. Front. Neurosci. 2021, 15, 622423. [Google Scholar] [CrossRef]
- Sari, Y. Serotonin receptors: From protein to physiological function and behavior. Neurosci. Biobehav. Rev. 2004, 28, 565–582. [Google Scholar] [CrossRef]
- Rutz, S.; Riegert, C.; Rothmaier, A.K.; Buhot, M.C.; Cassel, J.C.; Jackisch, R. Presynaptic serotonergic modulation of 5-HT and acetylcholine release in the hippocampus and the cortex of 5-HT1B-receptor knockout mice. Brain Res. Bull. 2006, 70, 81–93. [Google Scholar] [CrossRef]
- Olivier, B.; van Oorschot, R. 5-HT1B receptors and aggression: A review. Eur. J. Pharmacol. 2005, 526, 207–217. [Google Scholar] [CrossRef]
- Pierson, M.E.; Andersson, J.; Nyberg, S.; McCarthy, D.J.; Finnema, S.J.; Varnäs, K.; Takano, A.; Karlsson, P.; Gulyás, B.; Medd, A.M.; et al. [11C]AZ10419369: A selective 5-HT1B receptor radioligand suitable for positron emission tomography (PET). Characterization in the primate brain. Neuroimage 2008, 41, 1075–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, D.L.; Sobotka-Briner, C.; Ding, M.; Powell, M.E.; Jiang, Q.; Hill, G.; Heys, J.R.; Elmore, C.S.; Pierson, M.E.; Mrzljak, L. [N-methyl-3H3]AZ10419369 binding to the 5-HT 1B receptor: In vitro characterization and in vivo receptor occupancy. J. Pharmacol. Exp. Ther. 2009, 330, 342–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varnäs, K.; Nyberg, S.; Halldin, C.; Varrone, A.; Takano, A.; Karlsson, P.; Andersson, J.; McCarthy, D.; Smith, M.; Pierson, M.E.; et al. Quantitative Analysis of [ 11 C]AZ10419369 Binding to 5-HT 1B Receptors in Human Brain. J. Cereb. Blood Flow Metab. 2011, 31, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Nord, M.; Finnema, S.J.; Schain, M.; Halldin, C.; Farde, L. Test–retest reliability of [11C]AZ10419369 binding to 5-HT1B receptors in human brain. Eur. J. Nucl. Med. Mol. Imaging 2014, 41, 301–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiger, M.; Farde, L.; Rück, C.; Varrone, A.; Forsberg, A.; Lindefors, N.; Halldin, C.; Lundberg, J. Low serotonin1B receptor binding potential in the anterior cingulate cortex in drug-free patients with recurrent major depressive disorder. Psychiatry Res. Neuroimaging 2016, 253, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varrone, A.; Svenningsson, P.; Forsberg, A.; Varnäs, K.; Tiger, M.; Nakao, R.; Halldin, C.; Nilsson, L.-G.; Farde, L. Positron emission tomography imaging of 5-hydroxytryptamine1B receptors in Parkinson’s disease. Neurobiol. Aging 2014, 35, 867–875. [Google Scholar] [CrossRef]
- Lindberg, A.; Nag, S.; Schou, M.; Takano, A.; Matsumoto, J.; Amini, N.; Elmore, C.S.; Farde, L.; Pike, V.W.; Halldin, C. [11C]AZ10419096—A full antagonist PET radioligand for imaging brain 5-HT1B receptors. Nucl. Med. Biol. 2017, 54, 34–40. [Google Scholar] [CrossRef]
- Lindberg, A.; Arakawa, R.; Nogami, T.; Nag, S.; Schou, M.; Elmore, C.S.; Farde, L.; Pike, V.W.; Halldin, C. Potential for imaging the high-affinity state of the 5-HT1B receptor: A comparison of three PET radioligands with differing intrinsic activity. EJNMMI Res. 2019, 9, 100. [Google Scholar] [CrossRef] [Green Version]
- Lindberg, A.; Nag, S.; Schou, M.; Arakawa, R.; Nogami, T.; Moein, M.M.; Elmore, C.S.; Pike, V.W.; Halldin, C. Development of a 18F-labeled PET radioligand for imaging 5-HT1B receptors: [18F]AZ10419096. Nucl. Med. Biol. 2019, 78–79, 11–16. [Google Scholar] [CrossRef]
- Ridler, K.; Plisson, C.; Rabiner, E.A.; Gunn, R.N.; Easwaramoorthy, B.; Abi-Dargham, A.; Laruelle, M.; Slifstein, M. Characterization of in vivo pharmacological properties and sensitivity to endogenous serotonin of [11C] P943: A positron emission tomography study in Papio anubis. Synapse 2011, 65, 1119–1127. [Google Scholar] [CrossRef]
- Murrough, J.W.; Henry, S.; Hu, J.; Gallezot, J.-D.; Planeta-Wilson, B.; Neumaier, J.F.; Neumeister, A. Reduced ventral striatal/ventral pallidal serotonin1B receptor binding potential in major depressive disorder. Psychopharmacology 2011, 213, 547–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matuskey, D.; Bhagwagar, Z.; Planeta, B.; Pittman, B.; Gallezot, J.-D.; Chen, J.; Wanyiri, J.; Najafzadeh, S.; Ropchan, J.; Geha, P.; et al. Reductions in Brain 5-HT1B Receptor Availability in Primarily Cocaine-Dependent Humans. Biol. Psychiatry 2014, 76, 816–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Henry, S.; Gallezot, J.D.; Ropchan, J.; Neumaier, J.F.; Potenza, M.N.; Sinha, R.; Krystal, J.H.; Huang, Y.; Ding, Y.S.; et al. Serotonin 1B Receptor Imaging in Alcohol Dependence. Biol. Psychiatry 2010, 67, 800–803. [Google Scholar] [CrossRef] [Green Version]
- Leysen, J.E. 5-HT2 receptors. Curr. Drug Targets CNS Neurol. Disord. 2004, 3, 11–26. [Google Scholar] [CrossRef]
- Conn, P.J.; Sanders-Bush, E. Selective 5ht-2 antagonists inhibit serotonin stimulated phosphatidylinositol metabolism in cerebral cortex. Neuropharmacology 1984, 23, 993–996. [Google Scholar] [CrossRef]
- Kim, J.; Moon, B.S.; Lee, B.C.; Lee, H.Y.; Kim, H.J.; Choo, H.; Pae, A.N.; Cho, Y.S.; Min, S.J. A Potential PET Radiotracer for the 5-HT2C Receptor: Synthesis and in Vivo Evaluation of 4-(3-[18F]fluorophenethoxy)pyrimidine. ACS Chem. Neurosci. 2017, 8, 996–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, F.; Nye, J.A.; Voll, R.J.; Howell, L.; Goodman, M.M. Synthesis and Evaluation of Pyridyloxypyridyl Indole Carboxamides as Potential PET Imaging Agents for 5-HT2C Receptors. ACS Med. Chem. Lett. 2018, 9, 188–192. [Google Scholar] [CrossRef]
- Branchek, T.; Adham, N.; Macchi, M.; Kao, H.T.; Hartig, P.R. [3H]-DOB(4-bromo-2,5-dimethoxyphenylisopropylamine) and [3H] ketanserin label two affinity states of the cloned human 5-hydroxytryptamine2 receptor. Mol. Pharmacol. 1990, 38, 604–609. [Google Scholar]
- Nichols, D.E. Hallucinogens. Pharmacol. Ther. 2004, 101, 131–181. [Google Scholar] [CrossRef]
- Hall, H.; Farde, L.; Halldin, C.; Lundkvist, C.; Sedvall, G. Autoradiographic localization of 5-HT(2A) receptors in the human brain using [3H]M100907 and [11C]M100907. Synapse 2000, 38, 421–431. [Google Scholar] [CrossRef]
- Hasuo, H.; Matsuoka, T.; Akasu, T. Activation of presynaptic 5-hydroxytryptamine 2A receptors facilitates excitatory synaptic transmission via protein kinase C in the dorsolateral septal nucleus. J. Neurosci. 2002, 22, 7509–7517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gijsman, H.J.; Verkes, R.J.; Schouten-Verhagen, J.C.M.; Schoemaker, R.C.; Van Gerven, J.M.A.; De Rijk, R.H.; Van Kempen, G.M.J. A review of the role of serotonin receptors in psychiatric disorders. Hum. Psychopharmacol. 2000, 15, 397–415. [Google Scholar] [CrossRef]
- Berridge, M.; Comar, D.; Crouzel, C.; Baron, J.-C. 11C-Labeled ketanserin: A selective serotonin S2 antagonist. J. Label. Compd. Radiopharm. 1983, 20, 73–78. [Google Scholar] [CrossRef]
- Blin, J.; Pappata, S.; Kiyosawa, M.; Crouzel, C.; Baron, J.C. [18F]Setoperone: A new high-affinity ligand for positron emission tomography study of the serotonin-2 receptors in baboon brain in vivo. Eur. J. Pharmacol. 1988, 147, 73–82. [Google Scholar] [CrossRef]
- Blin, J.; Sette, G.; Fiorelli, M.; Bletry, O.; Elghozi, J.L.; Crouzel, C.; Baron, J.C. A Method for the In Vivo Investigation of the Serotonergic 5-HT2 Receptors in the Human Cerebral Cortex Using Positron Emission Tomography and 18F-Labeled Setoperone. J. Neurochem. 1990, 54, 1744–1754. [Google Scholar] [CrossRef]
- Meyer, J.H.; Kapur, S.; Houle, S.; DaSilva, J.; Owczarek, B.; Brown, G.M.; Wilson, A.A.; Kennedy, S.H. Prefrontal cortex 5-HT2 receptors in depression: An [18F]setoperone PET imaging study. Am. J. Psychiatry 1999, 156, 1029–1034. [Google Scholar] [CrossRef]
- Massou, J.M.; Trichard, C.; Attar-Levy, D.; Feline, A.; Corruble, E.; Beaufils, B.; Martinot, J.L. Frontal 5-HT(2A) receptors studied in depressive patients during chronic treatment by selective serotonin reuptake inhibitors. Psychopharmacology 1997, 133, 99–101. [Google Scholar] [CrossRef]
- Chabriat, H.; Vera, P.; Samson, Y.; Pappata, S.; Boullais, N.; Tehindrazanarivelo, A.; Bousser, M. 5HT2 receptors in cerebral cortex of migraineurs studied using PET and 18F-fluorosetoperone. Cephalalgia 1995, 15, 104–108. [Google Scholar] [CrossRef]
- Blin, J.; Baron, J.C.; Dubois, B.; Crouzel, C.; Fiorelli, M.; Attar-Lévy, D.; Pillon, B.; Fournier, D.; Vidailhet, M.; Agid, Y. Loss of brain 5-HT 2 receptors in Alzheimer’s disease. Brain 1993, 116, 497–510. [Google Scholar] [CrossRef]
- Lemaire, C.; Cantineau, R.; Guillaume, M.; Plenevaux, A.; Christiaens, L. Fluorine-18-altanserin: A radioligand for the study of serotonin receptors with PET: Radiolabeling and in vivo biologic behavior in rats. J. Nucl. Med. 1991, 32, 2266–2272. [Google Scholar]
- Biver, F.; Goldman, S.; Luxen, A.; Monclus, M.; Forestini, M.; Mendlewicz, J.; Lotstra, F. Multicompartmental study of fluorine-18 altanserin binding to brain 5HT2 receptors in humans using positron emission tomography. Eur. J. Nucl. Med. 1994, 21, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Price, J.C.; Lopresti, B.J.; Mason, N.S.; Holt, D.P.; Huang, Y.; Mathis, C.A. Analyses of [18F]altanserin bolus injection PET data. I: Consideration of radiolabeled metabolites in baboons. Synapse 2001, 41, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.Z.; Baldwin, R.M.; Van Dyck, C.H.; Al-Tikriti, M.; Roth, B.; Khan, N.; Charney, D.S.; Innis, R.B. Characterization of radioactive metabolites of 5-HT(2A) receptor PET ligand [18F]altanserin in human and rodent. Nucl. Med. Biol. 1999, 26, 601–608. [Google Scholar] [CrossRef]
- Adams, K.H.; Pinborg, L.H.; Svarer, C.; Hasselbalch, S.G.; Holm, S.; Haugbøl, S.; Madsen, K.; Frøkjær, V.; Martiny, L.; Paulson, O.B.; et al. A database of [18F]-altanserin binding to 5-HT2A receptors in normal volunteers: Normative data and relationship to physiological and demographic variables. Neuroimage 2004, 21, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Frank, G.K.; Kaye, W.H.; Meltzer, C.C.; Price, J.C.; Greer, P.; McConaha, C.; Skovira, K. Reduced 5-HT2A receptor binding after recovery from anorexia nervosa. Biol. Psychiatry 2002, 52, 896–906. [Google Scholar] [CrossRef]
- Rasmussen, H.; Frokjaer, V.G.; Hilker, R.W.; Madsen, J.; Anhøj, S.; Oranje, B.; Pinborg, L.H.; Glenthøj, B.; Knudsen, G.M. Low frontal serotonin 2A receptor binding is a state marker for schizophrenia? Eur. Neuropsychopharmacol. 2016, 26, 1248–1250. [Google Scholar] [CrossRef]
- Mintun, M.A.; Sheline, Y.I.; Moerlein, S.M.; Vlassenko, A.G.; Huang, Y.; Snyder, A.Z. Decreased hippocampal 5-HT2A receptor binding in major depressive disorder: In vivo measurement with [18F]altanserin positron emission tomography. Biol. Psychiatry 2004, 55, 217–224. [Google Scholar] [CrossRef]
- Marner, L.; Frokjaer, V.G.; Kalbitzer, J.; Lehel, S.; Madsen, K.; Baaré, W.F.C.; Knudsen, G.M.; Hasselbalch, S.G. Loss of serotonin 2A receptors exceeds loss of serotonergic projections in early Alzheimer’s disease: A combined [11C]DASB and [18F]altanserin-PET study. Neurobiol. Aging 2012, 33, 479–487. [Google Scholar] [CrossRef]
- Staley, J.K.; Van Dyck, C.H.; Tan, P.Z.; Al Tikriti, M.; Ramsby, Q.; Klump, H.; Ng, C.; Garg, P.; Soufer, R.; Baldwin, R.M.; et al. Comparison of [18F]altanserin and [18F]deuteroaltanserin for PET imaging of serotonin2A receptors in baboon brain: Pharmacological studies. Nucl. Med. Biol. 2001, 28, 271–279. [Google Scholar] [CrossRef]
- Van Dyck, C.H.; Soares, J.C.; Tan, P.Z.; Staley, J.K.; Baldwin, R.M.; Amici, L.A.; Fu, X.; Garg, P.K.; Seibyl, J.P.; Charney, D.S.; et al. Equilibrium modeling of 5-HT2A receptors with [18F]deuteroaltanserin and PET: Feasibility of a constant infusion paradigm. Nucl. Med. Biol. 2000, 27, 715–722. [Google Scholar] [CrossRef]
- Soares, J.C.; Van Dyck, C.H.; Tan, P.Z.; Zoghbi, S.S.; Garg, P.; Soufer, R.; Baldwin, R.M.; Fujita, M.; Staley, J.K.; Fu, X.; et al. Reproducibility of in vivo brain measures of 5-HT2A receptors with PET and [18F]deuteroaltanserin. Psychiatry Res. Neuroimaging 2001, 106, 81–93. [Google Scholar] [CrossRef]
- Santhosh, L.; Estok, K.M.; Vogel, R.S.; Tamagnan, G.D.; Baldwin, R.M.; Mitsis, E.M.; MacAvoy, M.G.; Staley, J.K.; van Dyck, C.H. Regional distribution and behavioral correlates of 5-HT2A receptors in Alzheimer’s disease with [18F]deuteroaltanserin and PET. Psychiatry Res. Neuroimaging 2009, 173, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Nyberg, S.; Halldin, C.; Lundkvist, C.; Farde, L. PET imaging of central 5-HT(2A) receptors with carbon-11-MDL 100,907. J. Nucl. Med. 1998, 39, 208–214. [Google Scholar] [PubMed]
- Lopez-Gimenez, J.F.; Vilaro, M.T.; Palacios, M.; Mengod, G. [3H] MDL 100, 907 labels 5-HT 2A serotonin receptors selectively in primate brain. Neuropharmacology 1998, 37, 1147–1158. [Google Scholar] [CrossRef] [Green Version]
- López-Giménez, J.F.; Mengod, G.; Palacios, J.M.; Vilaró, M.T. Selective visualization of rat brain 5-HT(2A) receptors by autoradiography with [3H]MDL 100,907. Naunyn Schmiedebergs Arch. Pharmacol. 1997, 356, 446–454. [Google Scholar] [CrossRef]
- Lundkvist, C.; Halldin, C.; Ginovart, N.; Nyberg, S.; Swahn, C.G.; Carr, A.A.; Brunner, F.; Farde, L. [11C]MDL 100907, a radioligand for selective imaging of 5-HT2A receptors with positron emission tomography. Life Sci. 1996, 58, 187–192. [Google Scholar] [CrossRef]
- Talbot, P.S.; Slifstein, M.; Hwang, D.R.; Huang, Y.; Scher, E.; Abi-Dargham, A.; Laruelle, M. Extended characterisation of the serotonin 2A (5-HT 2A) receptor-selective PET radiotracer 11C-MDL100907 in humans: Quantitative analysis, test-retest reproducibility, and vulnerability to endogenous 5-HT tone. Neuroimage 2012, 59, 271–285. [Google Scholar] [CrossRef] [Green Version]
- Bhagwagar, Z.; Hinz, R.; Taylor, M.; Fancy, S.; Cowen, P.; Grasby, P. Increased 5-HT2A receptor binding in euthymic, medication-free patients recovered from depression: A positron emission study with [11C]MDL 100,907. Am. J. Psychiatry 2006, 163, 1580–1587. [Google Scholar] [CrossRef]
- Girgis, R.R.; Slifstein, M.; Xu, X.; Frankle, W.G.; Anagnostou, E.; Wasserman, S.; Pepa, L.; Kolevzon, A.; Abi-Dargham, A.; Laruelle, M.; et al. The 5-HT 2A receptor and serotonin transporter in Asperger’s Disorder: A PET study with [11C]MDL 100907 and [11C]DASB. Psychiatry Res. Neuroimaging 2011, 194, 230–234. [Google Scholar] [CrossRef] [Green Version]
- Simpson, H.B.; Slifstein, M.; Bender, J.; Xu, X.; Hackett, E.; Maher, M.J.; Abi-Dargham, A. Serotonin 2A receptors in obsessive-compulsive disorder: A positron emission tomography study with [11C]MDL 100907. Biol. Psychiatry 2011, 70, 897–904. [Google Scholar] [CrossRef] [Green Version]
- Scott, D.O.; Heath, T.G. Investigation of the CNS penetration of a potent 5-HT(2a) receptor antagonist (MDL 100,907) and an active metabolite (MDL 105,725) using in vivo microdialysis sampling in the rat. J. Pharm. Biomed. Anal. 1998, 17, 17–25. [Google Scholar] [CrossRef]
- Ren, H.; Wey, H.Y.; Strebl, M.; Neelamegam, R.; Ritter, T.; Hooker, J.M. Synthesis and imaging validation of [18F]MDL100907 enabled by Ni-mediated fluorination. ACS Chem. Neurosci. 2014, 5, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Herth, M.M.; Piel, M.; Debus, F.; Schmitt, U.; Lüddens, H.; Rösch, F. Preliminary in vivo and ex vivo evaluation of the 5-HT2A imaging probe [18F]MH.MZ. Nucl. Med. Biol. 2009, 36, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Debus, F.; Herth, M.M.; Piel, M.; Buchholz, H.G.; Bausbacher, N.; Kramer, V.; Lüddens, H.; Rösch, F. 18F-Labeling and evaluation of novel MDL 100907 derivatives as potential 5-HT2A antagonists for molecular imaging. Nucl. Med. Biol. 2010, 37, 487–495. [Google Scholar] [CrossRef]
- Hansen, H.D.; Ettrup, A.; Herth, M.M.; Dyssegaard, A.; Ratner, C.; Gillings, N.; Knudsen, G.M. Direct comparison of [18F]MH.MZ and [18F]altanserin for 5-HT2A receptor imaging with PET. Synapse 2013, 67, 328–337. [Google Scholar] [CrossRef]
- Ettrup, A.; Palner, M.; Gillings, N.; Santini, M.A.; Hansen, M.; Kornum, B.R.; Rasmussen, L.K.; Någren, K.; Madsen, J.; Begtrup, M.; et al. Radiosynthesis and evaluation of11C-CIMBI-5 as a 5-HT 2A receptor agonist radioligand for PET. J. Nucl. Med. 2010, 51, 1763–1770. [Google Scholar] [CrossRef] [Green Version]
- Prabhakaran, J.; DeLorenzo, C.; Zanderigo, F.; Knudsen, G.M.; Gilling, N.; Pratap, M.; Jorgensen, M.J.; Daunais, J.; Kaplan, J.R.; Parsey, R.V.; et al. In vivo PET Imaging of [11C]CIMBI-5, a 5-HT2AR Agonist Radiotracer in Nonhuman Primates. J. Pharm. Pharm. Sci. 2019, 22, 352–364. [Google Scholar] [CrossRef]
- Ettrup, A.; Hansen, M.; Santini, M.A.; Paine, J.; Gillings, N.; Palner, M.; Lehel, S.; Herth, M.M.; Madsen, J.; Kristensen, J.; et al. Radiosynthesis and in vivo evaluation of a series of substituted 11C-phenethylamines as 5-HT2A agonist PET tracers. Eur. J. Nucl. Med. Mol. Imaging 2011, 38, 681–693. [Google Scholar] [CrossRef]
- Johansen, A.; Hansen, H.D.; Svarer, C.; Lehel, S.; Leth-Petersen, S.; Kristensen, J.L.; Gillings, N.; Knudsen, G.M. The importance of small polar radiometabolites in molecular neuroimaging: A PET study with [11C]Cimbi-36 labeled in two positions. J. Cereb. Blood Flow Metab. 2018, 38, 659–668. [Google Scholar] [CrossRef] [Green Version]
- Johansen, A.; Holm, S.; Dall, B.; Keller, S.; Kristensen, J.L.; Knudsen, G.M.; Hansen, H.D. Human biodistribution and radiation dosimetry of the 5-HT2A receptor agonist Cimbi-36 labeled with carbon-11 in two positions. EJNMMI Res. 2019, 9, 71. [Google Scholar] [CrossRef]
- Jørgensen, L.M.; Weikop, P.; Villadsen, J.; Visnapuu, T.; Ettrup, A.; Hansen, H.D.; Baandrup, A.O.; Andersen, F.L.; Bjarkam, C.R.; Thomsen, C.; et al. Cerebral 5-HT release correlates with [11C]Cimbi36 PET measures of 5-HT2A receptor occupancy in the pig brain. J. Cereb. Blood Flow Metab. 2017, 37, 425–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Cunha-Bang, S.; Ettrup, A.; Mc Mahon, B.; Skibsted, A.P.; Schain, M.; Lehel, S.; Dyssegaard, A.; Jørgensen, L.M.; Møller, K.; Gillings, N.; et al. Measuring endogenous changes in serotonergic neurotransmission with [11C]Cimbi-36 positron emission tomography in humans. Transl. Psychiatry 2019, 9, 134. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.C.; Stepanov, V.; Martinsson, S.; Ettrup, A.; Takano, A.; Knudsen, G.M.; Halldin, C.; Farde, L.; Finnema, S.J. Fenfluramine Reduces [11C]Cimbi-36 Binding to the 5-HT2A Receptor in the Nonhuman Primate Brain. Int. J. Neuropsychopharmacol. 2017, 20, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Erritzoe, D.; Ashok, A.H.; Searle, G.E.; Colasanti, A.; Turton, S.; Lewis, Y.; Huiban, M.; Moz, S.; Passchier, J.; Saleem, A.; et al. Serotonin release measured in the human brain: A PET study with [11C]CIMBI-36 and d-amphetamine challenge. Neuropsychopharmacology 2020, 45, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Ettrup, A.; Svarer, C.; McMahon, B.; da Cunha-Bang, S.; Lehel, S.; Møller, K.; Dyssegaard, A.; Ganz, M.; Beliveau, V.; Jørgensen, L.M.; et al. Serotonin 2A receptor agonist binding in the human brain with [11C]Cimbi-36: Test-retest reproducibility and head-to-head comparison with the antagonist [18F]altanserin. Neuroimage 2016, 130, 167–174. [Google Scholar] [CrossRef]
- Prabhakaran, J.; Underwood, M.D.; Kumar, J.S.D.; Simpson, N.R.; Kassir, S.A.; Bakalian, M.J.; Mann, J.J.; Arango, V. Synthesis and in vitro evaluation of [18F]FECIMBI-36: A potential agonist PET ligand for 5-HT2A/2C receptors. Bioorg. Med. Chem. Lett. 2015, 25, 3933–3936. [Google Scholar] [CrossRef] [Green Version]
- Prabhakaran, J.; Solingapuram Sai, K.K.; Zanderigo, F.; Rubin-Falcone, H.; Jorgensen, M.J.; Kaplan, J.R.; Tooke, K.I.; Mintz, A.; Mann, J.J.; Kumar, J.S.D. In vivo evaluation of [18F]FECIMBI-36, an agonist 5-HT2A/2Creceptor PET radioligand in nonhuman primate. Bioorg. Med. Chem. Lett. 2017, 27, 21–23. [Google Scholar] [CrossRef] [Green Version]
- Sargent, T.; Budinger, T.F.; Braun, G.; Shulgin, A.T.; Braun, U. An iodinated catecholamine congener for brain imaging and metabolic studies. J. Nucl. Med. 1978, 19, 71–76. [Google Scholar]
- Glennon, R.A.; Seggel, M.R.; Soine, W.H.; Herrick-Davis, K.; Lyon, R.A.; Titeler, M. Iodine-125 labeled 1-(2,5-dimethoxy-4-iodophenyl)-2-aminopropane: An iodinated radioligand that specifically labels the agonist high-affinity state of 5-HT2 serotonin receptors. J. Med. Chem. 1988, 31, 5–7. [Google Scholar] [CrossRef]
- Appel, M.; Mitchell, M.; Garlick, R.K.; Glennon, R.A.; Teitler, M.; Souza, B.D.E. Autoradiographic Characterization ([1251]DOI) Binding to 5-HT2 and 5-HT1 Receptors in Rat Brain. J. Pharmacol. Exp. Ther. 1990, 233, 843–857. [Google Scholar]
- Zea-Ponce, Y.; Kegeles, L.S.; Guo, N.; Raskin, L.; Bakthavachalam, V.; Laruelle, M. Pharmacokinetics and brain distribution in non human primate of R(-)[123I]DOI, A 5HT2A/2C serotonin agonist. Nucl. Med. Biol. 2002, 29, 575–583. [Google Scholar] [CrossRef]
- Terriere, D.; Janssen, P.M.; Gommeren, W.; Gysemans, M.; Mertens, J.J.; Leysen, J.E. Evaluation of radioiodo-4-amino-N-[1-[3-(4-fluorophenoxy)-propyl]-4- methyl-4-piperidinyl]-5-iodo-2-methoxybenzamide as a potential 5HT2 receptor tracer for SPE(C)T. Nucl. Med. Biol. 1995, 22, 1005–1010. [Google Scholar] [CrossRef]
- Abi-Dargham, A.; Zea-Ponce, Y.; Terriere, D.; Al-Tikriti, M.; Baldwin, R.M.; Hoffer, P.; Charney, D.; Leysen, J.E.; Laruelle, M.; Mertens, J.; et al. Preclinical evaluation of [123I]R93274 as a SPECT radiotracer for imaging 5-HT(2A) receptors. Eur. J. Pharmacol. 1997, 321, 285–293. [Google Scholar] [CrossRef]
- Busatto, G.F.; Pilowsky, L.S.; Costa, D.C.; Mertens, J.; Terriere, D.; Ell, P.J.; Mulligan, R.; Travis, M.J.; Leysen, J.E.; Lui, D.; et al. Initial evaluation of 123I-5-I-R91150, a selective 5-HT2A ligand for single-photon emission tomography, in healthy human subjects. Eur. J. Nucl. Med. 1997, 24, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Catafau, A.M.; Danus, M.; Bullich, S.; Llop, J.; Perich, J.; Cunningham, V.J.; Plaza, P.; Penengo, M.M.; Eersels, J.L.H.; Squassante, L.; et al. Characterization of the SPECT 5-HT2A receptor ligand 123I-R91150 in healthy volunteers: Part 1—Pseudoequilibrium interval and quantification methods. J. Nucl. Med. 2006, 47, 919–928. [Google Scholar]
- Baeken, C.; D’haenen, H.; Flamen, P.; Mertens, J.; Terriere, D.; Chavatte, K.; Boumon, R.; Bossuyt, A. 123I-5-I-R91150, a new single-photon emission tomography ligand for 5-HT(2A) receptors: Influence of age and gender in healthy subjects. Eur. J. Nucl. Med. 1998, 25, 1617–1622. [Google Scholar] [CrossRef]
- Travis, M.J.; Busatto, G.F.; Pilowsky, L.S.; Mulligan, R.; Acton, P.D.; Gacinovic, S.; Mertens, J.; Terrière, D.; Costa, D.C.; Ell, P.J.; et al. 5-HT 2A receptor blockade in patients with schizophrenia treated with risperidone or clozapine. Br. J. Psychiatry 1998, 173, 236–241. [Google Scholar] [CrossRef]
- Melse, M.; Tan, S.K.H.; Temel, Y.; van Kroonenburgh, M.J.P.G.; Leentjens, A.F.G. Changes in 5-HT2A Receptor Expression in Untreated, de novo Patients with Parkinson’s Disease. J. Parkinsons Dis. 2014, 4, 283–287. [Google Scholar] [CrossRef]
- Versijpt, J.; Van Laere, K.J.; Dumont, F.; Decoo, D.; Vandecapelle, M.; Santens, P.; Goethals, I.; Audenaert, K.; Slegers, G.; Dierckx, R.A.; et al. Imaging of the 5-HT2A system: Age-, gender-, and Alzheimer’s disease-related findings. Neurobiol. Aging 2003, 24, 553–561. [Google Scholar] [CrossRef]
- Baeken, C.; De Raedt, R.; Bossuyt, A.; Van Hove, C.; Mertens, J.; Dobbeleir, A.; Blanckaert, P.; Goethals, I. The impact of HF-rTMS treatment on serotonin2A receptors in unipolar melancholic depression. Brain Stimul. 2011, 4, 104–111. [Google Scholar] [CrossRef]
- Vermeire, S.T.; Audenaert, K.R.; Dobbeleir, A.A.; De Meester, R.H.; De Vos, F.J.; Peremans, K.Y. Evaluation of the brain 5-HT2A receptor binding index in dogs with anxiety disorders, measured with 123I-5I-R91150 and SPECT. J. Nucl. Med. 2009, 50, 284–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peremans, K.; De Spiegeleer, B.; Buntinx, E.; Dobbeleir, A.; Vermeire, S.; Vandermeulen, E.; De Vos, F.; Megens, A.; Eersels, J.; Audenaert, K. Evaluation of serotonin-2A receptor occupancy with 123I-5-I-R91150 and single-photon emission tomography before and after low-dose pipamperone administration in the canine brain. Nucl. Med. Commun. 2008, 29, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Goethals, I.; Vervaet, M.; Audenaert, K.; Jacobs, F.; Ham, H.; Van de Wiele, C.; Vandecapelle, M.; Slegers, G.; Dierckx, R.; van Heeringen, C. Differences of cortical 5-HT2A receptor binding index with SPECT in subtypes of anorexia nervosa: Relationship with personality traits? J. Psychiatr. Res. 2007, 41, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Audenaert, K.; Van Laere, K.; Dumont, F.; Vervaet, M.; Goethals, I.; Slegers, G.; Mertens, J.; van Heeringen, C.; Dierckx, R.A. Decreased 5-HT2a receptor binding in patients with anorexia nervosa. J. Nucl. Med. 2003, 44, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Van Heeringen, C.; Audenaert, K.; Van Laere, K.; Dumont, F.; Slegers, G.; Mertens, J.; Dierckx, R.A. Prefrontal 5-HT2a receptor binding index, hopelessness and personality characteristics in attempted suicide. J. Affect. Disord. 2003, 74, 149–158. [Google Scholar] [CrossRef]
- Dumas, N.; Moulin-Sallanon, M.; Ginovart, N.; Tournier, B.B.; Suzanne, P.; Cailly, T.; Fabis, F.; Rault, S.; Charnay, Y.; Millet, P. Small-animal single-photon emission computed tomographic imaging of the brain serotoninergic systems in wild-type and Mdr1a knockout rats. Mol. Imaging 2014, 13, 1–12. [Google Scholar] [CrossRef]
- Mühlhausen, U.; Ermert, J.; Coenen, H.H. Synthesis, labelling and first evaluation of [18F]R91150 as a serotonin 5-HT2A receptor antagonist for PET. J. Label. Compd. Radiopharm. 2009, 52, 13–22. [Google Scholar] [CrossRef]
- Samnick, S.; Remy, N.; Ametamey, S.; Bader, J.B.; Brandau, W.; Kirsch, C.-M. 123I-MSP and F[1 1C]MSP: New selective 5-HT2A receptor radiopharmaceuticals for in vivo studies of neuronal 5-HT2 serotonin receptors. Synthesis, in vitro binding study with unlabelled analogues and preliminary in vivo evaluation in mice. Life Sci. 1998, 63, 2001–2013. [Google Scholar] [CrossRef]
- Blanckaert, P.B.M.; Burvenich, I.; Wyffels, L.; De Bruyne, S.; Moerman, L.; De Vos, F. In vivo evaluation in rodents of [123I]-3-I-CO as a potential SPECT tracer for the serotonin 5-HT2A receptor. Nucl. Med. Biol. 2008, 35, 861–867. [Google Scholar] [CrossRef]
- Lummis, S.C.R. 5-HT 3 Receptors. J. Biol. Chem. 2012, 287, 40239–40245. [Google Scholar] [CrossRef] [Green Version]
- Niesler, B.; Kapeller, J.; Hammer, C.; Rappold, G. Serotonin type 3 receptor genes: HTR3A, B, C, D, E. Pharmacogenomics 2008, 9, 501–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilpatrick, G.J.; Jones, B.J.; Tyers, M.B. Identification and distribution of 5-HT3 receptors in rat brain using radioligand binding. Nature 1987, 330, 746–748. [Google Scholar] [CrossRef] [PubMed]
- Parker, R.M.C.; Barnes, J.M.; Ge, J.; Barber, P.C.; Barnes, N.M. Autoradiographic distribution of [3H]-(S)-zacopride-labelled 5-HT3 receptors in human brain. J. Neurol. Sci. 1996, 144, 119–127. [Google Scholar] [CrossRef]
- Thompson, A.J.; Lummis, S.C.R. The 5-HT3 receptor as a therapeutic target. Expert Opin. Ther. Targets 2007, 11, 527–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walstab, J.; Rappold, G.; Niesler, B. 5-HT3 receptors: Role in disease and target of drugs. Pharmacol. Ther. 2010, 128, 146–169. [Google Scholar] [CrossRef] [PubMed]
- Fakhfouri, G.; Rahimian, R.; Dyhrfjeld-Johnsen, J.; Zirak, M.R.; Beaulieu, J.M. 5-HT3 receptor antagonists in neurologic and neuropsychiatric disorders: The iceberg still lies beneath the surface. Pharmacol. Rev. 2019, 71, 383–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, A.J. Recent developments in 5-HT3 receptor pharmacology. Trends Pharmacol. Sci. 2013, 34, 100–109. [Google Scholar] [CrossRef]
- Paterson, L.M.; Kornum, B.R.; Nutt, D.J.; Pike, V.W.; Knudsen, G.M. 5-HT radioligands for human brain imaging with PET and SPECT. Med. Res. Rev. 2013, 33, 54–111. [Google Scholar] [CrossRef] [Green Version]
- Camsonne, R.; Barre, L.; Petit-Taboué, M.C.; Travére, J.M.; Jones, R.; Debruyne, D.; Moulin, M.A.; MacKenzie, E.T.; Baron, J.C. Positron emission tomographic studies of [11C]MDL 72222, a potential 5-HT3 receptor radioligand: Distribution, kinetics and binding in the brain of the baboon. Neuropharmacology 1993, 32, 65–71. [Google Scholar] [CrossRef]
- Ishiwata, K.; Saito, N.; Yanagawa, K.; Furuta, R.; Ishii, S.I.; Kiyosawa, M.; Homma, Y.; Ishii, K.; Suzuki, F.; Senda, M. Synthesis and evaluation of 5-HT3 receptor antagonist [11C]KF17643. Nucl. Med. Biol. 1996, 23, 285–290. [Google Scholar] [CrossRef]
- Besret, L.; Dauphin, F.; Guillouet, G.; Dhilly, M.; Gourand, F.; Blaizot, X.; Young, A.R.; Petit-taboué, M.C.; Mickala, P.; Barbelivien, A.; et al. [11C] S21OO7, a Putative Partial Agonist for 5-HT3 Receptors PET Studies. Rat and Primate in vio Biological Evaluation. Life Sci. 1998, 62, 115–129. [Google Scholar] [CrossRef]
- Katounina, T.; Besret, L.; Dhilly, M.; Petit-Taboué, M.C.; Barbelivien, A.; Baron, J.C.; Dauphin, F.; Barré, L. Synthesis and biological investigations of [18F]MR18445, a 5-HT3 receptor partial agonist. Bioorg. Med. Chem. 1998, 6, 789–795. [Google Scholar] [CrossRef]
- Smith, W.W.; Sancilio, L.F.; Owera-Atepo, J.B.; Naylor, R.J.; Lambert, L. Zacopride, a potent 5-HT3 antagonist. J. Pharm. Pharmacol. 1988, 301–302. [Google Scholar] [CrossRef] [PubMed]
- Hewlett, W.A.; Fridman, S.; Trivedi, B.L.; Schmidt, D.E.; De Paulis, T.; Ebert, M.H. Characterization of desamino-5-[125I]iodo-3-methoxy-zacopride ([125I]MIZAC) binding to 5-HT3 receptors in the rat brain. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1998, 22, 397–410. [Google Scholar] [CrossRef]
- Hewlett, W.A.; Trivedi, B.L.; Zhang, Z.; Lovinger, D.M.; Ansari, M.S.I.B.; Ebert, M.H.; Schmidt, D.E. Characterization of (S)-Des-4-amino-3-[125I]iodozacopride ([125I]DAIZAC), a Selective High-Affinity Radioligand for 5-hydroxytryptamine3 receptors. J. Pharmacol. Exp. Ther. 1999, 288, 221–231. [Google Scholar]
- Pithia, N.K.; Liang, C.; Pan, X.Z.; Pan, M.L.; Mukherjee, J. Synthesis and evaluation of (S)-[18F]fesetron in the rat brain as a potential PET imaging agent for serotonin 5-HT3 receptors. Bioorg. Med. Chem. Lett. 2016, 26, 1919–1924. [Google Scholar] [CrossRef] [Green Version]
- Mu, L.; Müller Herde, A.; Rüefli, P.M.; Sladojevich, F.; Milicevic Sephton, S.; Krämer, S.D.; Thompson, A.J.; Schibli, R.; Ametamey, S.M.; Lochner, M. Synthesis and Pharmacological Evaluation of [11C]Granisetron and [18F]Fluoropalonosetron as PET Probes for 5-HT3 Receptor Imaging. ACS Chem. Neurosci. 2016, 7, 1552–1564. [Google Scholar] [CrossRef]
- Ishiwata, K.; Ishii, K.; Ishii, S.I.; Senda, M. Synthesis of 5-HT3 receptor antagonists, [11C]Y-25130 and [11C]YM060. Appl. Radiat. Isot. 1995, 46, 907–910. [Google Scholar] [CrossRef]
- Thorell, J.O.; Stone-Elander, S.; Eriksson, L.; Ingvar, M. N-methylquipazine: Carbon-11 labelling of the 5-HT3 agonist and in vivo evaluation of its biodistribution using PET. Nucl. Med. Biol. 1997, 24, 405–412. [Google Scholar] [CrossRef]
- Gao, M.; Wang, M.; Hutchins, G.D.; Zheng, Q.H. Synthesis of new carbon-11 labeled benzoxazole derivatives for PET imaging of 5-HT3 receptor. Eur. J. Med. Chem. 2008, 43, 1570–1574. [Google Scholar] [CrossRef]
- Dumuis, A.; Bouhelal, R.; Sebben, M.; Cory, R.; Bockaert, J. A nonclassical 5-hydroxytryptamine receptor positively coupled with adenylate cyclase in the central nervous system. Mol. Pharmacol. 1988, 34, 880–887. [Google Scholar] [PubMed]
- Bockaert, J.; Claeysen, S.; Compan, V.; Dumuis, A. 5-HT4 receptors: History, molecular pharmacology and brain functions. Neuropharmacology 2008, 55, 922–931. [Google Scholar] [CrossRef] [PubMed]
- Hegde, S.S.; Eglen, R.M. Peripheral 5-HT 4 receptors. FASEB J. 1996, 10, 1398–1407. [Google Scholar] [CrossRef] [PubMed]
- Bockaert, J.; Claeysen, S.; Compan, V.; Dumuis, A. 5-HT4 receptors, a place in the sun: Act two. Curr. Opin. Pharmacol. 2011, 11, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Hagena, H.; Manahan-Vaughan, D. The serotonergic 5-HT4 receptor: A unique modulator of hippocampal synaptic information processing and cognition. Neurobiol. Learn. Mem. 2017, 138, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Claeysen, S.; Bockaert, J.; Giannoni, P. Serotonin: A New Hope in Alzheimer’s Disease? ACS Chem. Neurosci. 2015, 6, 940–943. [Google Scholar] [CrossRef] [Green Version]
- Lezoualc’h, F. 5-HT4 receptor and Alzheimer’s disease: The amyloid connection. Exp. Neurol. 2007, 205, 325–329. [Google Scholar] [CrossRef]
- Jean, A.; Conductier, G.; Manrique, C.; Bouras, C.; Berta, P.; Hen, R.; Charnay, Y.; Bockaert, J.; Compan, V. Anorexia induced by activation of serotonin 5-HT4 receptors is mediated by increases in CART in the nucleus accumbens. Proc. Natl. Acad. Sci. USA 2007, 104, 16335–16340. [Google Scholar] [CrossRef] [Green Version]
- Lucas, G.; Rymar, V.V.; Du, J.; Mnie-Filali, O.; Bisgaard, C.; Manta, S.; Lambas-Senas, L.; Wiborg, O.; Haddjeri, N.; Piñeyro, G.; et al. Serotonin4 (5-HT4) Receptor Agonists Are Putative Antidepressants with a Rapid Onset of Action. Neuron 2007, 55, 712–725. [Google Scholar] [CrossRef] [Green Version]
- Kaumann, A.J.; Gaster, L.M.; King, F.D.; Brown, A.M. Blockade of human atrial 5-HT4 receptors by SB 207710, a selective and high affinity 5-HT4 receptor antagonist. Naunyn Schmiedebergs Arch. Pharmacol. 1994, 349, 546–548. [Google Scholar] [CrossRef]
- Varnäs, K.; Halldin, C.; Pike, V.W.; Hall, H. Distribution of 5-HT4 receptors in the postmortem human brain—An autoradiographic study using [125I]SB 207710. Eur. Neuropsychopharmacol. 2003, 13, 228–234. [Google Scholar] [CrossRef]
- Patel, S.; Roberts, J.; Moorman, J.; Reavill, C. Localization of serotonin-4 receptors in the striatonigral pathway in rat brain. Neuroscience 1995, 69, 1159–1167. [Google Scholar] [CrossRef]
- Pike, V.W.; Halldin, C.; Nobuhara, K.; Hiltunen, J.; Mulligan, R.S.; Swahn, C.G.; Karlsson, P.; Olsson, H.; Hume, S.P.; Hirani, E.; et al. Radioiodinated SB 207710 as a radioligand in vivo: Imaging of brain 5-HT4 receptors with SPET. Eur. J. Nucl. Med. Mol. Imaging 2003, 30, 1520–1528. [Google Scholar] [CrossRef] [PubMed]
- Gee, A.D.; Martarello, L.; Passchier, J.; Wishart, M.; Parker, C.; Matthews, J.; Comley, R.; Hopper, R.; Gunn, R. Synthesis and evaluation of [11C]SB207145 as the first in vivo serotonin 5-HT4 receptor radioligand for PET imaging in man. Curr. Radiopharm. 2008, 1, 110–114. [Google Scholar] [CrossRef]
- Kornum, B.R.; Lind, N.M.; Gillings, N.; Marner, L.; Andersen, F.; Knudsen, G.M. Evaluation of the novel 5-HT4 receptor PET ligand [11C]SB207145 in the Göttingen minipig. J. Cereb. Blood Flow Metab. 2009, 29, 186–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marner, L.; Gillings, N.; Comley, R.A.; Baaré, W.F.C.; Rabiner, E.A.; Wilson, A.A.; Houle, S.; Hasselbalch, S.G.; Svarer, C.; Gunn, R.N.; et al. Kinetic modeling of11C-SB207145 binding to 5-HT4 receptors in the human brain in vivo. J. Nucl. Med. 2009, 50, 900–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marner, L.; Gillings, N.; Madsen, K.; Erritzoe, D.; Baaré, W.F.C.; Svarer, C.; Hasselbalch, S.G.; Knudsen, G.M. Brain imaging of serotonin 4 receptors in humans with [11C]SB207145-PET. Neuroimage 2010, 50, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Madsen, K.; Marner, L.; Haahr, M.; Gillings, N.; Knudsen, G.M. Mass dose effects and in vivo affinity in brain PET receptor studies—A study of cerebral 5-HT 4 receptor binding with [ 11C]SB207145. Nucl. Med. Biol. 2011, 38, 1085–1091. [Google Scholar] [CrossRef]
- Haahr, M.E.; Fisher, P.M.; Jensen, C.G.; Frokjaer, V.G.; McMahon, B.; Madsen, K.; Baaré, W.F.C.; Lehel, S.; Norremolle, A.; Rabiner, E.A.; et al. Central 5-HT4 receptor binding as biomarker of serotonergic tonus in humans: A [11C]SB207145 PET study. Mol. Psychiatry 2014, 19, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Perfalk, E.; da Cunha-Bang, S.; Holst, K.K.; Keller, S.; Svarer, C.; Knudsen, G.M.; Frokjaer, V.G. Testosterone levels in healthy men correlate negatively with serotonin 4 receptor binding. Psychoneuroendocrinology 2017, 81, 22–28. [Google Scholar] [CrossRef]
- Jakobsen, G.R.; Fisher, P.M.; Dyssegaard, A.; McMahon, B.; Holst, K.K.; Lehel, S.; Svarer, C.; Jensen, P.S.; Knudsen, G.M.; Frokjaer, V.G. Brain serotonin 4 receptor binding is associated with the cortisol awakening response. Psychoneuroendocrinology 2016, 67, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Deen, M.; Hansen, H.D.; Hougaard, A.; Nørgaard, M.; Eiberg, H.; Lehel, S.; Ashina, M.; Knudsen, G.M. High brain serotonin levels in migraine between attacks: A 5-HT4 receptor binding PET study. NeuroImage Clin. 2018, 18, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Hong, J.; Morse, C.L.; Pike, V.W. Synthesis, structure-affinity relationships, and radiolabeling of selective high-affinity 5-HT4 receptor ligands as prospective imaging probes for positron emission tomography. J. Med. Chem. 2010, 53, 7035–7047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terry, G.E.; Hirvonen, J.; Liow, J.S.; Zoghbi, S.S.; Gladding, R.; Tauscher, J.T.; Schaus, J.M.; Phebus, L.; Felder, C.C.; Morse, C.L.; et al. Imaging and quantitation of cannabinoid CB1 receptors in human and monkey brains using 18F-labeled inverse agonist radioligands. J. Nucl. Med. 2010, 51, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Lohith, T.G.; Xu, R.; Tsujikawa, T.; Morse, C.L.; Anderson, K.B.; Gladding, R.L.; Zoghbi, S.S.; Fujita, M.; Innis, R.B.; Pike, V.W. Evaluation in monkey of two candidate PET radioligands, [11C]RX-1 and [18F]RX-2, for imaging brain 5-HT4 receptors. Synapse 2014, 68, 613–623. [Google Scholar] [CrossRef]
- Caillé, F.; Morley, T.J.; Tavares, A.A.S.; Papin, C.; Twardy, N.M.; Alagille, D.; Lee, H.S.; Baldwin, R.M.; Seibyl, J.P.; Barret, O.; et al. Synthesis and biological evaluation of positron emission tomography radiotracers targeting serotonin 4 receptors in brain: [18F]MNI-698 and [18F]MNI-699. Bioorg. Med. Chem. Lett. 2013, 23, 6243–6247. [Google Scholar] [CrossRef]
- Tavares, A.A.S.; Caillé, F.; Barret, O.; Papin, C.; Lee, H.; Morley, T.J.; Fowles, K.; Holden, D.; Seibyl, J.P.; Alagille, D.; et al. Whole-body biodistribution and dosimetry estimates of a novel radiotracer for imaging of serotonin 4 receptors in brain: [18F]MNI-698. Nucl. Med. Biol. 2014, 41, 432–439. [Google Scholar] [CrossRef]
- Tavares, A.A.S.; Caillé, F.; Barret, O.; Papin, C.; Lee, H.; Morley, T.J.; Fowles, K.; Holden, D.; Seibyl, J.P.; Alagille, D.; et al. In vivo evaluation of 18F-MNI698: An 18F-labeled radiotracer for imaging of serotonin 4 receptors in brain. J. Nucl. Med. 2014, 55, 858–864. [Google Scholar] [CrossRef] [Green Version]
- Fresneau, N.; Dumas, N.; Tournier, B.B.; Fossey, C.; Ballandonne, C.; Lesnard, A.; Millet, P.; Charnay, Y.; Cailly, T.; Bouillon, J.P.; et al. Design of a serotonin 4 receptor radiotracer with decreased lipophilicity for single photon emission computed tomography. Eur. J. Med. Chem. 2015, 94, 386–396. [Google Scholar] [CrossRef]
- Monsma, F.J.; Shen, Y.; Ward, R.P.; Hamblin, M.W.; Sibley, D.R. Cloning and expression of a novel serotonin receptor with high affinity for tricyclic psychotropic drugs. Mol. Pharmacol. 1993, 43, 320–327. [Google Scholar]
- Ruat, M.; Traiffort, E.; Leurs, R.; Tardivel-Lacombe, J.; Diaz, J.; Arrang, J.M.; Schwartz, J.C. Molecular cloning, characterization, and localization of a high-affinity serotonin receptor (5-HT7) activating cAMP formation. Proc. Natl. Acad. Sci. USA 1993, 90, 8547–8551. [Google Scholar] [CrossRef] [Green Version]
- Kohen, R.; Metcalf, M.A.; Khan, N.; Druck, T.; Huebner, K.; Lachowicz, J.E.; Meltzer, H.Y.; Sibley, D.R.; Roth, B.L.; Hamblin, M.W. Cloning, characterization, and chromosomal localization of a human 5-HT6 serotonin receptor. J. Neurochem. 1996, 66, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, M.; Matsumoto, M.; Togashi, H.; Mori, K.; Saito, H. Central distribution and function of 5-HT6 receptor subtype in the rat brain. Life Sci. 1998, 62, 1473–1477. [Google Scholar] [CrossRef]
- Ward, R.P.; Hamblin, M.W.; Lachowicz, J.E.; Hoffman, B.J.; Sibley, D.R.; Dorsa, D.M. Localization of serotonin subtype 6 receptor messenger RNA in the rat brain by in situ hybridization histochemistry. Neuroscience 1995, 64, 1105–1111. [Google Scholar] [CrossRef]
- Gérard, C.; Martres, M.P.; Lefèvre, K.; Miquel, M.C.; Vergé, D.; Lanfumey, L.; Doucet, E.; Hamon, M.; El Mestikawy, S. Immune-localization of serotonin 5-HT6 receptor-like material in the rat central nervous system. Brain Res. 1997, 746, 207–219. [Google Scholar] [CrossRef]
- Roberts, J.C.; Reavill, C.; East, S.Z.; Harrison, P.J.; Patel, S.; Routledge, C.; Leslie, R.A. The distribution of 5-HT6 receptors in rat brain: An autoradiographic binding study using the radiolabelled 5-HT6 receptor antagonist [125I]SB-258585. Brain Res. 2002, 934, 49–57. [Google Scholar] [CrossRef]
- East, S.Z.; Burnet, P.W.J.; Leslie, R.A.; Roberts, J.C.; Harrison, P.J. 5-HT6 receptor binding sites in schizophrenia and following antipsychotic drug administration: Autoradiographic studies with [125I]SB-258585. Synapse 2002, 45, 191–199. [Google Scholar] [CrossRef]
- Barnes, N.M.; Ahern, G.P.; Becamel, C.; Bockaert, J.; Camilleri, M.; Chaumont-Dubel, S.; Claeysen, S.; Cunningham, K.A.; Fone, K.C.; Gershon, M.; et al. International union of basic and clinical pharmacology. Cx. Classification of receptors for 5-hydroxytryptamine; pharmacology and function. Pharmacol. Rev. 2021, 73, 310–520. [Google Scholar] [CrossRef]
- Meffre, J.; Chaumont-Dubel, S.; Mannoury la Cour, C.; Loiseau, F.; Watson, D.J.G.; Dekeyne, A.; Séveno, M.; Rivet, J.M.; Gaven, F.; Déléris, P.; et al. 5-HT6 receptor recruitment of mTOR as a mechanism for perturbed cognition in schizophrenia. EMBO Mol. Med. 2012, 4, 1043–1056. [Google Scholar] [CrossRef]
- Karila, D.; Freret, T.; Bouet, V.; Boulouard, M.; Dallemagne, P.; Rochais, C. Therapeutic Potential of 5-HT6 Receptor Agonists. J. Med. Chem. 2015, 58, 7901–7912. [Google Scholar] [CrossRef]
- Voigt, J.P.; Fink, H. Serotonin controlling feeding and satiety. Behav. Brain Res. 2015, 277, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.; Tousi, B.; Sabbagh, M.N. 5HT6 Antagonists in the Treatment of Alzheimer’s Dementia: Current Progress. Neurol. Ther. 2018, 7, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoury, R.; Grysman, N.; Gold, J.; Patel, K.; Grossberg, G.T. The role of 5-HT6-receptor antagonists in Alzheimer’s disease: An update. Expert Opin. Investig. Drugs 2018, 27, 523–533. [Google Scholar] [CrossRef]
- Ferrero, H.; Solas, M.; Francis, P.T.; Ramirez, M.J. Serotonin 5-HT6 Receptor Antagonsits in Alzheimer’s Disease: Therapeutic rationale and Current Development Status. CNS Drugs 2017, 31, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Hirst, W.D.; Minton, J.A.L.; Bromidge, S.M.; Moss, S.F.; Latter, A.J.; Riley, G.; Routledge, C.; Middlemiss, D.N.; Price, G.W. Characterization of [125I]-SB-258585 binding to human recombinant and native 5-HT6 receptors in rat, pig and human brain tissue. Br. J. Pharmacol. 2000, 130, 1597–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, S.; Verdurand, M.; Joseph, B.; Lemoine, L.; Daoust, A.; Billard, T.; Fournet, G.; Le Bars, D.; Zimmer, L. Synthesis and biological evaluation in rat and cat of [18F]12ST05 as a potential 5-HT6 PET radioligand. Nucl. Med. Biol. 2007, 34, 995–1002. [Google Scholar] [CrossRef]
- Zhou, P.; Yan, Y.; Bernotas, R.; Harrison, B.L.; Huryn, D.; Robichaud, A.J.; Zhang, G.M.; Smith, D.L.; Schechter, L.E. 4-(2-Aminoethoxy)-N-(phenylsulfonyl)indoles as novel 5-HT6 receptor ligands. Bioorg. Med. Chem. Lett. 2005, 15, 1393–1396. [Google Scholar] [CrossRef]
- Parker, C.A.; Gunn, R.N.; Rabiner, E.A.; Slifstein, M.; Comley, R.; Salinas, C.; Johnson, C.N.; Jakobsen, S.; Houle, S.; Laruelle, M.; et al. Radiosynthesis and characterization of 11C-GSK215083 as a PET radioligand for the 5-HT6 receptor. J. Nucl. Med. 2012, 53, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Parker, C.A.; Rabiner, E.A.; Gunn, R.N.; Searle, G.; Martarello, L.; Comley, R.A.; Davy, M.; Wilson, A.A.; Houle, S.; Mizrahi, R.; et al. Human kinetic modeling of the 5HT6 PET radioligand 11C-GSK215083 and its utility for determining occupancy at both 5HT6 and 5HT2A receptors by SB742457 as a potential therapeutic mechanism of action in Alzheimer disease. J. Nucl. Med. 2015, 56, 1901–1909. [Google Scholar] [CrossRef] [Green Version]
- Rosse, G. Quinoline Derivatives as 5-HT 6 Receptor PET Ligands. ACS Med. Chem. Lett. 2014, 5, 275–276. [Google Scholar] [CrossRef] [Green Version]
- Colomb, J.; Becker, G.; Fieux, S.; Zimmer, L.; Billard, T. Syntheses, radiolabelings, and in vitro evaluations of fluorinated pet radioligands of 5-HT6 serotoninergic receptors. J. Med. Chem. 2014, 57, 3884–3890. [Google Scholar] [CrossRef] [PubMed]
- Emery, S.; Fieux, S.; Vidal, B.; Courault, P.; Bouvard, S.; Tourvieille, C.; Iecker, T.; Billard, T.; Zimmer, L.; Lancelot, S. Preclinical validation of [18F]2FNQ1P as a specific PET radiotracer of 5-HT6 receptors in rat, pig, non-human primate and human brain tissue. Nucl. Med. Biol. 2020, 82–83, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Bard, J.A.; Zgombick, J.; Adham, N.; Vaysse, P.; Branchek, T.A.; Weinshank, R.L. Cloning of a novel human serotonin receptor (5-HT7) positively linked to adenylate cyclase. J. Biol. Chem. 1993, 268, 23422–23426. [Google Scholar] [CrossRef]
- Lovenberg, T.W.; Baron, B.M.; de Lecea, L.; Miller, J.D.; Prosser, R.A.; Rea, M.A.; Foye, P.E.; Racke, M.; Slone, A.L.; Siegel, B.W. A novel adenylyl cyclase-activating serotonin receptor (5-HT7) implicated in the regulation of mammalian circadian rhythms. Neuron 1993, 11, 449–458. [Google Scholar] [CrossRef]
- Varnäs, K.; Thomas, D.R.; Tupala, E.; Tiihonen, J.; Hall, H. Distribution of 5-HT7 receptors in the human brain: A preliminary autoradiographic study using [3H]SB-269970. Neurosci. Lett. 2004, 367, 313–316. [Google Scholar] [CrossRef]
- Cifariello, A.; Pompili, A.; Gasbarri, A. 5-HT7 receptors in the modulation of cognitive processes. Behav. Brain Res. 2008, 195, 171–179. [Google Scholar] [CrossRef]
- L’Estrade, E.T.; Erlandsson, M.; Edgar, F.G.; Ohlsson, T.; Knudsen, G.M.; Herth, M.M. Towards selective CNS PET imaging of the 5-HT7 receptor system: Past, present and future. Neuropharmacology 2020, 172, 107830–107848. [Google Scholar] [CrossRef]
- Kikuchi, C.; Nagaso, H.; Hiranuma, T.; Koyama, M. Tetrahydrobenzindoles: Selective Antagonists of the 5-HT 7 Receptor. J. Med. Chem. 1999, 42, 533–535. [Google Scholar] [CrossRef]
- Kikuchi, C.; Hiranuma, T.; Koyama, M. Tetrahydrothienopyridylbutyl-tetrahydrobenzindoles: New selective ligands of the 5-HT7 receptor. Bioorg. Med. Chem. Lett. 2002, 12, 2549–2552. [Google Scholar] [CrossRef]
- Zhang, M.R.; Haradahira, T.; Maeda, J.; Okauchi, T.; Kida, T.; Obayashi, S.; Suzuki, K.; Suhara, T. Synthesis and preliminary PET study of the 5-HT7 receptor antagonist [11C]DR4446. J. Label. Compd. Radiopharm. 2002, 45, 857–866. [Google Scholar] [CrossRef]
- Herth, M.M.; Volk, B.; Pallagi, K.; Kofoed Bech, L.; Antoni, F.A.; Knudsen, G.M.; Kristensen, J.L. Synthesis and in vitro evaluation of oxindole derivatives as potential radioligands for 5-HT7 receptor imaging with PET. ACS Chem. Neurosci. 2012, 3, 1002–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemoine, L.; Andries, J.; Le Bars, D.; Billard, T.; Zimmer, L. Comparison of 4 radiolabeled antagonists for serotonin 5-HT(7) receptor neuroimaging: Toward the first PET radiotracer. J. Nucl. Med. 2011, 52, 1811–1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tampio L’Estrade, E.; Xiong, M.; Shalgunov, V.; Edgar, F.G.; Volk, B.; Baerentzen, S.L.; Palner, M.; Erlandsson, M.; Ohlsson, T.; Knudsen, G.M.; et al. Development and Evaluation of Two Potential 5-HT7 Receptor PET Tracers: [18F]ENL09 and [18F]ENL10. ACS Chem. Neurosci. 2019, 10, 3961–3968. [Google Scholar] [CrossRef] [PubMed]
- Hansen, H.D.; Constantinescu, C.C.; Barret, O.; Herth, M.M.; Magnussen, J.H.; Lehel, S.; Dyssegaard, A.; Colomb, J.; Billard, T.; Zimmer, L.; et al. Evaluation of [18F]2FP3 in pigs and non-human primates. J. Label. Compd. Radiopharm. 2019, 62, 34–42. [Google Scholar] [CrossRef]
- Colomb, J.; Becker, G.; Forcellini, E.; Meyer, S.; Buisson, L.; Zimmer, L.; Billard, T. Synthesis and pharmacological evaluation of a new series of radiolabeled ligands for 5-HT7 receptor PET neuroimaging. Nucl. Med. Biol. 2014, 41, 330–337. [Google Scholar] [CrossRef]
- L’Estrade, E.T.; Edgar, F.G.; Xiong, M.; Shalgunov, V.; Baerentzen, S.L.; Erlandsson, M.; Ohlsson, T.G.; Palner, M.; Knudsen, G.M.; Herth, M.M. Synthesis, Radiolabeling, and in Vitro and in Vivo Evaluation of [18F]ENL30: A Potential PET Radiotracer for the 5-HT 7 Receptor. ACS Omega 2019, 4, 7344–7353. [Google Scholar] [CrossRef] [Green Version]
- L’Estrade, E.T.; Shalgunov, V.; Edgar, F.G.; Strebl-Bantillo, M.G.; Xiong, M.; Crestey, F.; Neelamegam, R.; Dyssegaard, A.; Lehel, S.; Erlandsson, M.; et al. Radiosynthesis and preclinical evaluation of [11C]Cimbi-701—Towards the imaging of cerebral 5-HT7 receptors. J. Label. Compd. Radiopharm. 2020, 63, 46–55. [Google Scholar] [CrossRef]
- Rudnick, G. Serotonin transporters—Structure and function. J. Membr. Biol. 2006, 213, 101–110. [Google Scholar] [CrossRef]
- Suehiro, M.; Scheffel, U.; Ravert, H.T.; Dannals, R.F.; Wagner, H.N. [11C](+)McN5652 as a radiotracer for imaging serotonin uptake sites with pet. Life Sci. 1993, 53, 883–892. [Google Scholar] [CrossRef]
- Szabo, Z.; Scheffel, U.; Suehiro, M.; Dannais, R.F.; Kim, S.E.; Ravert, H.T.; Ricaurte, G.A.; Wagner, H.N. Positron emission tomography of 5-HT transporter sites in the baboon brain with [11C]McN5652. J. Cereb. Blood Flow Metab. 1995, 15, 798–805. [Google Scholar] [CrossRef]
- Parsey, R.V.; Kegeles, L.S.; Hwang, D.R.; Simpson, N.; Abi-Dargham, A.; Mawlawi, O.; Slifstein, M.; Van Heertum, R.L.; Mann, J.J.; Laruelle, M. In vivo quantification of brain serotonin transporters in humans using [11C]McN 5652. J. Nucl. Med. 2000, 41, 1465–1477. [Google Scholar] [PubMed]
- Ichimiya, T.; Suhara, T.; Sudo, Y.; Okubo, Y.; Nakayama, K.; Nankai, M.; Inoue, M.; Yasuno, F.; Takano, A.; Maeda, J.; et al. Serotonin transporter binding in patients with mood disorders: A PET study with [11C](+)McN5652. Biol. Psychiatry 2002, 51, 715–722. [Google Scholar] [CrossRef]
- Simpson, H.B.; Lombardo, I.; Slifstein, M.; Huang, H.Y.; Hwang, D.R.; Abi-Dargham, A.; Liebowitz, M.R.; Laruelle, M. Serotonin transporters in obsessive-compulsive disorder: A positron emission tomography study with [11C]McN 5652. Biol. Psychiatry 2003, 54, 1414–1421. [Google Scholar] [CrossRef]
- Reivich, M.; Amsterdam, J.D.; Brunswick, D.J.; Yann Shiue, C. PET brain imaging with [11C](+)McN5652 shows increased serotonin transporter availability in major depression. J. Affect. Disord. 2004, 82, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Marjamäki, P.; Zessin, J.; Eskola, O.; Grönroos, T.; Haaparanta, M.; Bergman, J.; Lehikoinen, P.; Forsback, S.; Brust, P.; Steinbach, J.; et al. S-[18F]fluoromethyl-(+)-McN5652, a PET tracer for the serotonin transporter: Evaluation in rats. Synapse 2003, 47, 45–53. [Google Scholar] [CrossRef]
- Brust, P.; Zessin, J.; Kuwabara, H.; Pawelke, B.; Kretzschmar, M.; Hinz, R.; Bergman, J.; Eskola, O.; Solin, O.; Steinbach, J.; et al. Positron emission tomography imaging of the serotonin transporter in the pig brain using [11C](+)-McN5652 and S-([18F]fluoromethyl)-(+)-McN5652. Synapse 2003, 47, 143–151. [Google Scholar] [CrossRef]
- Brust, P.; Hinz, R.; Kuwabara, H.; Hesse, S.; Zessin, J.; Pawelke, B.; Stephan, H.; Bergmann, R.; Steinbach, J.; Sabri, O. In vivo Measurement of the Serotonin Transporter with (S)-([18F]fluoromethyl)-(+)-McN5652. Neuropsychopharmacology 2003, 28, 2010–2019. [Google Scholar] [CrossRef]
- Oya, S.; Kung, M.P.; Acton, P.D.; Mu, M.; Hou, C.; Kung, H.F. A new single-photon emission computed tomography imaging agent for serotonin transporters: [123I]IDAM, 5-Iodo-2-((2- ((dimethylamino)methyl)phenyl)thio)benzyl alcohol. J. Med. Chem. 1999, 42, 333–335. [Google Scholar] [CrossRef]
- Wilson, A.A.; Ginovart, N.; Schmidt, M.; Meyer, J.H.; Threlkeld, P.G.; Houle, S. Novel radiotracers for imaging the serotonin transporter by positron emission tomography: Synthesis, radiosynthesis, and in vitro and ex vivo evaluation of 11C-labeled 2-(phenylthio)araalkylamines. J. Med. Chem. 2000, 43, 3103–3110. [Google Scholar] [CrossRef] [Green Version]
- Houle, S.; Ginovart, N.; Hussey, D.; Meyer, J.H.; Wilson, A.A. Imaging the serotonin transporter with positron emission tomography: Initial human studies with [11C]DAPP and [11C]DASB. Eur. J. Nucl. Med. 2000, 27, 1719–1722. [Google Scholar] [CrossRef]
- Szabo, Z.; McCann, U.D.; Wilson, A.A.; Scheffel, U.; Owonikoko, T.; Mathews, W.B.; Ravert, H.T.; Hilton, J.; Dannals, R.F.; Ricaurte, G.A. Comparison of (+)-11C-McN5652 and 11C-DASB as serotonin transporter radioligands under various experimental conditions. J. Nucl. Med. 2002, 43, 678–692. [Google Scholar] [PubMed]
- Frankle, W.G.; Huang, Y.; Hwang, D.R.; Talbot, P.S.; Slifstein, M.; Van Heertum, R.; Abi-Dargham, A.; Laruelle, M. Comparative evaluation of serotonin transporter radioligands 11C-DASB and 11C-McN 5652 in healthy humans. J. Nucl. Med. 2004, 45, 682–694. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, M.J.; Simpson, N.R.; Wang, T.; Van Heertum, R.L.; Mann, J.J.; Parsey, R.V. Biodistribution and radiation dosimetry of [11C]DASB in baboons. Nucl. Med. Biol. 2004, 31, 1097–1102. [Google Scholar] [CrossRef]
- Wilson, A.A.; Ginovart, N.; Hussey, D.; Meyer, J.; Houle, S. In vitro and in vivo characterisation of [11C]-DASB: A probe for in vivo measurements of the serotonin transporter by positron emission tomography. Nucl. Med. Biol. 2004, 31, 1097–1102. [Google Scholar] [CrossRef]
- Ginovart, N.; Wilson, A.A.; Meyer, J.H.; Hussey, D.; Houle, S. [11C]-DASB, a tool for in vivo measurement of SSRI-induced occupancy of the serotonin transporter: PET characterization and evaluation in cats. Synapse 2003, 47, 123–133. [Google Scholar] [CrossRef]
- Meyer, J.H.; Wilson, A.A.; Ginovart, N.; Goulding, V.; Hussey, D.; Hood, K.; Houle, S. Occupancy of serotonin transporters by paroxetine and citalopram during treatment of depression: A [11C]DASB PET imaging study. Am. J. Psychiatry 2001, 158, 1843–1849. [Google Scholar] [CrossRef]
- Cannon, D.M.; Ichise, M.; Rollis, D.; Klaver, J.M.; Gandhi, S.K.; Charney, D.S.; Manji, H.K.; Drevets, W.C. Elevated Serotonin Transporter Binding in Major Depressive Disorder Assessed Using Positron Emission Tomography and [11C]DASB; Comparison with Bipolar Disorder. Biol. Psychiatry 2007, 62, 870–877. [Google Scholar] [CrossRef]
- Brown, A.K.; George, D.T.; Fujita, M.; Liow, J.S.; Ichise, M.; Hibbeln, J.; Ghose, S.; Sangare, J.; Hommer, D.; Innis, R.B. PET [11C]DASB imaging of serotonin transporters in patients with alcoholism. Alcohol. Clin. Exp. Res. 2007, 31, 28–32. [Google Scholar] [CrossRef]
- Matsumoto, R.; Ichise, M.; Ito, H.; Ando, T.; Takahashi, H.; Ikoma, Y.; Kosaka, J.; Arakawa, R.; Fujimura, Y.; Ota, M.; et al. Reduced serotonin transporter binding in the insular cortex in patients with obsessive-compulsive disorder: A [11C]DASB PET study. Neuroimage 2010, 49, 121–126. [Google Scholar] [CrossRef]
- Politis, M.; Wu, K.; Loane, C.; Kiferle, L.; Molloy, S.; Brooks, D.J.; Piccini, P. Staging of serotonergic dysfunction in Parkinson’s Disease: An in vivo 11C-DASB PET study. Neurobiol. Dis. 2010, 40, 216–221. [Google Scholar] [CrossRef]
- Kim, J.H.; Son, Y.D.; Kim, J.H.; Choi, E.J.; Lee, S.Y.; Lee, J.E.; Cho, Z.H.; Kim, Y.B. Serotonin transporter availability in thalamic subregions in schizophrenia: A study using 7.0-T MRI with [11C]DASB high-resolution PET. Psychiatry Res. Neuroimaging 2015, 231, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, B.H.; Kim, E.; Howes, O.D.; Cho, K.I.K.; Yoon, Y.B.; Kwon, J.S. Higher serotonin transporter availability in early-onset obsessive–compulsive disorder patients undergoing escitalopram treatment: A [11C]DASB PET study. Hum. Psychopharmacol. 2018, 33, 2–7. [Google Scholar] [CrossRef]
- Christensen, J.B.; Bechgaard, K.; Paquignon, G. Carbon-11 labelling of MADAM in two different positions: A highly selective PET radioligand for the serotonin transporter. J. Label. Compd. Radiopharm. 2001, 44, 1013–1023. [Google Scholar] [CrossRef]
- Halldin, C.; Lundberg, J.; Sóvágó, J.; Gulyás, B.; Guilloteau, D.; Vercouillie, J.; Emond, P.; Chalon, S.; Tarkiainen, J.; Hiltunen, J.; et al. [11C]MADAM, a new serotonin transporter radioligand characterized in the monkey brain by PET. Synapse 2005, 58, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.; Odano, I.; Olsson, H.; Halldin, C.; Farde, L. Quantification of 11C-MADAM binding to the serotonin transporter in the human brain. J. Nucl. Med. 2005, 46, 1505–1515. [Google Scholar] [PubMed]
- Lundberg, J.; Halldin, C.; Farde, L. Measurement of serotonin transporter binding with PET and [ 11C]MADAM: A test-retest reproducibility study. Synapse 2006, 60, 256–263. [Google Scholar] [CrossRef]
- Majuri, J.; Joutsa, J.; Johansson, J.; Voon, V.; Parkkola, R.; Alho, H.; Arponen, E.; Kaasinen, V. Serotonin transporter density in binge eating disorder and pathological gambling: A PET study with [11C]MADAM. Eur. Neuropsychopharmacol. 2017, 27, 1281–1288. [Google Scholar] [CrossRef] [Green Version]
- Oya, S.; Choi, S.R.; Hou, C.; Mu, M.; Kung, M.P.; Acton, P.D.; Siciliano, M.; Kung, H.F. 2-((2-((Dimethylamino)methyl)phenyl)thio)-5-iodophenylamine (ADAM): An improved serotonin transporter ligand. Nucl. Med. Biol. 2000, 27, 249–254. [Google Scholar] [CrossRef]
- Lin, K.J.; Ye, X.X.; Yen, T.C.; Wey, S.P.; Tzen, K.Y.; Ting, G.; Hwang, J.J. Biodistribution study of [123I] ADAM in mice: Correlation with whole body autoradiography. Nucl. Med. Biol. 2002, 29, 643–650. [Google Scholar] [CrossRef]
- Itti, E.; Klein, G.; Rosso, J.; Evangelista, E.; Monin, J.L.; Gueret, P.; Meignan, M.; Thirion, J.P. Assessment of myocardial reperfusion after myocardial infarction using automatic 3-dimensional quantification and template matching. J. Nucl. Med. 2004, 45, 1981–1988. [Google Scholar]
- Lin, K.J.; Yen, T.C.; Wey, S.P.; Hwang, J.J.; Ye, X.X.; Tzen, K.Y.; Fu, Y.K.; Chen, J.C. Characterization of the binding sites for 123I-ADAM and the relationship to the serotonin transporter in rat and mouse brains using quantitative autoradiography. J. Nucl. Med. 2004, 45, 673–681. [Google Scholar] [PubMed]
- Ye, X.X.; Hwang, J.J.; Hsieh, J.F.; Chen, J.C.; Chou, Y.T.; Tu, K.Y.; Wey, S.P.; Ting, G. In vivo quantification by SPECT of [ 123I] ADAM bound to serotonin transporters in the brains of rabbits. Nucl. Med. Biol. 2004, 31, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Kauppinen, T.A.; Bergström, K.A.; Heikman, P.; Hiltunen, J.; Ahonen, A.K. Biodistribution and radiation dosimetry of [123I]ADAM in healthy human subjects: Preliminary results. Eur. J. Nucl. Med. Mol. Imaging 2003, 30, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Newberg, A.B.; Plössl, K.; Mozley, P.D.; Stubbs, J.B.; Wintering, N.; Udeshi, M.; Alavi, A.; Kauppinen, T.; Kung, H.F. Biodistribution and imaging with 123I-ADAM: A serotonin transporter imaging agent. J. Nucl. Med. 2004, 45, 834–841. [Google Scholar]
- Catafau, A.M.; Pérez, V.; Penengo, M.M.; Bullich, S.; Danús, M.; Puigdemont, D.; Pascual, J.C.; Corripio, I.; Llop, J.; Perich, J.; et al. SPECT of serotonin transporters using123I-ADAM: Optimal imaging time after bolus injection and long-term test-retest in healthy volunteers. J. Nucl. Med. 2005, 46, 1301–1309. [Google Scholar] [PubMed]
- Frokjaer, V.G.; Pinborg, L.H.; Madsen, J.; De Nijs, R.; Svarer, C.; Wagner, A.; Knudsen, G.M. Evaluation of the serotonin transporter ligand 123I-ADAM for SPECT studies on humans. J. Nucl. Med. 2008, 49, 247–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, K.H.; Huang, W.S.; Huang, S.Y.; Cheng, C.Y.; Chen, C.Y.; Shen, L.H.; Liu, J.C.; Fu, Y.K. Imaging serotonin transporters using [123I]ADAM SPECT in a parkinsonian primate model. Appl. Radiat. Isot. 2008, 66, 1799–1803. [Google Scholar] [CrossRef]
- Van De Giessen, E.; Booij, J. The SPECT tracer [123I]ADAM binds selectively to serotonin transporters: A double-blind, placebo-controlled study in healthy young men. Eur. J. Nucl. Med. Mol. Imaging 2010, 37, 1507–1511. [Google Scholar] [CrossRef] [Green Version]
- Catafau, A.M.; Perez, V.; Plaza, P.; Pascual, J.C.; Bullich, S.; Suarez, M.; Penengo, M.M.; Corripio, I.; Puigdemont, D.; Danus, M.; et al. Serotonin transporter occupancy induced by paroxetine in patients with major depression disorder: A 123I-ADAM SPECT study. Psychopharmacology 2006, 189, 145–153. [Google Scholar] [CrossRef]
- Herold, N.; Uebelhack, K.; Franke, L.; Amthauer, H.; Luedemann, L.; Bruhn, H.; Felix, R.; Uebelhack, R.; Plotkin, M. Imaging of serotonin transporters and its blockade by citalopram in patients with major depression using a novel SPECT ligand [123I]-ADAM. J. Neural Transm. 2006, 113, 659–670. [Google Scholar] [CrossRef]
- Lundgren, J.D.; Newberg, A.B.; Allison, K.C.; Wintering, N.A.; Ploessl, K.; Stunkard, A.J. 123I-ADAM SPECT imaging of serotonin transporter binding in patients with Night Eating Syndrome: A preliminary report. Psychiatry Res. Neuroimaging 2008, 162, 214–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amsterdam, J.D.; Newberg, A.B.; Newman, C.F.; Shults, J.; Wintering, N.; Soeller, I. Change over time in brain serotonin transporter binding in major depression: Effects of therapy measured with [123I]-ADAM SPECT. J. Neuroimaging 2013, 23, 469–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.H.; Chang, C.S.; Yang, Y.K.; Shen, L.H.; Yao, W.J. Comparison of brain serotonin transporter using [I-123]-ADAM between obese and nonobese young adults without an eating disorder. PLoS ONE 2017, 12, e0170886. [Google Scholar] [CrossRef]
- Shiue, G.G.; Fang, P.; Shiue, C.Y. Synthesis of N,N-dimethyl-2-(2-amino-4-[18F]fluorophenylthio)benzylamine as a serotonin transporter imaging agent. Appl. Radiat. Isot. 2003, 58, 183–191. [Google Scholar] [CrossRef]
- Shiue, G.G.; Choi, S.R.; Fang, P.; Hou, C.; Acton, P.D.; Cardi, C.; Saffer, J.R.; Greenberg, J.H.; Karp, J.S.; Kung, H.F.; et al. N,N-dimethyl-2-(2-amino-4-18F-fluorophenylthio)-benzylamine (4-18F-ADAM): An improved PET radioligand for serotonin transporters. J. Nucl. Med. 2003, 44, 1890–1897. [Google Scholar] [PubMed]
- Chen, Y.A.; Huang, W.S.; Lin, Y.S.; Cheng, C.Y.; Liu, R.S.; Wang, S.J.; Li, I.H.; Huang, S.Y.; Shiue, C.Y.; Chen, C.Y.; et al. Characterization of 4-[18F]-ADAM as an imaging agent for SERT in non-human primate brain using PET: A dynamic study. Nucl. Med. Biol. 2012, 39, 279–285. [Google Scholar] [CrossRef]
- Yeh, Y.W.; Ho, P.S.; Chen, C.Y.; Kuo, S.C.; Liang, C.S.; Ma, K.H.; Shiue, C.Y.; Huang, W.S.; Cheng, C.Y.; Wang, T.Y.; et al. Incongruent reduction of serotonin transporter associated with suicide attempts in patients with major depressive disorder: A positron emission tomography study with 4-[18F]-ADAM. Int. J. Neuropsychopharmacol. 2015, 18, pyu065. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.W.; Ho, P.S.; Kuo, S.C.; Chen, C.Y.; Liang, C.S.; Yen, C.H.; Huang, C.C.; Ma, K.H.; Shiue, C.Y.; Huang, W.S.; et al. Disproportionate reduction of serotonin transporter may predict the response and adherence to antidepressants in patients with major depressive disorder: A positron emission tomography study with 4-[18F]-ADAM. Int. J. Neuropsychopharmacol. 2015, 18, pyu120. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.L.; Parhi, A.K.; Oya, S.; Lieberman, B.; Kung, H.F. In vivo characterization of a series of 18F-diaryl sulfides (18F-2-(2′-((dimethylamino)methyl)-4′-(fluoroalkoxy) phenylthio)benzenamine) for PET imaging of the serotonin transporter. J. Nucl. Med. 2009, 50, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Zhang, Y.; Wu, Z.; Zhu, L.; Choi, S.R.; Ploessl, K.; Kung, H.F. One-step preparation of [18F]FPBM for PET imaging of serotonin transporter (SERT) in the brain. Nucl. Med. Biol. 2016, 43, 470–477. [Google Scholar] [CrossRef]
- Innis, R.; Baldwin, R.; Sybirska, E.; Zea, Y.; Laruelle, M.; Al-Tikriti, M.; Charney, D.; Zoghbi, S.; Smith, E.; Wisniewski, G.; et al. Single photon emission computed tomography imaging of monoamine reuptake sites in primate brain with [123I]CIT. Eur. J. Pharmacol. 1991, 200, 369–370. [Google Scholar] [CrossRef]
- Neumeyer, J.L.; Wang, S.; Milius, R.A.; Baldwin, R.M.; Zea-Ponce, Y.; Hoffer, P.B.; Sybirska, E.; Al-Tikriti, M.; Charney, D.S. [123I]-2.beta.-carbomethoxy-3.beta.-(4-iodophenyl)tropane: High-affinity SPECT (single photon emission computed tomography) radiotracer of monoamine reuptake sites in brain. J. Med. Chem. 1991, 34, 3144–3146. [Google Scholar] [CrossRef] [PubMed]
- Kuikka, J.T.; Åkerman, K.; Bergström, K.A.; Karhu, J.; Hiltunen, J.; Haukka, J.; Heikkinen, J.; Tiihonen, J.; Wang, S.; Neumeyer, J.L. Iodine-123 labelledN-(2-fluoroethyl)-2β-carbomethoxy-3β-(4-iodophenyl)nortropane for dopamine transporter imaging in the living human brain. Eur. J. Nucl. Med. 1995, 22, 682–686. [Google Scholar] [CrossRef] [PubMed]
- Hiltunen, J.; Åkerman, K.K.; Kuikka, J.T.; Bergström, K.A.; Halldin, C.; Nikula, T.; Räsänen, P.; Tiihonen, J.; Vauhkonen, M.; Karhu, J.; et al. Iodine-123 labeled nor-β-CIT as a potential tracer for serotonin transporter imaging in the human brain with single-photon emission tomography. Eur. J. Nucl. Med. 1998, 25, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Zitterl, W.; Aigner, M.; Stompe, T.; Zitterl-Eglseer, K.; Gutierrez-Lobos, K.; Wenzel, T.; Zettinig, G.; Hornik, K.; Pirker, W.; Thau, K. Changes in thalamus-hypothalamus serotonin transporter availability during clomipramine administration in patients with obsessive-compulsive disorder. Neuropsychopharmacology 2008, 33, 3126–3134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makkonen, I.; Riikonen, R.; Kokki, H.; Airaksinen, M.M.; Kuikka, J.T. Serotonin and dopamine transporter binding in children with autism determined by SPECT. Dev. Med. Child Neurol. 2008, 50, 593–597. [Google Scholar] [CrossRef]
- Lehto, S.M.; Tolmunen, T.; Kuikka, J.; Valkonen-Korhonen, M.; Joensuu, M.; Saarinen, P.I.; Vanninen, R.; Ahola, P.; Tiihonen, J.; Lehtonen, J. Midbrain serotonin and striatum dopamine transporter binding in double depression: A one-year follow-up study. Neurosci. Lett. 2008, 441, 291–295. [Google Scholar] [CrossRef]
- Dam, J.H.; Bender, D.; Peters, D.; Någren, K. [11C]NS9531, [11C]NS9762 and [11C]NS6417, specific SERT tracers: Pre-clinical evaluation in pigs and optimization of synthesis conditions using [11C]methyl triflate. Nucl. Med. Biol. 2016, 43, 42–51. [Google Scholar] [CrossRef]
Figure 1.
WAY-100365 derivatives.

Figure 2.
CUMI-101 derivatives.

Figure 3.
F15999 derivatives.

Figure 4.
AZ10419369 derivatives.

Figure 5.
5-HT2CR selective radioligands.

Figure 6.
Ketanserin analogues as 5-HT2AR radiotracers.

Figure 7.
Volinanserin analogues as 5-HT2AR radiotracers.

Figure 8.
CIMBI-5 analogues as 5-HT2AR radiotracers.

Figure 9.
Miscellaneous 5-HT2AR SPECT imaging tracers.

Figure 10.
[11C]MDL72222 and [11C]KF17643.

Figure 11.
[11C]MDL72222 and [11C]KF17643.

Figure 12.
Zacopride derivatives as 5-HT3R radiotracers.

Figure 13.
Miscellaneous derivatives as 5-HT3R radiotracers.

Figure 14.
SB207710 derivatives as 5-HT4R radioligands.

Figure 15.
[125I]SB258585 and [18F]12ST05.

Figure 16.
[11C]GSK215083 and [18F]2FNQ1P.

Figure 17.
DR4004 and [11C]DR4446.

Figure 18.
[11C]CIMBI-717, [11C]CIMBI-712, and [18F]ENL10.

Figure 19.
Radiolabeled SB-269970 derivatives.

Figure 20.
[18F]ENL30 and [11C]CIMBI-701.

Figure 21.
(−)-[11C]McN5655, (+)-[11C]McN5652, and [18F]FMe-(+)-McN5652.

Figure 22.
[123I]IDAM derivatives.

Figure 23.
SERT tracer in the tropane series.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mangeant, R.; Dubost, E.; Cailly, T.; Collot, V. Radiotracers for the Central Serotoninergic System. Pharmaceuticals 2022, 15, 571. https://doi.org/10.3390/ph15050571
AMA Style
Mangeant R, Dubost E, Cailly T, Collot V. Radiotracers for the Central Serotoninergic System. Pharmaceuticals. 2022; 15(5):571. https://doi.org/10.3390/ph15050571
Chicago/Turabian StyleMangeant, Reynald, Emmanuelle Dubost, Thomas Cailly, and Valérie Collot. 2022. "Radiotracers for the Central Serotoninergic System" Pharmaceuticals 15, no. 5: 571. https://doi.org/10.3390/ph15050571
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.