
Autophagy-Related ncRNAs in Pancreatic Cancer

 , and
, and
Abstract
:1. Introduction
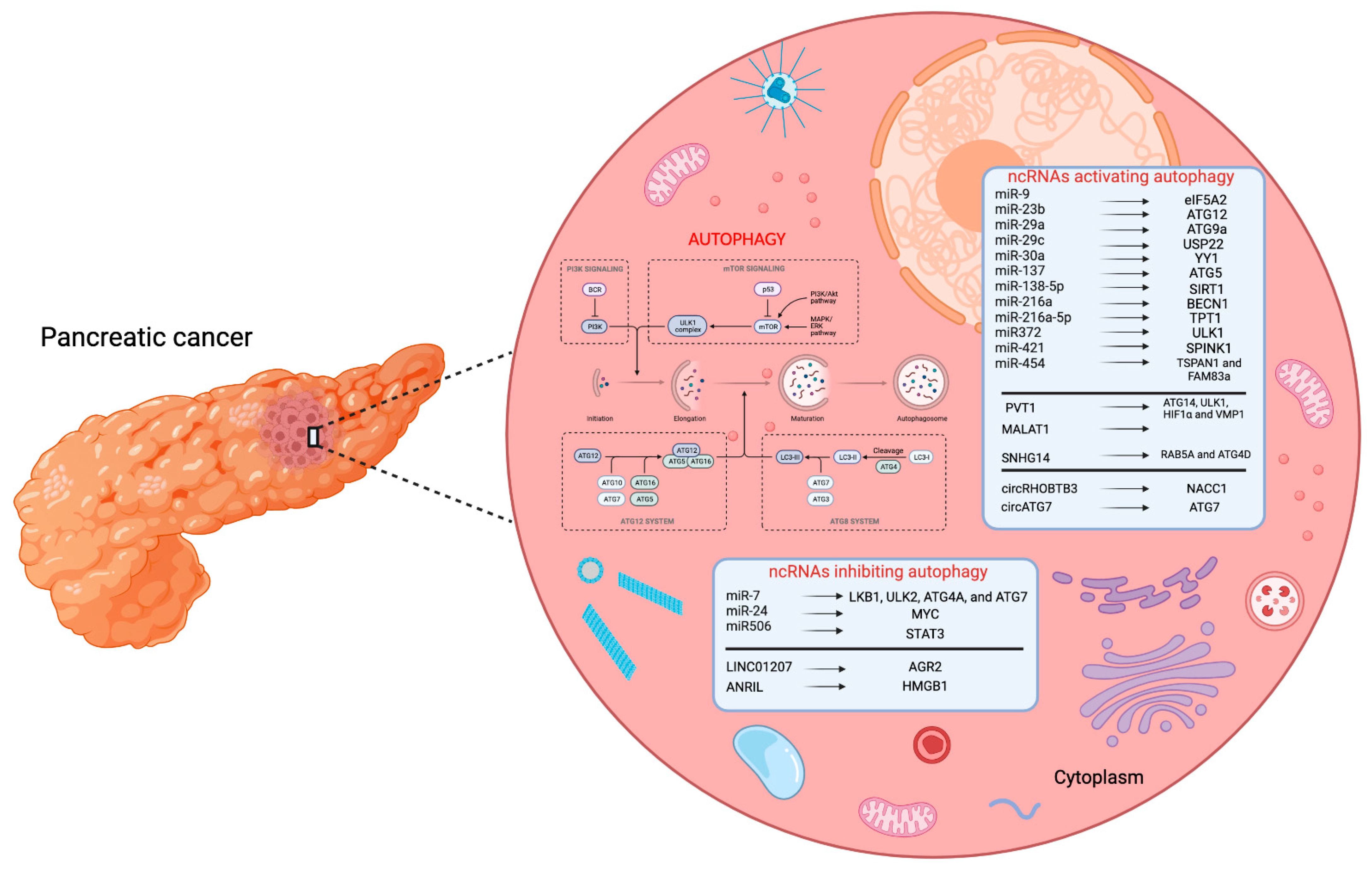
2. Autophagy-Related miRNAs in PC
3. Autophagy-Related lncRNAs in PC
4. Autophagy-Related circRNAs in PC
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Röder, P.V.; Wu, B.; Liu, Y.; Han, W. Pancreatic Regulation of Glucose Homeostasis. Exp. Mol. Med. 2016, 48, e219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, F.C.; Wright, C. Pancreas Organogenesis: From Bud to Plexus to Gland. Dev. Dyn. 2011, 240, 530–565. [Google Scholar] [CrossRef]
- Gittes, G.K. Developmental Biology of the Pancreas: A Comprehensive Review. Dev. Biol. 2009, 326, 4–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, T.J.; Hua, K.; Singh, A. Molecular Pathogenesis of Pancreatic Cancer. Prog. Mol. Biol. Transl. Sci. 2016, 144, 241–275. [Google Scholar] [CrossRef] [PubMed]
- Mahadevan, V. Anatomy of the Pancreas and Spleen. Surg. Oxf. 2019, 37, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Polonsky, K.S. The Past 200 Years in Diabetes. N. Engl. J. Med. 2012, 367, 1332–1340. [Google Scholar] [CrossRef]
- Braganza, J.M.; Lee, S.H.; McCloy, R.F.; McMahon, M.J. Chronic Pancreatitis. Lancet 2011, 377, 1184–1197. [Google Scholar] [CrossRef]
- Hezel, A.F.; Kimmelman, A.C.; Stanger, B.Z.; Bardeesy, N.; Depinho, R.A. Genetics and Biology of Pancreatic Ductal Adenocarcinoma. Genes Dev. 2006, 20, 1218–1249. [Google Scholar] [CrossRef] [Green Version]
- Klimstra, D.S. Nonductal Neoplasms of the Pancreas. Mod. Pathol. Off. J. U.S. Can. Acad. Pathol. Inc. 2007, 20 (Suppl. 1), S94–S112. [Google Scholar] [CrossRef] [Green Version]
- Luchini, C.; Grillo, F.; Fassan, M.; Vanoli, A.; Capelli, P.; Paolino, G.; Ingravallo, G.; Renzulli, G.; Doglioni, C.; D’Amuri, A.; et al. Malignant Epithelial/Exocrine Tumors of the Pancreas. Pathologica 2020, 112, 210–226. [Google Scholar] [CrossRef]
- Sridharan, V.; Hernandez-Barco, Y.G.; Ting, D.T. Landscape of Circulating Diagnostic Biomarkers in Pancreatic Malignancies. Ann. Pancreat. Cancer 2020, 3, 2616–2741. [Google Scholar] [CrossRef]
- Kanji, Z.S.; Gallinger, S. Diagnosis and Management of Pancreatic Cancer. CMAJ Can. Med. Assoc. J. J. Assoc. Medicale Can. 2013, 185, 1219–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costache, M.I.; Costache, C.A.; Dumitrescu, C.I.; Tica, A.A.; Popescu, M.; Baluta, E.A.; Anghel, A.C.; Saftoiu, A.; Dumitrescu, D. Which Is the Best Imaging Method in Pancreatic Adenocarcinoma Diagnosis and Staging—CT, MRI or EUS? Curr. Health Sci. J. 2017, 43, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Pekarek, L.; Fraile-Martinez, O.; Garcia-Montero, C.; Saez, M.A.; Barquero-Pozanco, I.; Del Hierro-Marlasca, L.; de Castro Martinez, P.; Romero-Bazán, A.; Alvarez-Mon, M.A.; Monserrat, J.; et al. Clinical Applications of Classical and Novel Biological Markers of Pancreatic Cancer. Cancers 2022, 14, 1866. [Google Scholar] [CrossRef]
- Wörmann, S.M.; Algül, H. Risk Factors and Therapeutic Targets in Pancreatic Cancer. Front. Oncol. 2013, 3, 282. [Google Scholar] [CrossRef] [Green Version]
- Risch, H.A.; Lu, L.; Kidd, M.S.; Wang, J.; Zhang, W.; Ni, Q.; Gao, Y.-T.; Yu, H. Helicobacter Pylori Seropositivities and Risk of Pancreatic Carcinoma. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2014, 23, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.M.; Li, D.; El-Deeb, A.S.; Wolff, R.A.; Bondy, M.L.; Davila, M.; Abbruzzese, J.L. Association Between Hepatitis B Virus and Pancreatic Cancer. J. Clin. Oncol. 2008, 26, 4557–4562. [Google Scholar] [CrossRef]
- El-Serag, H.B.; Engels, E.A.; Landgren, O.; Chiao, E.; Henderson, L.; Amaratunge, H.C.; Giordano, T.P. Risk of Hepatobiliary and Pancreatic Cancers after Hepatitis C Virus Infection: A Population-Based Study of U.S. Veterans. Hepatology 2009, 49, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Artinyan, A.; Soriano, P.A.; Prendergast, C.; Low, T.; Ellenhorn, J.D.I.; Kim, J. The Anatomic Location of Pancreatic Cancer Is a Prognostic Factor for Survival. HPB 2008, 10, 371–376. [Google Scholar] [CrossRef] [Green Version]
- Baur, A.D.J.; Pavel, M.; Prasad, V.; Denecke, T. Diagnostic Imaging of Pancreatic Neuroendocrine Neoplasms (PNEN): Tumor Detection, Staging, Prognosis, and Response to Treatment. Acta Radiol. 2016, 57, 260–270. [Google Scholar] [CrossRef]
- Viúdez, A.; De Jesus-Acosta, A.; Carvalho, F.L.; Vera, R.; Martín-Algarra, S.; Ramírez, N. Pancreatic Neuroendocrine Tumors: Challenges in an Underestimated Disease. Crit. Rev. Oncol. Hematol. 2016, 101, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Stathis, A.; Moore, M.J. Advanced Pancreatic Carcinoma: Current Treatment and Future Challenges. Nat. Rev. Clin. Oncol. 2010, 7, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Seufferlein, T.; Bachet, J.B.; Van Cutsem, E.; Rougier, P.; ESMO Guidelines Working Group. Pancreatic Adenocarcinoma: ESMO-ESDO Clinical Practice Guidelines for Diagnosis, Treatment and Follow-Up. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2012, 23 (Suppl. 7), vii33–vii40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zeng, L.; Chen, Y.; Lian, G.; Qian, C.; Chen, S.; Li, J.; Huang, K. Pancreatic Cancer Epidemiology, Detection, and Management. Gastroenterol. Res. Pract. 2016, 2016, 8962321. [Google Scholar] [CrossRef] [Green Version]
- D’Cruz, J.R.; Misra, S.; Shamsudeen, S. Pancreaticoduodenectomy. In StatPearls; StatPearls Publishing: Tampa, FL, USA, 2022. [Google Scholar]
- Edwards, P.; Kang, B.W.; Chau, I. Targeting the Stroma in the Management of Pancreatic Cancer. Front. Oncol. 2021, 11, 691185. [Google Scholar] [CrossRef]
- Zhu, H.; Li, T.; Du, Y.; Li, M. Pancreatic Cancer: Challenges and Opportunities. BMC Med. 2018, 16, 214. [Google Scholar] [CrossRef] [Green Version]
- Kelly, P.N. The Cancer Immunotherapy Revolution. Science 2018, 359, 1344–1345. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.H.; Jung, Y.-J.; Moon, S.-H. Immunotherapy for Pancreatic Cancer. World J. Clin. Cases 2021, 9, 2969–2982. [Google Scholar] [CrossRef]
- Adamska, A.; Domenichini, A.; Falasca, M. Pancreatic Ductal Adenocarcinoma: Current and Evolving Therapies. Int. J. Mol. Sci. 2017, 18, 1338. [Google Scholar] [CrossRef]
- Sohal, D.P.S.; Kennedy, E.B.; Cinar, P.; Conroy, T.; Copur, M.S.; Crane, C.H.; Garrido-Laguna, I.; Lau, M.W.; Johnson, T.; Krishnamurthi, S.; et al. Metastatic Pancreatic Cancer: ASCO Guideline Update. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2020, 38, JCO2001364. [Google Scholar] [CrossRef]
- Humphris, J.L.; Patch, A.-M.; Nones, K.; Bailey, P.J.; Johns, A.L.; McKay, S.; Chang, D.K.; Miller, D.K.; Pajic, M.; Kassahn, K.S.; et al. Hypermutation In Pancreatic Cancer. Gastroenterology 2017, 152, 68–74.e2. [Google Scholar] [CrossRef] [Green Version]
- Nevala-Plagemann, C.; Hidalgo, M.; Garrido-Laguna, I. From State-of-the-Art Treatments to Novel Therapies for Advanced-Stage Pancreatic Cancer. Nat. Rev. Clin. Oncol. 2020, 17, 108–123. [Google Scholar] [CrossRef] [PubMed]
- Mucciolo, G.; Roux, C.; Scagliotti, A.; Brugiapaglia, S.; Novelli, F.; Cappello, P. The Dark Side of Immunotherapy: Pancreatic Cancer. Cancer Drug Resist. 2020, 3, 491–520. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Kuma, A.; Kobayashi, Y.; Yamamoto, A.; Matsubae, M.; Takao, T.; Natsume, T.; Ohsumi, Y.; Yoshimori, T. Mouse Apg16L, a Novel WD-Repeat Protein, Targets to the Autophagic Isolation Membrane with the Apg12-Apg5 Conjugate. J. Cell Sci. 2003, 116, 1679–1688. [Google Scholar] [CrossRef] [Green Version]
- Perrotta, C.; Cattaneo, M.G.; Molteni, R.; De Palma, C. Autophagy in the Regulation of Tissue Differentiation and Homeostasis. Front. Cell Dev. Biol. 2020, 8, 602901. [Google Scholar] [CrossRef]
- Mathiassen, S.G.; De Zio, D.; Cecconi, F. Autophagy and the Cell Cycle: A Complex Landscape. Front. Oncol. 2017, 7, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klionsky, D.J. Autophagy: From Phenomenology to Molecular Understanding in Less than a Decade. Nat. Rev. Mol. Cell Biol. 2007, 8, 931–937. [Google Scholar] [CrossRef]
- Cao, W.; Li, J.; Yang, K.; Cao, D. An Overview of Autophagy: Mechanism, Regulation and Research Progress. Bull. Cancer 2021, 108, 304–322. [Google Scholar] [CrossRef]
- Yang, M.-C.; Wang, H.-C.; Hou, Y.-C.; Tung, H.-L.; Chiu, T.-J.; Shan, Y.-S. Blockade of Autophagy Reduces Pancreatic Cancer Stem Cell Activity and Potentiates the Tumoricidal Effect of Gemcitabine. Mol. Cancer 2015, 14, 179. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Wang, X.; Contino, G.; Liesa, M.; Sahin, E.; Ying, H.; Bause, A.; Li, Y.; Stommel, J.M.; Dell’antonio, G.; et al. Pancreatic Cancers Require Autophagy for Tumor Growth. Genes Dev. 2011, 25, 717–729. [Google Scholar] [CrossRef]
- Li, J.; Chen, X.; Kang, R.; Zeh, H.; Klionsky, D.J.; Tang, D. Regulation and Function of Autophagy in Pancreatic Cancer. Autophagy 2021, 17, 3275–3296. [Google Scholar] [CrossRef] [PubMed]
- Hwang, R.F.; Moore, T.; Arumugam, T.; Ramachandran, V.; Amos, K.D.; Rivera, A.; Ji, B.; Evans, D.B.; Logsdon, C.D. Cancer-Associated Stromal Fibroblasts Promote Pancreatic Tumor Progression. Cancer Res. 2008, 68, 918–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, S.; Nakata, K.; Ohuchida, K.; Takesue, S.; Nakayama, H.; Abe, T.; Koikawa, K.; Okumura, T.; Sada, M.; Horioka, K.; et al. Autophagy Is Required for Activation of Pancreatic Stellate Cells, Associated with Pancreatic Cancer Progression and Promotes Growth of Pancreatic Tumors in Mice. Gastroenterology 2017, 152, 1492–1506.e24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, A.; Kimmelman, A.C. Inhibition of Autophagy Attenuates Pancreatic Cancer Growth Independent of TP53/TRP53 Status. Autophagy 2014, 10, 1683–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, C.M.; Biancur, D.E.; Wang, X.; Halbrook, C.J.; Sherman, M.H.; Zhang, L.; Kremer, D.; Hwang, R.F.; Witkiewicz, A.K.; Ying, H.; et al. Pancreatic Stellate Cells Support Tumour Metabolism through Autophagic Alanine Secretion. Nature 2016, 536, 479–483. [Google Scholar] [CrossRef] [Green Version]
- Pipinikas, C.P.; Berner, A.M.; Sposito, T.; Thirlwell, C. The Evolving (Epi)Genetic Landscape of Pancreatic Neuroendocrine Tumours. Endocr. Relat. Cancer 2019, 26, R519–R544. [Google Scholar] [CrossRef] [Green Version]
- Marini, F.; Cianferotti, L.; Brandi, M.L. Epigenetic Mechanisms in Bone Biology and Osteoporosis: Can They Drive Therapeutic Choices? Int. J. Mol. Sci. 2016, 17, 1329. [Google Scholar] [CrossRef] [Green Version]
- Aurilia, C.; Donati, S.; Palmini, G.; Miglietta, F.; Iantomasi, T.; Brandi, M.L. The Involvement of Long Non-Coding RNAs in Bone. Int. J. Mol. Sci. 2021, 22, 3909. [Google Scholar] [CrossRef]
- Aurilia, C.; Donati, S.; Palmini, G.; Miglietta, F.; Falsetti, I.; Iantomasi, T.; Brandi, M.L. Are Non-Coding RNAs Useful Biomarkers in Parathyroid Tumorigenesis? Int. J. Mol. Sci. 2021, 22, 10465. [Google Scholar] [CrossRef]
- Marques, A.C.; Hughes, J.; Graham, B.; Kowalczyk, M.S.; Higgs, D.R.; Ponting, C.P. Chromatin Signatures at Transcriptional Start Sites Separate Two Equally Populated yet Distinct Classes of Intergenic Long Noncoding RNAs. Genome Biol. 2013, 14, R131. [Google Scholar] [CrossRef]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs Are the Predominant Transcript Isoform from Hundreds of Human Genes in Diverse Cell Types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.C.; Chang, H.Y. Molecular Mechanisms of Long Noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Q.; Guo, Q.; Xiao, Y.; Li, C.; Huang, Y.; Luo, X. Regulation of Bone Marrow Mesenchymal Stem Cell Fate by Long Non-Coding RNA. Bone 2020, 141, 115617. [Google Scholar] [CrossRef]
- Wilusz, J.E. A 360° View of Circular RNAs: From Biogenesis to Functions. Wiley Interdiscip. Rev. RNA 2018, 9, e1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of MiRNAs and SiRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynt, A.S.; Lai, E.C. Biological Principles of MicroRNA-Mediated Regulation: Shared Themes amid Diversity. Nat. Rev. Genet. 2008, 9, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. Elegans Heterochronic Gene Lin-4 Encodes Small RNAs with Antisense Complementarity to Lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional Regulation of the Heterochronic Gene Lin-14 by Lin-4 Mediates Temporal Pattern Formation in C. Elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Hsu, P.W.C.; Huang, H.-D.; Hsu, S.-D.; Lin, L.-Z.; Tsou, A.-P.; Tseng, C.-P.; Stadler, P.F.; Washietl, S.; Hofacker, I.L. MiRNAMap: Genomic Maps of MicroRNA Genes and Their Target Genes in Mammalian Genomes. Nucleic Acids Res. 2006, 34, D135–D139. [Google Scholar] [CrossRef] [Green Version]
- Malan-Müller, S.; Hemmings, S.M.J.; Seedat, S. Big Effects of Small RNAs: A Review of MicroRNAs in Anxiety. Mol. Neurobiol. 2013, 47, 726–739. [Google Scholar] [CrossRef]
- Huntzinger, E.; Izaurralde, E. Gene Silencing by MicroRNAs: Contributions of Translational Repression and MRNA Decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Ipsaro, J.J.; Joshua-Tor, L. From Guide to Target: Molecular Insights into Eukaryotic RNA-Interference Machinery. Nat. Struct. Mol. Biol. 2015, 22, 20–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donati, S.; Ciuffi, S.; Brandi, M.L. Human Circulating MiRNAs Real-Time QRT-PCR-Based Analysis: An Overview of Endogenous Reference Genes Used for Data Normalization. Int. J. Mol. Sci. 2019, 20, 4353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Li, X.; Gao, C.; Zhang, L. MicroRNA-Based Autophagy Inhibition as Targeted Therapy in Pancreatic Cancer. Biomed. Pharmacother. 2020, 132, 110799. [Google Scholar] [CrossRef]
- Mortoglou, M.; Tabin, Z.K.; Arisan, E.D.; Kocher, H.M.; Uysal-Onganer, P. Non-Coding RNAs in Pancreatic Ductal Adenocarcinoma: New Approaches for Better Diagnosis and Therapy. Transl. Oncol. 2021, 14, 101090. [Google Scholar] [CrossRef]
- Esmaeili, M.; Keshani, M.; Vakilian, M.; Esmaeili, M.; Peymani, M.; Seyed Forootan, F.; Chau, T.L.; Göktuna, S.I.; Zaker, S.R.; Nasr Esfahani, M.H.; et al. Role of Non-Coding RNAs as Novel Biomarkers for Detection of Colorectal Cancer Progression through Interaction with the Cell Signaling Pathways. Gene 2020, 753, 144796. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wu, H.; Liu, X.; Li, B.; Chen, Y.; Ren, X.; Liu, C.-G.; Yang, J.-M. Regulation of Autophagy by a Beclin 1-Targeted MicroRNA, MiR-30a, in Cancer Cells. Autophagy 2009, 5, 816–823. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Zhou, Y.; Zhi, X.; Ma, T.; Liu, H.; Chen, B.W.; Zheng, X.; Xie, S.; Zhao, B.; Feng, X.; et al. Delivery of MiR-212 by Chimeric Peptide-Condensed Supramolecular Nanoparticles Enhances the Sensitivity of Pancreatic Ductal Adenocarcinoma to Doxorubicin. Biomaterials 2019, 192, 590–600. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Z.; Lu, Y.; Song, K.; Liu, X.; Xia, F.; Sun, W. Downregulation of ULK1 by MicroRNA-372 Inhibits the Survival of Human Pancreatic Adenocarcinoma Cells. Cancer Sci. 2017, 108, 1811–1819. [Google Scholar] [CrossRef]
- Huang, L.; Hu, C.; Cao, H.; Wu, X.; Wang, R.; Lu, H.; Li, H.; Chen, H. MicroRNA-29c Increases the Chemosensitivity of Pancreatic Cancer Cells by Inhibiting USP22 Mediated Autophagy. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 47, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Pang, E.-J.; Yang, R.; Fu, X.; Liu, Y. Overexpression of Long Non-Coding RNA MALAT1 Is Correlated with Clinical Progression and Unfavorable Prognosis in Pancreatic Cancer. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2015, 36, 2403–2407. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, J.-O.; Zhou, W.-Y.; Chang, X.-Y.; Zhang, M.-M.; Zhang, Y.; Yang, X.-H. Long Non-Coding RNA LINC01207 Silencing Suppresses AGR2 Expression to Facilitate Autophagy and Apoptosis of Pancreatic Cancer Cells by Sponging MiR-143-5p. Mol. Cell. Endocrinol. 2019, 493, 110424. [Google Scholar] [CrossRef]
- He, Z.; Cai, K.; Zeng, Z.; Lei, S.; Cao, W.; Li, X. Autophagy-Associated CircRNA CircATG7 Facilitates Autophagy and Promotes Pancreatic Cancer Progression. Cell Death Dis. 2022, 13, 233. [Google Scholar] [CrossRef]
- Yang, T.; Shen, P.; Chen, Q.; Wu, P.; Yuan, H.; Ge, W.; Meng, L.; Huang, X.; Fu, Y.; Zhang, Y.; et al. FUS-Induced CircRHOBTB3 Facilitates Cell Proliferation via MiR-600/NACC1 Mediated Autophagy Response in Pancreatic Ductal Adenocarcinoma. J. Exp. Clin. Cancer Res. 2021, 40, 261. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.-M.; Jiang, M.-T.; Lin, P.; Yang, H.; Dang, Y.-W.; Yu, Q.; Liao, D.-Y.; Luo, D.-Z.; Chen, G. Potential CeRNA Networks Involved in Autophagy Suppression of Pancreatic Cancer Caused by Chloroquine Diphosphate: A Study Based on Differentially-Expressed CircRNAs, LncRNAs, MiRNAs and MRNAs. Int. J. Oncol. 2018, 54, 600–626. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gao, S.; Zhang, Y.; Yi, H.; Xu, M.; Xu, J.; Liu, H.; Ding, Z.; He, H.; Wang, H.; et al. MiR-216a-5p Inhibits Tumorigenesis in Pancreatic Cancer by Targeting TPT1/MTORC1 and Is Mediated by LINC01133. Int. J. Biol. Sci. 2020, 16, 2612–2627. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, J.; Zhang, L.; Zhu, Z.; Fan, J.; Chen, L.; Zhuang, L.; Luo, J.; Chen, H.; Liu, L.; et al. MicroRNA 23b Regulates Autophagy Associated with Radioresistance of Pancreatic Cancer Cells. Gastroenterology 2013, 145, 1133–1143.e12. [Google Scholar] [CrossRef]
- Zhang, X.; Shi, H.; Lin, S.; Ba, M.; Cui, S. MicroRNA-216a Enhances the Radiosensitivity of Pancreatic Cancer Cells by Inhibiting Beclin-1-Mediated Autophagy. Oncol. Rep. 2015, 34, 1557–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Zhang, J.-J.; Peng, Y.-P.; Zhu, Y.; Yin, L.-D.; Wei, J.-S.; Gao, W.-T.; Jiang, K.-R.; Miao, Y. A Yin-Yang 1/MiR-30a Regulatory Circuit Modulates Autophagy in Pancreatic Cancer Cells. J. Transl. Med. 2017, 15, 211. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Guo, X.; Yu, C.; Sun, C.; Jiang, J. MiR-138-5p Suppresses Autophagy in Pancreatic Cancer by Targeting SIRT1. Oncotarget 2016, 8, 11071–11082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.-C.; Huang, F.-Z.; Xu, H.-B.; Sun, J.-C.; Wang, C.-F. MicroRNA-137 Inhibits Autophagy and Chemosensitizes Pancreatic Cancer Cells by Targeting ATG5. Int. J. Biochem. Cell Biol. 2019, 111, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Shopit, A.; Li, X.; Tang, Z.; Awsh, M.; Shobet, L.; Niu, M.; Wang, H.; Mousa, H.; Alshwmi, M.; Tesfaldet, T.; et al. MiR-421 up-Regulation by the Oleanolic Acid Derivative K73-03 Regulates Epigenetically SPINK1 Transcription in Pancreatic Cancer Cells Leading to Metabolic Changes and Enhanced Apoptosis. Pharmacol. Res. 2020, 161, 105130. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liang, Y.; Zhou, L.; Yan, Y.; Liu, N.; Zhang, R.; Huang, Y.; Wang, M.; Tang, Y.; Ali, D.W.; et al. TSPAN1 Promotes Autophagy Flux and Mediates Cooperation between WNT-CTNNB1 Signaling and Autophagy via the MIR454-FAM83A-TSPAN1 Axis in Pancreatic Cancer. Autophagy 2021, 17, 3175–3195. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Chen, M.; Zhou, J.; Mo, M.-S.; Zhu, L.-H.; Liu, Y.-P.; Gui, Q.-J.; Zhang, L.; Li, G.-Q. MiR-7 Inhibits the Invasion and Metastasis of Gastric Cancer Cells by Suppressing Epidermal Growth Factor Receptor Expression. Oncol. Rep. 2014, 31, 1715–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Xue, J.-L.; Shen, Q.; Chen, J.; Tian, L. MicroRNA-7 Inhibits Tumor Growth and Metastasis by Targeting the Phosphoinositide 3-Kinase/Akt Pathway in Hepatocellular Carcinoma. Hepatology 2012, 55, 1852–1862. [Google Scholar] [CrossRef]
- Gu, D.-N.; Huang, Q.; Tian, L. The Molecular Mechanisms and Therapeutic Potential of MicroRNA-7 in Cancer. Expert Opin. Ther. Targets 2015, 19, 415–426. [Google Scholar] [CrossRef]
- Zhang, N.; Li, X.; Wu, C.W.; Dong, Y.; Cai, M.; Mok, M.T.S.; Wang, H.; Chen, J.; Ng, S.S.M.; Chen, M.; et al. MicroRNA-7 Is a Novel Inhibitor of YY1 Contributing to Colorectal Tumorigenesis. Oncogene 2013, 32, 5078–5088. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.-D.; Lu, Y.-Y.; Guo, H.; Xie, H.-H.; He, L.-J.; Shen, G.-F.; Zhou, J.-F.; Li, T.; Hu, S.-J.; Zhou, L.; et al. MicroRNA-7/NF-ΚB Signaling Regulatory Feedback Circuit Regulates Gastric Carcinogenesis. J. Cell Biol. 2015, 210, 613–627. [Google Scholar] [CrossRef] [Green Version]
- Gu, D.-N.; Jiang, M.-J.; Mei, Z.; Dai, J.-J.; Dai, C.-Y.; Fang, C.; Huang, Q.; Tian, L. MicroRNA-7 Impairs Autophagy-Derived Pools of Glucose to Suppress Pancreatic Cancer Progression. Cancer Lett. 2017, 400, 69–78. [Google Scholar] [CrossRef]
- Ye, Z.; Zou, C.; Chen, H.; Jiang, M.; Mei, Z.; Gu, D. MicroRNA-7 as a Potential Biomarker for Prognosis in Pancreatic Cancer. Dis. Markers 2020, 2020, 2782101. [Google Scholar] [CrossRef]
- Kwon, J.J.; Willy, J.A.; Quirin, K.A.; Wek, R.C.; Korc, M.; Yin, X.-M.; Kota, J. Novel Role of MiR-29a in Pancreatic Cancer Autophagy and Its Therapeutic Potential. Oncotarget 2016, 7, 71635–71650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Tang, Y.; Xie, S.; Zheng, X.; Zhang, S.; Mao, J.; Wang, B.; Hou, Y.; Hu, L.; Chai, K.; et al. Chimeric Peptide Supramolecular Nanoparticles for Plectin-1 Targeted MiRNA-9 Delivery in Pancreatic Cancer. Theranostics 2020, 10, 1151–1165. [Google Scholar] [CrossRef]
- Sun, L.; Hu, L.; Cogdell, D.; Lu, L.; Gao, C.; Tian, W.; Zhang, Z.; Kang, Y.; Fleming, J.B.; Zhang, W. MIR506 Induces Autophagy-Related Cell Death in Pancreatic Cancer Cells by Targeting the STAT3 Pathway. Autophagy 2017, 13, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Borchardt, H.; Ewe, A.; Morawski, M.; Weirauch, U.; Aigner, A. MiR24-3p Activity after Delivery into Pancreatic Carcinoma Cell Lines Exerts Profound Tumor-Inhibitory Effects through Distinct Pathways of Apoptosis and Autophagy Induction. Cancer Lett. 2021, 503, 174–184. [Google Scholar] [CrossRef]
- Bermúdez, M.; Aguilar-Medina, M.; Lizárraga-Verdugo, E.; Avendaño-Félix, M.; Silva-Benítez, E.; López-Camarillo, C.; Ramos-Payán, R. LncRNAs as Regulators of Autophagy and Drug Resistance in Colorectal Cancer. Front. Oncol. 2019, 9, 1008. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Cui, L.; Zhong, W.; Cai, Y. Autophagy-Associated LncRNAs: Promising Targets for Neurological Disease Diagnosis and Therapy. Neural Plast. 2020, 2020, e8881687. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jin, F.; Li, Y. A Novel Autophagy-Related LncRNA Prognostic Risk Model for Breast Cancer. J. Cell. Mol. Med. 2021, 25, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, H.; Shen, Q.; Feng, L.; Jin, H. Long Non-Coding RNAs Involved in Autophagy Regulation. Cell Death Dis. 2017, 8, e3073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Yi, C.; Yi, Y.; Qin, W.; Yan, Y.; Dong, X.; Zhang, X.; Huang, Y.; Zhang, R.; Wei, J.; et al. LncRNA PVT1 Promotes Gemcitabine Resistance of Pancreatic Cancer via Activating Wnt/β-Catenin and Autophagy Pathway through Modulating the MiR-619-5p/Pygo2 and MiR-619-5p/ATG14 Axes. Mol. Cancer 2020, 19, 118. [Google Scholar] [CrossRef]
- Liu, Y.-F.; Luo, D.; Li, X.; Li, Z.-Q.; Yu, X.; Zhu, H.-W. PVT1 Knockdown Inhibits Autophagy and Improves Gemcitabine Sensitivity by Regulating the MiR-143/HIF-1α/VMP1 Axis in Pancreatic Cancer. Pancreas 2021, 50, 227–234. [Google Scholar] [CrossRef]
- Jiao, F.; Hu, H.; Yuan, C.; Wang, L.; Jiang, W.; Jin, Z.; Guo, Z.; Wang, L. Elevated Expression Level of Long Noncoding RNA MALAT-1 Facilitates Cell Growth, Migration and Invasion in Pancreatic Cancer. Oncol. Rep. 2014, 32, 2485–2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Chen, H.; Gao, Y.; Wang, Y.-W.; Zhang, G.-Q.; Pan, S.-H.; Ji, L.; Kong, R.; Wang, G.; Jia, Y.-H.; et al. Long Noncoding RNA MALAT1 Promotes Aggressive Pancreatic Cancer Proliferation and Metastasis via the Stimulation of Autophagy. Mol. Cancer Ther. 2016, 15, 2232–2243. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhao, P.; Wang, C.; Xin, B. SNHG14 Enhances Gemcitabine Resistance by Sponging MiR-101 to Stimulate Cell Autophagy in Pancreatic Cancer. Biochem. Biophys. Res. Commun. 2019, 510, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Bi, R.; Li, L.; Zhou, K.; Yin, H. LncRNA ANRIL Aggravates the Chemoresistance of Pancreatic Cancer Cells to Gemcitabine by Targeting Inhibition of MiR-181a and Targeting HMGB1-Induced Autophagy. Aging 2021, 13, 19272–19281. [Google Scholar] [CrossRef] [PubMed]
- Dahlhaus, M.; Schult, C.; Lange, S.; Freund, M.; Junghanss, C. MicroRNA 181a Influences the Expression of HMGB1 and CD4 in Acute Leukemias. Anticancer Res. 2013, 33, 445–452. [Google Scholar] [PubMed]
- Tian, J.; Fu, C.; Zeng, X.; Fan, X.; Wu, Y. An Independent Prognostic Model Based on Ten Autophagy-Related Long Noncoding RNAs in Pancreatic Cancer Patients. Genet. Res. 2022, 2022, 3895396. [Google Scholar] [CrossRef]
- Hsiao, K.-Y.; Sun, H.S.; Tsai, S.-J. Circular RNA—New Member of Noncoding RNA with Novel Functions. Exp. Biol. Med. 2017, 242, 1136–1141. [Google Scholar] [CrossRef] [Green Version]
- Rong, Z.; Xu, J.; Shi, S.; Tan, Z.; Meng, Q.; Hua, J.; Liu, J.; Zhang, B.; Wang, W.; Yu, X.; et al. Circular RNA in Pancreatic Cancer: A Novel Avenue for the Roles of Diagnosis and Treatment. Theranostics 2021, 11, 2755–2769. [Google Scholar] [CrossRef]
- Shen, P.; Yang, T.; Chen, Q.; Yuan, H.; Wu, P.; Cai, B.; Meng, L.; Huang, X.; Liu, J.; Zhang, Y.; et al. CircNEIL3 Regulatory Loop Promotes Pancreatic Ductal Adenocarcinoma Progression via MiRNA Sponging and A-to-I RNA-Editing. Mol. Cancer 2021, 20, 51. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer Incidence and Mortality Worldwide: Sources, Methods and Major Patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Midha, S.; Chawla, S.; Garg, P.K. Modifiable and Non-Modifiable Risk Factors for Pancreatic Cancer: A Review. Cancer Lett. 2016, 381, 269–277. [Google Scholar] [CrossRef]
- Panico, A.; Tumolo, M.R.; Leo, C.G.; Donno, A.D.; Grassi, T.; Bagordo, F.; Serio, F.; Idolo, A.; Masi, R.D.; Mincarone, P.; et al. The Influence of Lifestyle Factors on MiRNA Expression and Signal Pathways: A Review. Epigenomics 2021, 13, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Wang, J.; Wang, D.; Buas, M.F.; Ren, X.; Freudenheim, J.L.; Belinsky, S.A.; Liu, S.; Ambrosone, C.B.; Higgins, M.J. Differences in MicroRNA Expression in Breast Cancer between Women of African and European Ancestry. Carcinogenesis 2019, 40, 61–69. [Google Scholar] [CrossRef]
- Ke, J.; Peng, X.; Mei, S.; Tian, J.; Ying, P.; Yang, N.; Wang, X.; Zou, D.; Yang, Y.; Zhu, Y.; et al. Evaluation of Polymorphisms in MicroRNA-Binding Sites and Pancreatic Cancer Risk in Chinese Population. J. Cell. Mol. Med. 2020, 24, 2252–2259. [Google Scholar] [CrossRef]
- Hu, P.; Qiao, O.; Wang, J.; Li, J.; Jin, H.; Li, Z.; Jin, Y. Rs1859168 A > C Polymorphism Regulates HOTTIP Expression and Reduces Risk of Pancreatic Cancer in a Chinese Population. World J. Surg. Oncol. 2017, 15, 155. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, S.; Van Buren, G.; Fisher, W.E. Pancreatic Cancer: Advances in Treatment. World J. Gastroenterol. 2014, 20, 9354–9360. [Google Scholar] [CrossRef] [PubMed]
- Ansari, D.; Gustafsson, A.; Andersson, R. Update on the Management of Pancreatic Cancer: Surgery Is Not Enough. World J. Gastroenterol. 2015, 21, 3157–3165. [Google Scholar] [CrossRef]
- Sapio, L.; Ragone, A.; Spina, A.; Salzillo, A.; Naviglio, S. AdipoRon and Pancreatic Ductal Adenocarcinoma: A Future Perspective in Overcoming Chemotherapy-Induced Resistance? Cancer Drug Resist. 2022, 5, 625–636. [Google Scholar] [CrossRef]
- Manley, S.J.; Olou, A.A.; Jack, J.L.; Ruckert, M.T.; Walsh, R.M.; Eades, A.E.; Bye, B.A.; Ambrose, J.; Messaggio, F.; Anant, S.; et al. Synthetic Adiponectin-Receptor Agonist, AdipoRon, Induces Glycolytic Dependence in Pancreatic Cancer Cells. Cell Death Dis. 2022, 13, 114. [Google Scholar] [CrossRef]
- Ragone, A.; Salzillo, A.; Spina, A.; Naviglio, S.; Sapio, L. Integrating Gemcitabine-Based Therapy With AdipoRon Enhances Growth Inhibition in Human PDAC Cell Lines. Front. Pharmacol. 2022, 13, 837503. [Google Scholar] [CrossRef] [PubMed]
- White, E. The Role for Autophagy in Cancer. J. Clin. Investig. 2015, 125, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Poillet-Perez, L.; Despouy, G.; Delage-Mourroux, R.; Boyer-Guittaut, M. Interplay between ROS and Autophagy in Cancer Cells, from Tumor Initiation to Cancer Therapy. Redox Biol. 2015, 4, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackl, M.; Heilmeier, U.; Weilner, S.; Grillari, J. Circulating MicroRNAs as Novel Biomarkers for Bone Diseases—Complex Signatures for Multifactorial Diseases? Mol. Cell. Endocrinol. 2016, 432, 83–95. [Google Scholar] [CrossRef]
- Li, J.; Ju, J.; Ni, B.; Wang, H. The Emerging Role of MiR-506 in Cancer. Oncotarget 2016, 7, 62778–62788. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Shelar, B.; Krishna, S. Versatile Role of MiR-24/24-1*/24-2* Expression in Cancer and Other Human Diseases. Am. J. Transl. Res. 2022, 14, 20–54. [Google Scholar]
- Korać, P.; Antica, M.; Matulić, M. MiR-7 in Cancer Development. Biomedicines 2021, 9, 325. [Google Scholar] [CrossRef]
- Shen, S.; Wang, Y.; Zhang, Y.; Dong, Z.; Xing, J. Long Non-Coding RNA Small Nucleolar RNA Host Gene 14, a Promising Biomarker and Therapeutic Target in Malignancy. Front. Cell Dev. Biol. 2021, 9, 746714. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Ashrafi Hafez, A.; Taheri, M. Metastasis Associated Lung Adenocarcinoma Transcript 1: An Update on Expression Pattern and Functions in Carcinogenesis. Exp. Mol. Pathol. 2020, 112, 104330. [Google Scholar] [CrossRef] [PubMed]
- Bajan, S.; Hutvagner, G. RNA-Based Therapeutics: From Antisense Oligonucleotides to MiRNAs. Cells 2020, 9, 137. [Google Scholar] [CrossRef] [Green Version]
- Ling, H.; Fabbri, M.; Calin, G.A. MicroRNAs and Other Non-Coding RNAs as Targets for Anticancer Drug Development. Nat. Rev. Drug Discov. 2013, 12, 847–865. [Google Scholar] [CrossRef] [PubMed]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA Therapeutics—Challenges and Potential Solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef] [PubMed]
- Tietze, R.; Zaloga, J.; Unterweger, H.; Lyer, S.; Friedrich, R.P.; Janko, C.; Pöttler, M.; Dürr, S.; Alexiou, C. Magnetic Nanoparticle-Based Drug Delivery for Cancer Therapy. Biochem. Biophys. Res. Commun. 2015, 468, 463–470. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| microRNA | Expression Profile in PC | Impact on Autophagy | Autophagy-Related Targets | References |
|---|---|---|---|---|
| miR-154-3p andmiR-663a-5p | ↓ | N.D. | 11 and 46 | [77] |
| miR-216a-5p | ↓ | + | Translationally-controlled 1 (TPT1) | [78] |
| miR-23b | ↓ | + | Autophagy-related genes 12 (ATG12) | [79] |
| miR-216a | ↓ | + | Beclin 1 (BECN1) | [80] |
| miR-30a | ↓ | + | Yin-Yang 1 (YY1) | [81] |
| miR-138-5p | ↓ | + | Silent mating type information regulation 2 homolog 1 (SIRT1) | [82] |
| miR-29c | ↓ | + | Ubiquitin-specific peptidase 22 (USP22) | [72] |
| miR-137 | ↓ | + | Autophagy-related genes 5 (ATG5) | [83] |
| miR-421 | ↓ | + | Serine protease inhibitor Kazal-type 1 (SPINK1) | [84] |
| miR-454 | ↓ | + | Tetraspanin 1 (TSPAN1) and FAM83A | [85] |
| miR-372 | ↓ | + | Unc-51-like autophagy activating kinase 1 (ULK1) | [71] |
| miR-7 | N.D. | − | Liver kinase B1 (LKB1), ULK2, ATG4A, and ATG7 | [91] |
| miR-7 | ↓ | − | N.D. | [92] |
| miR-29a | ↓ | − | Autophagy-related genes 9A (ATG9A) | [93] |
| miR-9 | ↓ | + | Eukaryotic Translation Initiation Factor 5A2 (eIF5A2) | [94] |
| miR-506 | ↓ | − | Signal transducer and activator of transcription 3 (STAT3) | [76] |
| miR-24 | ↓ | − | MYC | [96] |
| lncRNA | Expression Profile in PC | Impact on Autophagy | Autophagy-Related Targets (miRNA) | References |
|---|---|---|---|---|
| PVT1 | ↑ | + | ATG14 (miR-619-5p) | [101] |
| PVT1 | ↑ | + | HIF-1α and VMP1 (miR-143) | [102] |
| LINC01207 | ↑ | − | AGR2 (miR-143-5p) | [74] |
| MALAT1 | ↑ | + | TIA-1 | [104] |
| SNHG14 | ↑ | + | RAB5A and ATG4D | [105] |
| ANRIL | ↑ | − | HMGB1 (miR-181a) | [106] |
| AC245041.2, AC036176.1, LINC01089, LINC02257, FLVCR1-DT, AC006504.7, AC125494.2, AC012306.2, ST20-AS1, and AC005696.1 | N.D. | N.D. | N.D. | [108] |
| circRNA | Expression Profile in PC | Impact on Autophagy | Autophagy-Related Targets (miRNA) | References |
|---|---|---|---|---|
| circRHOBTB3 | ↑ | + | NACC1 (miR-600) | [76] |
| circATG7 | ↑ | + | ATG7 (miR-766-5p) | [75] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donati, S.; Aurilia, C.; Palmini, G.; Falsetti, I.; Iantomasi, T.; Brandi, M.L. Autophagy-Related ncRNAs in Pancreatic Cancer. Pharmaceuticals 2022, 15, 1547. https://doi.org/10.3390/ph15121547
Donati S, Aurilia C, Palmini G, Falsetti I, Iantomasi T, Brandi ML. Autophagy-Related ncRNAs in Pancreatic Cancer. Pharmaceuticals. 2022; 15(12):1547. https://doi.org/10.3390/ph15121547
Chicago/Turabian StyleDonati, Simone, Cinzia Aurilia, Gaia Palmini, Irene Falsetti, Teresa Iantomasi, and Maria Luisa Brandi. 2022. "Autophagy-Related ncRNAs in Pancreatic Cancer" Pharmaceuticals 15, no. 12: 1547. https://doi.org/10.3390/ph15121547