
Monoamine Neurotransmitters Control Basic Emotions and Affect Major Depressive Disorders
 ,
,  , and
, and 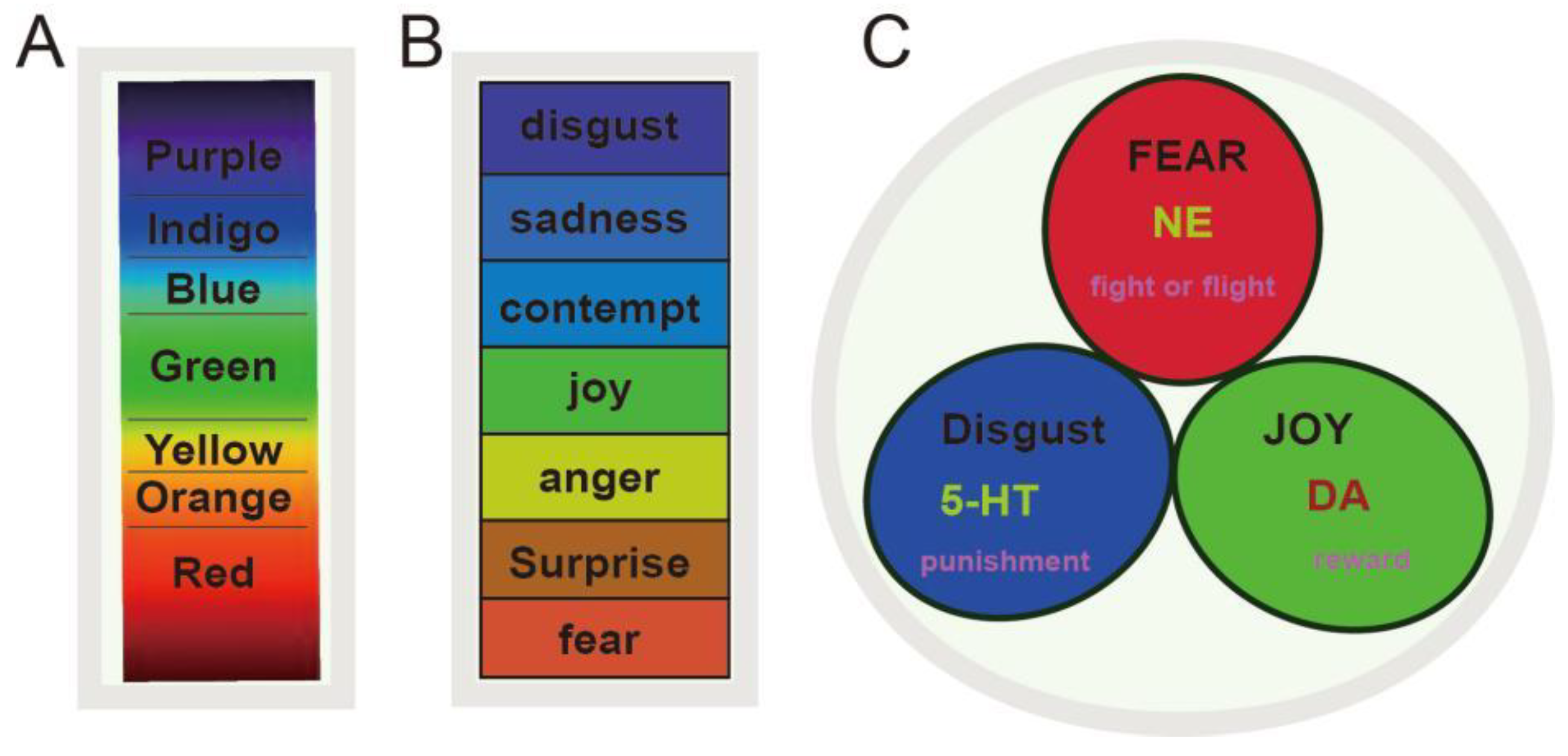{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Monoamine Neurotransmitters Mediate Three Core Effects
2.1. The Three Monoamines and Core Affects
2.2. The Significance of Basic Emotion Theory
3. Monoamine in the Digestive System
3.1. Monoamine and the Microorganisms in the Digestive System
3.2. Brain–Gut Axis
3.3. Brain–Liver Axis
4. Monoamines and Astrocyte in the Brain
4.1. Astrocytic Loss Might Be a Reason for MDD
4.2. Dysfunction of Buffering Ability in Astrocytes
4.3. Dysfunction of Glymphatic System
5. Monoamines and Microglial in the Brain
5.1. Microglia Cells
5.2. Early Life Stress Induces Microglia Vulnerability
5.3. Acute Stress
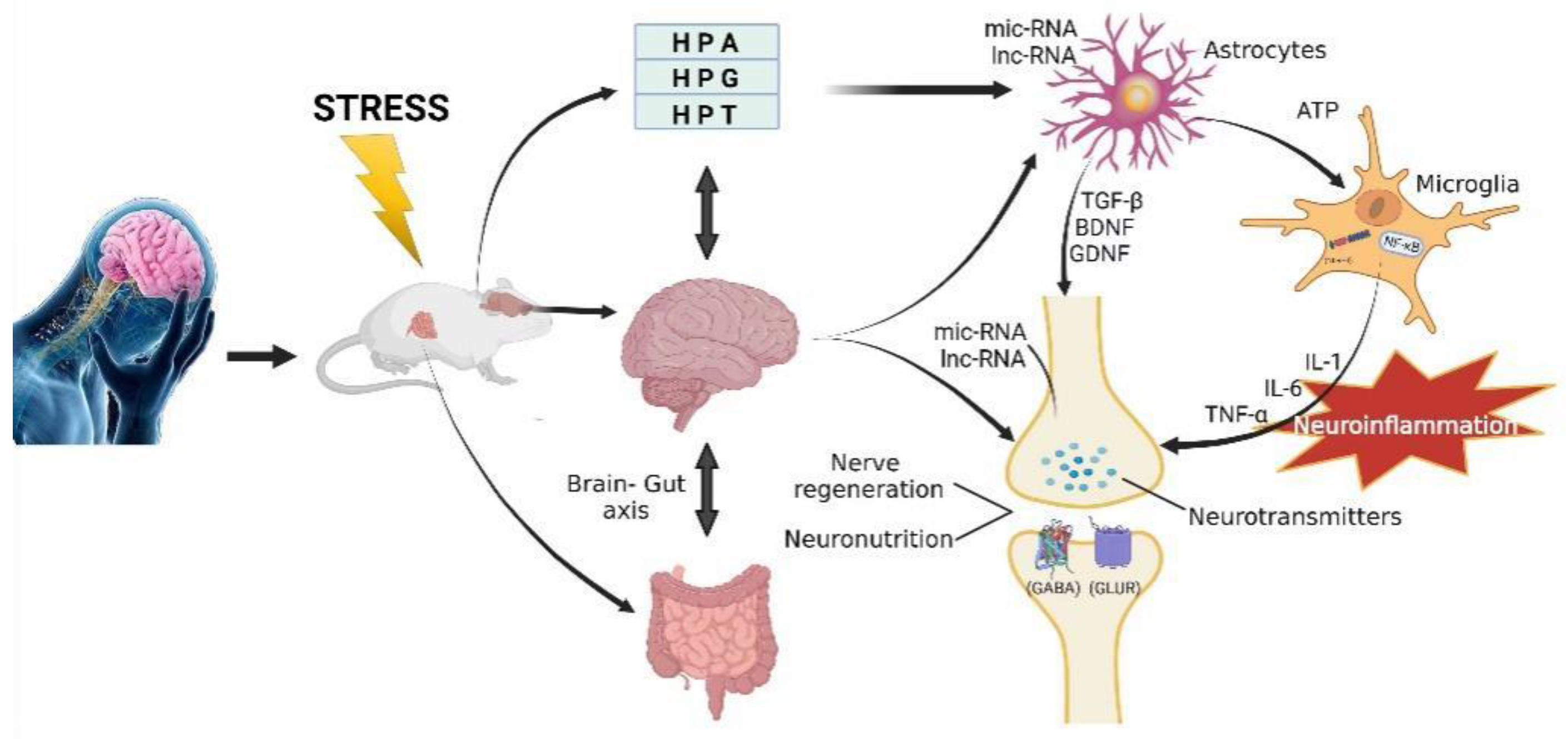
6. Monoamines and Neuroendocrine
6.1. Hypothalamic-Pituitary-Adrenal (HPA)
6.2. Hypothalamic-Pituitary-Gonadal Axis (HPG)
7. Neurotrophic, Nerve Regeneration, and Neuroplasticity
7.1. BDNF
7.2. Epigenetics
8. Monoamine Neurotransmitter and Other Neurotransmitters
9. Conclusions
10. Future Perspectives:
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Walker, E.R.; McGee, R.E.; Druss, B.G. Mortality in mental disorders and global disease burden implications: A systematic review and meta-analysis. JAMA Psychiatry 2015, 72, 334–341. [Google Scholar] [CrossRef]
- Pitsillou, E.; Bresnehan, S.M.; Kagarakis, E.A.; Wijoyo, S.J.; Liang, J.; Hung, A.; Karagiannis, T.C. The cellular and molecular basis of major depressive disorder: Towards a unified model for understanding clinical depression. Mol. Biol. Rep. 2020, 47, 753–770. [Google Scholar] [CrossRef]
- Disner, S.G.; Beevers, C.G.; Haigh, E.A.; Beck, A.T. Neural mechanisms of the cognitive model of depression. Nat. Rev. Neurosci. 2011, 12, 467–477. [Google Scholar] [CrossRef]
- Schildkraut, J.J. Biogenic amines and affective disorders. Annu. Rev. Med. 1974, 25, 333–348. [Google Scholar] [CrossRef]
- Schildkraut, J.J.; Schanberg, S.M.; Breese, G.R.; Kopin, I.J. Norepinephrine metabolism and drugs used in the affective disorders: A possible mechanism of action. Am. J. Psychiatry 1967, 124, 600–608. [Google Scholar] [CrossRef]
- Lövheim, H. A new three-dimensional model for emotions and monoamine neurotransmitters. Med. Hypotheses 2012, 78, 341–348. [Google Scholar] [CrossRef]
- Brown, J.; Li, B.; Yang, L. MAOI antidepressants: Could they be a next-generation ICB therapy? Front. Immunol. 2022, 13, 853624. [Google Scholar] [CrossRef]
- Brintzenhofe-Szoc, K.M.; Levin, T.T.; Li, Y.; Kissane, D.W.; Zabora, J.R. Mixed anxiety/depression symptoms in a large cancer cohort: Prevalence by cancer type. Psychosomatics 2009, 50, 383–391. [Google Scholar] [CrossRef]
- Gallego-Landin, I.; García-Baos, A.; Castro-Zavala, A.; Valverde, O. Reviewing the role of the endocannabinoid system in the pathophysiology of depression. Front. Pharmacol. 2021, 12, 762738. [Google Scholar] [CrossRef]
- Vasiliu, O. Investigational drugs for the treatment of depression (part 1): Monoaminergic, orexinergic, GABA-ergic, and anti-inflammatory agents. Front. Pharmacol. 2022, 13, 884143. [Google Scholar] [CrossRef]
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Bhatia, R.; Kumar, B.; Singh, G.; Monga, V. Design strategies, chemistry and therapeutic insights of multi-target directed ligands as antidepressant agents. Curr. Neuropharmacol. 2022, 20, 1329–1358. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Wang, F.; Patel, N.P.; Bourgeois, J.A.; Huang, J.H. A model for basic emotions using observations of behavior in drosophila. Front. Psychol. 2019, 10, 781. [Google Scholar] [CrossRef]
- He, Z.; Jiang, Y.; Gu, S.; Wu, D.; Qin, D.; Feng, G.; Ma, X.; Huang, J.H.; Wang, F. The aversion function of the limbic dopaminergic neurons and their roles in functional neurological disorders. Front. Cell Dev. Biol. 2021, 9, 713762. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Wang, F.; Cao, C.; Wu, E.; Tang, Y.Y.; Huang, J.H. An integrative way for studying neural basis of basic emotions with fMRI. Front. Neurosci. 2019, 13, 628. [Google Scholar] [CrossRef]
- Liang, F.; Feng, R.; Gu, S.; Jiang, S.; Zhang, X.; Li, N.; Xu, M.; Tang, Y.; Wang, F. Neurotransmitters and electrophysiological changes might work as biomarkers for diagnosing affective disorders. Dis. Markers 2021, 2021, 9116502. [Google Scholar] [CrossRef]
- Cavanagh, J.F.; Olguin, S.L.; Talledo, J.A.; Kotz, J.E.; Roberts, B.Z.; Nungaray, J.A.; Sprock, J.; Gregg, D.; Bhakta, S.G.; Light, G.A.; et al. Amphetamine alters an EEG marker of reward processing in humans and mice. Psychopharmacology 2022, 239, 923–933. [Google Scholar] [CrossRef]
- Kokkinou, M.; Ashok, A.H.; Howes, O.D. The effects of ketamine on dopaminergic function: Meta-analysis and review of the implications for neuropsychiatric disorders. Mol. Psychiatry 2018, 23, 59–69. [Google Scholar] [CrossRef]
- Irifune, M.; Shimizu, T.; Nomoto, M. Ketamine-induced hyperlocomotion associated with alteration of presynaptic components of dopamine neurons in the nucleus accumbens of mice. Pharmacol. Biochem. Behav. 1991, 40, 399–407. [Google Scholar] [CrossRef]
- Dunlop, B.W.; Nemeroff, C.B. The role of dopamine in the pathophysiology of depression. Arch. Gen. Psychiatry 2007, 64, 327–337. [Google Scholar] [CrossRef]
- Lumsden, E.W.; Troppoli, T.A.; Myers, S.J.; Zanos, P.; Aracava, Y.; Kehr, J.; Lovett, J.; Kim, S.; Wang, F.H.; Schmidt, S.; et al. Antidepressant-relevant concentrations of the ketamine metabolite (2R,6R)-hydroxynorketamine do not block NMDA receptor function. Proc. Natl. Acad. Sci. USA 2019, 116, 5160–5169. [Google Scholar] [CrossRef] [PubMed]
- Mitterauer, B.J. Ketamine may block NMDA receptors in astrocytes causing a rapid antidepressant effect. Front. Synaptic Neurosci. 2012, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Wojtas, A.; Bysiek, A.; Wawrzczak-Bargiela, A.; Szych, Z.; Majcher-Maślanka, I.; Herian, M.; Maćkowiak, M.; Gołembiowska, K. Effect of psilocybin and ketamine on brain neurotransmitters, glutamate receptors, DNA and rat behavior. Int. J. Mol. Sci. 2022, 23, 6713. [Google Scholar] [CrossRef]
- Yang, Y.; Cui, Y.; Sang, K.; Dong, Y.; Ni, Z.; Ma, S.; Hu, H. Ketamine blocks bursting in the lateral habenula to rapidly relieve depression. Nature 2018, 554, 317–322. [Google Scholar] [CrossRef]
- Hu, H. Reward and aversion. Annu. Rev. Neurosci. 2016, 39, 297–324. [Google Scholar] [CrossRef] [PubMed]
- Dienstbier, R.A. Arousal and physiological toughness: Implications for mental and physical health. Psychol. Rev. 1989, 96, 84–100. [Google Scholar] [CrossRef] [PubMed]
- Kopin, I.J. Avenues of investigation for the role of catecholamines in anxiety. Psychopathology 1984, 17 (Suppl. S1), 83–97. [Google Scholar] [CrossRef]
- Kohm, A.P.; Sanders, V.M. Norepinephrine and beta 2-adrenergic receptor stimulation regulate CD4+ T and B lymphocyte function in vitro and in vivo. Pharmacol. Rev. 2001, 53, 487–525. [Google Scholar]
- Bola, R.A.; Kiyatkin, E.A. Inflow of oxygen and glucose in brain tissue induced by intravenous norepinephrine: Relationships with central metabolic and peripheral vascular responses. J. Neurophysiol. 2018, 119, 499–508. [Google Scholar] [CrossRef]
- Uhel, F.; van der Poll, T. Norepinephrine in septic shock: A mixed blessing. Am. J. Respir. Crit. Care Med. 2020, 202, 788–789. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Wang, W.; Wang, F.; Huang, J.H. Neuromodulator and emotion biomarker for stress induced mental disorders. Neural Plast. 2016, 2016, 2609128. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Gu, S.; Lei, Y.; Lu, S.; Wang, W.; Li, Y.; Wang, F. Safety needs mediate stressful events induced mental disorders. Neural Plast. 2016, 2016, 8058093. [Google Scholar] [CrossRef]
- Kabitzke, P.; Silva, L.; Wiedenmayer, C.P. Norepinephrine mediates contextual fear learning and hippocampal pCREB in juvenile rats exposed to predator odor. Neurobiol. Learn. Mem. 2011, 96, 166–172. [Google Scholar] [CrossRef]
- Toussaint, L.; Sirois, F.; Hirsch, J.; Kohls, N.; Weber, A.; Schelling, J.; Vajda, C.; Offenbäecher, M. Anger rumination mediates differences between fibromyalgia patients and healthy controls on mental health and quality of life. Personal. Ment. Health 2019, 13, 119–133. [Google Scholar] [CrossRef]
- Wang, F.; Pan, F.; Tang, Y.Y.; Huang, J.H. Editorial: Uncertainty induced emotional disorders during the COVID-19. Front. Psychol. 2022, 13, 943966. [Google Scholar] [CrossRef]
- Gu, S.; He, Z.; Xu, Q.; Dong, J.; Xiao, T.; Liang, F.; Ma, X.; Wang, F.; Huang, J.H. The relationship between 5-hydroxytryptamine and its metabolite changes with post-stroke depression. Front. Psychiatry 2022, 13, 871754. [Google Scholar] [CrossRef]
- Artigas, F. Serotonin receptors involved in antidepressant effects. Pharmacol. Ther. 2013, 137, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Horton, R.E.; Apple, D.M.; Owens, W.A.; Baganz, N.L.; Cano, S.; Mitchell, N.C.; Vitela, M.; Gould, G.G.; Koek, W.; Daws, L.C. Decynium-22 enhances SSRI-induced antidepressant-like effects in mice: Uncovering novel targets to treat depression. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 10534–10543. [Google Scholar] [CrossRef]
- De Vry, J.; Schreiber, R.; Melon, C.; Dalmus, M.; Jentzsch, K.R. 5-HT1A receptors are differentially involved in the anxiolytic- and antidepressant-like effects of 8-OH-DPAT and fluoxetine in the rat. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2004, 14, 487–495. [Google Scholar] [CrossRef]
- Barnes, N.M.; Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 1999, 38, 1083–1152. [Google Scholar] [CrossRef]
- Shah, P.A.; Park, C.J.; Shaughnessy, M.P.; Cowles, R.A. Serotonin as a mitogen in the gastrointestinal tract: Revisiting a familiar molecule in a new role. Cell. Mol. Gastroenterol. Hepatol. 2021, 12, 1093–1104. [Google Scholar] [CrossRef]
- Jenkins, T.A.; Nguyen, J.C.; Polglaze, K.E.; Bertrand, P.P. Influence of tryptophan and serotonin on mood and cognition with a possible role of the gut-brain axis. Nutrients 2016, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Goda, M.; Kanda, M.; Yoshioka, T.; Yoshida, A.; Murai, Y.; Zamami, Y.; Aizawa, F.; Niimura, T.; Hamano, H.; Okada, N.; et al. Effects of 5-HT₃ receptor antagonists on cisplatin-induced kidney injury. Clin. Transl. Sci. 2021, 14, 1906–1916. [Google Scholar] [CrossRef] [PubMed]
- Lucas-Osma, A.M.; Li, Y.; Murray, K.; Lin, S.; Black, S.; Stephens, M.J.; Ahn, A.H.; Heckman, C.J.; Fenrich, K.K.; Fouad, K.; et al. 5-HT1D receptors inhibit the monosynaptic stretch reflex by modulating C-fiber activity. J. Neurophysiol. 2019, 121, 1591–1608. [Google Scholar] [CrossRef] [PubMed]
- Azizi, H.; Hwang, J.; Suen, V.; Kang, N.; Somvanshi, R.K.; Tadavarty, R.; Kumar, U.; Sastry, B.S. Sleep deprivation induces changes in 5-HT actions and 5-HT1A receptor expression in the rat hippocampus. Neurosci. Lett. 2017, 655, 151–155. [Google Scholar] [CrossRef]
- Gul, S.; Saleem, D.; Haleem, M.A.; Haleem, D.J. Inhibition of hormonal and behavioral effects of stress by tryptophan in rats. Nutr. Neurosci. 2019, 22, 409–417. [Google Scholar] [CrossRef]
- Gul, S.; Khan, A.; Raza, A.; Khan, I.; Ehtisham, S. Association of XPD Lys751Gln gene polymorphism with susceptibility and clinical outcome of colorectal cancer in Pakistani population: A case-control pharmacogenetic study. Genes Genom. 2020, 42, 1389–1398. [Google Scholar] [CrossRef]
- Pattij, T.; Schoffelmeer, A.N. Serotonin and inhibitory response control: Focusing on the role of 5-HT(1A) receptors. Eur. J. Pharmacol. 2015, 753, 140–145. [Google Scholar] [CrossRef]
- Ekman, P. An argument for basic emotions. Cogn. Emot. 1992, 6, 169–200. [Google Scholar] [CrossRef]
- Ortony, A.; Turner, T.J. What’s basic about basic emotions? Psychol. Rev. 1990, 97, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Ekman, P.; Cordaro, D. What is meant by calling emotions basic. Emot. Rev. 2011, 3, 364–370. [Google Scholar] [CrossRef]
- Levenson, R.W. Basic emotion questions. Emot. Rev. 2011, 3, 379–386. [Google Scholar] [CrossRef]
- Ekman, P. Emotions inside out. 130 years after Darwin’s “The expression of the emotions in man and animal”. Ann. N. Y. Acad. Sci. 2003, 1000, 1–6. [Google Scholar] [CrossRef]
- Gu, S.; Gao, M.; Yan, Y.; Wang, F.; Tang, Y.Y.; Huang, J.H. The neural mechanism underlying cognitive and emotional processes in creativity. Front. Psychol. 2018, 9, 1924. [Google Scholar] [CrossRef]
- MahmoudianDehkordi, S.; Bhattacharyya, S.; Brydges, C.R.; Jia, W.; Fiehn, O.; Rush, A.J.; Dunlop, B.W.; Kaddurah-Daouk, R. Gut microbiome-linked metabolites in the pathobiology of major depression with or without anxiety-a role for bile acids. Front. Neurosci. 2022, 16, 937906. [Google Scholar] [CrossRef]
- Mayer, E.A.; Tillisch, K.; Gupta, A. Gut/brain axis and the microbiota. J. Clin. Investig. 2015, 125, 926–938. [Google Scholar] [CrossRef]
- Foster, J.A.; McVey Neufeld, K.A. Gut-brain axis: How the microbiome influences anxiety and depression. Trends Neurosci. 2013, 36, 305–312. [Google Scholar] [CrossRef]
- Cheung, S.G.; Goldenthal, A.R.; Uhlemann, A.C.; Mann, J.J.; Miller, J.M.; Sublette, M.E. Systematic review of gut microbiota and major depression. Front. Psychiatry 2019, 10, 34. [Google Scholar] [CrossRef]
- Endle, H.; Horta, G.; Stutz, B.; Muthuraman, M.; Tegeder, I.; Schreiber, Y.; Snodgrass, I.F.; Gurke, R.; Liu, Z.W.; Sestan-Pesa, M.; et al. AgRP neurons control feeding behaviour at cortical synapses via peripherally derived lysophospholipids. Nat. Metab. 2022, 4, 683–692. [Google Scholar] [CrossRef]
- Goertsen, D.; Flytzanis, N.C.; Goeden, N.; Chuapoco, M.R.; Cummins, A.; Chen, Y.; Fan, Y.; Zhang, Q.; Sharma, J.; Duan, Y.; et al. AAV capsid variants with brain-wide transgene expression and decreased liver targeting after intravenous delivery in mouse and marmoset. Nat. Neurosci. 2022, 25, 106–115. [Google Scholar] [CrossRef]
- Xu, Q.; Jiang, M.; Gu, S.; Zhang, X.; Feng, G.; Ma, X.; Xu, S.; Wu, E.; Huang, J.H.; Wang, F. Metabolomics changes in brain-gut axis after unpredictable chronic mild stress. Psychopharmacology 2022, 239, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Dayan, P. Instrumental vigour in punishment and reward. Eur. J. Neurosci. 2012, 35, 1152–1168. [Google Scholar] [CrossRef] [PubMed]
- Sanada, K.; Nakajima, S.; Kurokawa, S.; Barceló-Soler, A.; Ikuse, D.; Hirata, A.; Yoshizawa, A.; Tomizawa, Y.; Salas-Valero, M.; Noda, Y.; et al. Gut microbiota and major depressive disorder: A systematic review and meta-analysis. J. Affect. Disord. 2020, 266, 1–13. [Google Scholar] [CrossRef]
- Estes, M.L.; McAllister, A.K. Maternal immune activation: Implications for neuropsychiatric disorders. Science 2016, 353, 772–777. [Google Scholar] [CrossRef]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef]
- Yang, J.; Zheng, P.; Li, Y.; Wu, J.; Tan, X.; Zhou, J.; Sun, Z.; Chen, X.; Zhang, G.; Zhang, H.; et al. Landscapes of bacterial and metabolic signatures and their interaction in major depressive disorders. Sci. Adv. 2020, 6, eaba8555. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, H.; Gui, S.; Zeng, B.; Pu, J.; Zheng, P.; Zeng, L.; Luo, Y.; Wu, Y.; Zhou, C.; et al. Proteomics analysis of the gut-brain axis in a gut microbiota-dysbiosis model of depression. Transl. Psychiatry 2021, 11, 568. [Google Scholar] [CrossRef]
- Zhang, L.; Qing, P.; Yang, H.; Wu, Y.; Liu, Y.; Luo, Y. Gut microbiome and metabolites in aystemic lupus erythematosus: Link, mechanisms and intervention. Front. Immunol. 2021, 12, 686501. [Google Scholar] [CrossRef]
- Schluter, J.; Peled, J.U.; Taylor, B.P.; Markey, K.A.; Smith, M.; Taur, Y.; Niehus, R.; Staffas, A.; Dai, A.; Fontana, E.; et al. The gut microbiota is associated with immune cell dynamics in humans. Nature 2020, 588, 303–307. [Google Scholar] [CrossRef]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P. Psychobiotics and the manipulation of bacteria-gut-brain signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef]
- Zhang, X.; Lei, Y.; Wang, F. The influence of oxytocin, progesterone and estrogen on disgust and its neurophysiological mechanism. Adv. Psychol. Sci. 2022, 30, 85–97. [Google Scholar] [CrossRef]
- Val-Laillet, D. Review: Impact of food, gut-brain signals and metabolic status on brain activity in the pig model: 10 years of nutrition research using in vivo brain imaging. Anim. Int. J. Anim. Biosci. 2019, 13, 2699–2713. [Google Scholar] [CrossRef]
- Li, Q.F.; Lu, W.T.; Zhang, Q.; Zhao, Y.D.; Wu, C.Y.; Zhou, H.F. Proprietary medicines containing bupleurum chinense DC. (Chaihu) for depression: Network meta-analysis and network pharmacology prediction. Front. Pharmacol. 2022, 13, 773537. [Google Scholar] [CrossRef]
- Hsu, J.H.; Chien, I.C.; Lin, C.H. Increased risk of chronic liver disease in patients with major depressive disorder: A population-based study. J. Affect. Disord. 2019, 251, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Shao, Q.; Wu, Y.; Ji, J.; Xu, T.; Yu, Q.; Ma, C.; Liao, X.; Cheng, F.; Wang, X. Interaction mechanisms between major depressive disorder and non-alcoholic fatty liver disease. Front. Psychiatry 2021, 12, 711835. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, A.A.; Kaplan, G.G.; Sharkey, K.A.; Lethebe, B.C.; Swain, M.G. Impact of major depression and antidepressant use on alcoholic and non-alcoholic fatty liver disease: A population-based study. Liver Int. Off. J. Int. Assoc. Study Liver 2021, 41, 2308–2317. [Google Scholar] [CrossRef]
- Paliokha, R.; Grinchii, D.; Khoury, T.; Nejati Bervanlou, R.; Dremencov, E. Inhibition of cytochrome P450 with proadifen alters the excitability of brain catecholamine-secreting neurons. Gen. Physiol. Biophys. 2022, 41, 255–262. [Google Scholar] [CrossRef]
- Lazarus, R.S. Progress on a cognitive-motivational-relational theory of emotion. Am. Psychol. 1991, 46, 819–834. [Google Scholar] [CrossRef]
- Nezlek, J.B.; Vansteelandt, K.; Van Mechelen, I.; Kuppens, P. Appraisal-emotion relationships in daily life. Emotion 2008, 8, 145–150. [Google Scholar] [CrossRef]
- Fullana, N.; Gasull-Camós, J.; Tarrés-Gatius, M.; Castañé, A.; Bortolozzi, A.; Artigas, F. Astrocyte control of glutamatergic activity: Downstream effects on serotonergic function and emotional behavior. Neuropharmacology 2020, 166, 107914. [Google Scholar] [CrossRef]
- Cao, X.; Li, L.P.; Wang, Q.; Wu, Q.; Hu, H.H.; Zhang, M.; Fang, Y.Y.; Zhang, J.; Li, S.J.; Xiong, W.C.; et al. Astrocyte-derived ATP modulates depressive-like behaviors. Nat. Med. 2013, 19, 773–777. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Jie, W.; Liu, J.H.; Yang, J.M.; Gao, T.M. An astroglial basis of major depressive disorder? An overview. Glia 2017, 65, 1227–1250. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Smith, N.A.; Xu, Q.; Goldman, S.; Peng, W.; Huang, J.H.; Takano, T.; Nedergaard, M. Photolysis of caged Ca2+ but not receptor-mediated Ca2+ signaling triggers astrocytic glutamate release. J. Neurosci. Off. J. Soc. Neurosci. 2013, 33, 17404–17412. [Google Scholar] [CrossRef] [PubMed]
- Rudzki, L.; Maes, M. From “leaky gut” to impaired glia-neuron communication in depression. Adv. Exp. Med. Biol. 2021, 1305, 129–155. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Smith, N.A.; Xu, Q.; Fujita, T.; Baba, A.; Matsuda, T.; Takano, T.; Bekar, L.; Nedergaard, M. Astrocytes modulate neural network activity by Ca²+-dependent uptake of extracellular K+. Sci. Signal. 2012, 5, ra26. [Google Scholar] [CrossRef]
- Wang, F.; Xu, Q.; Wang, W.; Takano, T.; Nedergaard, M. Bergmann glia modulate cerebellar Purkinje cell bistability via Ca2+-dependent K+ uptake. Proc. Natl. Acad. Sci. USA 2012, 109, 7911–7916. [Google Scholar] [CrossRef]
- Ding, F.; O’Donnell, J.; Xu, Q.; Kang, N.; Goldman, N.; Nedergaard, M. Changes in the composition of brain interstitial ions control the sleep-wake cycle. Science 2016, 352, 550–555. [Google Scholar] [CrossRef]
- Ding, F.; O’Donnell, J.; Thrane, A.S.; Zeppenfeld, D.; Kang, H.; Xie, L.; Wang, F.; Nedergaard, M. α1-Adrenergic receptors mediate coordinated Ca2+ signaling of cortical astrocytes in awake, behaving mice. Cell Calcium 2013, 54, 387–394. [Google Scholar] [CrossRef]
- Marathe, S.V.; D’almeida, P.L.; Virmani, G.; Bathini, P.; Alberi, L. Effects of monoamines and antidepressants on astrocyte physiology: Implications for monoamine hypothesis of depression. J. Exp. Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Rangroo, T.V.; Thrane, A.S.; Wang, F.; Cotrina, M.L.; Smith, N.A.; Chen, M.; Xu, Q.; Kang, N.; Fujita, T.; Nagelhus, E.A.; et al. Ammonia triggers neuronal disinhibition and seizures by impairing astrocyte potassium buffering. Nat. Med. 2013, 19, 1643–1648. [Google Scholar] [CrossRef]
- Mestre, H.; Mori, Y.; Nedergaard, M. The brain’s glymphatic system: Current controversies. Trends Neurosci. 2020, 43, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yang, J.; Pan, F.; Ho, R.C.; Huang, J.H. Editorial: Neurotransmitters and emotions. Front. Psychol. 2020, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Iliff, J.J.; Lee, H.; Yu, M.; Feng, T.; Logan, J.; Nedergaard, M.; Benveniste, H. Brain-wide pathway for waste clearance captured by contrast-enhanced MRI. J. Clin. Investig. 2013, 123, 1299–1309. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Yang, L.; Sun, G.; Qi, S.; Li, B. Mechanism of depression as a risk factor in the development of Alzheimer’s disease: The function of AQP4 and the glymphatic system. Psychopharmacology 2017, 234, 365–379. [Google Scholar] [CrossRef]
- Liu, X.; Hao, J.; Yao, E.; Cao, J.; Zheng, X.; Yao, D.; Zhang, C.; Li, J.; Pan, D.; Luo, X.; et al. Polyunsaturated fatty acid supplement alleviates depression-incident cognitive dysfunction by protecting the cerebrovascular and glymphatic systems. Brain Behav. Immun. 2020, 89, 357–370. [Google Scholar] [CrossRef]
- Liu, Y.; Feng, S.; Subedi, K.; Wang, H. Attenuation of ischemic stroke-caused brain injury by a monoamine oxidase inhibitor involves improved proteostasis and reduced neuroinflammation. Mol. Neurobiol. 2020, 57, 937–948. [Google Scholar] [CrossRef]
- Yan, T.; Qiu, Y.; Yu, X.; Yang, L. Glymphatic dysfunction: A bridge between sleep disturbance and mood disorders. Front. Psychiatry 2021, 12, 658340. [Google Scholar] [CrossRef]
- Christensen, J.; Yamakawa, G.R.; Shultz, S.R.; Mychasiuk, R. Is the glymphatic system the missing link between sleep impairments and neurological disorders? Examining the implications and uncertainties. Prog. Neurobiol. 2021, 198, 101917. [Google Scholar] [CrossRef]
- Jia, X.; Gao, Z.; Hu, H. Microglia in depression: Current perspectives. Sci. China Life Sci. 2021, 64, 911–925. [Google Scholar] [CrossRef]
- Colasanto, M.; Madigan, S.; Korczak, D.J. Depression and inflammation among children and adolescents: A meta-analysis. J. Affect. Disord. 2020, 277, 940–948. [Google Scholar] [CrossRef]
- Peters, A.T.; Ren, X.; Bessette, K.L.; George, N.; Kling, L.R.; Thies, B.; West, A.E.; Langenecker, S.A.; Pandey, G.N. Inflammation, depressive symptoms, and emotion perception in adolescence. J. Affect. Disord. 2021, 295, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Moons, W.G.; Eisenberger, N.I.; Taylor, S.E. Anger and fear responses to stress have different biological profiles. Brain Behav. Immun. 2010, 24, 215–219. [Google Scholar] [CrossRef]
- Yirmiya, R.; Rimmerman, N.; Reshef, R. Depression as a microglial disease. Trends Neurosci. 2015, 38, 637–658. [Google Scholar] [CrossRef]
- Barlow, M.A.; Wrosch, C.; Gouin, J.P.; Kunzmann, U. Is anger, but not sadness, associated with chronic inflammation and illness in older adulthood? Psychol. Aging 2019, 34, 330–340. [Google Scholar] [CrossRef]
- El Oussini, H.; Bayer, H.; Scekic-Zahirovic, J.; Vercruysse, P.; Sinniger, J.; Dirrig-Grosch, S.; Dieterlé, S.; Echaniz-Laguna, A.; Larmet, Y.; Müller, K.; et al. Serotonin 2B receptor slows disease progression and prevents degeneration of spinal cord mononuclear phagocytes in amyotrophic lateral sclerosis. Acta Neuropathol. 2016, 131, 465–480. [Google Scholar] [CrossRef]
- Montero-Menei, C.N.; Sindji, L.; Garcion, E.; Mege, M.; Couez, D.; Gamelin, E.; Darcy, F. Early events of the inflammatory reaction induced in rat brain by lipopolysaccharide intracerebral injection: Relative contribution of peripheral monocytes and activated microglia. Brain Res. 1996, 724, 55–66. [Google Scholar] [CrossRef]
- Meulendyke, K.A.; Ubaida-Mohien, C.; Drewes, J.L.; Liao, Z.; Gama, L.; Witwer, K.W.; Graham, D.R.; Zink, M.C. Elevated brain monoamine oxidase activity in SIV- and HIV-associated neurological disease. J. Infect. Dis. 2014, 210, 904–912. [Google Scholar] [CrossRef]
- Hodes, G.E.; Kana, V.; Menard, C.; Merad, M.; Russo, S.J. Neuroimmune mechanisms of depression. Nat. Neurosci. 2015, 18, 1386–1393. [Google Scholar] [CrossRef] [PubMed]
- Johnson, F.K.; Kaffman, A. Early life stress perturbs the function of microglia in the developing rodent brain: New insights and future challenges. Brain Behav. Immun. 2018, 69, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Nestler, E.J. The molecular neurobiology of depression. Nature 2008, 455, 894–902. [Google Scholar] [CrossRef]
- Gong, X.; Chen, Y.; Chang, J.; Huang, Y.; Cai, M.; Zhang, M. Environmental enrichment reduces adolescent anxiety- and depression-like behaviors of rats subjected to infant nerve injury. J. Neuroinflamm. 2018, 15, 262. [Google Scholar] [CrossRef]
- Bedrosian, T.A.; Fonken, L.K.; Walton, J.C.; Haim, A.; Nelson, R.J. Dim light at night provokes depression-like behaviors and reduces CA1 dendritic spine density in female hamsters. Psychoneuroendocrinology 2011, 36, 1062–1069. [Google Scholar] [CrossRef]
- Kitayama, S.; Park, J.; Boylan, J.M.; Miyamoto, Y.; Levine, C.S.; Markus, H.R.; Karasawa, M.; Coe, C.L.; Kawakami, N.; Love, G.D.; et al. Expression of anger and ill health in two cultures: An examination of inflammation and cardiovascular risk. Psychol. Sci. 2015, 26, 211–220. [Google Scholar] [CrossRef]
- Territo, P.R.; Zarrinmayeh, H. P2X7 receptors in neurodegeneration: Potential therapeutic applications from basic to clinical approaches. Front. Cell. Neurosci. 2021, 15, 617036. [Google Scholar] [CrossRef]
- Zha, Z.; Gao, Y.F.; Ji, J.; Sun, Y.Q.; Li, J.L.; Qi, F.; Zhang, N.; Jin, L.Y.; Xue, B.; Yang, T.; et al. Bu Shen Yi Sui capsule alleviates neuroinflammation and demyelination by promoting microglia toward M2 polarization, Which correlates with vhanges in miR-124 and miR-155 in experimental autoimmune encephalomyelitis. Oxidative Med. Cell. Longev. 2021, 2021, 5521503. [Google Scholar] [CrossRef]
- Smythies, L.E.; Smythies, J.R. Microbiota, the immune system, black moods and the brain-melancholia updated. Front. Hum. Neurosci. 2014, 8, 720. [Google Scholar] [CrossRef]
- Wei, Z.X.; Xie, G.J.; Mao, X.; Zou, X.P.; Liao, Y.J.; Liu, Q.S.; Wang, H.; Cheng, Y. Exosomes from patients with major depression cause depressive-like behaviors in mice with involvement of miR-139-5p-regulated neurogenesis. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2020, 45, 1050–1058. [Google Scholar] [CrossRef]
- Jiang, N.M.; Cowan, M.; Moonah, S.N.; Petri, W.A., Jr. The impact of systemic inflammation on neurodevelopment. Trends Mol. Med. 2018, 24, 794–804. [Google Scholar] [CrossRef]
- Bauer, M.E.; Teixeira, A.L. Inflammation in psychiatric disorders: What comes first? Ann. N. Y. Acad. Sci. 2019, 1437, 57–67. [Google Scholar] [CrossRef]
- al’Absi, M.; Bongard, S.; Lovallo, W.R. Adrenocorticotropin responses to interpersonal stress: Effects of overt anger expression style and defensiveness. Int. J. Psychophysiol. Off. J. Int. Organ. Psychophysiol. 2000, 37, 257–265. [Google Scholar] [CrossRef]
- Albert, K.M.; Boyd, B.D.; Taylor, W.D.; Newhouse, P.A. Differential effects of estradiol on neural and emotional stress response in postmenopausal women with remitted major depressive disorder. J. Affect. Disord. 2021, 293, 355–362. [Google Scholar] [CrossRef]
- Szymanska, M.; Schneider, M.; Chateau-Smith, C.; Nezelof, S.; Vulliez-Coady, L. Psychophysiological effects of oxytocin on parent-child interactions: A literature review on oxytocin and parent-child interactions. Psychiatry Clin. Neurosci. 2017, 71, 690–705. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Mao, J.; Li, J. Oxytocin alters the effect of payoff but not base rate in emotion perception. Psychoneuroendocrinology 2020, 114, 104608. [Google Scholar] [CrossRef] [PubMed]
- Zalachoras, I.; Houtman, R.; Atucha, E.; Devos, R.; Tijssen, A.M.; Hu, P.; Lockey, P.M.; Datson, N.A.; Belanoff, J.K.; Lucassen, P.J.; et al. Differential targeting of brain stress circuits with a selective glucocorticoid receptor modulator. Proc. Natl. Acad. Sci. USA 2013, 110, 7910–7915. [Google Scholar] [CrossRef] [PubMed]
- Zannas, A.S.; Jia, M.; Hafner, K.; Baumert, J.; Wiechmann, T.; Pape, J.C.; Arloth, J.; Ködel, M.; Martinelli, S.; Roitman, M.; et al. Epigenetic upregulation of FKBP5 by aging and stress contributes to NF-κB-driven inflammation and cardiovascular risk. Proc. Natl. Acad. Sci. USA 2019, 116, 11370–11379. [Google Scholar] [CrossRef]
- Brent, D.; Melhem, N.; Ferrell, R.; Emslie, G.; Wagner, K.D.; Ryan, N.; Vitiello, B.; Birmaher, B.; Mayes, T.; Zelazny, J.; et al. Association of FKBP5 polymorphisms with suicidal events in the treatment of resistant depression in adolescents (TORDIA) study. Am. J. Psychiatry 2010, 167, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, S.; Anderzhanova, E.A.; Bajaj, T.; Wiechmann, S.; Dethloff, F.; Weckmann, K.; Heinz, D.E.; Ebert, T.; Hartmann, J.; Geiger, T.M.; et al. Stress-primed secretory autophagy promotes extracellular BDNF maturation by enhancing MMP9 secretion. Nat. Commun. 2021, 12, 4643. [Google Scholar] [CrossRef]
- Zheng, D.; Sabbagh, J.J.; Blair, L.J.; Darling, A.L.; Wen, X.; Dickey, C.A. MicroRNA-511 binds to FKBP5 mRNA, Which encodes a chaperone protein, and regulates neuronal differentiation. J. Biol. Chem. 2016, 291, 17897–17906. [Google Scholar] [CrossRef]
- Ozsoy, F.; Nursal, A.F.; Karakus, N.; Demir, M.O.; Yigit, S. Estrogen receptor 1 gene rs22346939 and rs9340799 variants are associated with major depressive disorder and its clinical features. Curr. Neurovascular Res. 2021, 18, 12–19. [Google Scholar] [CrossRef]
- Rahman, T.; Patrick, C.J.; Ma, C.X.; Nicol, G.E.; Reynolds, C.F.; Mulsant, B.H.; Hartz, S.M.; Yingling, M.D.; Lenze, E.J. Prolactin and estrogen levels in postmenopausal women receiving aripiprazole augmentation treatment for depression. J. Clin. Psychopharmacol. 2021, 41, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Ercan, C.M.; Coksuer, H.; Aydogan, U.; Alanbay, I.; Keskin, U.; Karasahin, K.E.; Baser, I. Sexual dysfunction assessment and hormonal correlations in patients with polycystic ovary syndrome. Int. J. Impot. Res. 2013, 25, 127–132. [Google Scholar] [CrossRef] [Green Version]
- Magiakou, M.A.; Mastorakos, G.; Webster, E.; Chrousos, G.P. The hypothalamic-pituitary-adrenal axis and the female reproductive system. Ann. N. Y. Acad. Sci. 1997, 816, 42–56. [Google Scholar] [CrossRef] [PubMed]
- Roa, J.; Garcia-Galiano, D.; Varela, L.; Sánchez-Garrido, M.A.; Pineda, R.; Castellano, J.M.; Ruiz-Pino, F.; Romero, M.; Aguilar, E.; López, M.; et al. The mammalian target of rapamycin as novel central regulator of puberty onset via modulation of hypothalamic Kiss1 system. Endocrinology 2009, 150, 5016–5026. [Google Scholar] [CrossRef]
- Andreescu, C.E.; Alwani, R.A.; Hofland, J.; Looijenga, L.H.; de Herder, W.W.; Hofland, L.J.; Feelders, R.A. Adrenal cushing’s syndrome during pregnancy. Eur. J. Endocrinol. 2017, 177, K13–K20. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.Q.; Yin, Y.; Guo, J.; Gong, X.; Tian, Y.; Shi, L. MTOR-mediated interaction between the oocyte and granulosa cells regulates the development and function of both compartments in mice†. Biol. Reprod. 2022, 107, 76–84. [Google Scholar] [CrossRef]
- Juruena, M.F.; Bocharova, M.; Agustini, B.; Young, A.H. Atypical depression and non-atypical depression: Is HPA axis function a biomarker? A systematic review. J. Affect. Disord. 2018, 233, 45–67. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, J.; You, Z. Switching of the microglial activation phenotype is a possible treatment for depression disorder. Front. Cell. Neurosci. 2018, 12, 306. [Google Scholar] [CrossRef]
- Schmidt, H.D.; Duman, R.S. The role of neurotrophic factors in adult hippocampal neurogenesis, antidepressant treatments and animal models of depressive-like behavior. Behav Pharmacol. 2007, 18, 391–418. [Google Scholar] [CrossRef]
- Prickaerts, J.; De Vry, J.; Boere, J.; Kenis, G.; Quinton, M.S.; Engel, S.; Melnick, L.; Schreiber, R. Differential BDNF responses of triple versus dual reuptake inhibition in neuronal and astrocytoma cells as well as in rat hippocampus and prefrontal cortex. J. Mol. Neurosci. MN 2012, 48, 167–175. [Google Scholar] [CrossRef]
- Phillips, C. Brain-derived neurotrophic factor, depression, and physical activity: Making the neuroplastic connection. Neural Plast. 2017, 2017, 7260130. [Google Scholar] [CrossRef]
- Juric, D.M.; Miklic, S.; Carman-Krzan, M. Monoaminergic neuronal activity up-regulates BDNF synthesis in cultured neonatal rat astrocytes. Brain Res. 2006, 1108, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Mosiołek, A.; Mosiołek, J.; Jakima, S.; Pięta, A.; Szulc, A. Effects of antidepressant treatment on neurotrophic factors (BDNF and IGF-1) in patients with major depressive disorder (MDD). J. Clin. Med. 2021, 10, 3377. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Du, X.; Yin, G.; Zhang, Y.; Li, X.; Cai, J.; Huang, X.; Ning, Y.; Soares, J.C.; Wu, F.; et al. Effects of smoking on cognition and BDNF levels in a male Chinese population: Relationship with BDNF Val66Met polymorphism. Sci. Rep. 2019, 9, 217. [Google Scholar] [CrossRef] [PubMed]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef]
- Wheeler, A.L.; Felsky, D.; Viviano, J.D.; Stojanovski, S.; Ameis, S.H.; Szatmari, P.; Lerch, J.P.; Chakravarty, M.M.; Voineskos, A.N. BDNF-dependent effects on Amygdala-Cortical circuitry and depression risk in children and youth. Cereb. Cortex (New York, NY: 1991) 2018, 28, 1760–1770. [Google Scholar] [CrossRef] [PubMed]
- Kajitani, N.; Hisaoka-Nakashima, K.; Morioka, N.; Okada-Tsuchioka, M.; Kaneko, M.; Kasai, M.; Shibasaki, C.; Nakata, Y.; Takebayashi, M. Antidepressant acts on astrocytes leading to an increase in the expression of neurotrophic/growth factors: Differential regulation of FGF-2 by noradrenaline. PLoS ONE 2012, 7, e51197. [Google Scholar] [CrossRef]
- Zuccarello, D.; Sorrentino, U.; Brasson, V.; Marin, L.; Piccolo, C.; Capalbo, A.; Andrisani, A.; Cassina, M. Epigenetics of pregnancy: Looking beyond the DNA code. J. Assist. Reprod. Genet. 2022, 39, 801–816. [Google Scholar] [CrossRef]
- Vineis, P.; Stringhini, S.; Porta, M. The environmental roots of non-communicable diseases (NCDs) and the epigenetic impacts of globalization. Environ. Res. 2014, 133, 424–430. [Google Scholar] [CrossRef]
- Wang, F.; Pan, F.; Tang, Y.; Huang, J.H. Editorial: Early life stress-induced epigenetic changes involved in mental disorders. Front. Genet. 2021, 12, 684844. [Google Scholar] [CrossRef]
- Misztak, P.; Pańczyszyn-Trzewik, P.; Nowak, G.; Sowa-Kućma, M. Epigenetic marks and their relationship with BDNF in the brain of suicide victims. PLoS ONE 2020, 15, e0239335. [Google Scholar] [CrossRef]
- Ye, H.; Ji, M.; Wang, C.; Wang, C.; Li, Y.; Chen, Y.; Cheng, L.; Li, Y.; Yang, J. Integrated functional neuroimaging, monoamine neurotransmitters, and behavioral score on depressive tendency in intensive care unit medical staffs induced by sleep deprivation after night shift work. Front. Psychiatry 2022, 13, 848709. [Google Scholar] [CrossRef] [PubMed]
- Jarrin, S.; Pandit, A.; Roche, M.; Finn, D.P. Differential role of anterior cingulate cortical glutamatergic neurons in pain-related aversion learning and nociceptive behaviors in male and female rats. Front. Behav. Neurosci. 2020, 14, 139. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.; Upadhya, D.; Hattiangady, B.; Kim, D.K.; An, S.Y.; Shuai, B.; Prockop, D.J.; Shetty, A.K. Intranasal MSC-derived A1-exosomes ease inflammation, and prevent abnormal neurogenesis and memory dysfunction after status epilepticus. Proc. Natl. Acad. Sci. USA 2017, 114, E3536–E3545. [Google Scholar] [CrossRef] [PubMed]
- Buschman, T.J.; Miller, E.K. Top-down versus bottom-up control of attention in the prefrontal and posterior parietal cortices. Science (New York, NY) 2007, 315, 1860–1862. [Google Scholar] [CrossRef]
- Aguilar-Raab, C.; Eckstein, M.; Geracitano, S.; Prevost, M.; Gold, I.; Heinrichs, M.; Bilderbeck, A.; Ehlert, U.; Ditzen, B. Oxytocin modulates the cognitive appraisal of the own and others close intimate relationships. Front. Neurosci. 2019, 13, 714. [Google Scholar] [CrossRef]
- Taylor, C.; Fricker, A.D.; Devi, L.A.; Gomes, I. Mechanisms of action of antidepressants: From neurotransmitter systems to signaling pathways. Cell. Signal. 2005, 17, 549–557. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Zou, D.; Li, Y.; Gu, S.; Dong, J.; Ma, X.; Xu, S.; Wang, F.; Huang, J.H. Monoamine Neurotransmitters Control Basic Emotions and Affect Major Depressive Disorders. Pharmaceuticals 2022, 15, 1203. https://doi.org/10.3390/ph15101203
Jiang Y, Zou D, Li Y, Gu S, Dong J, Ma X, Xu S, Wang F, Huang JH. Monoamine Neurotransmitters Control Basic Emotions and Affect Major Depressive Disorders. Pharmaceuticals. 2022; 15(10):1203. https://doi.org/10.3390/ph15101203
Chicago/Turabian StyleJiang, Yao, Di Zou, Yumeng Li, Simeng Gu, Jie Dong, Xianjun Ma, Shijun Xu, Fushun Wang, and Jason H. Huang. 2022. "Monoamine Neurotransmitters Control Basic Emotions and Affect Major Depressive Disorders" Pharmaceuticals 15, no. 10: 1203. https://doi.org/10.3390/ph15101203