
Aptamer Applications in Emerging Viral Diseases

 , and
, and
Abstract
:1. Introduction
2. Aptamers Targeting Influenza Virus
3. Aptamers against Dengue
4. Aptamers against Rift Valley Fever Virus
5. Aptamers against the ZIKA Virus
6. Aptamers against Japanese Encephalitis Virus
7. Aptamers against Tick-Borne Encephalitis Virus
8. Aptamer Applications in Coronavirus Diseases
9. Aptamers as Diagnostic Tools for SARS-CoV-2 Monitoring and Detection
10. Aptamers as Therapeutics in COVID-19 Disease

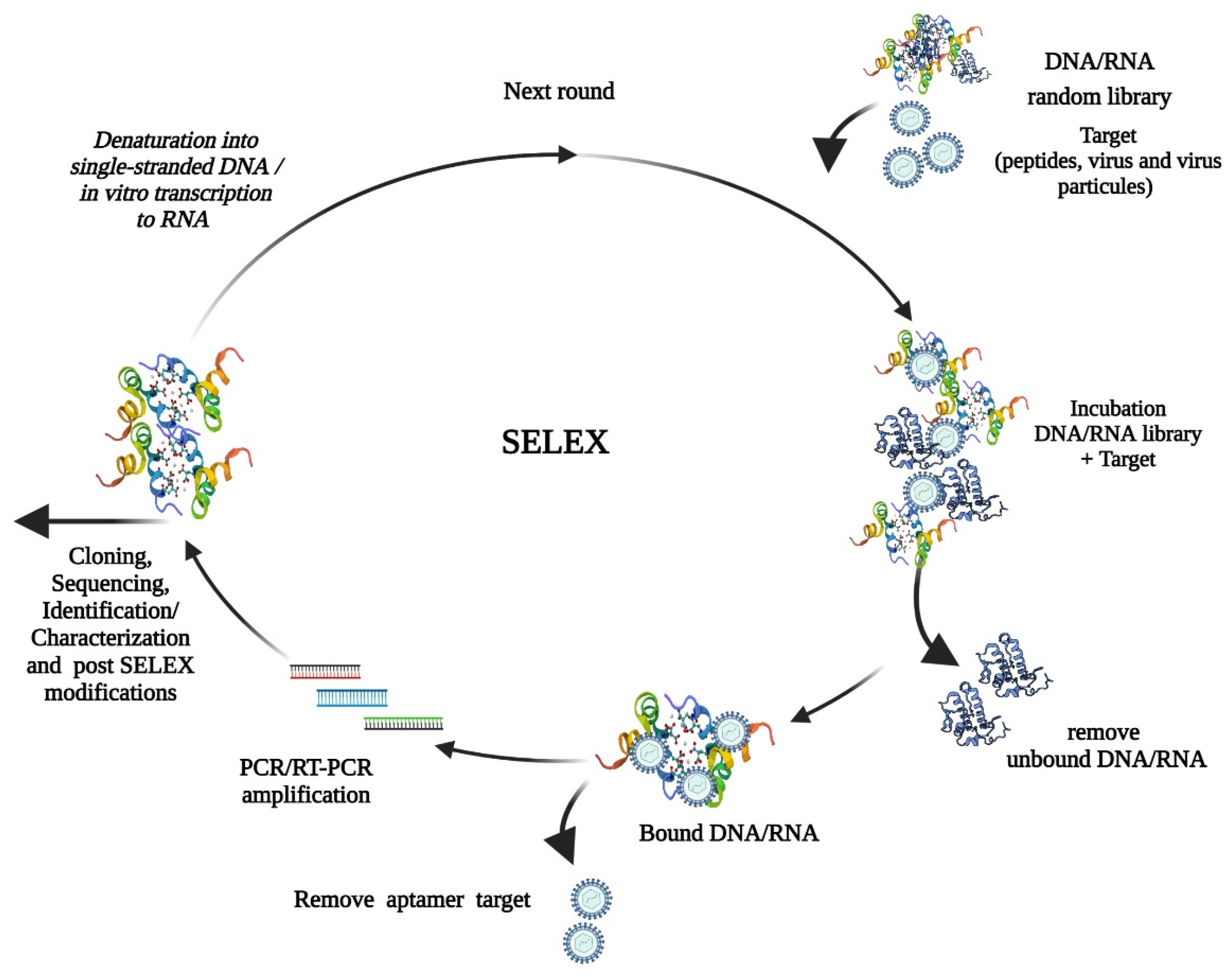{kind=link}
{kind=link}
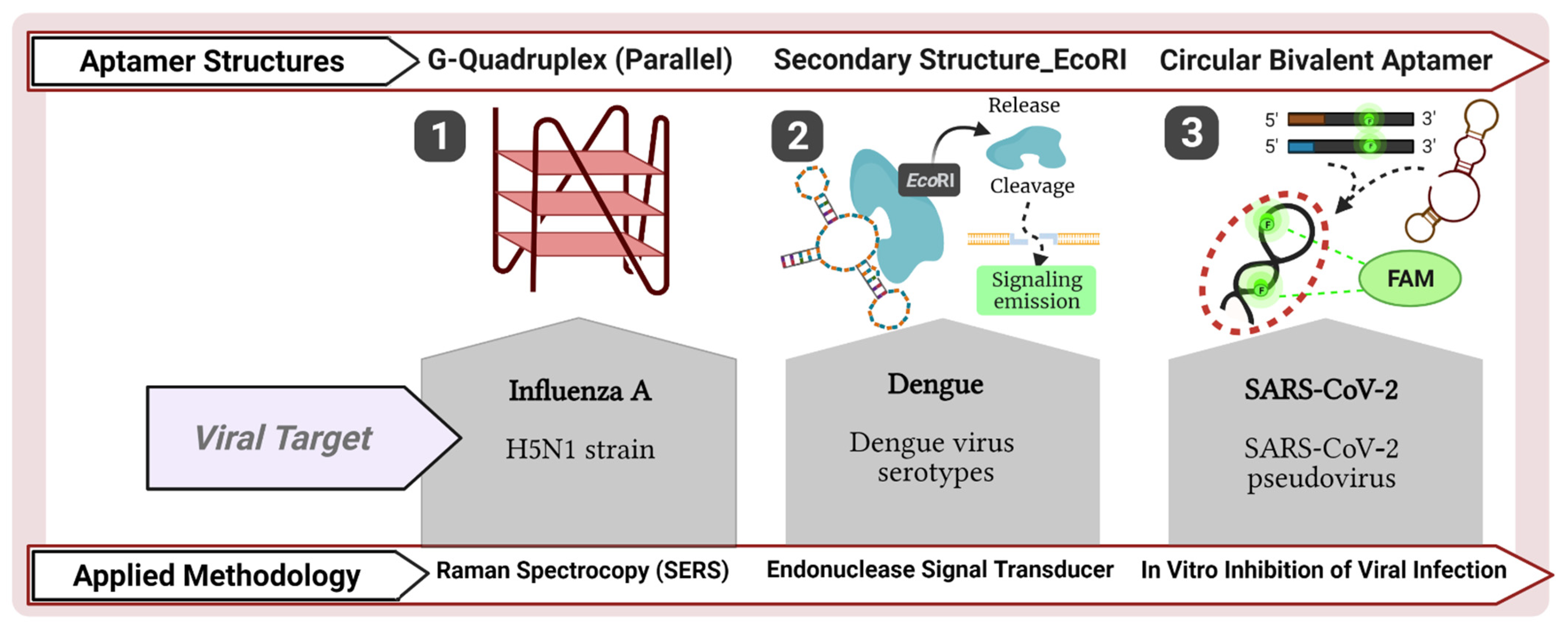{kind=link}
| Name | KD/KM | Type | Length (nt) | Limit of Detection /IC50 | Target | Reference |
|---|---|---|---|---|---|---|
| SARS-CoV therapeutic aptamer applications | ||||||
| ES15 | N.A. | RNA | 107–110 | 1.2 nM (helicase) 77 nM (ATPase) | SARS-CoV nsp10 | [117] |
| NG8 | 5.4 nM | DNA | 93 | 91 nM | SARS-CoV helicase | [119] |
| NG8 modified | 26.8 nM | DNA | 93 | 17.5 nM | SARS-CoV helicase | [119] |
| SARS-CoV diagnostic aptamer applications | ||||||
| Aptamer 1 | 1.65 ± 0.41 nM | RNA | 83 | 20 pg/mL (CLISA) 2 pg/mL (nanoarray chip) | SARS-CoV N protein | [114] |
| Aptamer 1 | 4.93 ± 0.30 nM | DNA | 88 | N.A. | SARS-CoV N protein | [115] |
| Aptamer 1 | See [114] | RNA | 83 | 0.1 pg/mL | SARS-CoV N protein | [116] |
| SARS-CoV-2 therapeutic aptamer applications | ||||||
| cb-CoV2-6C3 | 0.13 nM | DNA | 46 | 0.42 ± 0.15 nM (authentic virus) | SARS-CoV-2 S protein | [51] |
| Aptamer-1 | 6.05 ± 2.06 nM | DNA | 40 | 5.2 nM (inhibition of binding) | SARS-CoV-2 S protein | [127] |
| Aptamer-2 | 6.95 ± 1.10 nM | DNA | 40 | 4.4 nM (inhibition of binding) | SARS-CoV-2 S protein | [127] |
| Aptamer used from [122] | See [122] | DNA | 67 | 130 fg/mL (antigen) 8 particles/mL (virus) | SARS-CoV-2 S protein | [128] |
| SP6 | 21 ± 46 nM | DNA | 87 | 0.2–1 µM (pseudo-virus) | SARS-CoV-2 S protein | [129] |
| SARS-CoV-2 diagnostic aptamer applications | ||||||
| Aptamer 1-3 | N.A. | DNA | 88–57 | 10 ng/mL (ELAA) | SARS-CoV-2 N protein | [121] |
| CoV2-RBD-1C | 5.8 ± 0.8 nM | DNA | 51 | N.A. | SARS-CoV-2 S protein | [122] |
| CoV2-RBD-4C | 19.9 ± 2.6 nM | DNA | 67 | N.A. | SARS-CoV-2 S protein | [122] |
| MG aptamer | N.A. | RNA | 38 | 0.1 aM (in vitro) 1 aM (clinical samples) | Aptamer stabilizes fluorogenic dye | [123] |
| Broccoli | N.A. | RNA | 65 | |||
| Broccoli | N.A. | RNA | 65 | 82 RNA copies (in vitro) 100 RNA copies (SWAB) 500 RNA copies (food) | Aptamer stabilizes fluorogenic dye | [124] |
| A48 | 0.49 ± 0.05 nM | DNA | 58 | 20 pM (recombinant protein) | SARS-CoV-2 N protein | [125] |
| 1C,5′(biotin) | See [122] CoV2-RBD-1C | DNA | 51 | 0.03 – 0.32 fM | SARS-CoV-2 S protein | [111] |
| See [121,122] | N.A. | DNA | 94, 57 | 1.50 pM (N, in vitro) 1.31 pM (S, in vitro) 5.27 pM (N, saliva) 6.31 pM (S, saliva) | SARS-CoV-2 N and S protein | [126] |
11. From Aptamer Selection towards In Vivo Applications Combating Virus Disease
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pimentel, D.; Cooperstein, S.; Randell, H.; Filiberto, D.; Sorrentino, S.; Kaye, B.; Nicklin, C.; Yagi, J.; Brian, J.; O’Hern, J.; et al. Ecology of increasing diseases: Population growth and environmental degradation. Hum. Ecol. Interdiscip. J. 2007, 35, 653–668. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.K.; Kwiatkowski, D.P.; Sabeti, P.C. Natural selection and infectious disease in human populations. Nat. Rev. Genet. 2014, 15, 379–393. [Google Scholar] [CrossRef]
- Wilson, B.A.; Garud, N.R.; Feder, A.F.; Assaf, Z.J.; Pennings, P.S. The population genetics of drug resistance evolution in natural populations of viral, bacterial and eukaryotic pathogens. Mol. Ecol. 2016, 25, 42–66. [Google Scholar] [CrossRef]
- Li, H.-Y.; Jia, W.-N.; Li, X.-Y.; Zhang, L.; Liu, C.; Wu, J. Advances in detection of infectious agents by aptamer-based technologies. Emerg. Microbes Infect. 2020, 9, 1671–1681. [Google Scholar] [CrossRef] [PubMed]
- Goud, K.Y.; Reddy, K.K.; Khorshed, A.; Kumar, V.S.; Mishra, R.K.; Oraby, M.; Ibrahim, A.H.; Kim, H.; Gobi, K.V. Electrochemical diagnostics of infectious viral diseases: Trends and challenges. Biosens. Bioelectron. 2021, 180, 113112. [Google Scholar] [CrossRef]
- Madhav, N.; Oppenheim, B.; Gallivan, M.; Mulembakani, P.; Rubin, E.; Wolfe, N. Pandemics: Risks, impacts, and mitigation. In Disease Control Priorities: Improving Health and Reducing Poverty; Jamison, D.T., Gelband, H., Horton, S., Jha, P., Laxminarayan, R., Mock, C.N., Nugent, R., Eds.; The International Bank for Reconstruction and Development/The World Bank: Washington, DC, USA, 2017; ISBN 978-1-4648-0527-1. [Google Scholar]
- Wędrowska, E.; Wandtke, T.; Piskorska, E.; Kopiński, P. The latest achievements in the construction of influenza virus detection aptasensors. Viruses 2020, 12. [Google Scholar] [CrossRef]
- Labib, M.; Berezovski, M.V. Electrochemical aptasensors for microbial and viral pathogens. Adv. Biochem. Eng. Biotechnol. 2014, 140, 155–181. [Google Scholar] [CrossRef]
- Zhuo, Z.; Yu, Y.; Wang, M.; Li, J.; Zhang, Z.; Liu, J.; Wu, X.; Lu, A.; Zhang, G.; Zhang, B. Recent advances in SELEX technology and aptamer applications in biomedicine. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [Green Version]
- Nery, A.A.; Wrenger, C.; Ulrich, H. Recognition of biomarkers and cell-specific molecular signatures: Aptamers as capture agents. J. Sep. Sci. 2009, 32, 1523–1530. [Google Scholar] [CrossRef]
- Acquah, C.; Danquah, M.K.; Agyei, D.; Moy, C.K.S.; Sidhu, A.; Ongkudon, C.M. Deploying aptameric sensing technology for rapid pandemic monitoring. Crit. Rev. Biotechnol. 2016, 36, 1010–1022. [Google Scholar] [CrossRef] [Green Version]
- Vidic, M.; Smuc, T.; Janez, N.; Blank, M.; Accetto, T.; Mavri, J.; Nascimento, I.C.; Nery, A.A.; Ulrich, H.; Lah, T.T. In silico selection approach to develop dna aptamers for a stem-like cell subpopulation of non-small lung cancer adenocarcinoma cell line A549. Radiol. Oncol. 2018, 52, 152–159. [Google Scholar] [CrossRef] [Green Version]
- Kumar Kulabhusan, P.; Hussain, B.; Yüce, M. Current perspectives on aptamers as diagnostic tools and therapeutic agents. Pharmaceutics 2020, 12, 646. [Google Scholar] [CrossRef]
- Nascimento, I.C.; Nery, A.A.; Bassaneze, V.; Krieger, J.E.; Ulrich, H. Applications of aptamers in flow and imaging cytometry. Methods Mol. Biol. 2016, 1380, 127–134. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, H.; Jain, A.; Zhang, L.; Liu, C.; Cheng, K. Discovery of aptamer ligands for hepatic stellate cells using SELEX. Theranostics 2017, 7, 2982–2995. [Google Scholar] [CrossRef] [Green Version]
- Ulrich, H.; Martins, A.H.B.; Pesquero, J.B. RNA and DNA aptamers in cytomics analysis. Cytom. A 2004, 59, 220–231. [Google Scholar] [CrossRef]
- Faria, M.; Ulrich, H. Sugar boost: When ribose modifications improve oligonucleotide performance. Curr. Opin. Mol. Ther. 2008, 10, 168–175. [Google Scholar]
- Gulyaeva, L.F.; Kushlinskiy, N.E. Regulatory mechanisms of MicroRNA expression. J. Transl. Med. 2016, 14, 143. [Google Scholar] [CrossRef] [Green Version]
- Westhof, E.; Fritsch, V. RNA folding: Beyond watson-crick pairs. Structure 2000, 8, R55–R65. [Google Scholar] [CrossRef] [Green Version]
- Germer, K.; Leonard, M.; Zhang, X. RNA Aptamers and their therapeutic and diagnostic applications. Int. J. Biochem. Mol. Biol. 2013, 4, 27–40. [Google Scholar]
- Wang, T.; Chen, C.; Larcher, L.M.; Barrero, R.A.; Veedu, R.N. Three decades of nucleic acid aptamer technologies: Lessons learned, progress and opportunities on aptamer development. Biotechnol. Adv. 2019, 37, 28–50. [Google Scholar] [CrossRef]
- Ulrich, H.; Wrenger, C. Disease-specific biomarker discovery by aptamers. Cytometry A 2009, 75, 727–733. [Google Scholar] [CrossRef]
- Liang, Z.-M.; Peng, Y.-H.; Chen, Y.; Long, L.-L.; Luo, H.-J.; Chen, Y.-J.; Liang, Y.-L.; Tian, Y.-H.; Li, S.-J.; Shi, Y.-S.; et al. The BACE1-specific DNA aptamer A1 rescues amyloid-β pathology and behavioral deficits in a mouse model of Alzheimer’s disease. Nucleic Acid Ther. 2019, 29, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.L.; Nascimento, I.C.; Santos, A.P.; Ogusuku, I.E.Y.; Lameu, C.; Mayer, G.; Ulrich, H. Aptamers: Novelty tools for cancer biology. Oncotarget 2018, 9, 26934–26953. [Google Scholar] [CrossRef] [Green Version]
- Yasmeen, F.; Seo, H.; Javaid, N.; Kim, M.S.; Choi, S. Therapeutic interventions into innate immune diseases by means of aptamers. Pharmaceutics 2020, 12, 955. [Google Scholar] [CrossRef]
- Ulrich, H.; Magdesian, M.H.; Alves, M.J.M.; Colli, W. In vitro selection of RNA aptamers that bind to cell adhesion receptors of trypanosoma cruzi and inhibit cell invasion. J. Biol. Chem. 2002, 277, 20756–20762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimbres, F.M.; Tárnok, A.; Ulrich, H.; Wrenger, C. Aptamers: Novel molecules as diagnostic markers in bacterial and viral infections? Biomed. Res. Int. 2013, 2013, 731516. [Google Scholar] [CrossRef] [Green Version]
- Delač, M.; Motaln, H.; Ulrich, H.; Lah, T.T. Aptamer for imaging and therapeutic targeting of brain tumor glioblastoma. Cytom. A 2015, 87, 806–816. [Google Scholar] [CrossRef] [Green Version]
- Linzke, M.; Yan, S.L.R.; Tárnok, A.; Ulrich, H.; Groves, M.R.; Wrenger, C. Live and Let Dye: Visualizing the cellular compartments of the malaria parasite plasmodium falciparum. Cytometry A 2020, 97, 694–705. [Google Scholar] [CrossRef]
- Bai, H.; Wang, R.; Hargis, B.; Lu, H.; Li, Y. A SPR aptasensor for detection of avian influenza virus H5N1. Sensors 2012, 12, 12506–12518. [Google Scholar] [CrossRef] [Green Version]
- Wandtke, T.; Woźniak, J.; Kopiński, P. Aptamers in diagnostics and treatment of viral infections. Viruses 2015, 7, 751–780. [Google Scholar] [CrossRef] [Green Version]
- Ospina-Villa, J.D.; López-Camarillo, C.; Castañón-Sánchez, C.A.; Soto-Sánchez, J.; Ramírez-Moreno, E.; Marchat, L.A. Advances on aptamers against protozoan parasites. Genes 2018, 9, 584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, P.; Li, W.; Li, J. Applications of aptasensors in clinical diagnostics. Sensors 2012, 12, 1181–1193. [Google Scholar] [CrossRef]
- Singh, R.; Hong, S.; Jang, J. Label-free detection of influenza viruses using a reduced graphene oxide-based electrochemical immunosensor integrated with a microfluidic platform. Sci. Rep. 2017, 7, 42771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseinzadeh, L.; Mazloum-Ardakani, M. Advances in aptasensor technology. Adv. Clin. Chem. 2020, 99, 237–279. [Google Scholar] [CrossRef]
- Ambartsumyan, O.; Gribanyov, D.; Kukushkin, V.; Kopylov, A.; Zavyalova, E. SERS-based biosensors for virus determination with oligonucleotides as recognition elements. Int. J. Mol. Sci. 2020, 21, 3373. [Google Scholar] [CrossRef]
- Zou, X.; Wu, J.; Gu, J.; Shen, L.; Mao, L. Application of aptamers in virus detection and antiviral therapy. Front. Microbiol. 2019, 10, 1462. [Google Scholar] [CrossRef] [Green Version]
- Van den Kieboom, C.H.; van der Beek, S.L.; Mészáros, T.; Gyurcsányi, R.E.; Ferwerda, G.; de Jonge, M.I. Aptasensors for viral diagnostics. Trends Analyt. Chem. 2015, 74, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.J.; Jin, F.G.; Wang, P.; Wang, M.; Zhu, J.M. Isolation of influenza C virus from pigs and experimental infection of pigs with influenza C virus. J. Gen. Virol. 1983, 64 Pt 1, 177–182. [Google Scholar] [CrossRef]
- Horimoto, T.; Kawaoka, Y. Influenza: Lessons from past pandemics, warnings from current incidents. Nat. Rev. Microbiol. 2005, 3, 591–600. [Google Scholar] [CrossRef]
- Taubenberger, J.K.; Morens, D.M. The pathology of influenza virus infections. Ann. Rev. Pathol. 2008, 3, 499–522. [Google Scholar] [CrossRef]
- Sherman, A.C.; Mehta, A.; Dickert, N.W.; Anderson, E.J.; Rouphael, N. The Future of flu: A review of the human challenge model and systems biology for advancement of influenza vaccinology. Front. Cell. Infect. Microbiol. 2019, 9, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.-J.; Yeh, C.-Y.; Cheng, J.-C.; Huang, Y.-Q.; Hsu, K.-C.; Lin, Y.-F.; Lu, C.-H. Potent sialic acid inhibitors that target influenza A virus hemagglutinin. Sci. Rep. 2021, 11, 8637. [Google Scholar] [CrossRef]
- Woźniak-Kosek, A.; Kempińska-Mirosławska, B.; Hoser, G. Detection of the influenza virus yesterday and now. Acta Biochim. Pol. 2014, 61, 465–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labella, A.M.; Merel, S.E. Influenza. Med. Clin. N. Am. 2013, 97, 621–645. [Google Scholar] [CrossRef]
- Tam, P.D.; Van Hieu, N.; Chien, N.D.; Le, A.-T.; Anh Tuan, M. DNA sensor development based on multi-wall carbon nanotubes for label-free influenza virus (type A) detection. J. Immunol. Methods 2009, 350, 118–124. [Google Scholar] [CrossRef]
- Gribanyov, D.; Zhdanov, G.; Olenin, A.; Lisichkin, G.; Gambaryan, A.; Kukushkin, V.; Zavyalova, E. SERS-based colloidal aptasensors for quantitative determination of influenza virus. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Bizyaeva, A.A.; Bunin, D.A.; Moiseenko, V.L.; Gambaryan, A.S.; Balk, S.; Tashlitsky, V.N.; Arutyunyan, A.M.; Kopylov, A.M.; Zavyalova, E.G. The functional role of loops and flanking sequences of G-quadruplex aptamer to the hemagglutinin of influenza a virus. Int. J. Mol. Sci. 2021, 22, 2409. [Google Scholar] [CrossRef]
- Dong, S.; Zhao, R.; Zhu, J.; Lu, X.; Li, Y.; Qiu, S.; Jia, L.; Jiao, X.; Song, S.; Fan, C.; et al. Electrochemical DNA biosensor based on a tetrahedral nanostructure probe for the detection of avian influenza A (H7N9) virus. ACS Appl. Mater. Interfaces 2015, 7, 8834–8842. [Google Scholar] [CrossRef]
- Fletcher, S.J.; Phillips, L.W.; Milligan, A.S.; Rodda, S.J. Toward specific detection of dengue virus serotypes using a novel modular biosensor. Biosens. Bioelectron. 2010, 26, 1696–1700. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Liu, S.; Wei, X.; Wan, S.; Huang, M.; Song, T.; Lu, Y.; Weng, X.; Lin, Z.; Chen, H.; et al. Aptamer blocking strategy inhibits SARS-CoV-2 virus infection. Angew. Chem. Int. Ed. Engl. 2021, 60, 10266–10272. [Google Scholar] [CrossRef]
- Lum, J.; Wang, R.; Lassiter, K.; Srinivasan, B.; Abi-Ghanem, D.; Berghman, L.; Hargis, B.; Tung, S.; Lu, H.; Li, Y. Rapid detection of avian influenza H5N1 virus using impedance measurement of immuno-reaction coupled with RBC amplification. Biosens. Bioelectron. 2012, 38, 67–73. [Google Scholar] [CrossRef]
- Karash, S.; Wang, R.; Kelso, L.; Lu, H.; Huang, T.J.; Li, Y. Rapid detection of avian influenza virus H5N1 in chicken tracheal samples using an impedance aptasensor with gold nanoparticles for signal amplification. J. Virol. Methods 2016, 236, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Park, J.A.; Kim, J.; Kim, S.M.; Sohn, H.; Park, C.; Kim, T.-H.; Lee, J.-H.; Lee, M.-H.; Lee, T. Fabrication of electrochemical influenza virus (H1N1) biosensor composed of multifunctional DNA four-way junction and molybdenum disulfide hybrid material. Materials 2021, 14, 343. [Google Scholar] [CrossRef]
- Pang, Y.; Rong, Z.; Wang, J.; Xiao, R.; Wang, S. A fluorescent aptasensor for H5N1 influenza virus detection based-on the core-shell nanoparticles metal-enhanced fluorescence (MEF). Biosens. Bioelectron. 2015, 66, 527–532. [Google Scholar] [CrossRef]
- Zhang, J.; Tian, J.; He, Y.; Chen, S.; Jiang, Y.; Zhao, Y.; Zhao, S. Protein-binding aptamer assisted signal amplification for the detection of influenza A (H1N1) DNA sequences based on quantum dot fluorescence polarization analysis. Analyst 2013, 138, 4722–4727. [Google Scholar] [CrossRef]
- Tseng, Y.-T.; Wang, C.-H.; Chang, C.-P.; Lee, G.-B. Integrated microfluidic system for rapid detection of influenza H1N1 virus using a sandwich-based aptamer assay. Biosens. Bioelectron. 2016, 82, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-H.; Chang, C.-P.; Lee, G.-B. Integrated microfluidic device using a single universal aptamer to detect multiple types of influenza viruses. Biosens. Bioelectron. 2016, 86, 247–254. [Google Scholar] [CrossRef]
- Kiilerich-Pedersen, K.; Daprà, J.; Cherré, S.; Rozlosnik, N. High sensitivity point-of-care device for direct virus diagnostics. Biosens. Bioelectron. 2013, 49, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Kirkegaard, J.; Rozlosnik, N. Screen-printed all-polymer aptasensor for impedance based detection of influenza A virus. Methods Mol. Biol. 2017, 1572, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.; Lu, Z.; Jiang, H.; Yang, Z.; Liu, X.; Ding, H.; Li, H.; Dong, J.; Huang, A.; Fang, T.; et al. Aptamer selection and application in multivalent binding-based electrical impedance detection of inactivated H1N1 virus. Biosens. Bioelectron. 2018, 110, 162–167. [Google Scholar] [CrossRef]
- Liu, X.; Cheng, Z.; Fan, H.; Ai, S.; Han, R. Electrochemical detection of avian influenza virus H5N1 gene sequence using a DNA aptamer immobilized onto a hybrid nanomaterial-modified electrode. Electrochim. Acta 2011, 56, 6266–6270. [Google Scholar] [CrossRef]
- Bhardwaj, J.; Sharma, A.; Jang, J. Vertical flow-based paper immunosensor for rapid electrochemical and colorimetric detection of influenza virus using a different pore size sample pad. Biosens. Bioelectron. 2019, 126, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wang, R.; Kelso, L.C.; Ying, Y.; Li, Y. A target-responsive and size-dependent hydrogel aptasensor embedded with QD fluorescent reporters for rapid detection of avian influenza virus H5N1. Sens. Actuators B Chem. 2016, 234, 98–108. [Google Scholar] [CrossRef]
- Lai, W.-A.; Lin, C.-H.; Yang, Y.-S.; Lu, M.S.-C. Ultrasensitive and label-free detection of pathogenic avian influenza DNA by using CMOS impedimetric sensors. Biosens. Bioelectron. 2012, 35, 456–460. [Google Scholar] [CrossRef]
- Zhu, X.; Ai, S.; Chen, Q.; Yin, H.; Xu, J. Label-free electrochemical detection of avian influenza virus genotype utilizing multi-walled carbon nanotubes–Cobalt phthalocyanine–PAMAM nanocomposite modified glassy carbon electrode. Electrochem. Commun. 2009, 7, 1543–1546. [Google Scholar] [CrossRef]
- Kushwaha, A.; Takamura, Y.; Nishigaki, K.; Biyani, M. Competitive non-SELEX for the selective and rapid enrichment of DNA aptamers and its use in electrochemical aptasensor. Sci. Rep. 2019, 9, 6642. [Google Scholar] [CrossRef] [PubMed]
- Moulick, A.; Richtera, L.; Milosavljevic, V.; Cernei, N.; Haddad, Y.; Zitka, O.; Kopel, P.; Heger, Z.; Adam, V. Advanced nanotechnologies in avian influenza: Current status and future trends—A review. Anal. Chim. Acta 2017, 983, 42–53. [Google Scholar] [CrossRef]
- Turner, D.; Baldwin, E.; Russell, K.; Wells, L.A. DNA-crosslinked alginate and layered microspheres to modulate the release of encapsulated FITC-dextran. Eur. J. Pharm. Biopharm. 2021, 158, 313–322. [Google Scholar] [CrossRef]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antiviral. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [Green Version]
- Argondizzo, A.P.C.; Silva, D.; Missailidis, S. Application of aptamer-based assays to the diagnosis of arboviruses important for public health in Brazil. Int. J. Mol. Sci. 2020, 22, 159. [Google Scholar] [CrossRef]
- Gubler, D.; Kuno, G.; Markoff, L. Flaviviruses. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 2007; pp. 1153–1251. [Google Scholar]
- Lindenbach, B.; Thiel, H.J.; Rice, C.M. Flaviviridae: The viruses and their replication. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 2007; pp. 1101–1151. [Google Scholar]
- Henderson, B.R.; Saeedi, B.J.; Campagnola, G.; Geiss, B.J. Analysis of RNA binding by the dengue virus NS5 RNA capping enzyme. PLoS ONE 2011, 6, e25795. [Google Scholar] [CrossRef]
- Figueiredo, L.T. The brazilian flaviviruses. Microbes Infect. 2000, 2, 1643–1649. [Google Scholar] [CrossRef]
- Lopes, N.; Nozawa, C.; Linhares, R.E.C. General features and epidemiology of emerging arboviruses in Brazil/Características gerais e epidemiologia dos arbovírus emergentes no Brasil. Rev. Pan Amaz Saude 2014, 5, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Gandham, S.H.A.; Volk, D.E.; Lokesh, G.L.R.; Neerathilingam, M.; Gorenstein, D.G. Thioaptamers targeting dengue virus Type-2 envelope protein domain III. Biochem. Biophys. Res. Commun. 2014, 453, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-L.; Hsiao, W.-H.; Lee, H.-C.; Wu, S.-C.; Cheng, J.-W. Selection and characterization of DNA aptamers targeting all four serotypes of dengue viruses. PLoS ONE 2015, 10, e0131240. [Google Scholar] [CrossRef]
- Balkhy, H.H.; Memish, Z.A. Rift Valley Fever: An uninvited zoonosis in the arabian peninsula. Int. J. Antimicrob. Agents 2003, 21, 153–157. [Google Scholar] [CrossRef]
- Kilgore, P.E.; Ksiazek, T.G.; Rollin, P.E.; Mills, J.N.; Villagra, M.R.; Montenegro, M.J.; Costales, M.A.; Paredes, L.C.; Peters, C.J. Treatment of bolivian hemorrhagic fever with intravenous ribavirin. Clin. Infect. Dis. 1997, 24, 718–722. [Google Scholar] [CrossRef] [Green Version]
- McCormick, J.B.; King, I.J.; Webb, P.A.; Scribner, C.L.; Craven, R.B.; Johnson, K.M.; Elliott, L.H.; Belmont-Williams, R. Lassa fever. Effective therapy with ribavirin. N. Engl. J. Med. 1986, 314, 20–26. [Google Scholar] [CrossRef]
- Monath, T.P. Treatment of yellow fever. Antivir. Res. 2008, 78, 116–124. [Google Scholar] [CrossRef]
- Ellenbecker, M.; Sears, L.; Li, P.; Lanchy, J.-M.; Lodmell, J.S. Characterization of RNA aptamers directed against the nucleocapsid protein of Rift Valley Fever virus. Antivir. Res. 2012, 93, 330–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.H.; Zeng, H. Aptamer-based ELISA assay for highly specific and sensitive detection of Zika NS1 protein. Anal. Chem. 2017, 89, 12743–12748. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.T.H.; Bao, D.T.; Park, H.; Ngoc, N.M.; Yeo, S.-J. Development of a novel peptide aptamer-based immunoassay to detect Zika virus in serum and urine. Theranostics 2018, 8, 3629–3642. [Google Scholar] [CrossRef]
- Lessler, J.; Chaisson, L.H.; Kucirka, L.M.; Bi, Q.; Grantz, K.; Salje, H.; Carcelen, A.C.; Ott, C.T.; Sheffield, J.S.; Ferguson, N.M.; et al. Assessing the global threat from Zika virus. Science 2016, 353, aaf8160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Linden, V.; Pessoa, A.; Dobyns, W.; Barkovich, A.J.; Júnior, H. van der L.; Filho, E.L.R.; Ribeiro, E.M.; Leal, M.d.C.; Coimbra, P.P.d.A.; Aragão, M.d.F.V.V.; et al. Description of 13 infants born during October 2015–January 2016 with congenital Zika Virus infection without microcephaly at birth—Brazil. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Göertz, G.P.; Vogels, C.B.F.; Geertsema, C.; Koenraadt, C.J.M.; Pijlman, G.P. Mosquito co-infection with Zika and chikungunya virus allows simultaneous transmission without affecting vector competence of Aedes Aegypti. PLoS Negl Trop. Dis. 2017, 11, e0005654. [Google Scholar] [CrossRef] [Green Version]
- Saraf, N.; Villegas, M.; Willenberg, B.J.; Seal, S. Multiplex viral detection platform based on a aptamers-integrated microfluidic channel. ACS Omega 2019, 4, 2234–2240. [Google Scholar] [CrossRef] [PubMed]
- Souf, S. Recent advances in diagnostic testing for viral infections. Biosci. Horiz. Int. J. Stud. Res. 2016, 9. [Google Scholar] [CrossRef]
- Schiøler, K.L.; Samuel, M.; Wai, K.L. Vaccines for preventing Japanese encephalitis. In Cochrane Database System Reviews; John Wiley & Sons: Hoboken, NJ, USA, 2007. [Google Scholar] [CrossRef]
- Han, S.R.; Lee, S.-W. Inhibition of Japanese encephalitis virus (JEV) replication by specific RNA APTAMER against JEV methyltransferase. Biochem. Biophys. Res. Commun. 2017, 483, 687–693. [Google Scholar] [CrossRef]
- Jung, J.I.; Han, S.R.; Lee, S.-W. Development of RNA aptamer that inhibits methyltransferase activity of dengue virus. Biotechnol. Lett. 2018, 40, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, L.; Vapalahti, O. Tick-borne encephalitis. Lancet 2008, 371, 1861–1871. [Google Scholar] [CrossRef]
- Kondratov, I.G.; Khasnatinov, M.A.; Potapova, U.V.; Potapov, V.V.; Levitskii, S.A.; Leonova, G.N.; Pavlenko, E.V.; Solovarov, I.S.; Denikina, N.N.; Kulakova, N.V.; et al. Obtaining aptamers to a fragment of surface protein E of tick-borne encephalitis virus. Dokl. Biochem. Biophys. 2013, 448, 19–21. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.; Drosten, C.; Müller, M.A. Serological assays for emerging coronaviruses: Challenges and pitfalls. Virus Res. 2014, 194, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.M.; Cavanagh, D. The Molecular biology of coronaviruses. Adv. Virus Res. 1997, 48, 1–100. [Google Scholar]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, genetic recombination, and pathogenesis of coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [Green Version]
- Weiss, S.R.; Navas-Martin, S. Coronavirus pathogenesis and the emerging pathogen severe acute respiratory syndrome coronavirus. Microbiol. Mol. Biol. Rev. 2005, 69, 635–664. [Google Scholar] [CrossRef] [Green Version]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taxonomy. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 2 May 2021).
- Wertheim, J.O.; Chu, D.K.W.; Peiris, J.S.M.; Kosakovsky Pond, S.L.; Poon, L.L.M. A case for the ancient origin of coronaviruses. J. Virol. 2013, 87, 7039–7045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, A.; Drosten, C.; Doerr, H.W.; Stürmer, M.; Preiser, W. Severe acute respiratory syndrome (SARS)—Paradigm of an Emerging Viral Infection. J. Clin. Virol. 2004, 29, 13–22. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, K.; Perlman, S. Coronaviruses, including severe acute respiratory syndrome (SARS) and Middle East respiratory syndrome (MERS). Mandell Douglas Bennetts Princ. Pract. Infect. Dis. 2015, 1928–1936.e2. [Google Scholar] [CrossRef]
- Tyrrell, D.A.; Bynoe, M.L. Cultivation of a novel type of common-cold virus in organ cultures. Br. Med. J. 1965, 1, 1467–1470. [Google Scholar] [CrossRef] [Green Version]
- Anonymous. Virology: Coronaviruses. Nature 1968, 220, 650. [Google Scholar] [CrossRef] [Green Version]
- Lau, S.K.P.; Lee, P.; Tsang, A.K.L.; Yip, C.C.Y.; Tse, H.; Lee, R.A.; So, L.-Y.; Lau, Y.-L.; Chan, K.-H.; Woo, P.C.Y.; et al. Molecular epidemiology of human coronavirus OC43 reveals evolution of different genotypes over time and recent emergence of a novel genotype due to natural recombination. J. Virol. 2011, 85, 11325–11337. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.-L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef]
- Acquah, C.; Jeevanandam, J.; Tan, K.X.; Danquah, M.K. Engineered aptamers for enhanced COVID-19 Theranostics. Cell. Mol. Bioeng. 2021, 1–13. [Google Scholar] [CrossRef]
- Stanborough, T.; Given, F.M.; Koch, B.; Sheen, C.R.; Stowers-Hull, A.B.; Waterland, M.R.; Crittenden, D.L. Optical detection of CoV-SARS-2 viral proteins to sub-picomolar concentrations. ACS Omega 2021, 6, 6404–6413. [Google Scholar] [CrossRef]
- Byun, J. Recent Progress and Opportunities for Nucleic Acid Aptamers. Life 2021, 11, 193. [Google Scholar] [CrossRef]
- Torabi, R.; Ranjbar, R.; Halaji, M.; Heiat, M. Aptamers, the bivalent agents as probes and therapies for coronavirus infections: A systematic review. Mol. Cell. Probes 2020, 53, 101636. [Google Scholar] [CrossRef] [PubMed]
- Ahn, D.-G.; Jeon, I.-J.; Kim, J.D.; Song, M.-S.; Han, S.-R.; Lee, S.-W.; Jung, H.; Oh, J.-W. RNA aptamer-based sensitive detection of SARS coronavirus nucleocapsid protein. Analyst 2009, 134, 1896–1901. [Google Scholar] [CrossRef]
- Cho, S.-J.; Woo, H.-M.; Kim, K.-S.; Oh, J.-W.; Jeong, Y.-J. Novel system for detecting SARS coronavirus nucleocapsid protein using an SsDNA aptamer. J. Biosci. Bioeng. 2011, 112, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Roh, C.; Jo, S.K. Quantitative and sensitive detection of SARS coronavirus nucleocapsid protein using quantum dots-conjugated RNA aptamer on chip. J. Chem. Technol. Biotechnol. 2011, 86, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Jang, K.J.; Lee, N.-R.; Yeo, W.-S.; Jeong, Y.-J.; Kim, D.-E. Isolation of inhibitory RNA aptamers against severe acute respiratory syndrome (SARS) coronavirus NTPase/Helicase. Biochem. Biophys. Res. Commun. 2008, 366, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhou, Q.; Li, Y.; Garner, L.V.; Watkins, S.P.; Carter, L.J.; Smoot, J.; Gregg, A.C.; Daniels, A.D.; Jervey, S.; et al. Research and development on therapeutic agents and vaccines for COVID-19 and related human coronavirus diseases. ACS Cent. Sci. 2020, 6, 315–331. [Google Scholar] [CrossRef]
- Shum, K.T.; Tanner, J.A. Differential inhibitory activities and stabilisation of DNA aptamers against the SARS coronavirus helicase. ChemBioChem 2008, 9, 3037–3045. [Google Scholar] [CrossRef]
- Rutschke, N.; Zimmermann, J.; Möller, R.; Klöck, G.; Winterhalter, M.; Leune, A. Hot start reverse transcriptase: An approach for improved real-time RT-PCR performance. J. Anal. Sci. Technol. 2015, 6, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Wu, Q.; Chen, J.; Ni, X.; Dai, J. A DNA aptamer based method for detection of SARS-CoV-2 nucleocapsid protein. Virol. Sin. 2020, 35, 351–354. [Google Scholar] [CrossRef]
- Song, Y.; Song, J.; Wei, X.; Huang, M.; Sun, M.; Zhu, L.; Lin, B.; Shen, H.; Zhu, Z.; Yang, C. Discovery of aptamers targeting the receptor-binding domain of the SARS-CoV-2 spike glycoprotein. Anal. Chem. 2020, 92, 9895–9900. [Google Scholar] [CrossRef]
- Woo, C.H.; Jang, S.; Shin, G.; Jung, G.Y.; Lee, J.W. Sensitive fluorescence detection of SARS-CoV-2 RNA in clinical samples via one-pot isothermal ligation and transcription. Nat. Biomed. Eng. 2020, 4, 1168–1179. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; Chen, J.; Wang, M.; Zhang, T.; Luo, W.; Li, Y.; Wu, Y.; Zeng, B.; Zhang, K.; et al. Detection of SARS-CoV-2 and its mutated variants via CRISPR-Cas13-based transcription amplification. Anal. Chem. 2021, 93, 3393–3402. [Google Scholar] [CrossRef]
- Zhang, L.; Fang, X.; Liu, X.; Ou, H.; Zhang, H.; Wang, J.; Li, Q.; Cheng, H.; Zhang, W.; Luo, Z. Discovery of sandwich type COVID-19 nucleocapsid protein DNA aptamers. Chem. Commun. 2020, 56, 10235–10238. [Google Scholar] [CrossRef]
- Singh, N.K.; Ray, P.; Carlin, A.F.; Magallanes, C.; Morgan, S.C.; Laurent, L.C.; Aronoff-Spencer, E.S.; Hall, D.A. Hitting the diagnostic sweet spot: Point-of-care SARS-CoV-2 salivary antigen testing with an off-the-shelf glucometer. Biosens. Bioelectron. 2021, 180, 113111. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Y.-L.; Wu, J.; Qi, J.; Zeng, Z.; Wan, Q.; Chen, Z.; Manandhar, P.; Cavener, V.S.; Boyle, N.R.; et al. Neutralizing aptamers block S/RBD-ACE2 interactions and prevent host cell infection. Angew. Chem. Int. Ed. Engl. 2021, 60, 10273–10278. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, A.; Gao, Y.; Patibandla, S.; Mitra, D.; McCandless, M.G.; Fassero, L.A.; Gates, K.; Tandon, R.; Ray, P.C. Aptamer conjugated gold nanostar-based distance-dependent nanoparticle surface energy transfer spectroscopy for ultrasensitive detection and inactivation of corona virus. J. Phys. Chem. Lett. 2021, 12, 2166–2171. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, A.; Weber, A.; Bayin, M.; Breuers, S.; Fieberg, V.; Famulok, M.; Mayer, G. A SARS-CoV-2 spike binding DNA aptamer that inhibits pseudovirus infection by an RBD-independent mechanism *. Angew. Chem. Int. Ed. Engl. 2021, 60, 10279–10285. [Google Scholar] [CrossRef]
- Dimitrov, D.S. Virus entry: Molecular mechanisms and biomedical applications. Nat. Rev. Microbiol. 2004, 2, 109–122. [Google Scholar] [CrossRef]
- Kim, T.-H.; Lee, S.-W. Aptamers for anti-viral therapeutics and diagnostics. Int. J. Mol. Sci. 2021, 22, 4168. [Google Scholar] [CrossRef] [PubMed]
- De Stefano, J.J.; Nair, G.R. Novel aptamer inhibitors of human immunodeficiency virus reverse transcriptase. Oligonucleotides 2008, 18, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Bellecave, P.; Cazenave, C.; Rumi, J.; Staedel, C.; Cosnefroy, O.; Andreola, M.-L.; Ventura, M.; Tarrago-Litvak, L.; Astier-Gin, T. Inhibition of hepatitis C virus (HCV) RNA polymerase by DNA aptamers: Mechanism of inhibition of in vitro RNA synthesis and effect on HCV-infected cells. Antimicrob. Agents Chemother. 2008, 52, 2097–2110. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J.P.; Kent, K.; Bird, J.; Fishback, J.; Froehler, B. Modified deoxyoligonucleotides stable to exonuclease degradation in serum. Nucleic Acids Res. 1991, 19, 747–750. [Google Scholar] [CrossRef] [Green Version]
- Wesselhoeft, R.A.; Kowalski, P.S.; Anderson, D.G. Engineering circular RNA for potent and stable translation in eukaryotic cells. Nat. Commun. 2018, 9, 2629. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Kim, M.Y.; Lee, J.H.; You, J.C.; Jeong, S. Selection and stabilization of the RNA aptamers against the human immunodeficiency virus type-1 nucleocapsid protein. Biochem. Biophys. Res. Commun. 2002, 291, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Kuai, H.; Zhao, Z.; Mo, L.; Liu, H.; Hu, X.; Fu, T.; Zhang, X.; Tan, W. Circular bivalent aptamers enable in vivo stability and recognition. J. Am. Chem. Soc. 2017, 139, 9128–9131. [Google Scholar] [CrossRef]
- Ulrich, H.; Trujillo, C.A.; Nery, A.A.; Alves, J.M.; Majumder, P.; Resende, R.R.; Martins, A.H. DNA and RNA aptamers: From tools for basic research towards therapeutic applications. Comb. Chem. High Throughput Screen 2006, 9, 619–632. [Google Scholar] [CrossRef]
- Ohashi, S.; Hashiya, F.; Abe, H. Variety of nucleotide polymerase mutants aiming to synthesize modified RNA. ChemBioChem 2021, 22, 1–10. [Google Scholar] [CrossRef]
- Volk, D.E.; Lokesh, G.L.R. Development of phosphorothioate DNA and DNA thioaptamers. Biomedicines 2017, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Eulberg, D.; Klussmann, S. Spiegelmers: Biostable Aptamers. ChemBioChem 2003, 4, 979–983. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, H.; Kasahara, Y.; Kuwahara, M.; Obika, S. DNA polymerase variants with high processivity and accuracy for encoding and decoding locked nucleic acid sequences. J. Am. Chem. Soc. 2020, 142, 21530–21537. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.I.; Holliger, P. Selecting fully-modified XNA aptamers using synthetic genetics. Curr. Protoc. Chem. Biol. 2018, 10, e44. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhang, K. PEGylation of therapeutic oligonucletides: From linear to highly branched PEG architectures. Nano Res. 2018, 11, 5519–5534. [Google Scholar] [CrossRef]
- Dheyab, M.A.; Khaniabadi, P.M.; Aziz, A.A.; Jameel, M.S.; Mehrdel, B.; Oglat, A.A.; Khaleel, H.A. Focused role of nanoparticles against COVID-19: Diagnosis and treatment. Photodiagn. Photodyn. Ther. 2021, 34, 102287. [Google Scholar] [CrossRef]
- Balkourani, G.; Brouzgou, A.; Archonti, M.; Papandrianos, N.; Song, S.; Tsiakaras, P. Emerging materials for the electrochemical detection of COVID-19. J. Electroanal. Chem. 2021, 115289. [Google Scholar] [CrossRef]
- Li, J.; You, J.; Dai, Y.; Shi, M.; Han, C.; Xu, K. Gadolinium oxide nanoparticles and aptamer-functionalized silver nanoclusters-based multimodal molecular imaging nanoprobe for optical/magnetic resonance cancer cell imaging. Anal. Chem. 2014, 86, 11306–11311. [Google Scholar] [CrossRef]
- Koudrina, A.; McConnell, E.M.; Zurakowski, J.A.; Cron, G.O.; Chen, S.; Tsai, E.C.; DeRosa, M.C. Exploring the unique contrast properties of aptamer-gadolinium conjugates in magnetic resonance imaging for targeted imaging of thrombi. ACS Appl. Mater. Interfaces 2021, 13, 9412–9424. [Google Scholar] [CrossRef] [PubMed]
- Moghadam, S.M.M.; Alibolandi, M.; Babaei, M.; Mosafer, J.; Saljooghi, A.S.; Ramezani, M. Fabrication of deferasirox-decorated aptamer-targeted superparamagnetic iron oxide nanoparticles (SPION) as a therapeutic and magnetic resonance imaging agent in cancer therapy. J. Biol. Inorg. Chem. 2021, 26, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.M.; Aartsma-Rus, A.; Alves, S.; Borgos, S.E.; Buijsen, R.A.M.; Collin, R.W.J.; Covello, G.; Denti, M.A.; Desviat, L.R.; Echevarría, L.; et al. Delivery of oligonucleotide-based therapeutics: Challenges and opportunities. EMBO Mol. Med. 2021, 13, e13243. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Xiang, J. Aptamer-functionalized nanoparticles in targeted delivery and cancer therapy. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Jang, S.; Jang, S.; Yang, J.; Seo, S.W.; Jung, G.Y. RNA-based dynamic genetic controllers: Development strategies and applications. Curr. Opin. Biotechnol. 2018, 53, 1–11. [Google Scholar] [CrossRef]
- Ketzer, P.; Kaufmann, J.K.; Engelhardt, S.; Bossow, S.; von Kalle, C.; Hartig, J.S.; Ungerechts, G.; Nettelbeck, D.M. Artificial riboswitches for gene expression and replication control of DNA and RNA viruses. Proc. Natl. Acad. Sci. USA 2014, 111, E554–E562. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krüger, A.; de Jesus Santos, A.P.; de Sá, V.; Ulrich, H.; Wrenger, C. Aptamer Applications in Emerging Viral Diseases. Pharmaceuticals 2021, 14, 622. https://doi.org/10.3390/ph14070622
Krüger A, de Jesus Santos AP, de Sá V, Ulrich H, Wrenger C. Aptamer Applications in Emerging Viral Diseases. Pharmaceuticals. 2021; 14(7):622. https://doi.org/10.3390/ph14070622
Chicago/Turabian StyleKrüger, Arne, Ana Paula de Jesus Santos, Vanessa de Sá, Henning Ulrich, and Carsten Wrenger. 2021. "Aptamer Applications in Emerging Viral Diseases" Pharmaceuticals 14, no. 7: 622. https://doi.org/10.3390/ph14070622