
Discovery of New Chemical Tools against Leishmania amazonensis via the MMV Pathogen Box


Abstract
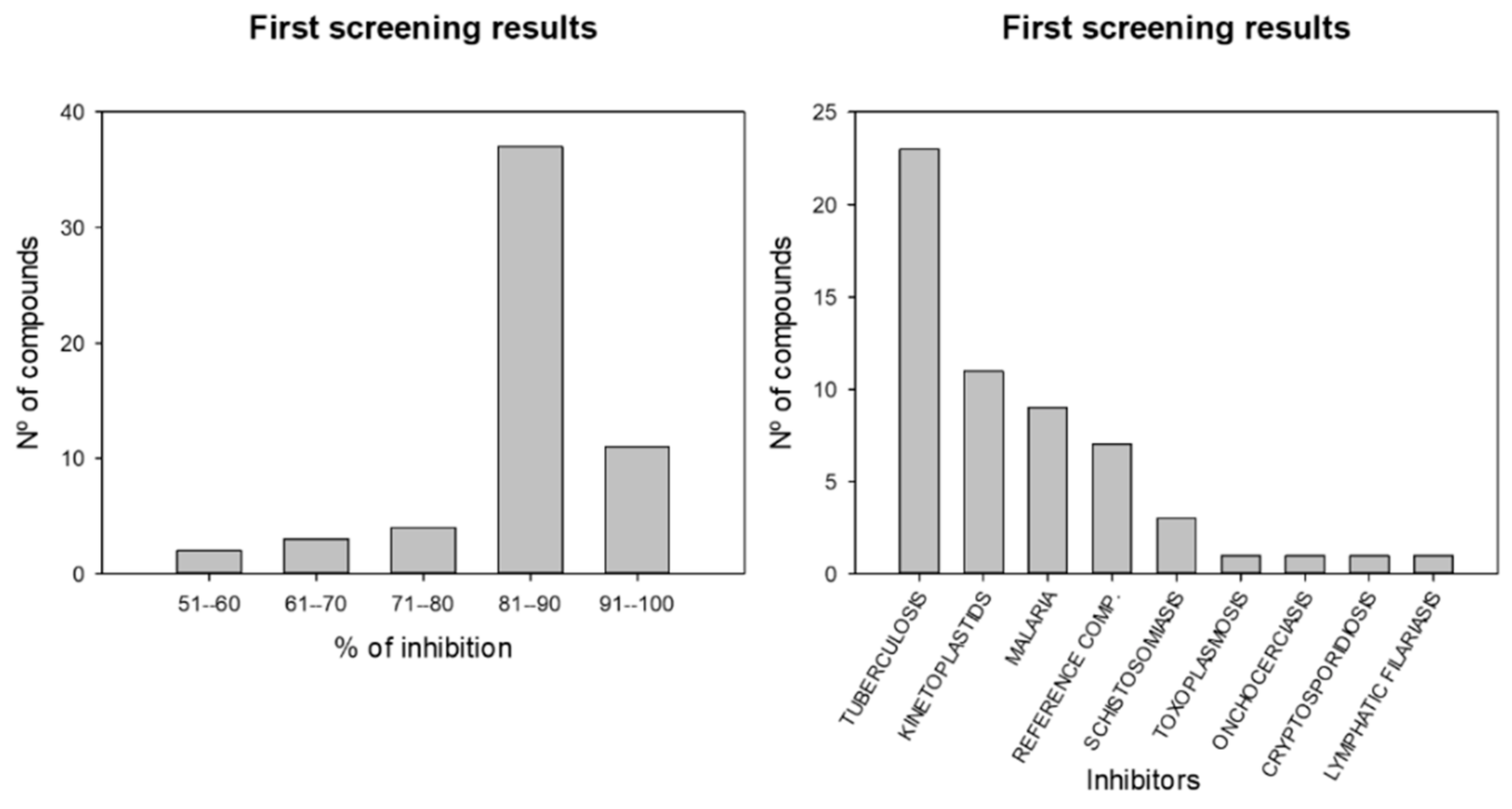
:1. Introduction
2. Results
2.1. Characterization of the Compounds
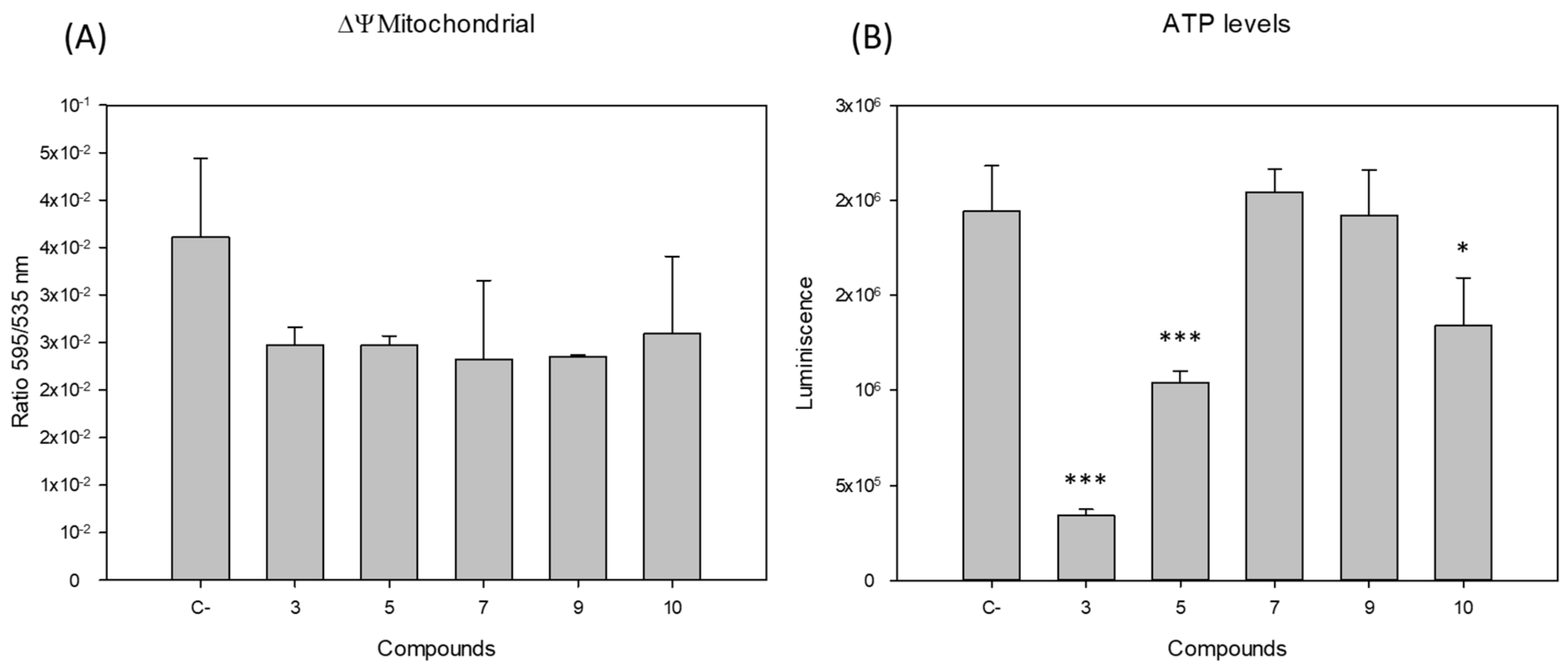
2.2. Mitochondrial Function
2.3. Oxidative Stress
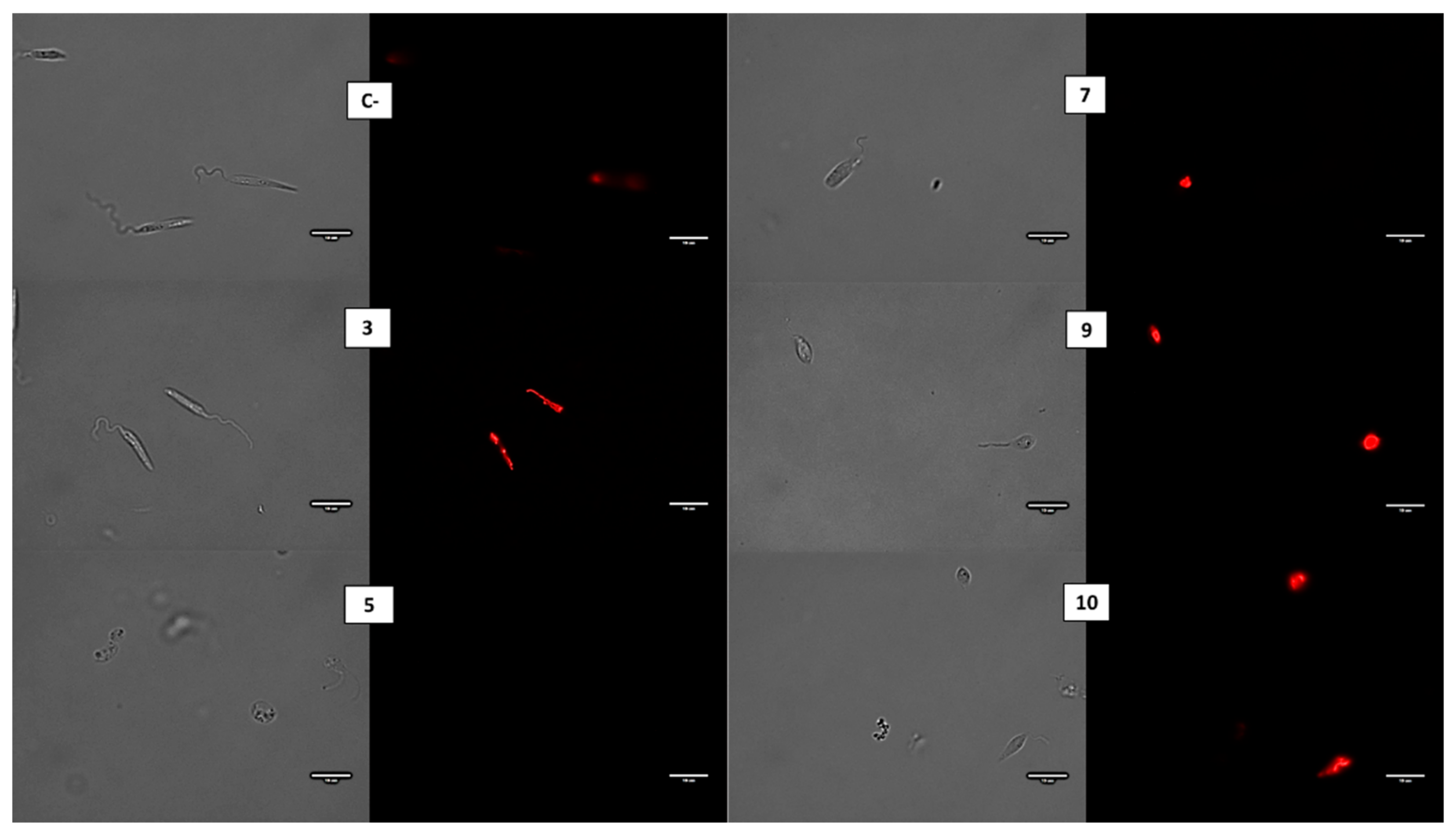
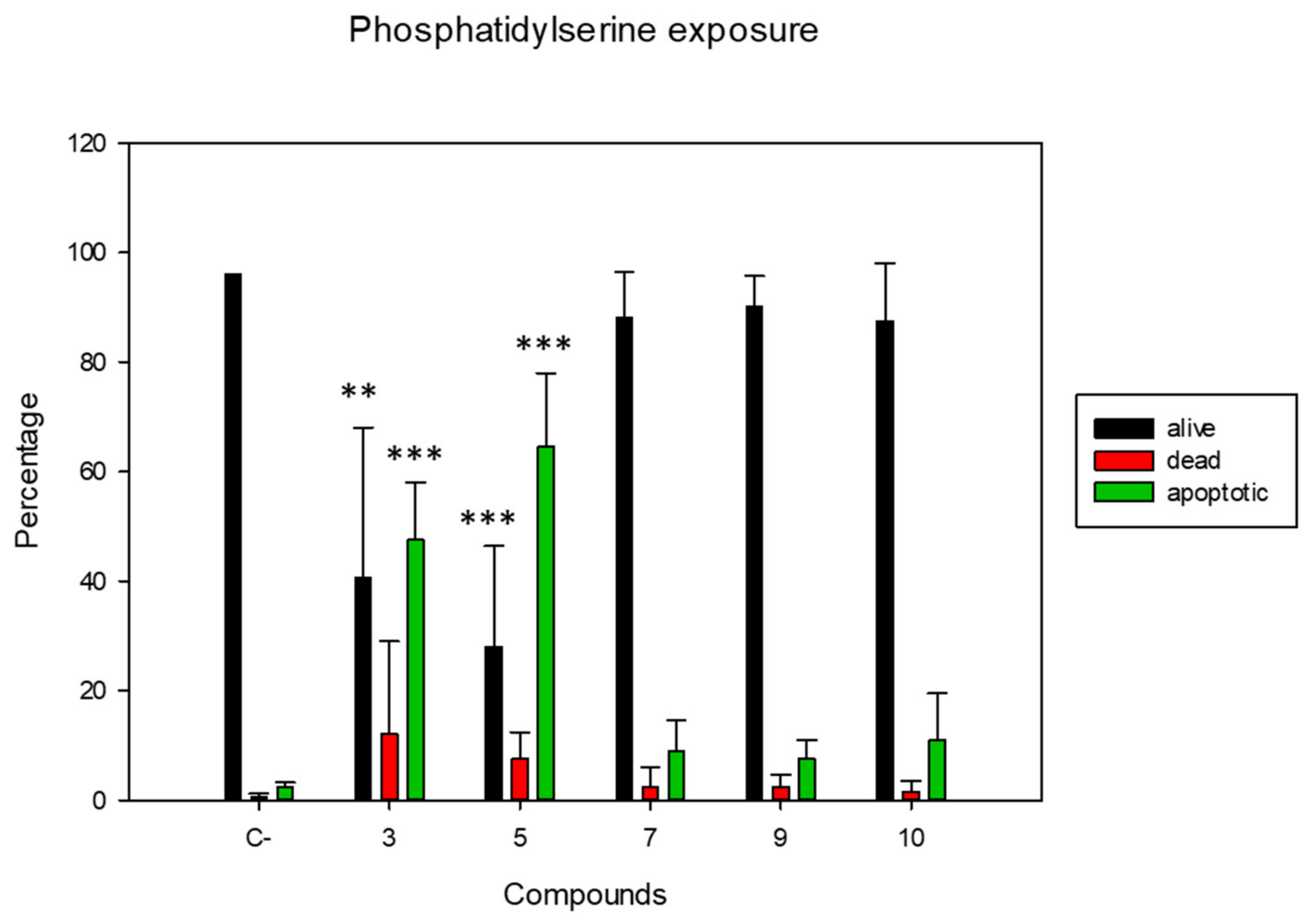
2.4. Membrane Alterations
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Strains
4.3. Leishmanicidal Activity
4.4. Cytotoxic Activity
4.5. ATP Levels
4.6. Mitochondrial Membrane Potential Disruption
4.7. Reactive Oxygen Species (ROS) Detection
4.8. Phosphatidylserine Externalization
4.9. Plasmatic Membrane Permeability
4.10. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Available online: http://www.who.int/leishmaniasis/en/ (accessed on 15 May 2021).
- Lee, S.-M.; Kim, M.-S.; Hayat, F.; Shin, D. Recent Advances in the Discovery of Novel Antiprotozoal Agents. Molecules 2019, 24, 3886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, H.W.; Berman, J.D.; Davies, C.R.; Saravia, N.G. Advances in leishmaniasis. Lancet 2005, 366, 1561–1577. [Google Scholar] [CrossRef]
- Camacho, M.; Phillipson, J.; Croft, S.; Solis, P.; Marshall, S.; Ghazanfar, S. Screening of plant extracts for antiprotozoal and cytotoxic activities. J. Ethnopharmacol. 2003, 89, 185–191. [Google Scholar] [CrossRef]
- Sen, R.; Chatterjee, M. Plant derived therapeutics for the treatment of Leishmaniasis. Phytomedicine 2011, 18, 1056–1069. [Google Scholar] [CrossRef]
- Ponte-Sucre, A.; Gamarro, F.; Dujardin, J.-C.; Barrett, M.P.; López-Vélez, R.; García-Hernández, R.; Pountain, A.; Mwenechanya, R.; Papadopoulou, B. Drug resistance and treatment failure in leishmaniasis: A 21st century challenge. PLoS Neglected Trop. Dis. 2017, 11, e0006052. [Google Scholar] [CrossRef]
- Das, M.; Mukherjee, S.B.; Shaha, C. Hydrogen peroxide induces apoptosis-like death in Leishmania donovani promastigotes. J. Cell Sci. 2001, 114, 2461–2469. [Google Scholar] [CrossRef]
- Roy, A.; Ganguly, A.; BoseDasgupta, S.; Das, B.B.; Pal, C.; Jaisankar, P.; Majumder, H.K. Mitochondria-Dependent Reactive Oxygen Species-Mediated Programmed Cell Death Induced by 3,3′-Diindolylmethane through Inhibition of F0F1-ATP Synthase in Unicellular Protozoan ParasiteLeishmania donovani. Mol. Pharmacol. 2008, 74, 1292–1307. [Google Scholar] [CrossRef]
- Reece, S.E.; Pollitt, L.C.; Colegrave, N.; Gardner, A. The Meaning of Death: Evolution and Ecology of Apoptosis in Protozoan Parasites. PLoS Pathog. 2011, 7, e1002320. [Google Scholar] [CrossRef] [Green Version]
- Sifaoui, I.; Reyes-Batlle, M.; López-Arencibia, A.; Chiboub, O.; Bethencourt-Estrella, C.J.; Nicolás-Hernández, D.S.; Expósito, R.L.R.; Rizo-Liendo, A.; Piñero, J.E.; Lorenzo-Morales, J. Screening of the pathogen box for the identification of anti-Acanthamoeba agents. Exp. Parasitol. 2019, 201, 90–92. [Google Scholar] [CrossRef]
- Rice, C.A.; Lares-jiménez, L.F.; Lares-villa, F.; Kyle, D.E. In Vitro Screening of the Open-Source Medicines for Malaria. Antimicrob. Agents Chemother. 2020, 64, 1–6. [Google Scholar]
- Dans, M.G.; Weiss, G.E.; Wilson, D.; Sleebs, B.E.; Crabb, B.S.; de Koning-Ward, T.F.; Gilson, P.R. Screening the Medicines for Malaria Venture Pathogen Box for invasion and egress inhibitors of the blood stage of Plasmodium falciparum reveals several inhibitory compounds. Int. J. Parasitol. 2020, 50, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Spalenka, J.; Escotte-Binet, S.; Bakiri, A.; Hubert, J.; Renault, J.-H.; Velard, F.; Duchateau, S.; Aubert, D.; Huguenin, A.; Villena, I. Discovery of New Inhibitors of Toxoplasma gondii via the Pathogen Box. Antimicrob. Agents Chemother. 2018, 62, e01640-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machicado, C.; Soto, M.P.; Timoteo, O.; Vaisberg, A.; Pajuelo, M.; Ortiz, P.; Marcos, L.A. Screening the Pathogen Box for Identification of New Chemical Agents with Anti- Fasciola hepatica Activity. Antimicrob. Agents Chemother. 2019, 63, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, L.; de Paula, J.C.; Baréa, P.; Sarragiotto, M.H.; Ueda-Nakamura, T.; Alonso, A.; Fernandes, N.D.S.; Lancheros, C.A.C.; Volpato, H.; Lazarin-Bidóia, D.; et al. Membrane dynamics in Leishmania amazonensis and antileishmanial activities of β-carboline derivatives. Biochim. Biophys. Acta (BBA)-Biomembr. 2021, 1863, 183473. [Google Scholar] [CrossRef] [PubMed]
- Trinconi, C.; Reimão, J.; Coelho, A.C.; Uliana, S.R.B. Efficacy of tamoxifen and miltefosine combined therapy for cutaneous leishmaniasis in the murine model of infection withLeishmania amazonensis. J. Antimicrob. Chemother. 2016, 71, 1314–1322. [Google Scholar] [CrossRef] [Green Version]
- Proto, W.R.; Coombs, G.H.; Mottram, J. Cell death in parasitic protozoa: Regulated or incidental? Nat. Rev. Genet. 2012, 11, 58–66. [Google Scholar] [CrossRef]
- Menna-Barreto, R.F.S. Cell death pathways in pathogenic trypanosomatids: Lessons of (over)kill. Cell Death Dis. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Piacenza, L.; Irigoín, F.; Alvarez, M.N.; Peluffo, G.; Taylor, M.; Kelly, J.M.; Wilkinson, S.R.; Radi, R. Mitochondrial superoxide radicals mediate programmed cell death in Trypanosoma cruzi: Cytoprotective action of mitochondrial iron superoxide dismutase overexpression. Biochem. J. 2007, 403, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Yang, Z.; Xu, Y.; Chen, Y.; Yu, Q. Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol. Cancer 2015, 14, 1–14. [Google Scholar] [CrossRef] [Green Version]
- López-Arencibia, A.; Nicolás-Hernández, D.S.; Bethencourt-Estrella, C.J.; Sifaoui, I.; Reyes-Batlle, M.; Rodríguez-Expósito, R.L.; Rizo-Liendo, A.; Lorenzo-Morales, J.; Bazzocchi, I.L.; Piñero, J.E.; et al. Withanolides from Withania aristata as Antikinetoplastid Agents through Induction of Programmed Cell Death. Pathogens 2019, 8, 172. [Google Scholar] [CrossRef] [Green Version]
- López-Arencibia, A.; Reyes-Batlle, M.; Freijo, M.B.; Sifaoui, I.; Bethencourt-Estrella, C.J.; Rizo-Liendo, A.; Chiboub, O.; McNaughton-Smith, G.; Lorenzo-Morales, J.; Abad-Grillo, T.; et al. In vitro activity of 1H-phenalen-1-one derivatives against Leishmania spp. and evidence of programmed cell death. Parasit. Vectors 2019, 12, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freijo, M.B.; Arencibia, A.L.; Piñero, J.; McNaughton-Smith, G.; Abad-Grillo, T. Design, synthesis and evaluation of amino-substituted 1H-phenalen-1-ones as anti-leishmanial agents. Eur. J. Med. Chem. 2018, 143, 1312–1324. [Google Scholar] [CrossRef]
- Diaz-Marrero, A.R.; López-Arencibia, A.; Bethencout-Estrella, C.J.; Cen-Pacheco, F.; Sifaoui, I.; Creus, A.H.; Duque-Ramírez, M.C.; Souto, M.L.; Daranas, A.H.; Lorenzo-Morales, J.; et al. Antiprotozoal activities of marine polyether triterpenoids. Bioorganic Chem. 2019, 92, 103276. [Google Scholar] [CrossRef] [PubMed]
- Actor, P.; Chow, A.W.; Dutko, F.J.; McKinlay, M.A. Chemotherapeutics. Ullmann Encycl. Ind. Chem. 2000. [Google Scholar] [CrossRef]
- Tadele, M.; Abay, S.M.; Makonnen, E.; Hailu, A. Identification of Leishmania donovani inhibitors from pathogen box compounds of Medicine for Malaria Venture. bioRxiv 2019, 1307–1317. [Google Scholar] [CrossRef] [Green Version]
- Duffy, S.; Sykes, M.L.; Jones, A.J.; Shelper, T.B.; Simpson, M.; Lang, R.; Poulsen, S.-A.; Sleebs, B.E.; Avery, V.M. Screening the Medicines for Malaria Venture Pathogen Box across Multiple Pathogens Reclassifies Starting Points for Open-Source Drug Discovery. Antimicrob. Agents Chemother. 2017, 61, e00379-17. [Google Scholar] [CrossRef] [Green Version]
- Musonda, C.C.; Whitlock, G.A.; Witty, M.J.; Brun, R.; Kaiser, M. Synthesis and evaluation of 2-pyridyl pyrimidines with in vitro antiplasmodial and antileishmanial activity. Bioorganic Med. Chem. Lett. 2009, 19, 401–405. [Google Scholar] [CrossRef]
- Canever, M.F.; Miletti, L.C. Screening and Identification of Pathogen Box® Compounds with anti-Trypanosoma evansi Activity. Acta Trop. 2020, 206, 105421. [Google Scholar] [CrossRef]
- Borbély, G.; Szabadkai, I.; Horváth, Z.; Markó, P.; Varga, Z.; Breza, N.; Baska, F.; Vántus, T.; Huszár, M.; Geiszt, M.; et al. Small-Molecule Inhibitors of NADPH Oxidase 4. J. Med. Chem. 2010, 53, 6758–6762. [Google Scholar] [CrossRef]
- McRobert, L.; Taylor, C.J.; Deng, W.; Fivelman, Q.L.; Cummings, R.M.; Polley, S.D.; Billker, O.; Baker, D.A. Gametogenesis in Malaria Parasites Is Mediated by the cGMP-Dependent Protein Kinase. PLoS Biol. 2008, 6, e139. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Chong, C.R.; Shi, L.; Yoshimoto, T.; Sullivan, D.J.; Liu, J.O. Inhibitors of Plasmodium falciparum methionine aminopeptidase 1b possess antimalarial activity. Proc. Natl. Acad. Sci. USA 2006, 103, 14548–14553. [Google Scholar] [CrossRef]
- Rocco-Machado, N.; Cosentino-Gomes, D.; Nascimento, M.; Paes-Vieira, L.; Khan, Y.; Mittra, B.; Andrews, N.; Meyer-Fernandes, J. Leishmania amazonensis ferric iron reductase (LFR1) is a bifunctional enzyme: Unveiling a NADPH oxidase activity. Free. Radic. Biol. Med. 2019, 143, 341–353. [Google Scholar] [CrossRef]
- Jaeger, T.; Flohé, L. The thiol-based redox networks of pathogens: Unexploited targets in the search for new drugs. BioFactors 2006, 27, 109–120. [Google Scholar] [CrossRef]
- Fiorillo, A.; Colotti, G.; Boffi, A.; Baiocco, P.; Ilari, A. The Crystal Structures of the Tryparedoxin-Tryparedoxin Peroxidase Couple Unveil the Structural Determinants of Leishmania Detoxification Pathway. PLoS Neglected Trop. Dis. 2012, 6, e1781. [Google Scholar] [CrossRef] [Green Version]
- Auld, U.S.; Veith, H.; Cali, J.J. Bioluminescent Assays for Cytochrome P450 Enzymes. Adv. Struct. Saf. Stud. 2013, 987, 1–9. [Google Scholar] [CrossRef]
- Blair, I.A. Analysis of endogenous glutathione-adducts and their metabolites. Biomed. Chromatogr. 2009, 24, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Mikus, J.; Steverding, D. A simple colorimetric method to screen drug cytotoxicity against Leishmania using the dye Alamar Blue®. Parasitol. Int. 2000, 48, 265–269. [Google Scholar] [CrossRef]
- Jain, S.K.; Sahu, R.; Walker, L.A.; Tekwani, B.L. A Parasite Rescue and Transformation Assay for Antileishmanial Screening Against Intracellular Leishmania donovani Amastigotes in THP1 Human Acute Monocytic Leukemia Cell Line. J. Vis. Exp. 2012, e4054. [Google Scholar] [CrossRef] [Green Version]
- López-Arencibia, A.; Martín-Navarro, C.; Sifaoui, I.; Reyes-Batlle, M.; Wagner, C.; Lorenzo-Morales, J.; Maciver, S.K.; Piñero, J.E. Perifosine Mechanisms of Action in Leishmania Species. Antimicrob. Agents Chemother. 2017, 61, 61. [Google Scholar] [CrossRef] [Green Version]
- Marinho, F.A.; Gonçalves, K.C.S.; Oliveira, S.S.C.; Gonçalves, D.D.S.; Matteoli, F.P.; Seabra, S.H.; Oliveira, A.C.; Bellio, M.; Oliveira, S.S.; Souto-Padrón, T.; et al. The Calpain Inhibitor MDL28170 Induces the Expression of Apoptotic Markers in Leishmania amazonensis Promastigotes. PLoS ONE 2014, 9, e87659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeouk, I.; Sifaoui, I.; López-Arencibia, A.; Reyes-Batlle, M.; Bethencourt-Estrella, C.J.; Bazzocchi, I.L.; Bekhti, K.; Lorenzo-Morales, J.; Jiménez, I.A.; Piñero, J.E. Sesquiterpenoids and flavonoids from Inula viscosa induce programmed cell death in kinetoplastids. Biomed. Pharmacother. 2020, 130, 110518. [Google Scholar] [CrossRef]
- López-Arencibia, A.; García-Velázquez, D.; Martín-Navarro, C.M.; Sifaoui, I.; Reyes-Batlle, M.; Lorenzo-Morales, J.; Gutiérrez-Ravelo, Á.; Piñero, J. In VitroActivities of Hexaazatrinaphthylenes against Leishmania spp. Antimicrob. Agents Chemother. 2015, 59, 2867–2874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesquita, J.T.; da Costa-Silva, T.A.; Borborema, S.E.T.; Tempone, A.G. Activity of imidazole compounds on Leishmania (L.) infantum chagasi: Reactive oxygen species induced by econazole. Mol. Cell. Biochem. 2014, 389, 293–300. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Leishmania amazonensis Promastigotes IC50 (µM) | Macrophages CC50 (µM) | Selectivity Index |
|---|---|---|---|
| 1 | 6.43 ± 1.06 | >10 | >1.56 |
| 2 | 4.03 ± 0.73 | >10 | >2.48 |
| 3 | 1.33 ± 0.12 | 8.02 ± 0.93 | 5.66 |
| 4 | 1.44 ± 0.24 | 4.30 ± 0.16 | 2.94 |
| 5 | 0.59 ± 0.08 | >10 | >17.06 |
| 6 | 3.93 ± 0.64 | >10 | >2.54 |
| 7 | 1.84 ± 0.14 | >10 | >5.44 |
| 8 | 2.62 ± 0.40 | 8.01 ± 1.13 | 3.06 |
| 9 | 0.77 ± 0.01 | 7.22 ± 2.36 | 9.35 |
| 10 | 1.85 ± 0.39 | >10 | >5.40 |
| Miltefosine | 6.48 ± 0.24 | 72.19 ± 3.06 | 11.14 |
| Comp. 1 | Comp. 2 |
 |  |
| Comp. 3 | Comp. 4 |
 |  |
| Comp. 5 | Comp. 6 |
 |  |
| Comp. 7 | Comp. 8 |
 |  |
| Comp. 9 | Comp. 10 |
 |  |
| Compound | Leishmania amazonensis Intracellular Amastigote IC50 (µM) | Selectivity Index |
|---|---|---|
| 1 | >10 | nd |
| 2 | >10 | nd |
| 3 | 4.78 ± 1.20 | 1.68 |
| 4 | >10 | nd |
| 5 | 1.25 ± 0.28 | >8.00 |
| 6 | 8.52 ± 0.67 | >1.17 |
| 7 | 2.50 ± 0.27 | >4.00 |
| 8 | 6.85 ± 0.75 | 1.17 |
| 9 | 6.90 ± 1.79 | 1.05 |
| 10 | 1.33 ± 0.36 | >7.52 |
| Miltefosine | 3.12 ± 0.30 | 23.14 |
| MMV Identifier a | CHEMBL Identifier and Name | ADME a | ||
|---|---|---|---|---|
| CYP2C9 IC50 (μM) | CYP2D6 IC50 (μM) | Glutathione Reactivity a | ||
| MMV676388 (Compound 3) | CHEMBL3637869 5-(Benzylsulfonyl)-1-(3-methoxyphenyl)-1H-tetrazole | >20 | 15 | Medium |
| MMV690103 (Compound 5) | CHEMBL3637900 2-N-Methyl-2-N-[(1R)-1-(5,6,7,8-tetrahydronaphthalen-2-yl)ethyl]pyrimido [4,5-d]pyrimidine-2,5,7-triamine | >20 | >20 | Medium |
| MMV022029 (Compound 7) | CHEMBL545853 N-Benzyl-4-[3-[[(1-methylpiperidin-4-yl)amino]methyl]phenyl]benzenesulfonamide | >20 | >20 | No GSH adduct |
| MMV022478 (Compound 9) | CHEMBL534797 3-(3-Chlorophenyl)-N-(4-piperazin-1-ylphenyl)pyrazolo [1,5-a]pyrimidine-5-carboxamide | >20 | >20 | No GSH adduct |
| MMV021013 (Compound 10) | CHEMBL530275 N-Cyclohexyl-6-cyclopropyl-2-pyridin-2-ylpyrimidin-4-amine | >20 | >20 | No GSH adduct |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Arencibia, A.; Sifaoui, I.; Reyes-Batlle, M.; Bethencourt-Estrella, C.J.; San Nicolás-Hernández, D.; Lorenzo-Morales, J.; Piñero, J.E. Discovery of New Chemical Tools against Leishmania amazonensis via the MMV Pathogen Box. Pharmaceuticals 2021, 14, 1219. https://doi.org/10.3390/ph14121219
López-Arencibia A, Sifaoui I, Reyes-Batlle M, Bethencourt-Estrella CJ, San Nicolás-Hernández D, Lorenzo-Morales J, Piñero JE. Discovery of New Chemical Tools against Leishmania amazonensis via the MMV Pathogen Box. Pharmaceuticals. 2021; 14(12):1219. https://doi.org/10.3390/ph14121219
Chicago/Turabian StyleLópez-Arencibia, Atteneri, Ines Sifaoui, María Reyes-Batlle, Carlos J. Bethencourt-Estrella, Desirée San Nicolás-Hernández, Jacob Lorenzo-Morales, and José E. Piñero. 2021. "Discovery of New Chemical Tools against Leishmania amazonensis via the MMV Pathogen Box" Pharmaceuticals 14, no. 12: 1219. https://doi.org/10.3390/ph14121219