

The Explanation of Photopic Luminous Efficiency Curve by Using Both of the Cones’ Optical Fiber Coupling Effects and the Absorption of L Cones

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Beam Propagation Method (BPM)
2.2. Monitoring and Analysis of the Simulation Results
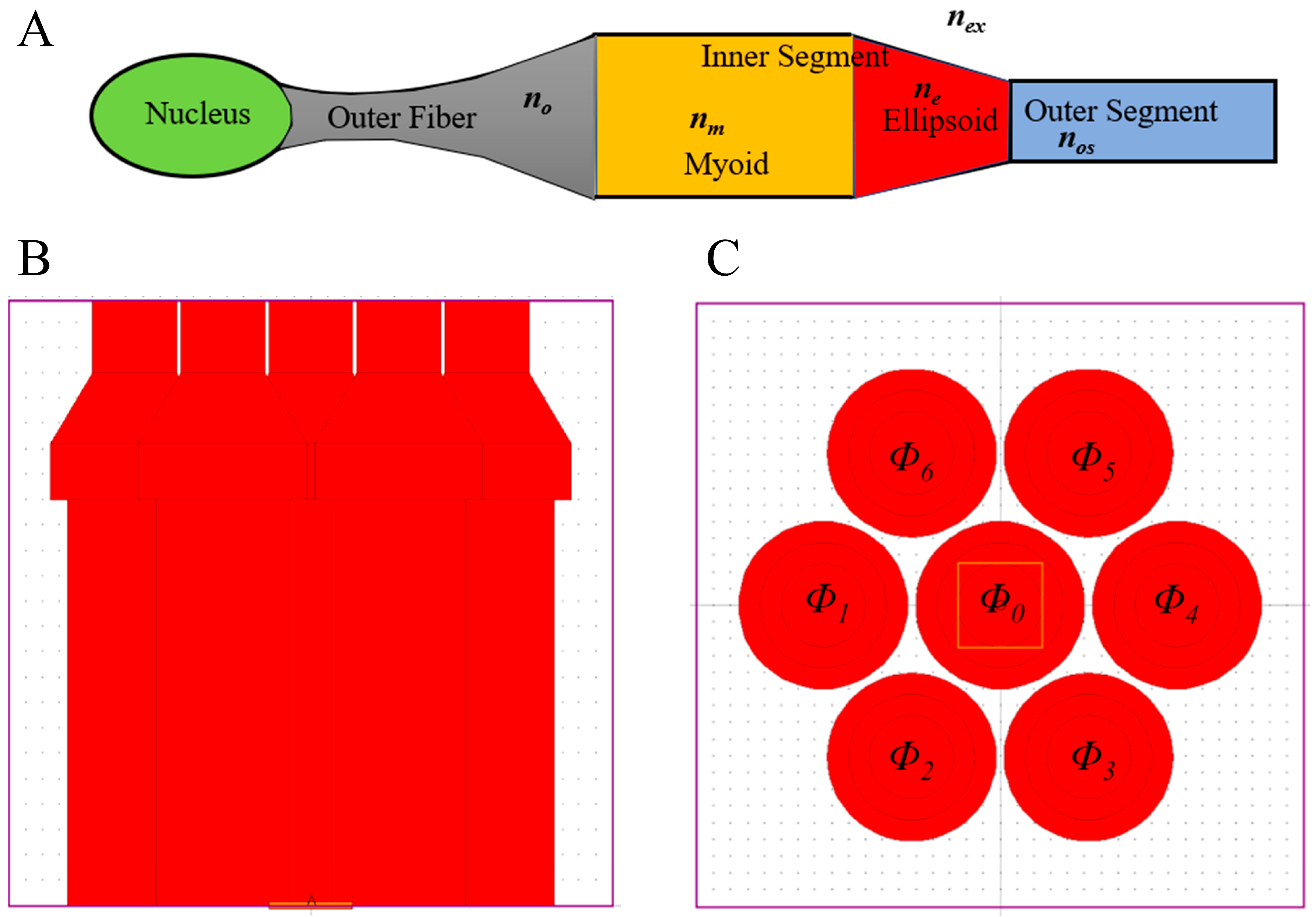
2.3. Parameters of the Models
2.4. Splitting Ratio
3. Results
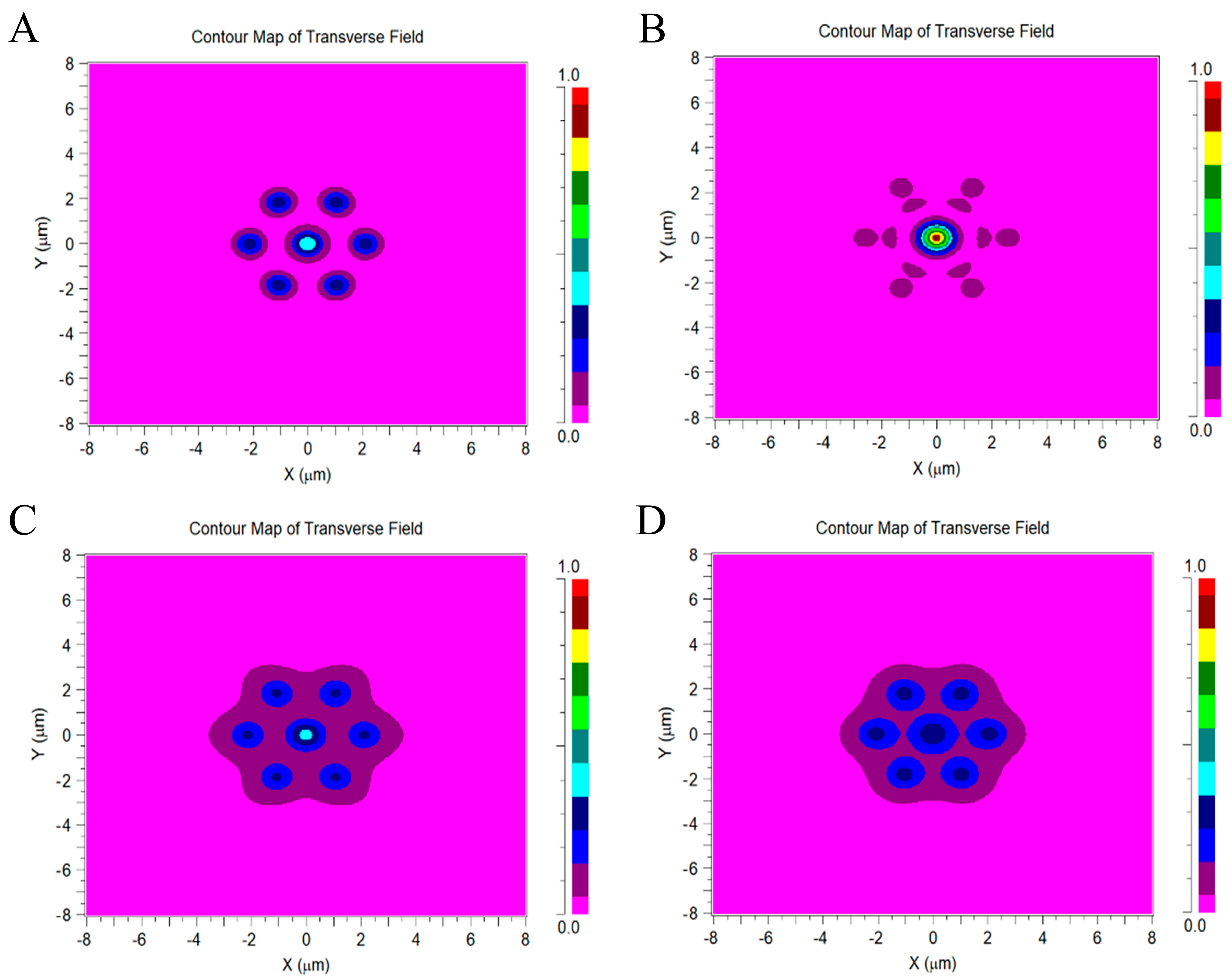
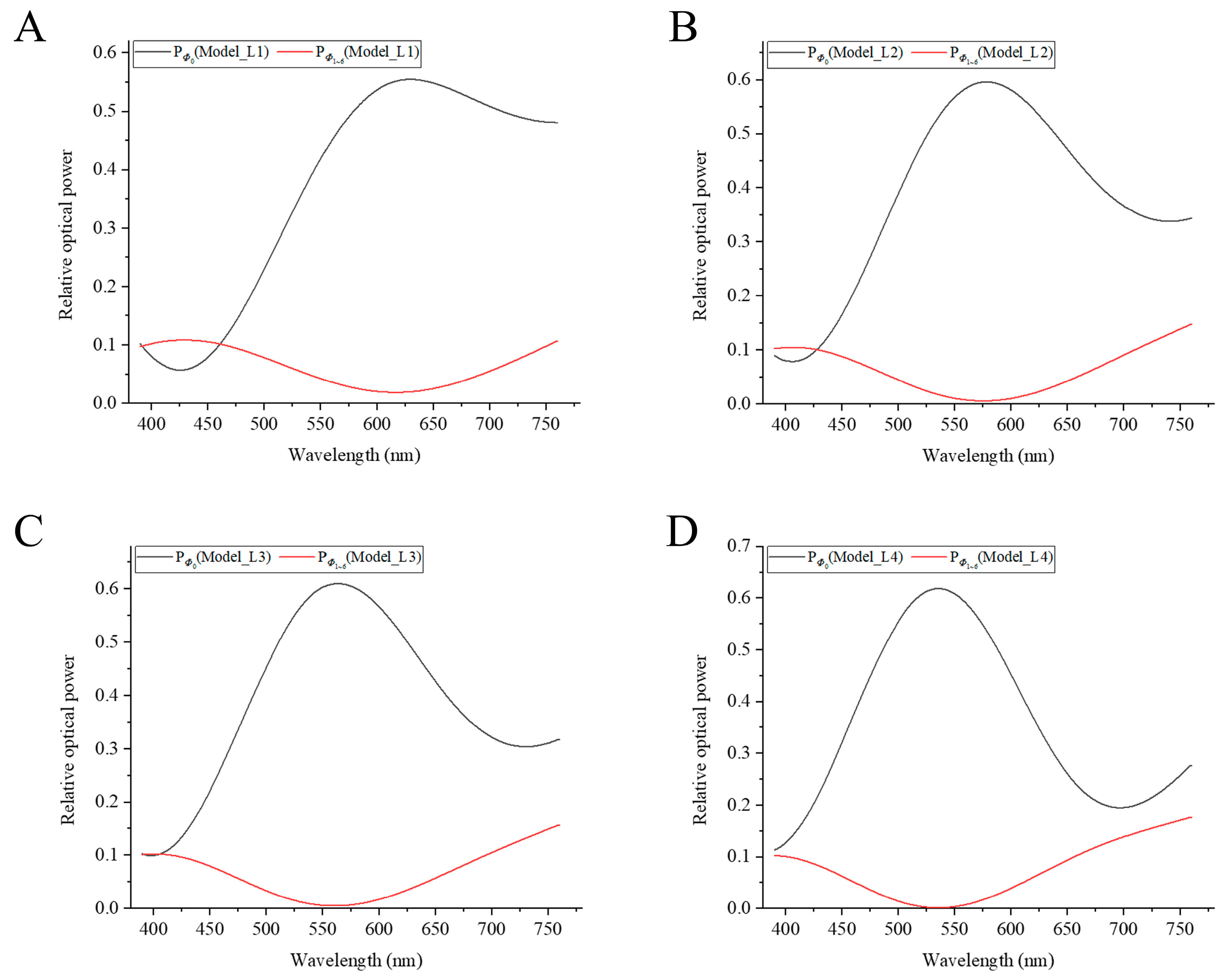
3.1. Relative Optical Power at the Output of the Outer Segment
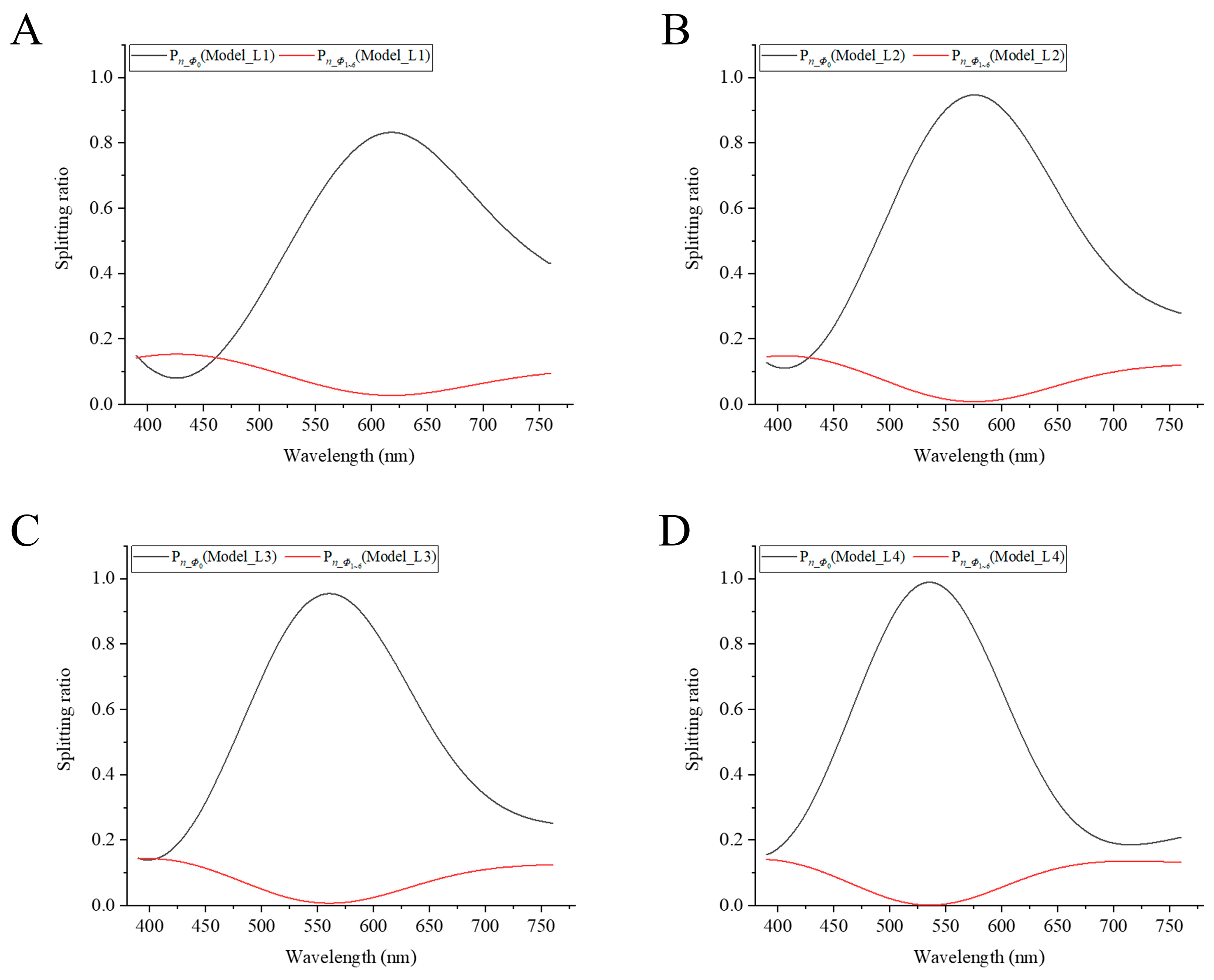
3.2. Relationship between Splitting Ratio and Wavelength
3.3. Photopic Luminous Efficiency
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Enoch, J.M. Visualization of wave-guide modes in retinal receptors. Am. J. Ophthalmol. 1961, 51, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Harte, R.A. Receptor elements of the human retina as semiconductors. J. Opt. Soc. Am. 1961, 51, 1275–1278. [Google Scholar] [CrossRef] [PubMed]
- Liang, A. Photoreceptors of Animals Are Quantum-Well Detectors. In Proceedings of the Japan Society of Applied Physics 59th Autumn Meeting, Hiroshima, Japan, 25–28 September 1998. [Google Scholar]
- Liang, A.; Hu, L. Novel optical waveguide theory and novel electrical circuit theory of photoreceptors in the human retina. Invited talk. In Proceedings of the Progress in Electromagnetics Research Symposium, Stockholm, Sweden, 12–15 August 2013. [Google Scholar]
- Liang, A.; Li, W.; Luo, X.; Gu, J.; Liu, Y.; Hu, L. Strong optical coupling between neighboring cones on human retina. In Proceedings of the OSA Fall Vision Meeting, Philadelphia, PA, USA, 22–24 October 2014. [Google Scholar]
- Liang, A.; Hu, L.; Liang, Z. Mode-Field-Diameter and the Coupling Loss between Inner and Outer Segment of Photoreceptors. Opt. Photonics J. 2015, 5, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Liang, A.; Meng, Z. The explanation of color vision based on fiber coupler theory of foveal cones. Chin. Sci. Bull. 2016, 61, 2952–2959. (In Chinese) [Google Scholar] [CrossRef] [Green Version]
- Liang, A.; Wang, X.; Meng, Z. Existence of strong optical coupling among seven identical human foveal cones. In Proceedings of the OSA Fall Vision Meeting, Washington, DC, USA, 13–15 October 2017. [Google Scholar]
- Liang, A. Smart and advanced PIC on human retina and optical fibers in animal bodies. In Proceedings of the SPIE, San Francisco, CA, USA, 31 July 2018. [Google Scholar]
- Liang, A.H. Biological Optical AI and Bionic Optical AI. Plenary speech. In Proceedings of the 2020 5th International Conference on Robotics and Automation Engineering (ICRAE), Singapore, 20–22 November 2020; pp. 76–80. [Google Scholar]
- Ball, J.M.; Chen, S.; Li, W. Mitochondria in cone photoreceptors act as microlenses to enhance photon delivery and confer directional sensitivity to light. Sci. Adv. 2022, 8, eabn2070. [Google Scholar] [CrossRef]
- Xin, H.; Li, Y.; Liu, X.; Li, B. Escherichia coli-based biophotonic waveguides. Nano Lett. 2013, 13, 3408–3413. [Google Scholar] [CrossRef]
- Xin, H.; Li, Y.; Li, B. Bacteria-based branched structures for bionanophotonics. Laser Photonics Rev. 2015, 9, 554–563. [Google Scholar] [CrossRef]
- Pan, T.; Lu, D.; Xin, H.; Li, B. Biophotonic probes for bio-detection and imaging. Light Sci. Appl. 2021, 10, 124. [Google Scholar] [CrossRef]
- Lee, H.J.; Ha, J.H.; Kim, S.G.; Choi, H.K.; Kim, Z.H.; Han, Y.J.; Kim, J.I.; Oh, Y.; Fragoso, V.; Shin, K.; et al. Stem-piped light activates phytochrome B to trigger light responses in Arabidopsis thaliana roots. Sci. Signal. 2016, 9, ra106. [Google Scholar] [CrossRef]
- Xiaoling, L. Visual Neurophysiology; People’s Medical Publishing House: Beijing, China, 2018. [Google Scholar]
- Brainard, D.H. Color, Pattern, and the Retinal Cone Mosaic. Curr. Opin. Behav. Sci. 2019, 30, 41–47. [Google Scholar] [CrossRef]
- Stockman, A. Cone fundamentals and CIE standards. Curr. Opin. Behav. Sci. 2019, 30, 87–93. [Google Scholar] [CrossRef]
- Stockman, A.; Sharpe, L.T. The spectral sensitivities of the middle- and long-wavelength-sensitive cones derived from measurements in observers of known genotype. Vis. Res. 2000, 40, 1711–1737. [Google Scholar] [CrossRef] [Green Version]
- Stockman, A.; Sharpe, L.T.; Fach, C. The spectral sensitivity of the human short-wavelength sensitive cones derived from thresholds and color matches. Vis. Res. 1999, 39, 2901–2927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Yao, Q.; Jiang, L. Comparisons of Scotopic/Photopic Ratios Using 2- and 10-Degree Spectral Sensitivity Curves. Appl. Sci. 2019, 9, 4471. [Google Scholar] [CrossRef] [Green Version]
- Snyder, A.W.; Hamer, M. The light-capture area of a photoreceptor. Vis. Res. 1972, 12, 1749–1753. [Google Scholar] [CrossRef]
- Gorrand, J.-M.; Delori, F.C. A model for assessment of cone directionality. J. Mod. Optic. 2009, 44, 473–491. [Google Scholar] [CrossRef]
- Snyder, A.W.; Pask, C. The Stiles-Crawford effect—Explanation and consequences. Vis. Res. 1973, 13, 1115–1137. [Google Scholar] [CrossRef]
- Liu, Z.; Kocaoglu, O.P.; Turner, T.L.; Miller, D.T. Modal content of living human cone photoreceptors. Biomed. Opt. Express 2015, 6, 3378–3404. [Google Scholar] [CrossRef] [Green Version]
- Barer, R. Refractometry and interferometry of living cells. J. Opt. Soc. Am. 1957, 47, 545–556. [Google Scholar] [CrossRef]
- Enoch, J.M.; Tobey, F.L., Jr. Use of the waveguide parameter V to determine the difference in the index of refraction between the rat rod outer segment and the interstitial matrix. J. Opt. Soc. Am. 1978, 68, 1130–1134. [Google Scholar] [CrossRef]
- Spaide, R.F.; Curcio, C.A. Anatomical correlates to the bands seen in the outer retina by optical coherence tomography: Literature review and model. Retin. J. Ret. Vit. Dis. 2011, 31, 1609–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thylén, L. The beam propagation method: An analysis of its applicability. Opt. Quant. Electron. 1983, 15, 433–439. [Google Scholar] [CrossRef]
- Feit, M.D.; Fleck, J.A., Jr. Light propagation in graded-index optical fibers. Appl. Opt. 1978, 17, 3990–3998. [Google Scholar] [CrossRef] [PubMed]
- Labin, A.M.; Safuri, S.K.; Ribak, E.N.; Perlman, I. Müller cells separate between wavelengths to improve day vision with minimal effect upon night vision. Nat. Commun. 2014, 5, 4319. [Google Scholar] [CrossRef] [Green Version]
- Kreysing, M.; Boyde, L.; Guck, J.; Chalut, K.J. Physical insight into light scattering by photoreceptor cell nuclei. Opt. Lett. 2010, 35, 2639–2641. [Google Scholar] [CrossRef]
- Sawides, L.; de Castro, A.; Burns, S.A. The organization of the cone photoreceptor mosaic measured in the living human retina. Vis. Res. 2017, 132, 34–44. [Google Scholar] [CrossRef]
- Litts, K.M.; Cooper, R.F.; Duncan, J.L.; Carroll, J. Photoreceptor-Based Biomarkers in AOSLO Retinal Imaging. Investig. Ophthalmol. Vis. Sci. 2017, 58, BIO255–BIO267. [Google Scholar] [CrossRef] [Green Version]
- Foote, K.G.; Loumou, P.; Griffin, S.; Qin, J.; Ratnam, K.; Porco, T.C.; Roorda, A.; Duncan, J.L. Relationship Between Foveal Cone Structure and Visual Acuity Measured with Adaptive Optics Scanning Laser Ophthalmoscopy in Retinal Degeneration. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3385–3393. [Google Scholar] [CrossRef]
- Burns, S.A.; Elsner, A.E.; Sapoznik, K.A.; Warner, R.L.; Gast, T.J. Adaptive optics imaging of the human retina. Prog. Retin. Eye Res. 2019, 68, 1–30. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, A.; Yu, K.; Min, X.; Li, J.; Li, J.; Zuo, X.; Yao, Y. The Explanation of Photopic Luminous Efficiency Curve by Using Both of the Cones’ Optical Fiber Coupling Effects and the Absorption of L Cones. Sensors 2023, 23, 1523. https://doi.org/10.3390/s23031523
Liang A, Yu K, Min X, Li J, Li J, Zuo X, Yao Y. The Explanation of Photopic Luminous Efficiency Curve by Using Both of the Cones’ Optical Fiber Coupling Effects and the Absorption of L Cones. Sensors. 2023; 23(3):1523. https://doi.org/10.3390/s23031523
Chicago/Turabian StyleLiang, Anhui, Kexin Yu, Xiaolin Min, Jing Li, Jianyu Li, Xiaoling Zuo, and Youli Yao. 2023. "The Explanation of Photopic Luminous Efficiency Curve by Using Both of the Cones’ Optical Fiber Coupling Effects and the Absorption of L Cones" Sensors 23, no. 3: 1523. https://doi.org/10.3390/s23031523