Gaussia Luciferase as a Reporter for Quorum Sensing in Staphylococcus aureus
by
 , , , and
, , , and
Isobel Blower
1,† ,
,
Carmen Tong
1,2,†,‡,
Xiaohui Sun
2,3,
Ewan Murray
1,
Jeni Luckett
1,
Weng Chan
4,
Paul Williams
1 and
Philip Hill
1,2,* 1
School of Life Sciences, Biodiscovery Institute, University of Nottingham, University Park, Nottingham NG7 2RD, UK
2
School of Biosciences, Sutton Bonington Campus, University of Nottingham, Loughborough LE12 5RD, UK
3
College of Chemical Engineering, Huaqiao University, 668 Jimei Blvd, Xiamen 361021, China
4
School of Pharmacy, Biodiscovery Institute, University of Nottingham, University Park, Nottingham NG7 2RD, UK
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
‡
Present address: Sygnature Discovery Limited, BioCity, Pennyfoot Street, Nottingham NG1 1GR, UK.
Sensors 2020, 20(15), 4305; https://doi.org/10.3390/s20154305
Submission received: 30 June 2020
/
Revised: 23 July 2020
/
Accepted: 30 July 2020
/
Published: 1 August 2020
(This article belongs to the Special Issue Advances in Optical, Fluorescent and Luminescent Biosensors)
Abstract
:Gaussia luciferase (GLuc) is a secreted protein with significant potential for use as a reporter of gene expression in bacterial pathogenicity studies. To date there are relatively few examples of its use in bacteriology. In this study we show that GLuc can be functionally expressed in the human pathogen Staphylococcus aureus and furthermore show that it can be used as a biosensor for the agr quorum sensing (QS) system which employs autoinducing peptides to control virulence. GLuc was linked to the P3 promoter of the S. aureus agr operon. Biosensor strains were validated by evaluation of chemical agent-mediated activation and inhibition of agr. Use of GLuc enabled quantitative assessment of agr activity. This demonstrates the utility of Gaussia luciferase for in vitro monitoring of agr activation and inhibition.
1. Introduction
Staphylococcus aureus has been identified by the World Health Organization as a high priority pathogen [1]. It is one of the most common causes of bacterial infections in both hospitals and in the community [2]. Infections caused by S. aureus are difficult to treat due to the ability of this bacterium to acquire resistance to multiple antibiotics rapidly and to grow as a biofilm which limits the efficacy of diverse antibiotics and biocides [3,4]. There is, therefore, an urgent need to develop antimicrobials with novel targets which are effective against resistant strains. As well as traditional drugs, alternative therapies are being investigated which are less likely to drive the development of resistance [5]. These include compounds which will target functions essential for infection such as colonization and virulence factor production without affecting bacterial growth [6].
In S. aureus the accessory gene regulator (agr) quorum sensing system is a key regulator of virulence factor expression [2,7]. Quorum sensing (QS) is a means of cell to cell communication in bacterial populations. It coordinates cooperative behavior by enabling bacteria at high population densities to respond to altered environments [8]. Bacteria communicate using secreted signal molecules called autoinducers. When the extracellular concentration of the autoinducer reaches a threshold level, a signal transduction pathway is activated, leading to targeted gene activation or repression [7].
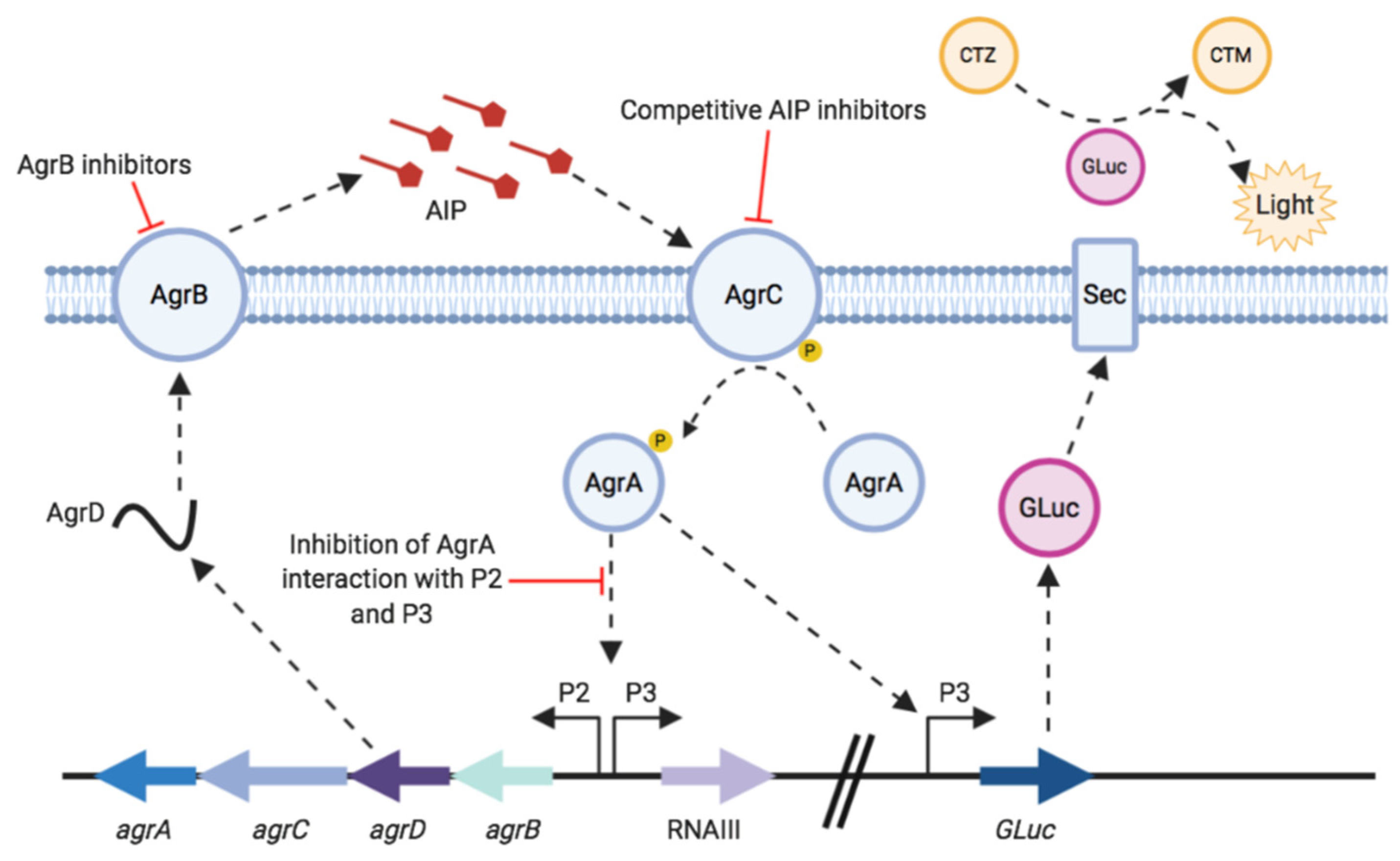
For the S. aureus agr system, as outlined in Figure 1, the quorum sensing signal molecule is an autoinducing peptide (AIP) [9]. AIPs are produced by post-translational N- and C-terminal processing of the AgrD pre-propeptide by AgrB and exported from the cell [7,10]. Once an AIP reaches a threshold concentration outside the cell it binds to and activates AgrC in the cell membrane. This autophosphorylates in response to AIP binding and in turn phosphorylates AgrA, the response regulator. Activated AgrA binds to the P2 and P3 promoters, upregulating agrBCDA and RNAIII expression [10,11,12]. RNAIII is the effector of the agr system, it encodes δ-haemolysin and also acts as a regulatory RNA that controls expression of multiple virulence genes post-transcriptionally [7].
There is increasing evidence to suggest that blocking S. aureus virulence gene expression attenuates infection in certain experimental animal infection models [14,15]. This has been achieved through mutation of agr or inhibition of agr via various natural and synthetic compounds. Inhibitors of agr target different components within this quorum sensing system including those which target AgrB to block AIP generation, competitive AIP inhibitors (also known as AgrC antagonists) which prevent AgrC activation and inhibition of P2 and P3 activation by AgrA (Figure 1) [13]. Attenuating virulence early in infection could allow the host immune system to respond and clear the pathogen, sparing exposure to antibiotics [8]. Virulence inhibitors could also be used as adjuvants with existing antibiotics, or with therapies which boost the host immune response [16,17].
Currently it is unclear when or where staphylococcal agr is expressed within the infection site and whether agr inhibitors could be used against established infections or as prophylactics. In order to investigate temporal agr expression and agr inhibition it is important to develop methods for real time monitoring of agr activity which can be used in vivo [18]. This can be performed using bioluminescent reporters linked to promoters of the agr system [19]. In previous studies, the Photorhabdus luminescens lux operon linked to the agrP3 promoter has been used to measure temporal agr activation in infection models and assess agr inhibitors in vivo [14,15]. agrP3 lux based systems have also been used for high-throughput screening of agr inhibitors [12] though it has been reported that some compounds such as Pseudomonas aeruginosa quorum sensing molecule 3-oxo-C12-HSL directly inhibit Lux-based bioluminescence, potentially leading to false positive hits [20]. Lux-based systems also require oxygen and so are potentially compromised in environmental niches where oxygen levels are low. In this study, a novel bioluminescent reporter linked to the P3 promoter of the agr system was validated for use in S. aureus.
Luciferases are often used as biological reporters due to their high sensitivity, broad dynamic range and operational simplicity. Luciferases have been used to monitor multiple cellular processes, such as gene expression and protein interactions as well as tracking of bacterial and viral infection [21,22]. Traditionally, bacterial luciferases have been used as reporter genes in staphylococci, however this system requires substrate and energy supply from the cells, at the point of live-cell assay. Novel luciferases such as the Gaussia luciferase, which is small, secreted and ATP-independent, broaden the potential applications and provide the ability to detect bioluminescence extracellularly and without the need for extra energy from live cells at the point of the assay [23].
Gaussia luciferase (GLuc) is a naturally secreted enzyme from the marine copepod Gaussia princeps [24]. GLuc is a 185 amino acid, 19.9 kDa luciferase which catalyses oxidative decarboxylation of the substrate coelenterazine (CTZ) producing coelenteramide (CTM), emitting blue light at 480 nm [25]. GLuc is nontoxic and is naturally secreted from the cell therefore is useful for monitoring biological processes without the requirement for cellular energy to drive the luminescent reaction [26]. GLuc is a useful reporter due to its small size and high light intensity, and hence delivering assays with high sensitivity. GLuc is also stable at a wide range of pHs and is resistant to heat shock and oxidative stress [27]. GLuc has been shown to be an effective biological reporter in bacteria, such as Salmonella enterica and Mycobacterium smegmatis [26,28]. It is a highly sensitive reporter for nondestructive quantification of in vivo biological processes ex vivo, as bioluminescence can be detected from the blood or urine of in vivo models using a luminometer [29,30].
The aim of this work was to demonstrate the use of Gaussia luciferase as a reporter in S. aureus and for monitoring activation of the agr quorum sensing system (see Figure 1). Reporter strains were constructed with GLuc chromosomally encoded under the control of the agrP3 promoter in S. aureus USA300 strain JE2 and an isogenic Δagr mutant. These were validated by evaluation of activation and inhibition of the agr system using the cognate AIP as agonist and synthetic (Ala5)AIP-1 as an antagonist and by testing both nonpeptidic AgrC allosteric and AgrB inhibitors. The data obtained show that GLuc can be used successfully for in vitro monitoring of agr activation and inhibition.
2. Materials and Methods
Staphylococcal strains were cultured in tryptone soy broth (TSB) (Oxoid) and E. coli in lysogeny broth (LB) (Oxoid) with antibiotics added as required. All strains were grown at 37 °C and overnight cultures were grown with aeration by shaking at 200 rpm. AIPs, N-(3-oxododecanoyl)-L-homoserine lactone (3-oxo-C12-HSL, (S)-3-dodecanoyl-5-(2-hydroxyethyl) tetramic acid (C12-TMA) and 3-dodecanoyltetronic acid (C12-TOA) were synthesized as described before. Ambuic acid was purchased from Cayman Chemicals.
2.1. Construction of agrP3 Reporter S. aureus Strains
A transcriptional reporter fusion was constructed with Gaussia encoded chromosomally under the control of the agrP3 promoter. All strains and primers used are listed in Table 1 and Table 2, respectively. To ensure GLuc would be secreted by S. aureus, the protein A (SpA) leader was engineered upstream of a synthetic GLuc gene (kind gift of S. Wiles, University of Auckland, Auckland, New Zealand). This replaced the sequence of the original native eukaryotic leader to direct the translated protein to the S. aureus Sec secretory pathway. This fragment was assembled into pUNKdestR3R4 flanked by the Pxyl/tet promoter and the rrnB1B2 transcriptional terminator to create plasmid pUNK1 pXyl/tet:SpA leader:GLuc using Gateway cloning [31] and provided for constitutive expression of Gluc in S. aureus. The GLuc gene was then amplified by PCR from this vector using primer pair CT16 and CT17 using high fidelity Phusion DNA polymerase (Thermofisher). The agrP3 promoter was also amplified by PCR from S. aureus 8325-4 genomic DNA using primer pair CT14 and CT15 and assembled upstream of GLuc into pUNK1 using Hifi DNA assembly (NEB). This was used to transform electrocompetent E. coli DC10B. Plasmids were extracted using a miniprep kit (Qiagen) before confirmation by sequencing (Source Bioscience).
Chromosomal integration of agrP3-GLuc into S. aureus was performed as outlined by Lei et al. [33] using integration vector pLL102. Plasmids pUNK1 agrP3:SpA leader:GLuc and pLL102 were digested with SalI and fragments ligated using T4 DNA ligase (NEB) and used to transform electrocompetent E. coli DC10B to create plasmid pLL102 agrP3:SpA leader:GLuc. This plasmid was used to transform electrocompetent S. aureus RN4220 attB2. Chromosomal integration was checked by colony PCR using primer pairs OU9R10 and SCV4, and SCV8 and OU9R7 and the correct strain named S. aureus RN4220 attB2 agrP3 SpA leader GLuc. This strain was used as a donor for phage transduction into S. aureus USA300 strain JE2 using phage phi11 (Φ11). Further phage transduction was performed with a Δagr Φ11 phage lysate. This resulted in the construction of reporter strains USA300 agrP3-GLuc and USA300 Δagr P3 GLuc.
2.2. Measuring agrP3 Promoter Activity Using GAUSSIA Luciferase Bioluminescence
When testing bioluminescence output from GLuc over time, a single colony of the test strain was used to inoculate 50 mL of TSB, which was incubated at 37 °C, 200 rpm. An OD600 reading of a 1 mL sample was taken every hour for 12 h. The supernatant fraction was separated from the cells by centrifugation twice at 13,000× g for 2 min. Samples were stored at −20 °C prior to analysis. Freeze thawing has been shown to have no effect on the stability of Gaussia luciferase [38]. When testing the effect of AIP-1 and ambuic acid, a single colony was grown in 5 mL of TSB broth for 16 h with test compounds added at the desired concentrations at the start of incubation. When testing the effect of Ala5(AIP-1) and 3-oxo-C12-HSL analogues these compounds were added to broth with a single colony as before and incubated for 6 h. Cultures were incubated at 37 °C at 200 rpm and samples taken as described previously.
Samples were thawed at room temperature and 100 µL loaded into a black, flat clear-bottomed 96 well plate (Greiner). Bioluminescence was recorded as Relative Light Units (RLU) using a 96 well-plate reader and injector (Infinite F200 Tecan): 50 µL of 20 µM coelenterazine (CTZ) substrate was injected at 200 µL/s and bioluminescence read with an integration time of 100 ms after a wait time of 1 s. A portion of 20 µM CTZ was used as this was shown to saturate the enzyme. CTZ was injected into each well and bioluminescence read in turn, 3 technical repeats were recorded for each sample. Statistical analysis was performed using an unpaired t-test with Welch’s correction.
3. Results
3.1. Induction of agr with Respect to Growth Phase
In S. aureus, agr-dependent quorum sensing is activated at high cell population densities when the autoinducing AIP reaches a threshold level. This has been reported to occur in vitro in the late exponential/early stationary phase of growth [11]. When the AIP reaches the threshold concentration the agrP3 promoter is activated, therefore GLuc expression will be induced via agrP3. In order to determine whether the production of GLuc bioluminescence occurs as a function of the growth phase consistent with agr induction, bioluminescence and OD600 were quantified over 12 h using S. aureus USA300 agrP3-GLuc.
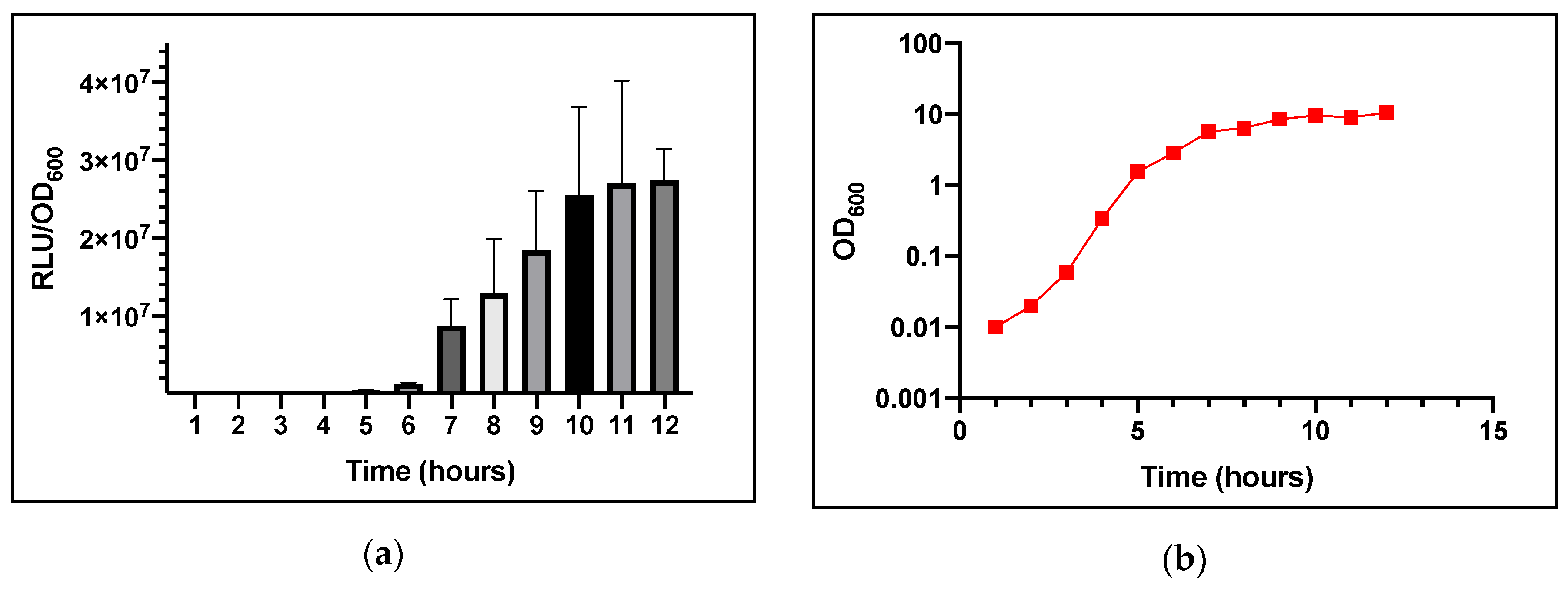
A single colony was inoculated into 50 mL TSB broth with 0.01% v/v DMSO added as a solvent control. Growth did not appear to be attenuated. As shown in Figure 2a, light output was observed after 6–7 h of growth. From 5–7 h, the level of bioluminescence increased substantially, from ~4.83 × 105 to 8.74 × 106 RLU/OD600 (Relative Light Units/Optical Density at 600 nm). The increase in bioluminescence reached a plateau after 10 h at ~2.7 × 107 RLU/OD600 which is consistent with agr expression.
3.2. Activation of USA300 Δagr P3-GLuc pSKermP2 agrAC with Synthetic AIP-1
To confirm the AIP-dependent expression of the agrP3-Gluc construct we set out to demonstrate that the reporter can be activated with synthetic, exogenous AIP-1. The agrA and agrC genes which constitute the agr two-component signal transduction system, were introduced into USA300 Δagr P3-GLuc on plasmid pSKermP2 agrAC [36]. Genetic complementation with agrA and agrC would enable the strain to respond to exogenous AIP-1 by upregulation of the agrP3-GLuc promoter due to signal transduction from AgrC and AgrA [36]. Due to the lack of AgrB and AgrD the strain could not itself synthesize AIP-1.
The response of this strain to synthetic AIP-1 was demonstrated by the addition of 100 nM AIP-1 to an overnight culture. As a control, 0.01% v/v DMSO, the solvent used for AIP-1 was tested, as well as USA300 Δagr P3-GLuc with addition of 100 nM AIP-1.
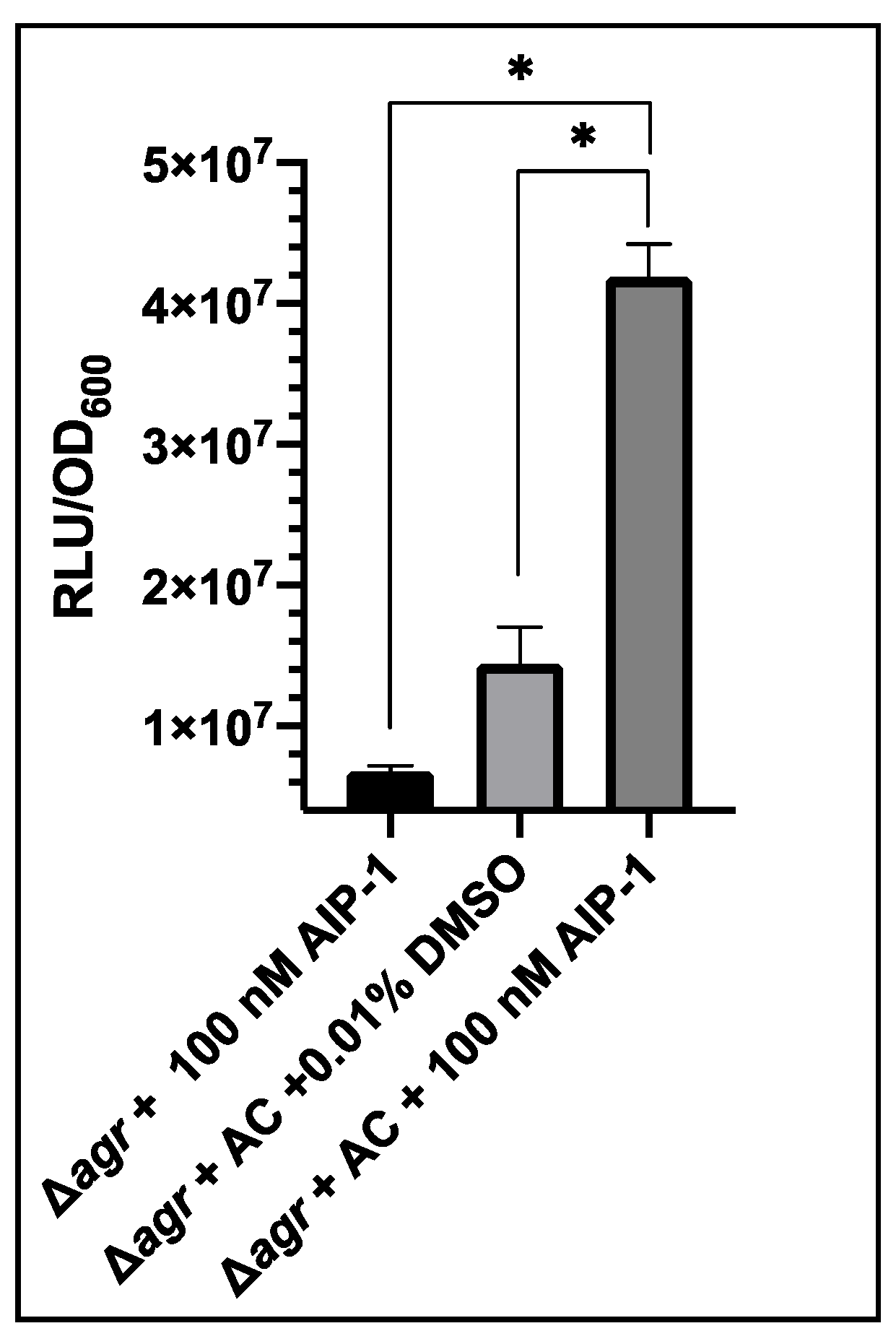
As shown in Figure 3 after addition of AIP-1 to USA300 Δagr P3-GLuc the level of bioluminescence was ~6.78 × 106 RLU/OD600. This demonstrates that AIP-1 does not activate quorum sensing in the Δagr strain. When complemented with agrA and agrC, the 0.01% v/v DMSO control produced bioluminescence of ~1.4 × 107 RLU/OD600, indicating the levels of background luminescence when the AgrA transcription activator was present. On exogenous provision of 100 nM AIP-1 this increased to ~4.19 × 107 RLU/OD600. This was a significant increase showing the strain responded productively to exogenous AIP-1.
3.3. Inhibition of S. aureus USA300 agrP3 by an AgrC Antagonist (Ala5)AIP-1
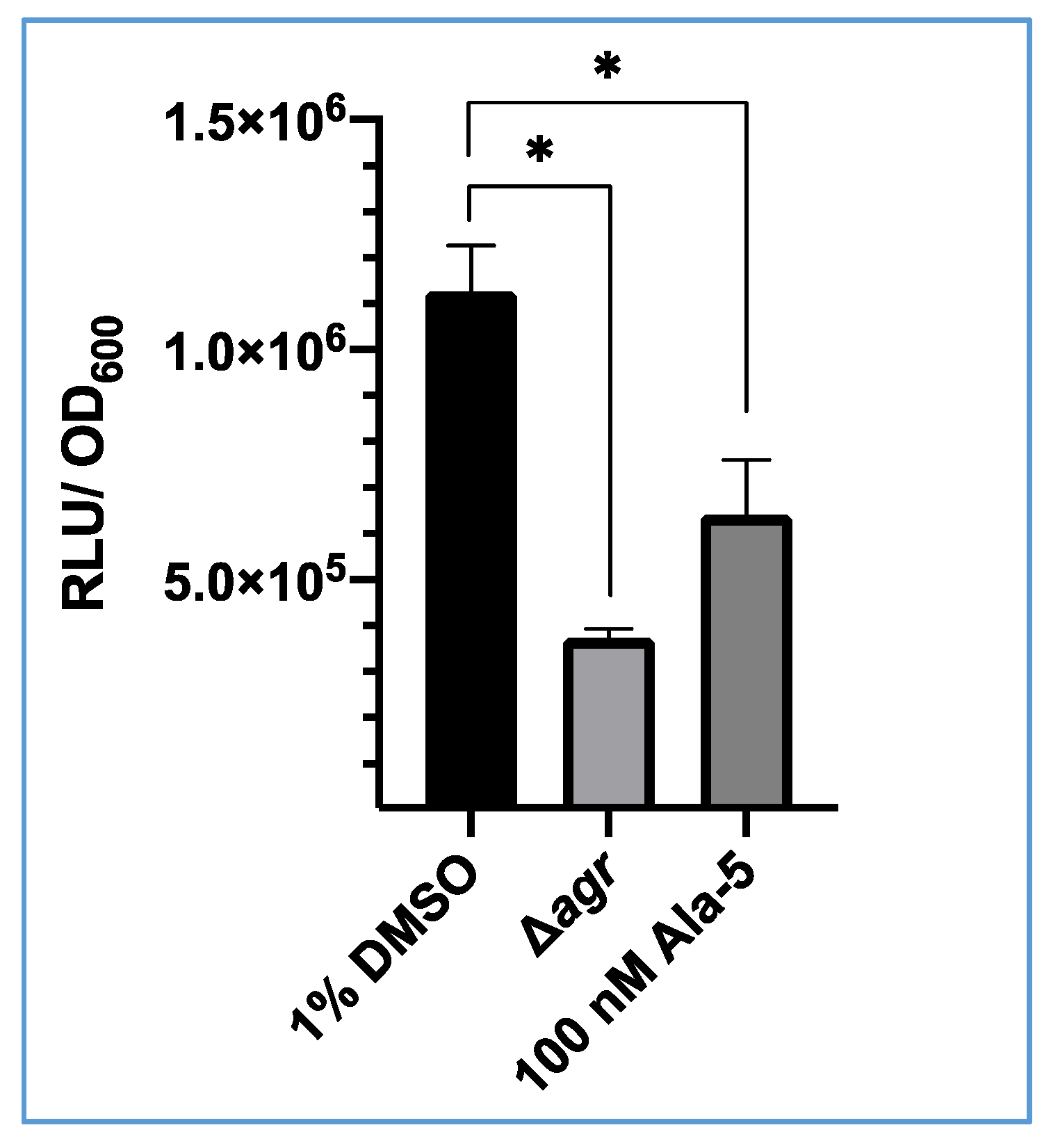
To demonstrate that GLuc expression is under the control of the agrP3 promoter, S. aureus USA300 agrP3-GLuc was treated with (Ala5)AIP-1, a competitive inhibitor of the agr system [39]. Bioluminescence was recorded after incubation for 6 h. As shown in Figure 4, after 6 h there was a significant decrease in bioluminescence observed when treated with 100 nM (Ala5)AIP-1 (~6.4 × 105 RLU) compared with the 1% v/v DMSO control (1.13 × 106 RLU). This demonstrates that bioluminescence from this strain can be inhibited with addition of a competitive antagonist of the agr system.
3.4. Evaluation of an AgrB Inhibitor with USA300 agrP3-GLuc
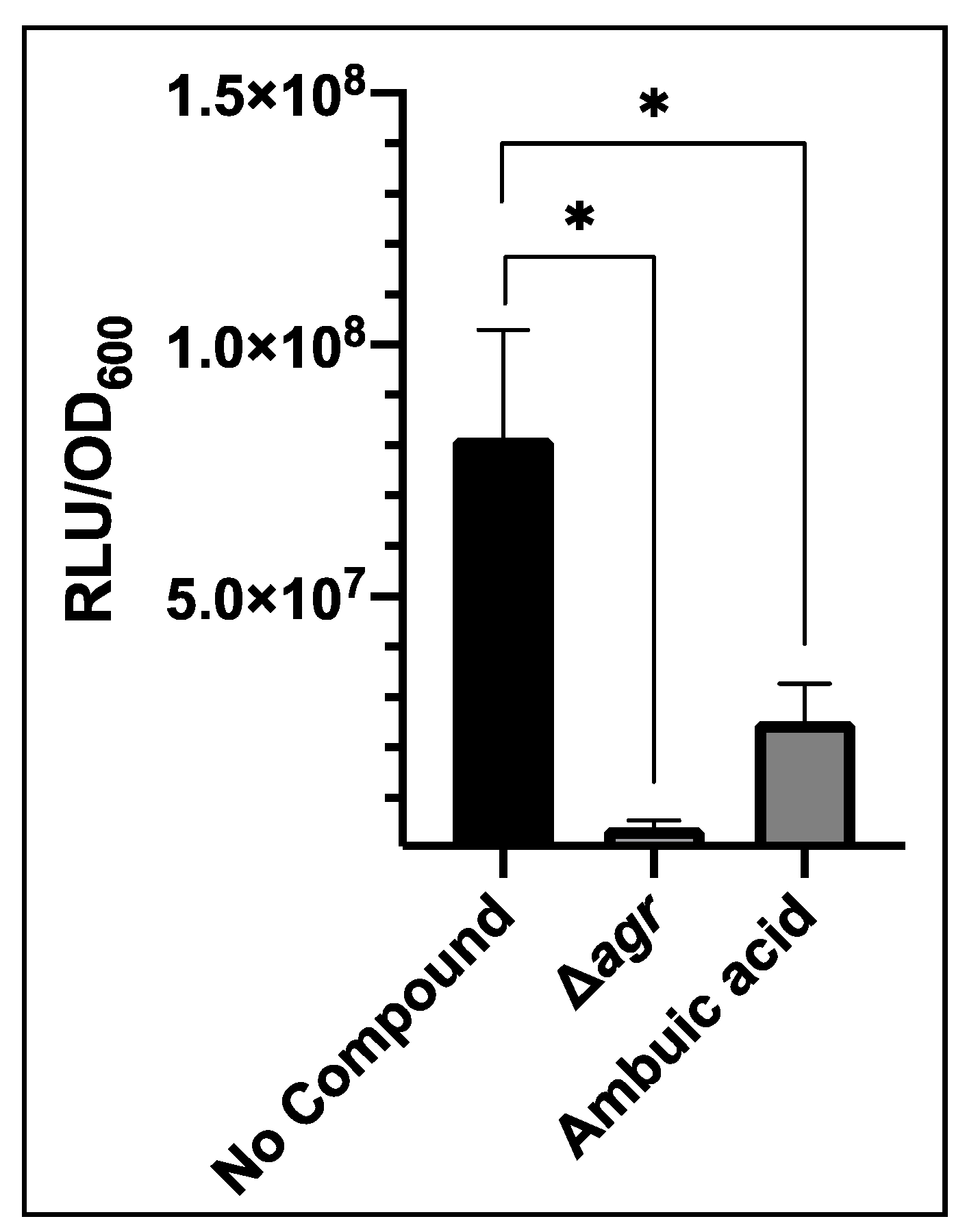
Inhibitors of other targets in the agr system have been identified, such as ambuic acid which blocks AIP biosynthesis by targeting AgrB [15]. This was tested against USA300 agrP3-GLuc. Addition of 40 µM ambuic acid significantly reduced bioluminescence compared with the control to 2.52 × 107 RLU/OD600 (Figure 5). This was almost 4-fold lower than the control although bioluminescence was not reduced to the level of the agr mutant.
3.5. 3-Oxo-C12-HSL Has Differential Effects on GLuc and Lux Bioluminescence
The molecule 3-Oxo-C12-HSL is a Pseudomonas aeruginosa quorum sensing signal molecule which has inhibitory effects on the S. aureus agr quorum sensing system [40]. Murray et al. screened analogues of 3-oxo-C12-HSL and identified C12-TMA and C12-tetronic acid (C12-TOA) as the most potent inhibitors. They used a blaZ based agrP3 reporter as 3-oxo-C12-HSL affected lux-based bioluminescence making conventional lux-based agrP3 reporter systems unsuitable [40].
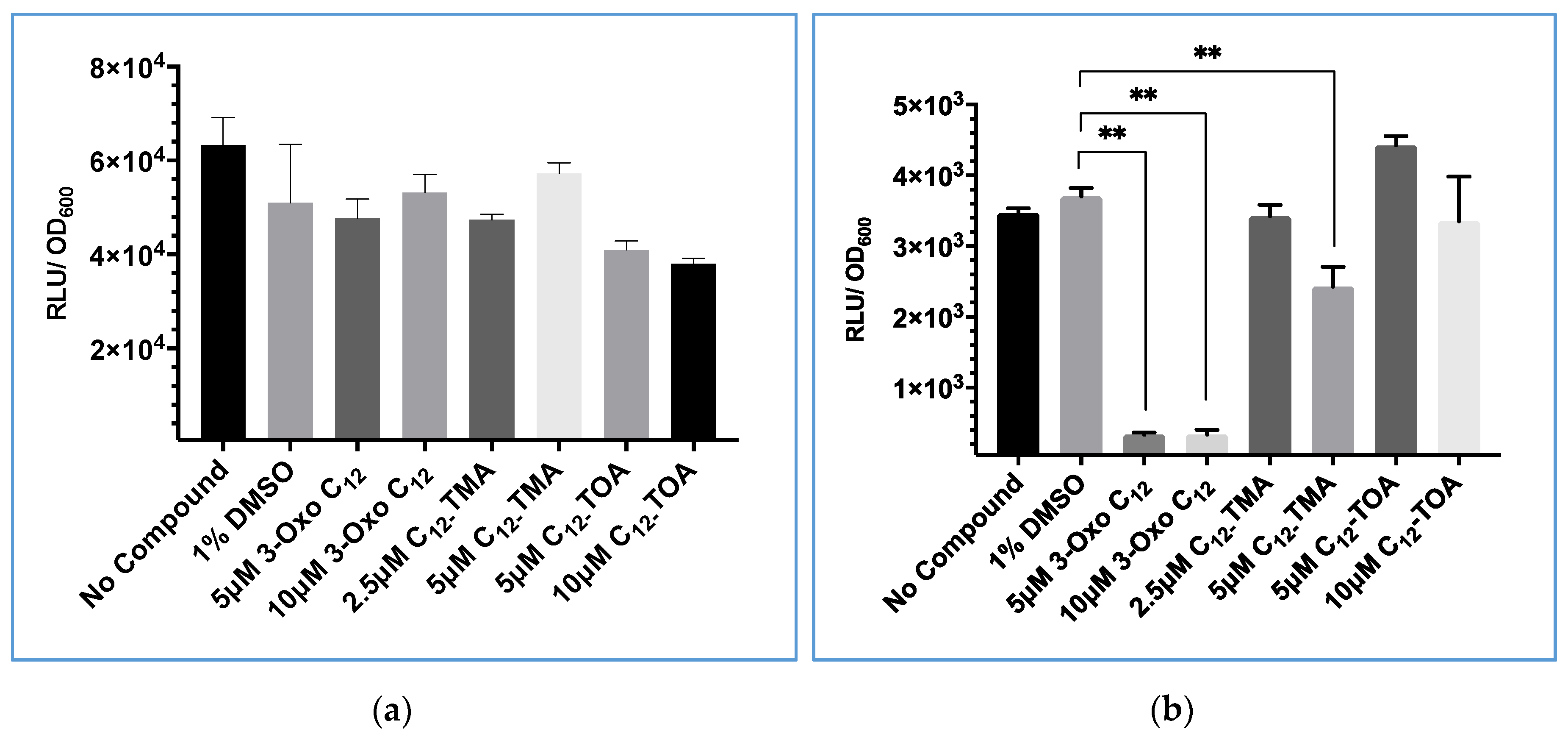
To determine whether a GLuc-based reporter could be used to screen for quorum sensing inhibitory activity with compounds that inhibited Lux-based bioluminescence, 3-oxo-C12-HSL, C12-TMA and C12-TOA were mixed with Gaussia luciferase in the supernatant of Gluc-expressing S. aureus to investigate whether they directly influenced the GLuc biochemical reaction. They were also tested on cultures of constitutively bioluminescent S. aureus expressing lux to compare the effect on lux-based bioluminescence.
As shown in Figure 6, neither 3-oxo-C12-HSL nor C12-TMA significantly affected GLuc bioluminescence compared with a 1% v/v DMSO solvent control. However, when these two compounds were incubated with Lux + S. aureus at 5 µM, light output was significantly reduced. This confirmed that lux-based bioluminescence was directly inhibited by these compounds. C12-TOA was not shown to significantly decrease either GLuc or lux-based bioluminescence at both concentrations tested.
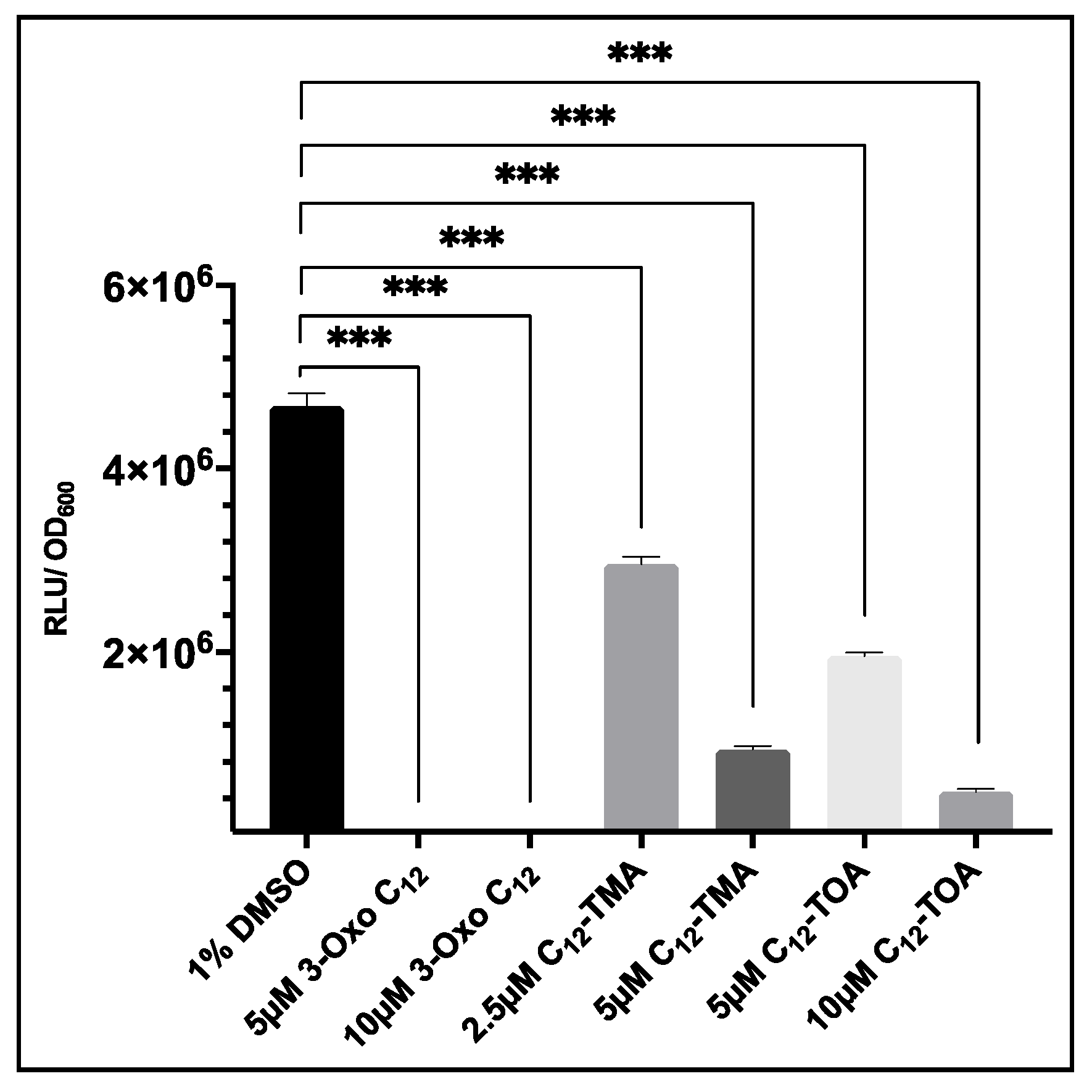
3.6. Evaluation of USA300 agrP3 GLuc as a Screen for agr Inhibitors Related to 3-oxo-C12-HSL
To evaluate whether S. aureus USA300 agrP3 GLuc could be used to screen analogues of 3-oxo-C12-HSL, this strain was cultured in the presence or absence of a range of concentrations of 3-oxo-C12-HSL, C12-TMA and C12-TOA and bioluminescence read after 6 h. As shown in Figure 7, addition of 5 µM or 10 µM of 3-oxo-C12-HSL significantly reduced bioluminescence compared to the 1% DMSO control. The addition of C12-TMA and C12-TOA also reduced bioluminescence which confirmed findings of agr inhibition by these compounds reported by Murray et al. (2014). This demonstrates that S. aureus USA300 agrP3 GLuc can be used to screen inhibitors of agr-dependent quorum sensing, where screening by Lux is precluded due to direct inhibition of Lux bioluminescence.
4. Discussion
Antibiotic resistance in many bacterial pathogens including S. aureus is increasing and antibiotics of last resort are increasingly used to treat these infections, which in turn is leading to the emergence of resistance to these high-value antibiotics. There is an urgent need for novel therapeutics which do not succumb to conventional mechanisms of antibiotic resistance. One method is through attenuation of bacterial virulence. In S. aureus, pathogenicity is largely controlled by the agr quorum sensing system, therefore this is a potential target for antivirulence drugs [13,41]. It is therefore important to develop simple methods for monitoring of agr activity in order to facilitate screening for inhibitors and gain information about agr activation in infection models in vivo. In this study we have demonstrated that Gaussia luciferase can be used as a reporter for agr dependent quorum sensing in S. aureus. The ability of GLuc to report agr activation in vitro was demonstrated by comparing bioluminescence over time with OD600. This showed that bioluminescence increased in the late log phase of growth when agr is activated [11]. This confirmed that the reporter functioned as anticipated and is likely to be useful for monitoring temporal agr activation both in vitro and potentially in vivo in experimental animal infection models.
To demonstrate that GLuc activation is under the control of the agr system, the response to synthetic AIP-1 was first evaluated. Activation of the agr system in vitro in wild type S. aureus strains by addition of exogenous synthetic AIP-1 can be difficult to observe because of other regulators of the agr system. For example, CodY represses agr activation during exponential growth in rich media, preventing agr activation before depletion of key nutrients [42]. Therefore, in order to evaluate the response of the reporter to exogenous AIP-1, agrA and agrC which make up the two-component signal transduction system were introduced into USA300 Δagr P3-GLuc on plasmid pSKermP2. This plasmid encodes agrC and agrA under the control of the P2 promoter of the agr system [36]. Complementation of agrA and agrC enabled the Δagr strain to respond to exogenous AIP-1. This was shown by the increased light output on provision of AIP-1 compared with the DMSO control.
It was demonstrated that for USA300 agrP3-GLuc the expression of Gaussia luciferase was dependent on activation of the agr QS system. This was further validated by addition of a competitive antagonist of the agr system. Addition of (Ala5)AIP-1 reduced bioluminescence. This confirmed that inhibition of the agr system also inhibited GLuc activation, which was consistent with the control of light output through activation of QS. Similar results were obtained with C12-TMA and C12-TOA which are noncompetitive, negative allosteric modulators of AgrC [40].
This work also demonstrates the potential of the GLuc reporter in identifying inhibitors of other agr system components. Ambuic acid, an AgrB inhibitor, was tested against USA300 agrP3-GLuc. Ambuic acid has been tested previously in in vivo models with an agrP3-lux reporter where real time reduction of agr activity was seen [15]. This shows that GLuc can be used as a reporter to screen for agr inhibitors and evaluate their potential as prophylactic and therapeutic agents.
Previously, bacterial lux-based reporter systems have been used to study agr quorum sensing however it has been demonstrated that some agr inhibitors can interfere with the luciferase activity giving false results. Here it was demonstrated that 3-oxo-C12 HSL interfered with the lux signal however did not interfere with GLuc bioluminescence. It was shown that QS inhibitors which have an effect on lux-based bioluminescence could be reliably assayed using GLuc, demonstrating that it was a useful alternative output for bacterial reporter systems. Additionally, the output from lux-based reporters may be affected by metabolic effects decreasing the availability of bioluminescence substrates such as FMNH2 at the point of assay, therefore any reduction in bioluminescence could be due to metabolic factors rather than a reduction of agr activity [43]. GLuc does not require cellular cofactors during assay therefore directly provides information about agr activation, avoiding the reduction in specificity associated with the loss of metabolites. This work demonstrates that agrP3 GLuc reporters can be used as a useful alternative or adjunct to screens for inhibitors of the agr system in vitro.
Together these data show that the agrP3-GLuc reporter can be activated by exogenous cognate AIP-1 and inhibited by antagonists of the agr system similar to agrP3-lux and agrP3-blaZ reporters [36,39,44]. This highlights the use of the Gaussia luciferase as a novel reporter of agr activity in S. aureus in vitro and for potential screening for activators and inhibitors of the agr system.
Author Contributions
Conceptualization, P.H., P.W. and W.C.; methodology, P.H., W.C. and P.W.; investigation, I.B., C.T., X.S., J.L. and E.M.; formal analysis, I.B., C.T., J.L. and E.M.; visualization, I.B. and C.T.; writing—original draft preparation, I.B. and C.T.; writing—review and editing, I.B., C.T., P.H., P.W. and W.C.; supervision, P.H., J.L., P.W. and W.C.; funding acquisition, P.W., W.C. and P.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded via the Wellcome Trust and Medical Research Council UK postgraduate studentships and by a Medical Research Council Programme grant (MR/N010477/1).
Acknowledgments
Thanks to Siouxsie Wiles for providing synthetic Gluc gene and Alex Truman for synthesizing C12-TMA and C12-TOA.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Asokan, G.V.; Ramadhan, T.; Ahmed, E.; Sanad, H. WHO Global Priority Pathogens List: A Bibliometric Analysis of Medline-PubMed for Knowledge Mobilization to Infection Prevention and Control Practices in Bahrain. Oman Med. J. 2019, 34, 184–193. [Google Scholar] [CrossRef]
- Queck, S.Y.; Jameson-Lee, M.; Villaruz, A.E.; Bach, T.H.; Khan, B.A.; Sturdevant, D.E.; Ricklefs, S.M.; Li, M.; Otto, M. RNAIII-independent target gene control by the agr quorum-sensing system: Insight into the evolution of virulence regulation in Staphylococcus aureus. Mol. Cell 2008, 32, 150–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boles, B.R.; Horswill, A.R. Agr-mediated dispersal of Staphylococcus aureus biofilms. PLoS Pathog. 2008, 4, e1000052. [Google Scholar] [CrossRef] [PubMed]
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin Resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar] [PubMed]
- Aminov, R.I. A brief history of the antibiotic era: Lessons learned and challenges for the future. Front. Microbiol. 2010, 1, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clatworthy, A.E.; Pierson, E.; Hung, D.T. Targeting virulence: A new paradigm for antimicrobial therapy. Nat. Chem. Biol. 2007, 3, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Quorum-sensing control in Staphylococci—A target for antimicrobial drug therapy? FEMS Microbiol. Lett. 2004, 241, 135–141. [Google Scholar] [CrossRef]
- Gray, B.; Hall, P.; Gresham, H. Targeting agr- and agr-Like quorum sensing systems for development of common therapeutics to treat multiple gram-positive bacterial infections. Sensors 2013, 13, 5130–5166. [Google Scholar] [CrossRef] [Green Version]
- Novick, R.P.; Projan, S.J.; Kornblum, J.; Ross, H.F.; Ji, G.; Kreiswirth, B.; Vandenesch, F.; Moghazeh, S. The agr P2 operon: An autocatalytic sensory transduction system in Staphylococcus aureus. Mol. Gen. Genet. 1995, 248, 446–458. [Google Scholar] [CrossRef]
- Novick, R.P. Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol. Microbiol. 2003, 48, 1429–1449. [Google Scholar] [CrossRef]
- Sloan, T.J.; Murray, E.; Yokoyama, M.; Massey, R.C.; Chan, W.C.; Bonev, B.B.; Williams, P. Timing is Everything: Impact of Naturally Occurring Staphylococcus aureus AgrC Cytoplasmic Domain Adaptive Mutations on Autoinduction. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desouky, S.E.; Nishiguchi, K.; Zendo, T.; Igarashi, Y.; Williams, P.; Sonomoto, K.; Nakayama, J. High-throughput screening of inhibitors targeting Agr/Fsr quorum sensing in Staphylococcus aureus and Enterococcus faecalis. Biosci. Biotechnol. Biochem. 2013, 77, 923–927. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.P.; Williams, P.; Chan, W.C. Attenuating Staphylococcus aureus virulence gene regulation: A medicinal chemistry perspective. J. Med. Chem. 2013, 56, 1389–1404. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.S., III; Jin, R.; Novick, R.P. Transient interference with Staphylococcal Quorum Sensing blocks abscess formation. Proc. Natl. Acad. Sci. USA 2005, 102, 1691–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, D.A.; Parlet, C.P.; Crosby, H.A.; Malone, C.L.; Heilmann, K.P.; Horswill, A.R.; Cech, N.B. Signal Biosynthesis Inhibition with Ambuic Acid as a Strategy To Target Antibiotic-Resistant Infections. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Daly, S.M.; Elmore, B.O.; Kavanaugh, J.S.; Triplett, K.D.; Figueroa, M.; Raja, H.A.; El-Elimat, T.; Crosby, H.A.; Femling, J.K.; Cech, N.B.; et al. Omega-Hydroxyemodin limits staphylococcus aureus quorum sensing-mediated pathogenesis and inflammation. Antimicrob. Agents Chemother. 2015, 59, 2223–2235. [Google Scholar] [CrossRef] [Green Version]
- Sully, E.K.; Malachowa, N.; Elmore, B.O.; Alexander, S.M.; Femling, J.K.; Gray, B.M.; DeLeo, F.R.; Otto, M.; Cheung, A.L.; Edwards, B.S.; et al. Selective chemical inhibition of agr quorum sensing in Staphylococcus aureus promotes host defense with minimal impact on resistance. PLoS Pathog. 2014, 10, e1004174. [Google Scholar] [CrossRef]
- Kavanaugh, J.S.; Horswill, A.R. Impact of Environmental Cues on Staphylococcal Quorum Sensing and Biofilm Development. J. Biol. Chem. 2016, 291, 12556–12564. [Google Scholar] [CrossRef] [Green Version]
- Alksne, L.E.; Projan, S.J. Bacterial virulence as a target for antimicrobial chemotherapy. Curr. Opin. Biotechnol. 2000, 11, 625–636. [Google Scholar] [CrossRef]
- Qazi, S.; Middleton, B.; Muharram, S.H.; Cockayne, A.; Hill, P.; O’Shea, P.; Chhabra, S.R.; Cámara, M.; Williams, P. N-acylhomoserine lactones antagonize virulence gene expression and quorum sensing in Staphylococcus aureus. Infect. Immun. 2006, 74, 910–919. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, k.; Vidugiris, G.; Machleidt, T.; et al. Engineered luciferase reporter from a deep sea shrimp utilizing a novel imidazopyrazinone substrate. ACS Chem. Biol. 2012, 7, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Shi, P.; Song, W.; Bi, S. Chemiluminescence and Bioluminescence Imaging for Biosensing and Therapy: In Vitro and In Vivo Perspectives. Theranostics 2019, 9, 4047–4065. [Google Scholar] [CrossRef] [PubMed]
- England, C.G.; Ehlerding, E.B.; Cai, W. NanoLuc: A Small Luciferase Is Brightening Up the Field of Bioluminescence. Bioconjug. Chem. 2016, 27, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Maguire, C.A.; Deliolanis, N.C.; Pike, L.; Niers, J.M.; Tjon-Kon-Fat, L.A.; Sena-Esteves, M.; Tannous, B.A. Gaussia luciferase variant for high-throughput functional screening applications. Anal. Chem. 2009, 81, 7102–7106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathnayaka, T.; Tawa, M.; Sohya, S.; Yohda, M.; Kuroda, Y. Biophysical characterization of highly active recombinant Gaussia luciferase expressed in Escherichia coli. Biochim. Biophys. Acta 2010, 1804, 1902–1907. [Google Scholar] [CrossRef]
- Wu, N.; Rathnayaka, T.; Kuroda, Y. Bacterial expression and re-engineering of Gaussia princeps luciferase and its use as a reporter protein. Biochim. Biophys. Acta 2015, 1854 Pt A, 1392–1399. [Google Scholar] [CrossRef]
- Wiles, S.; Ferguson, K.; Stefanidou, M.; Young, D.B.; Robertson, B.D. Alternative luciferase for monitoring bacterial cells under adverse conditions. Appl. Environ. Microbiol. 2005, 71, 3427–3432. [Google Scholar] [CrossRef] [Green Version]
- Wille, T.; Blank, K.; Schmidt, C.; Vogt, V.; Gerlach, R.G. Gaussia princeps luciferase as a reporter for transcriptional activity, protein secretion, and protein-protein interactions in Salmonella enterica serovar typhimurium. Appl. Environ. Microbiol. 2012, 78, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Tannous, B.A. Gaussia luciferase reporter assay for monitoring biological processes in culture and in vivo. Nat. Protoc. 2009, 4, 582–591. [Google Scholar] [CrossRef]
- Wurdinger, T.; Badr, C.; Pike, L.; de Kleine, R.; Weissleder, R.; Breakefield, X.O.; Tannous, B.A. A secreted luciferase for ex vivo monitoring of in vivo processes. Nat. Methods 2008, 5, 171–173. [Google Scholar] [CrossRef] [Green Version]
- Perehinec, T.M.; Qazi, S.N.; Gaddipati, S.R.; Salisbury, V.; Rees, C.E.; Hill, P.J. Construction and evaluation of multisite recombinatorial (Gateway) cloning vectors for Gram-positive bacteria. BMC Mol. Biol. 2007, 8, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novick, R. Properties of a cryptic high-frequency transducing phage in Staphylococcus aureus. Virology 1967, 33, 155–166. [Google Scholar] [CrossRef]
- Lei, M.G.; Cue, D.; Alba, J.; Junecko, J.; Graham, J.W.; Lee, C.Y. A single copy integration vector that integrates at an engineered site on the Staphylococcus aureus chromosome. BMC Res. Notes 2012, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fey, P.D.; Endres, J.L.; Yajjala, V.K.; Widhelm, T.J.; Boissy, R.J.; Bose, J.L.; Bayles, K.W. A genetic resource for rapid and comprehensive phenotype screening of nonessential Staphylococcus aureus genes. Mbio 2013, 4, e00537-12. [Google Scholar] [CrossRef] [Green Version]
- McDougal, L.K.; Steward, C.D.; Killgore, G.E.; Chaitram, J.M.; McAllister, S.K.; Tenover, F.C. Pulsed-field gel electrophoresis typing of oxacillin-resistant Staphylococcus aureus isolates from the United States: Establishing a national database. J. Clin. Microbiol. 2003, 41, 5113–5120. [Google Scholar] [CrossRef] [Green Version]
- Jensen, R.O.; Winzer, K.; Clarke, S.R.; Chan, W.C.; Williams, P. Differential recognition of Staphylococcus aureus quorum-sensing signals depends on both extracellular loops 1 and 2 of the transmembrane sensor AgrC. J. Mol. Biol. 2008, 381, 300–309. [Google Scholar] [CrossRef]
- Monk, I.R.; Shah, I.M.; Xu, M.; Tan, M.W.; Foster, T.J. Transforming the untransformable: Application of direct transformation to manipulate genetically Staphylococcus aureus and Staphylococcus epidermidis. MBio 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Wider, D.; Picard, D. Secreted dual reporter assay with Gaussia luciferase and the red fluorescent protein mCherry. PLoS ONE 2017, 12, e0189403. [Google Scholar] [CrossRef] [Green Version]
- McDowell, P.; Affas, Z.; Reynolds, C.; Holden, M.T.; Wood, S.J.; Saint, S.; Cockayne, A.; Hill, P.J.; Dodd, C.E.; Bycroft, B.W.; et al. Structure, activity and evolution of the group I thiolactone peptide quorum-sensing system of Staphylococcus aureus. Mol. Microbiol. 2001, 41, 503–512. [Google Scholar] [CrossRef] [Green Version]
- Murray, E.J.; Crowley, R.C.; Truman, A.; Clarke, S.R.; Cottam, J.A.; Jadhav, G.P.; Steele, V.R.; O’Shea, P.; Lindholm, C.; Cockayne, A.; et al. Targeting Staphylococcus aureus quorum sensing with nonpeptidic small molecule inhibitors. J. Med. Chem. 2014, 57, 2813–2819. [Google Scholar] [CrossRef]
- Baldry, M.; Kitir, B.; Frokiaer, H.; Christensen, S.B.; Taverne, N.; Meijerink, M.; Franzyk, H.; Olsen, C.A.; Wells, J.M.; Ingmer, H. The agr Inhibitors Solonamide B and Analogues Alter Immune Responses to Staphylococccus aureus but Do Not Exhibit Adverse Effects on Immune Cell Functions. PLoS ONE 2016, 11, e0145618. [Google Scholar] [CrossRef] [PubMed]
- Roux, A.; Todd, D.A.; Velazquez, J.V.; Cech, N.B.; Sonenshein, A.L. CodY-mediated regulation of the Staphylococcus aureus Agr system integrates nutritional and population density signals. J. Bacteriol. 2014, 196, 1184–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Blinn, C.; McLeod, S.M.; Wiseman, J.W.; Newman, J.V.; Fisher, S.L.; Walkup, G.K. Secreted Gaussia princeps luciferase as a reporter of Escherichia coli replication in a mouse tissue cage model of infection. PLoS ONE 2014, 9, e90382. [Google Scholar] [CrossRef] [PubMed]
- Mayville, P.; Ji, G.; Beavis, R.; Yang, H.; Goger, M.; Novick, R.P.; Muir, T.W. Structure-activity analysis of synthetic autoinducing thiolactone peptides from Staphylococcus aureus responsible for virulence. Proc. Natl. Acad. Sci. USA 1999, 96, 1218–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Outline of the staphylococcal agr system and the P3 GLuc reporter. Critical control points of the agr system, which are amenable to targeting by inhibitors, are indicated by blunt arrows. CTZ represents coelenterazine and CTM coelenteramide. Adapted from [13].
Figure 1.
Outline of the staphylococcal agr system and the P3 GLuc reporter. Critical control points of the agr system, which are amenable to targeting by inhibitors, are indicated by blunt arrows. CTZ represents coelenterazine and CTM coelenteramide. Adapted from [13].

Figure 2.
Bioluminescence from S. aureus strain USA300 agrP3-GLuc over time. (a) Bioluminescence output. (b) Bacterial growth (OD600). Data are representative of three technical repeats and three biological repeats for each sample with error bars displaying standard deviations.
Figure 2.
Bioluminescence from S. aureus strain USA300 agrP3-GLuc over time. (a) Bioluminescence output. (b) Bacterial growth (OD600). Data are representative of three technical repeats and three biological repeats for each sample with error bars displaying standard deviations.

Figure 3.
Impact of the addition of exogenous AIP-1 on bioluminescence produced by S. aureus USA300 Δagr P3-Gluc pSKermP2 agrAC. Δagr represents USA300 Δagr P3-GLuc and Δagr + AC represents USA300 Δagr P3-Gluc pSKermP2 agrAC. The data represent three technical repeats and three biological repeats for each sample and error bars displaying standard deviation. * p < 0.01.
Figure 3.
Impact of the addition of exogenous AIP-1 on bioluminescence produced by S. aureus USA300 Δagr P3-Gluc pSKermP2 agrAC. Δagr represents USA300 Δagr P3-GLuc and Δagr + AC represents USA300 Δagr P3-Gluc pSKermP2 agrAC. The data represent three technical repeats and three biological repeats for each sample and error bars displaying standard deviation. * p < 0.01.

Figure 4.
Impact of Ala5(AIP-1) on bioluminescence output from S. aureus USA300 agrP3-GLuc. The 1% v/v DMSO represents USA300 agrP3 GLuc with 1% DMSO, Δagr represents USA300 Δagr P3-GLuc and 100 nM Ala-5 represents USA300 agrP3 GLuc with addition of 100 nM (Ala5)AIP-1. The data shown are the means with error bars displaying standard deviation of triplicate technical repeats and is representative of biological triplicate repeats. * p < 0.01.
Figure 4.
Impact of Ala5(AIP-1) on bioluminescence output from S. aureus USA300 agrP3-GLuc. The 1% v/v DMSO represents USA300 agrP3 GLuc with 1% DMSO, Δagr represents USA300 Δagr P3-GLuc and 100 nM Ala-5 represents USA300 agrP3 GLuc with addition of 100 nM (Ala5)AIP-1. The data shown are the means with error bars displaying standard deviation of triplicate technical repeats and is representative of biological triplicate repeats. * p < 0.01.

Figure 5.
Impact of the AIP biosynthesis inhibitor ambuic acid (40 µM) on bioluminescence output from S. aureus USA300 agrP3-GLuc. ‘No compound’ represents USA300 agrP3 GLuc without inhibitor, Δagr represents USA300 Δagr P3-GLuc without inhibitor and ambuic acid represents USA300 agrP3 GLuc with the addition of 40 µM ambuic acid. The graph displays the mean of three technical and three biological repeats with error bars displaying standard deviation. * p < 0.01.
Figure 5.
Impact of the AIP biosynthesis inhibitor ambuic acid (40 µM) on bioluminescence output from S. aureus USA300 agrP3-GLuc. ‘No compound’ represents USA300 agrP3 GLuc without inhibitor, Δagr represents USA300 Δagr P3-GLuc without inhibitor and ambuic acid represents USA300 agrP3 GLuc with the addition of 40 µM ambuic acid. The graph displays the mean of three technical and three biological repeats with error bars displaying standard deviation. * p < 0.01.

Figure 6.
Comparison of the effect of 3-oxo-C12-HSL, C12-TMA and C12-TOA on S. aureus constitutively expressing bioluminescence based on (a) GLuc or (b) Lux. The data shown are the means with error bars displaying standard deviation of triplicate technical repeats and are representative of biological triplicate repeats. ** p < 0.05.
Figure 6.
Comparison of the effect of 3-oxo-C12-HSL, C12-TMA and C12-TOA on S. aureus constitutively expressing bioluminescence based on (a) GLuc or (b) Lux. The data shown are the means with error bars displaying standard deviation of triplicate technical repeats and are representative of biological triplicate repeats. ** p < 0.05.

Figure 7.
The effect of 3-oxo-C12-HSL, C12-TMA and C12-TOA on agrP3-GLuc expression in USA300 agrP3 GLuc. For 3-oxo-C12-HSL, low luminescence values of 1.9 × 104 RLU/OD600 (5 µM) and 2.45 × 104 RLU/OD600 (10 µM) were recorded. The data show the means with error bars displaying standard deviation of triplicate technical repeats and are representative of biological triplicate repeats. *** p < 0.0001.
Figure 7.
The effect of 3-oxo-C12-HSL, C12-TMA and C12-TOA on agrP3-GLuc expression in USA300 agrP3 GLuc. For 3-oxo-C12-HSL, low luminescence values of 1.9 × 104 RLU/OD600 (5 µM) and 2.45 × 104 RLU/OD600 (10 µM) were recorded. The data show the means with error bars displaying standard deviation of triplicate technical repeats and are representative of biological triplicate repeats. *** p < 0.0001.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bacterial strains used in this study.
| Strain | Description | Reference |
|---|---|---|
| S. aureus 8325-4 | Derivative of S. aureus NCTC 8325 | [32] |
| S. aureus RN4220 attB2 | Cloning intermediate | [33] |
| S. aureus USA300 JE2 | Plasmid-cured derivative of the CA-MRSA strain USA300 LAC | [34,35] |
| S. aureus RN4220 pXyl/tet: SpA leader: GLuc | Constitutively expressing SpA leader:GLuc | This study |
| S. aureus RN4220 Pxyl/tet:::luxABCDE | Constitutively bioluminescent strain | [20] |
| S. aureus USA300 agrP3 Gluc | USA300 with chromosomal integration of SpA leader and GLuc under the control of agrP3 | This study |
| S. aureus USA300 Δagr P3 Gluc | USA300 Δagr with chromosomal integration of SpA leader and GLuc under the control of agrP3. | This study |
| S. aureus ROJ143 | ROJ143 carrying plasmid pSKermP2 agrC1 agrA | [36] |
| S. aureus USA300 Δagr P3 Gluc pSKermP2 agrC1 agrA | USA300 Δagr P3 Gluc carrying pSKermP2 agrC1 agrA | This study |
| E. coli DC10B | Cloning intermediate dc | [37] |
Table 2.
Primers used in this study. All primers were sourced from Sigma Aldrich (UK).
| Primer | Sequence | Function | Reference |
|---|---|---|---|
| CT12 (F) | CTAGTAGGAGGAAAAACATATGATGACTTTACA | Amplification of SpA:GLuc | This study |
| CT13 (R) | ATTTGTCGACCTCAGGAGAGCGTTCACC | ||
| CT14 (F) | AGTGAATTCCCGGGGATCCGACACGTCGACCCTCACTG | Amplification of agrP3 | This study |
| CT15 (R) | CTCCTACTAGCCATCACATCTCTGTGATCTAG | ||
| CT16 (F) | GATGTGATGGCTAGTAGGAGGAAAAACATATGATG | Amplification of SpA:GLuc | This study |
| CT17 (R) | TCGATAAGCTTGGCTGCAGGATTTGTCGACCTCAGGAG | ||
| OU9R10 | CATACTACATATCAACGAAATCAG | Forward primer at RN4220 attB2 integration site | [33] |
| SCV4 | ACCCAGTTTGTAATTCCAGGAG | Reverse primer at 5′ end of pLL102 to RN4220 attB2 | [33] |
| SCV8 | GCACATAATTGCTCACAGCCA | Forward primer at 3′ end of pLL102 to RN4220 attB2 | [33] |
| OU9R7 | ATGGGTGGTAAAACACAAATTTC | Reverse primer at RN4220 attB2 integration site | [33] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Blower, I.; Tong, C.; Sun, X.; Murray, E.; Luckett, J.; Chan, W.; Williams, P.; Hill, P. Gaussia Luciferase as a Reporter for Quorum Sensing in Staphylococcus aureus. Sensors 2020, 20, 4305. https://doi.org/10.3390/s20154305
AMA Style
Blower I, Tong C, Sun X, Murray E, Luckett J, Chan W, Williams P, Hill P. Gaussia Luciferase as a Reporter for Quorum Sensing in Staphylococcus aureus. Sensors. 2020; 20(15):4305. https://doi.org/10.3390/s20154305
Chicago/Turabian StyleBlower, Isobel, Carmen Tong, Xiaohui Sun, Ewan Murray, Jeni Luckett, Weng Chan, Paul Williams, and Philip Hill. 2020. "Gaussia Luciferase as a Reporter for Quorum Sensing in Staphylococcus aureus" Sensors 20, no. 15: 4305. https://doi.org/10.3390/s20154305
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.