
Limobrevibacterium gyesilva gen. nov., sp. nov., Isolated from Forest Soil

Department of Life Science, College of Natural Sciences, Kyonggi University, Suwon 16227, Gyeonggi-do, Republic of Korea
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(5), 684; https://doi.org/10.3390/d15050684
Submission received: 8 April 2023
/
Revised: 9 May 2023
/
Accepted: 15 May 2023
/
Published: 19 May 2023
(This article belongs to the Special Issue Feature Papers in Microbial Diversity and Culture Collections)
Abstract
:An aerobic, oxidase-positive, catalase-negative, non-motile, oval/rod-shaped, and Gram-negative strain RN2-1T was isolated from a Korean forest soil sample. Cells grew in a pH range of 6–9 (optimum range: 7–8), in a temperature range of 20–40 °C (optimum range: 30–35 °C), and in the presence of 0–1% (w/v) NaCl. The major polar lipid profile consisted of phosphatidylglycerol, diphosphatidyl glycerol, phosphatidylethanolamine, phosphatidylcholine, and a group of phospholipids, aminolipids, and polar lipids with unidentified structures. The major quinone was ubiquinone 10. The main fatty acids were summed feature 8 (C18:1 ω7c/C18:1 ω6c), C16:0, Cyclo-C17:0, cyclo C19:0 ω8c, and C18:1 2OH. Phylogenetic similarities based on 16S rDNA sequences showed that strain RN2-1T grouped in the family Acetobacteraceae included the genera Rhodovastum, Acidisoma, Acidisphaera, and Acidibrevibacterium. The aerobic phototrophic bacterium Rhodovastum atsumiense G2-11T was the closest relative to RN2-1T (96.04% sequence similarity). Genomic DNA revealed 68.7% G+C content. Whole genome analysis and phenotypic data of the strain RN2-1T confirmed its distinctive phylogenetic location within the family Acetobacteraceae. Because of these results, strain RN2-1T represents the novel species of a new genus for which the name Limobrevibacterium gyesilva gen. nov., sp. nov. was proposed. The type of strain was strain RN2-1T (KACC 23001 = JCM 35944).
1. Introduction
The family Acetobacteraceae was established in 1980 [1] and is involved in the order Rhodospirillales in the class Alphaproteobacteria. The family Acetobacteraceae presently comprises 46 genera with validly published names (https://lpsn.dsmz.de/family/acetobacteraceae (accessed on 3 April 2023)) [2]. Based on single-strand conformation polymorphism analysis, the 16S-SSCP profiles of DNA from Cladonia arbuscula indicated that members of the family Acetobacteraceae had a close phylogenetic relationship to the genera Gluconacetobacter and Acidisphaera, as well as to the species Rhodovastum atsumiense [3,4].
The habitats of genera in the family Acetobacteraceae were considerably diverse, and some members were characterized by the presence of bacteriochlorophyll a [4,5,6]. The genera of the acidophilic group were isolated from acidic mine drainage [7], acidic hot springs, mine drainage [6], acidic-dominated tundra, and wetlands [8]. A number of representatives of this family have been found to be components of symbiotic microbial communities of plants, such as sugarcane [9], flowers [10,11,12], and the thalli of lichens [4]. The desert [13,14], hot springs [15,16], and air-conditioning system [17] were no exception, as well as some soil such as sediment [18] and paddy soil [5]. Acetobacteraceae could be classified into two groups: acetate-producing bacteria and non-acetate-producing bacteria [7]. The acetate-producing group is obligated with aerobic bacteria within the family Acetobacteracea and has two crucial enzymes of alcohol, dehydrogenase, and aldehyde dehydrogenase that oxidize sugars, sugar alcohols, and ethanol to release the corresponding metabolites (aldehydes, ketones, and organic acids) into the media. These bacteria were mostly members of Acetobacter, Gluconacetobacter, and Gluconobacter and were used commercially in beverage production and food fermentation. The non-acetate-producing group mainly included acidophilic or neutrophilic bacteria, which are widely used in the bioprocessing of minerals and bioremediation due to their versatile metabolisms. These bacteria phylogenetically formed an independent lineage in the family Acetobacteraceae, such as Acidiphilium, Acidocella, Acidicaldus, Acidisphaera, and Acidisoma [7].
In this study, we reported the polyphasic features of strain RN2-1T isolated from a forest soil sample in Seongnam, Gyeonggi-do, Republic of Korea, and classified as a member of the acetate-producing group with the presence of enzyme alcohol dehydrogenase. Therefore, strain RN2-1T can be considered to represent a novel species of a new genus within the family Acetobacteraceae on the basis of its phenotypic, chemotaxonomic, and genotypic features.
2. Material and Methods
2.1. Isolation and Ecology
Strain RN2-1T was isolated from the forest soil at Sangjeok-dong, Sujeong-gu, Seongnam, Gyeonggi-do, Republic of Korea (37°24′52.6″ N 127°02′29.4″ E). Isolation was performed using a culture method modified with a six-transwell plate (Corning Inc., Corning, NY, USA) on 1% R2A and soil extract (1% R2A-SE). To prepare SE, 400 g of the air-dried garden soil was sterilized in 1000 mL DW for one hour at 121 °C and was allowed to sediment for a few hours at room temperature. The supernatant was centrifuged, and 0.032 g R2A per 1000 mL was added before adjusting the pH to 7.5 and sterilizing. A debris-free sieved soil sample (3 g) was placed on the bottom of the transwell plate, and 3 mL of 1% R2A-SE was added. Next, 100 µL of the soil suspension was added to the insert. Then, the plate was kept in a shaking incubator at 130 rpm at 28 °C for 4 weeks. After incubation, 100 μL of each diluted culture was spread on a 1% R2A-SE agar plate. Each colony was picked and streaked separately on 1% R2A-SE plates until pure colonies were grown. The pure isolates were then subcultured on 50% to 100% R2A and were stored at −70 °C in R2A broth, including 20% (v/v) glycerol. Strain RN2-1T was deposited at the KACC and JCM (under KACC 23001 and JCM 35944) for the purpose of systematic research. Rhodovastum atsumiense KCTC 5708, Lichennicoccus roseus KCTC 72321, Acidibrevibacterium fodinaquatile KCTC 62275, Acidisphaera rubrifaciens JCM 10600, Acidisoma tundrae DSM 19999 were provided from the Korean Collection for Type Cultures (KCTC), Japan Collection of Microorganisms (JCM) and Deutsche Sammlung von Mikroorganismen und Zellkulturen (DSMZ), respectively. They were used as reference organisms.
2.2. Physiology and Chemotaxonomy
The morphology of colonies and cells was observed after cultivation on R2A agar plates at 30 °C for 3 days. Cell morphology was visualized by light microscopy (BX50; Olympus) and transmission electron microscopy (Bio-TEM, Hitachi, H-7650). Gram staining was performed by Hucker’s method [19]. Mobile ability was observed in an R2A medium containing 0.4% agar after 72–96 h at 30 °C. The range of temperature for growth was determined at different temperatures (4, 10, 15, 20, 25, 28, 30, 32, 35, 37, 40, 41, 42, and 45 °C) on R2A for 7 days. The range of pH for growth was obtained at 30 °C in R2A broth and was adjusted in the range of pH 3–12 (pH increase by 0.5) using citrate/NaH2PO4 buffer (pH 3.0–5.5), Sorensen’s phosphate buffer (pH 6.5–8), Tris buffer (pH 8.5–9), carbonate buffer (pH 9.5–10.0) and 5 M NaOH (pH 10.5–11) [20]. NaCl tolerance was measured in R2A supplemented with increasing NaCl concentrations (0 to 7.0%) in 0.5% increments during a 7-day incubation at 30 °C. Cellular growth at various pH values and salinity levels was monitored by the optical density at 600 nm.
Carbon source utilization was determined in test tubes filled with a modified MM2V (Table S1) broth medium without adding glucose, to which the different carbon sources were filter-sterilized and supplemented to a final concentration of 1.0 g/L. The control also was prepared in the same way but without a substrate. The nitrogen-fixing activity was determined in a modified MM2V broth medium in which (NH4)2SO4 and glucose were replaced with glutamine and peptone, respectively, through H2 gas production in test tubes with Durham tubes as being positive [21]. The anaerobic growth of cells was tested on an R2A agar at 30 °C for 10 days using the BD GasPak EZ Gas Generating Pouch System under the darkness or the light at 2000 lux (Multi Thermo Chamber HB-305S). Acetone/methanol (7:2, v/v) extracts of RN2-1T cell pellets were scanned within the rank of 300 and 800 nm with a slit of 10 nm on a spectrophotometer (Biochrom-libra-s4+) to find the absorption spectra [7,22]. The activity of oxidase was tested using 1% (w/v) tetramethyl-p-phenylenediamine. The activity of catalase was examined based on gas bubbles after adding a drop of 3% (v/v) hydrogen peroxide (H2O2). The hydrolysis of casein and starch was tested as described by Tindall et al. [23], and Tween 80 hydrolysis was estimated by the method of Smibert and Krieg [24]. Nitrate reduction, indole production, gelatin and aesculin hydrolysis, and the assimilation of different substrates were examined with API 20NE strips (bioMérieux). Various enzyme activities and acid formation from sugar were further tested using API ZYM test strips (bioMérieux). Using both kits, API 20NE and API ZYM were performed in accordance with the manufacturer’s instructions.
For the determination of respiratory quinones and polar lipids, cells of strain RN2-1T that were grown on R2A plates were harvested, freeze-dried, and used after 5 days of growth. Respiratory quinones were extracted with methanol and petroleum ether from 100 mg of freeze-dried cells, and quinone purification was performed according to the method of Minnikin [25]. Then, the pure quinone was analyzed by a reversed-phase HPLC system [solvent MeOH/isopropanol (7:5, v/v), 1.0 mL min−1 flow rate, 270 nm wavelength] [25]. Polar lipids were extracted according to the protocol of Bligh and Dyer [26]. The total lipids profiles were stained with 10% (w/v) ethanolic molybdatophosphoric acid (Sigma-Aldrich), and amino lipids, phospholipids, and glycolipids were discovered with 0.4% (w/v) solution of ninhydrin (Sigma Life Science) in butanol with a Zinzadze reagent (molybdenum blue spray reagent, 1.3%; Sigma Life Science), and α-naphthol reagent (0.5%, w/v), respectively.
2.3. 16S rRNA Phylogeny
The 16S rRNA gene of strain RN2-1T was PCR-amplified from genomic DNA using the primers 27F and 1492R, as mentioned by Frank et al. [27]. Then, the 16S rRNA gene was sequenced and compiled with the EzBioCloud database (www.ezbiocloud.net/eztaxon (accessed on 15 March 2023)) [28] to find the closest phylogenetic neighbors. The phylogenetically closest neighbors were found from the NCBI GenBank database (www.ncbi.nlm.nih.gov/ (accessed on 15 March 2023)) search using 16S rRNA gene sequences. Multiple alignments of sequence data were carried out using the SILVA aligner (www.arb-silva.de/aligner/ (accessed on 16 March 2023)). Three major phylogenetic trees (neighbor-joining (NJ), maximum-likelihood (ML), and maximum-parsimony (MP)) were rebuilt using mega version 7.0.26 software [29]. Bootstrap analyses were performed based on 1000 sample replications [30], and the evolutionary distances were determined according to the Kimura two-parameter method [31].
2.4. Genome Features
Genomic DNA was extracted using DNeasy Blood and Tissue kits (Qiagen, Hilden, Germany). The whole genome shotgun sequencing of strains was performed by Macrogen (Seoul, Republic of Korea) using the Illumina HiSeq platform and assembled using SPAdes version 3.13.0 [32]. To analyze the evolutionary divergence according to whole-genome sequences, a phylogenomic tree of strain RN2-1T and the species of 15 genera within the family Acetobacteraceae from the NCBI (www.ncbi.nlm.nih.gov/ (accessed on 17 March 2023)) was rebuilt in silico with the concatenated alignment of 92 core genes using the UBCG pipeline [33]. The DNA G+C content and average nucleotide identity (ANI) between the whole genome sequence of strain RN2-1T and two closely related genera Rhodovastum atsumiense G2-11T and Acidibrevibacterium fodinaquatile G45-3T were estimated using the OrthoANIu algorithm (www.ezbiocloud.net/tools/ani (accessed on 20 March 2023) [34]. The digital DNA–DNA hybridization (dDDH) values were obtained through the GGDC web server (http://ggdc.dsmz.de/ggdc.php (accessed on 7 April 2023)) [35]. The genome sequence of the target strain was annotated by Rapid Annotation utilizing Subsystem Technology (RAST) server, version 2.0 [36]. The COG (Clusters of Orthologous Group) functional categories were performed by searching the KEGG (Kyoto Encyclopedia of Genes and Genomes) database [37]. The annotation and analysis of secondary metabolite biosynthesis genes were carried out using anti-SMASH server version 5.0 [38]. Finally, a genome map of strain RN2-1T was generated using the CGView Server (http://cgview.ca (accessed on 25 March 2023)) [31].
3. Results and Discussion
3.1. Isolation of Uncultured Strain
General isolation steps of bacterial strains proceed in the following order using conventional media: (1) enrichment culture, (2) serial dilution, (3) plating, and (4) pure culture. However, this study introduced a modified method using a six-transwell plate and a diluted R2A plus soil extract medium to isolate uncultured soil bacteria, including the strain RN2-1T for enrichment culture as well as plating with the same medium.
3.2. Physiology and Chemotaxonomy
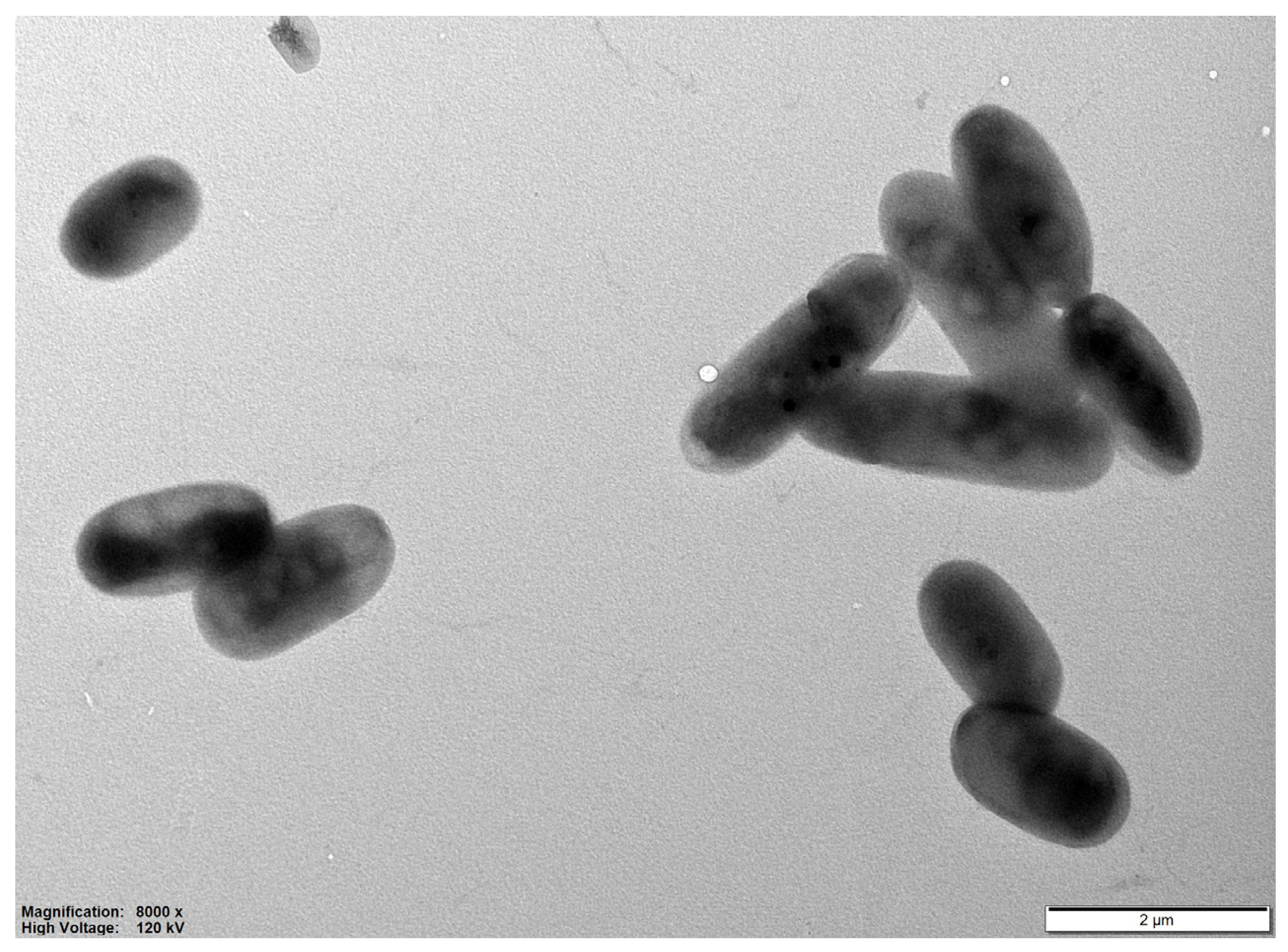
On medium agar R2A, the colonies color of strain RN2-1T varied from silver watercolor to white, circular, convex, slimy, and smooth surface after 5 day-incubation at 30 °C. Strain RN2-1T was Gram-stain-negative, non-spore-forming, non-motile, ovals and rods, 1.5–3.2 µm long and 0.8–1.1 µm wide (Figure 1). The temperature range of growth was 20–40 °C (optimum: 30–35 °C), and the pH range for growth was 6–9 (optimum: 7–8). Strain RN2-1T grew at 0–1% (w/v) NaCl. The growth of the strain was inhibited by NaCl concentrations higher than 1%.
For the utilization of carbon sources, the optimal growth of RN2-1T was observed on casamino acid, glutamine, pyruvate, and citrate; other usable carbon sources were asparagine, peptone, ethanol, propanol, alanine, fumarate, succinate, gluconate, malate, malic acid; little or ambiguous growth was observed with mannose, sorbitol, glycerol, methanol, butanol, sucrose, glucose, L-arabinose, and D-mannitol; acetate, lactate, aspartate, glutamate, formate, propionate, butyrate, and benzoate were not utilized (Table 1 and Table S2). In API ZYM strip tests, the production of alkaline phosphatase, esterase (C4), leucine arylamidase, acid phosphatase, and naphthol-AS-BI-phosphate was positive. RN2-1T could not carry out nitrogen fixation because there was no observation of H2 gas production using glutamine as the sole nitrogen source. The strain was not able to grow under anaerobic conditions with or without light. Bacteriochlorophyll a was not detected under the experimental conditions. Oxidase-positive and catalase-negative activities were observed. Nitrate was converted to nitrite by reduction, but indole production and glucose fermentation did not occur. The hydrolyses of starch, Tween 80, casein, arginine, urease, aesculin, and gelatin were all negative. The differences in the physiological and chemotaxonomy of strain RN2-1T with close phylogenetically related species showed that the strain RN2-1T was different from its close neighbors (Table 1 and Table S3).
For the determination of the fatty acid content, the cells of strain RN2-1T, Rhodovastum atsumiense KCTC 5708; Acidisphaera rubrifaciens JCM 10600; Lichennicoccus roseus KCTC 72321; Acidibrevibacterium fodinaquatile KCTC 62275; and Acidisoma tundrae DSM 19999 were grown on the R2A, MSYS, GNYS, MM2V, GNYS and acidosoma medium (DSMZ), respectively and were harvested after 5–7 days of growth. Fatty acids were extracted, saponified, and methylated following the protocol of the Sherlock Microbial Identification System (MIDI) and analyzed by the capillary GC system (Hewlett Packard 6890) using the Microbial Identification software package with the Sherlock system MIDI 6.3 and the Sherlock Aerobic Bacterial Database (TSBA6.21) [39]. The major fatty acids were C16:0, Cyclo-C17:0, cyclo C19:0 ω8c, C18:1 2OH, and summed feature 8 (C18:1 ω7c/C18:1 ω6c) (Table 2 and Table S4). The major polar lipids of strain RN2-1T consisted of phosphatidylglycerol, diphosphatidylglycerol, phosphatidylethanolamine, and phosphatidylcholine. Additionally, the amounts of phospholipids, amino acids, and unknown polar lipids were detected (Figure S1). Similar to the genera Acidisphaera, Lichennicoccus, Acidibrevibacterium, and Acidisoma, the prime quinone of strain RN2-1T was ubiquinone 10; however, there was a difference between the closest genera Rhodovastum, which had both ubiquinone 10 and rhodoquinone 10.
3.3. 16S rRNA Phylogeny
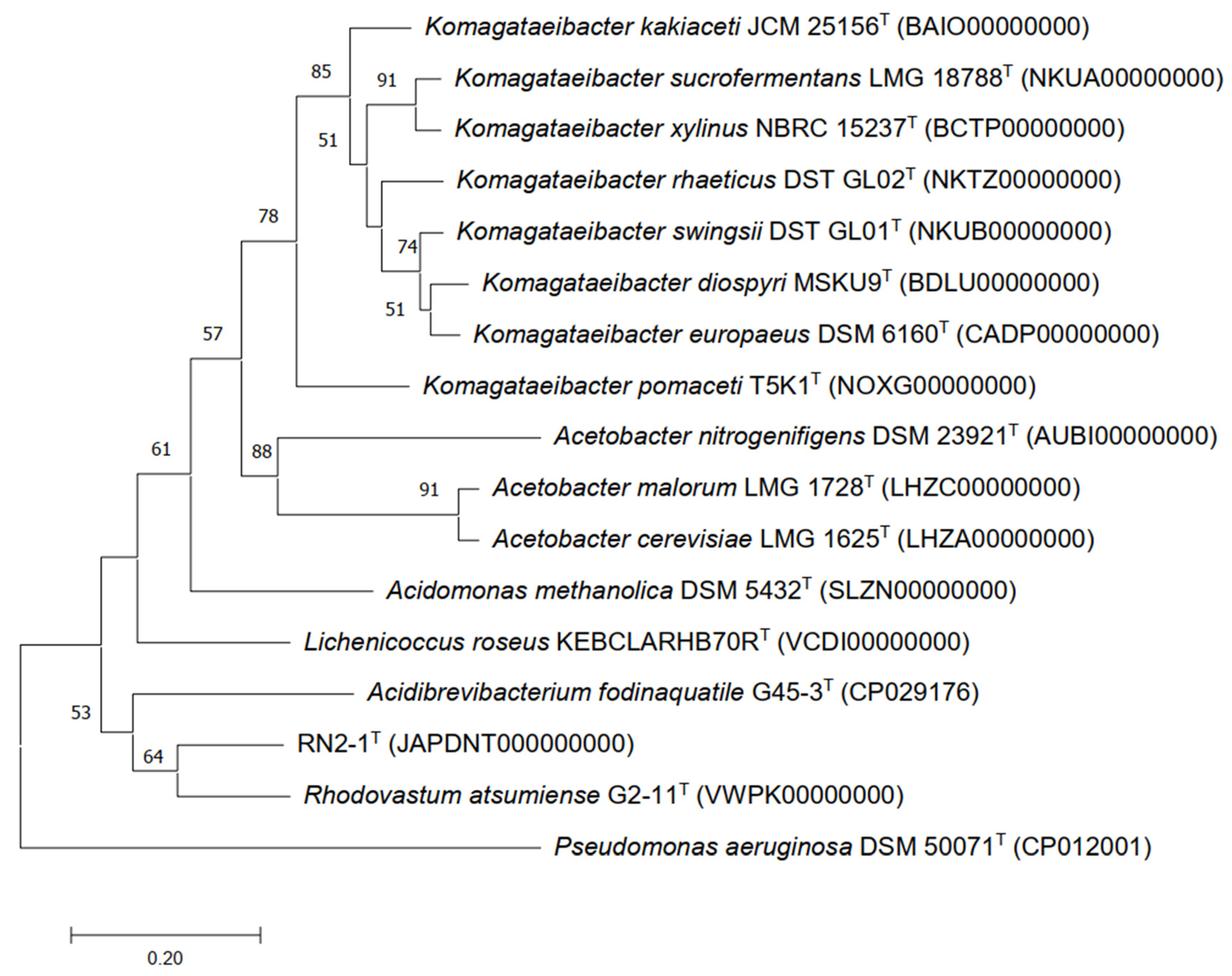
According to 16S rRNA gene sequences, phylogenetic analysis indicated that the novel strain was a member of the family Acetobacteraceae, and its closet type strains were Rhodovastum atsumiense G2-11T and Acidisphaera rubrifaciens HS-AP3T, with 96.04% and 95.39% similarity, respectively. Phylogenetic trees were reconstructed using the NJ algorithm, which displayed that strain RN2-1T was clustered in a group of the genera Rhodovastum, Acidisoma, Acidisphaera, and Acidibrevibacterium in which RN2-1T and phototrophic alphaproteobacterium Rhodovastum atsumiense formed a sub-lineage outside of these genera (Figure 2). The NJ tree was topologically supported by those of the MP (Figure S2) and ML trees (Figure S3).
3.4. Genome Features
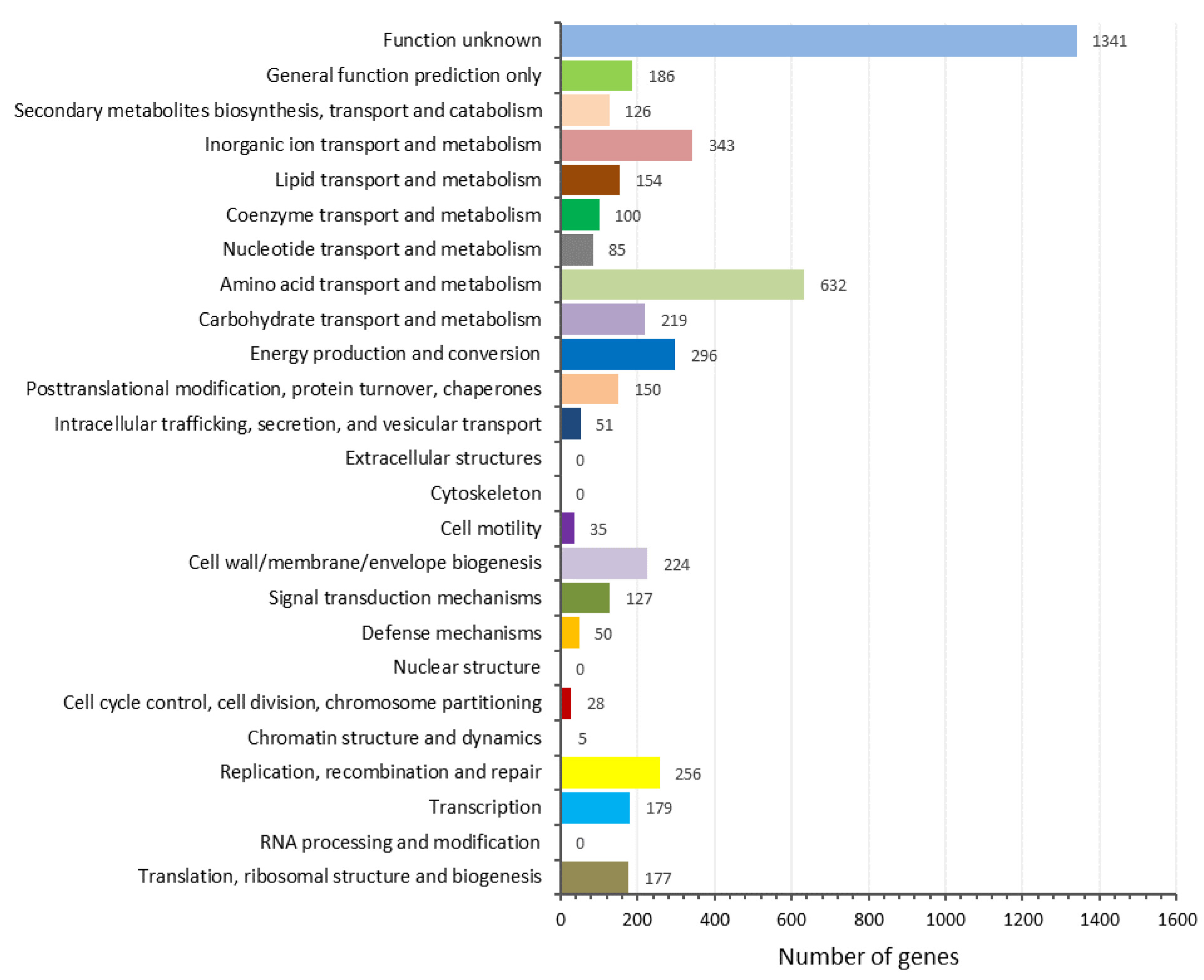
The results indicated that strain RN2-1T also clustered with the genera Rhodovastum and Acidibrevibacterium (Figure 3). The complete genome of strain RN2-1T was 5,373,152 bp in size, the N50 of the filtered reads was 173,752 kb, and the genome coverage was 23.45×. The G+C content of strain RN2-1T was computed as 68.7mol%. Rast analysis revealed the presence of 5057 coding sequences, 53 RNAs, and 448 subsystems in the genome of strain RN2-1T. A total of 5019 protein-coding genes were identified, and 4764 (94.91%) were assigned to COG categories. The COG category assignment of genes to general function prediction was only 3.9% (186 genes), and other known functions included amino acid transport and metabolism (13.3%: 632 genes), inorganic ion transport and metabolism (7.2%: 343 genes), and energy production and conversion (6.2%: 296 genes), whereas 28.1% of genes functioned unknown into the COGs (Figure 4). The function genes showed that RN2-1T might be classified as a member of the acetate-producing group with the presence of alcohol dehydrogenase (ADH) enzymes consisting of the alcohol dehydrogenase genes adh_1, adh_2, adhA_1, adhA_2, and adhT. These proteins, in relation to ADHs, were detected in the genome of strain RN2-1T (Table S5). The genome properties and statistics are summarized in Figure 5.
Pairwise ANI values between strain RN2-1T and Rhodovastum atsumiense G2-11T, and Acidibrevibacterium fodinaquatile G45-3T were 76.82 and 72.70%, respectively, and strain RN2-1T had dDDH values ranging from 16.4 to 21.0% with Rhodovastum atsumiense G2-11T and 13.5 to 18.7% Acidibrevibacterium fodinaquatile G45-3T. The ANI and dDDH values were much lower than the generally accepted species threshold of 95 and 70%, respectively [34,40], indicating that the genomic of RN2-1T was different from the existing members of the family Acetobacteraceae.
4. Conclusions
The phylogenetic trees showed that RN2-1T had a relatively close phylogenetic relationship to phototrophic alphaproteobacterium Rhodovastum atsumiense G2-11T. However, morphologically our isolates were clearly distinguishable from the R. atsumiense G2-11T. R. atsumiense G2-11T was grown under anaerobic light conditions, whereas RN2-1T did not grow under the same conditions. Cells of R. atsumiense G2-11T were capable of motion by means of a single polar flagellum, and the cell extract of R. atsumiense G2-11T had bacteriochlorophyll a and carotenoids, but RN2-1T did not. Additionally, there are vital differences between the isolate and R. atsumiense G2-11T in chemotaxonomic analysis, such as quinone and fatty acid composition. Additionally, by additional phenotypic traits, strain RN2-1T was clearly differentiated from the existing genera, as shown in Table 1 and Table 2. Moreover, pairwise ANI values between the strain RN2-1T and R. atsumiense G2-11T were 76.82, which is much less than the commonly recognized species threshold of 95%. The result of digital DNA–DNA hybridization (dDDH) between strain RN2-1T and R. atsumiense G2-11T of 16.4 to 21.0% showed the genomic differences among them. In summary, strain RN2-1T possessed numerous characteristics that apparently distinguished it from its most closely related phylogenetic neighbor R. atsumiense G2-11T, and other phylogenetically close members of the family Acetobacteraceae. Based on the polyphasic data, as shown above, strain RN2-1T could be considered representative of a novel species of a new genus Limobrevibacterium, for which the name Limobrevibacterium gyesilva gen. nov., sp. nov. is proposed.
Further research may include further genomic studies in detail to find unknown functions with this strain which has 1341 unknown genes (Figure 4).
- Description of Limobrevibacterium gen. nov.
Limobrevibacterium (Li.mo.bre.vi.bac.te′ri.um. limosa slimy; brevis short; bacterium a small rod; Limobrevibacterium, a genus of short rod-shaped slimy bacteria).
The cells were non-motile, non-spore-forming, and Gram-stain-negative ovals or rods. BChl a and carotenoids were absent when catalase-negative and oxidase-positive. The main respiratory quinone was ubiquinone 10. The polar lipid profile included diphosphatidylglycerol, phosphatidylglycerol, phosphatidylethanolamine, phosphatidylcholine, and a group of unidentified structures of phospholipids, amino lipids, and polar lipids. The major fatty acids were C16:0, Cyclo-C17:0, cyclo C19:0 ω8c, and C18:1 2OH. The G+C content of the genomic DNA was 68.7%. A novel genus of the family Acetobacteraceae was identified, and the type species was Limobrevibacterium gyesilva RN2-1T.
- Description of Limobrevibacterium gyesilva sp. nov.
Limobrevibacterium gyesilva (gy.o.sil.va. gye Gyeonggi, isolated place; silva from forest land; gyesilva isolated from forest land of Gyeonggi).
The description is the same for the genus, with the following additional characteristics. The cells were 0.8–1.1 μm wide and 1.5–3.2 μm long. Colonies varied from silver watercolor to a white, circular, convex, slimy, and smooth surface. The minimum and maximum temperatures for growth were 20 °C and 40 °C, and this grew optimally at 30–35 °C. The optimum pH range for growth is 7–8. Does not grow anaerobically with or without light. Nitrogen fixation could not be carried out. Nitrate was converted to nitrite by reduction. Starch, casein, gelatin, and Tween 80 were not hydrolyzed. The carbon sources optimal for growth included casamino acid, glutamine, pyruvate, and citrate. Additional carbon sources that induced growth were asparagine, peptone, ethanol, propanol, alanine, fumarate, succinate, gluconate, malate, and malic acid. Little or ambiguous growth occurred with mannose, sorbitol, glycerol, methanol, butanol, sucrose, glucose, L-arabinose, and D-mannitol. No growth was observed on acetate, lactate, aspartate, glutamate, formate, propionate, butyrate, and benzoate. The enzyme activities included alkaline phosphate, esterase, leucine arylamidase, acid phosphatase, and Naphthol-AS-BI- phosphate (API ZYM test).
The type of strain was RN2-1T (=KCTC 23001 = JCM 35944), which was isolated from the forest soil at Sangjeok-dong, Sujeong-gu, Seongnam, and Gyeonggi-do, Republic of Korea. The DNA G+C content of the type of strain was 68.7 mol%. GenBank accession numbers for sequences of the strain RN2-1T were JAPDNT000000000 (genome) and PRJNA892442 (16S rRNA gene nucleotide sequence).
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/d15050684/s1, Figure S1: Polar lipid profile of strain RN2-1T; Figure S2: Phylogenetic tree reconstructed with the Minimum Evolution method based on 16S rRNA gene sequences of strain RN2-1T and type species of the family Acetobacteraceae; Figure S3: Phylogenetic tree reconstructed with the Maximum Likelihood method based on 16S rRNA gene sequences of strain RN2-1T and type species of the family Acetobacteraceae; Table S1: Composition of modified MM2V medium; Table S2: Results from API ZYM, API 20NE test; Table S3: Results from carbon source utilization;Table S4: Detailed cellular fatty acid profiles (% of totals) of strain RN2-1T and closely related reference strains; Table S5: Alcohol dehydrogenase proteins in RN2-1T genome.
Author Contributions
N.L.T.T. envisioned, designed, and performed all the experiments. N.L.T.T. and J.K. interpreted all the data, and checked, discussed, edited, and approved the final draft of the manuscript. J.K. coordinated and supervised this study. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Kyonggi University Research Grant (2020-036).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gillis, M.; de Ley, J. Intra- and intergeneric similarities of the ribosomal ribonucleic acid cistrons of Acetobacter and Gluconobacter. Int. J. Syst. Bacteriol. 1980, 30, 7–27. [Google Scholar] [CrossRef]
- Parte, A.C. LPSN—List of prokaryotic names withstanding in nomenclature (bacterio.net), 20 years on. Int. J. Syst. Evol. Microbiol. 2018, 68, 1825–1829. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, M.; de Castro, J.V., Jr.; Müller, H.; Berg, G.; Grube, M. In situ analysis of the bacterial community associated with the reindeer lichen Cladonia arbuscula reveals predominance of Alphaproteobacteria. FEMS Microbiol. Ecol. 2008, 66, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Pankratov, T.A.; Grouzdev, D.S.; Patutina, E.O.; Kolganova, T.V.; Berestovskaya, J.J.; Ashikhmin, A.A. Lichenicoccus roseus gen. nov., sp. nov., the first bacteriochlorophyll a-containing, psychrophilic and acidophilic Acetobacteraceae bacteriobiont of lichen Cladonia species. Int. J. Syst. Evol. Microbiol. 2020, 70, 4591–4601. [Google Scholar] [CrossRef] [PubMed]
- Okamura, K.; Hisada, T.; Kanbe, T.; Hiraishi, A. Rhodovastum atsumiense gen. nov., sp. nov., a phototrophic alphaproteobacterium isolated from paddy soil. J. Gen. Appl. Microbiol. 2009, 55, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Hiraishi, A.; Matsuzawa, Y.; Kanbe, T.; Wakao, N. Acidisphaera rubrifaciens gen. nov., sp. nov., an aerobic bacteriochlorophyll-containing bacterium isolated from acidic environments. Int. J. Syst. Evol. Microbiol. 2000, 50, 1539–1546. [Google Scholar] [CrossRef]
- Muhadesi, J.B.; Huang, Y.; Wang, B.J.; Jiang, C.Y.; Liu, S.J. Acidibrevibacterium fodinaquatile gen. nov., sp. nov., isolated from acidic mine drainage. Int. J. Syst. Evol. Microbiol. 2019, 69, 3243–3250. [Google Scholar] [CrossRef]
- Belova, S.E.; Pankratov, T.A.; Detkova, E.N.; Kaparullina, E.N.; Dedysh, S.N. Acidisoma tundrae gen. nov., sp. nov. and Acidisoma sibiricum sp. nov., two acidophilic, psychrotolerant members of the Alphaproteobacteria from acidic northern wetlands. Int. J. Syst. Evol. Microbiol. 2009, 59, 2283–2290. [Google Scholar] [CrossRef]
- Muthukumarasamy, R.; Revathi, G.; Seshadri, S.; Lakshminarasimhan, C. Gluconacetobacter diazotrophicus (syn. Acetobacter diazotrophicus), a promising diazotrophic endophyte in tropics. Curr. Sci. 2002, 83, 137–145. [Google Scholar]
- Yamada, Y.; Katsura, K.; Kawasaki, H.; Widyastuti, Y.; Saono, S.; Seki, T.; Uchimura, T.; Komagata, K. Asaia bogorensis gen. nov., sp. nov., an unusual acetic acid bacterium in the alpha-Proteobacteria. Int. J. Syst. Evol. Microbiol. 2000, 50, 823–829. [Google Scholar] [CrossRef]
- Yukphan, P.; Malimas, T.; Muramatsu, Y.; Takahashi, M.; Kaneyasu, M.; Potacharoen, W.; Tanasupawat, S.; Nakagawa, Y.; Hamana, K.; Tahara, Y.; et al. Ameyamaea chiangmaiensis gen. nov., sp. nov., an acetic acid bacterium in the alpha-Proteobacteria. Biosci. Biotechnol. Biochem. 2009, 73, 2156–2162. [Google Scholar] [CrossRef] [PubMed]
- Yukphan, P.; Malimas, T.; Muramatsu, Y.; Potacharoen, W.; Tanasupawat, S.; Nakagawa, Y.; Tanticharoen, M.; Yamada, Y. Neokomagataea gen. nov., with descriptions of Neokomagataea thailandica sp. nov. and Neokomagataea tanensis sp. nov., osmotolerant acetic acid bacteria of the alpha-Proteobacteria. Biosci. Biotechnol. Biochem. 2011, 75, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.W.; Salam, N.; Hua, Z.S.; Liu, B.B.; Han, M.X.; Fang, B.Z.; Wang, D.; Xiao, M.; Hozzein, W.N.; Li, W.J. Siccirubricoccus deserti gen. nov., sp. nov., a proteobacterium isolated from a desert sample. Int. J. Syst. Evol. Microbiol. 2017, 67, 4862–4867. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.L.; Li, S.; Wei, Q.C.; Lian, W.H.; Lu, C.Y.; Xu, L.; Huang, L.N.; Dong, L.; Li, W.J. Sabulicella rubraurantiaca gen. nov., sp. nov., a new member of the family Acetobacteraceae, isolated from desert soil. Arch. Microbiol. 2021, 204, 1. [Google Scholar] [CrossRef] [PubMed]
- Ming, H.; Duan, Y.Y.; Yin, Y.R.; Meng, X.L.; Li, S.; Zhou, E.M.; Huang, J.R.; Nie, G.X.; Li, W.J. Crenalkalicoccus roseus gen. nov., sp. nov., a thermophilic bacterium isolated from alkaline hot springs. Int. J. Syst. Evol. Microbiol. 2016, 66, 2319–2326. [Google Scholar] [CrossRef]
- Albuquerque, L.; Rainey, F.A.; Nobre, M.F.; da Costa, M.S. Elioraea tepidiphila gen. nov., sp. nov., a slightly thermophilic member of the Alphaproteobacteria. Int. J. Syst. Evol. Microbiol. 2008, 58, 773–778. [Google Scholar] [CrossRef]
- Khan, S.A.; Jeong, S.E.; Jung, H.S.; Quan, Z.X.; Jeon, C.O. Roseicella frigidaeris gen. nov., sp. nov., isolated from an air-conditioning system. Int. J. Syst. Evol. Microbiol. 2019, 69, 1384–1389. [Google Scholar] [CrossRef]
- Kim, W.H.; Kim, D.H.; Kang, K.; Ahn, T.Y. Dankookia rubra gen. nov., sp. nov., an alphaproteobacterium isolated from sediment of a shallow stream. J. Microbiol. 2016, 54, 420–425. [Google Scholar] [CrossRef]
- Lányi, B. Classical and Rapid Identification Method for Medically Important Bacteria. In Method in Microbiology 19; Cowell, R., Ed.; Academic Press: Oak Ridge, TN, USA, 1987; pp. 1–65. [Google Scholar]
- Trinh, N.H.; Kim, J. Paraburkholderia flava sp. nov., isolated from cool temperate forest soil. Int. J. Syst. Evol. Microbiol. 2020, 70, 2509–2514. [Google Scholar] [CrossRef]
- Okamura, K.; Hisada, T.; Hiraishi, A. Characterization of thermotolerant purple nonsulfur bacteria isolated from hot-spring Chloroflexus mats and the reclassification of “Rhodopseudomonas cryptolactis” Stadtwald-Demchick et al. 1990 as Rhodoplanes cryptolactis nom. rev., comb. nov. J. Gen. Appl. Microbiol. 2007, 53, 357–361. [Google Scholar] [CrossRef]
- Le Olson, T.; Meene, A.M.; Francis, J.N.; Pierson, B.K.; Blankenship, R.E. Pigment analysis of “Candidatus Chlorothrix halophila”, a green filamentous anoxygenic phototrophic bacterium. J. Bacteriol. 2007, 189, 4187–4195. [Google Scholar] [CrossRef] [PubMed]
- Tindall, B.J.; Sikorski, J.; Smibert, R.A.; Krieg, N.R. Phenotypic Characterization and the Principles of Comparative Systematics. In Methods for General and Molecular Microbiology; Reddy, C.A., Ed.; ASM Press: Washington, DC, USA, 2007; pp. 330–393. [Google Scholar]
- Smibert, R.M.; Krieg, N.R. Phenotypic characterization. In Methods for General and Molecular Bacteriology; Gerhardt, P., Murray, R.G.E., Wood, W.A., Krieg, N.R., Eds.; American Society for Microbiology: Washington, DC, USA, 1994; pp. 607–654. [Google Scholar]
- Minnikin, D.E.; O’Donnell, A.G.; Goodfellow, M.; Alderson, G.; Athalye, M.; Schaal, A.; Parlett, J. An integrated procedure for the extraction of bacterial isoprenoid quinones and polar lipids. J. Microbiol. Methods 1984, 2, 233–241. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically United database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Soc. Study Evol. 1985, 39, 783–791. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Na, S.I.; Kim, Y.O.; Yoon, S.H.; Ha, S.M.; Baek, I.; Chun, J. UBCG: Up-to-date bacterial core gene set and pipeline for phylogenomic tree reconstruction. J. Microbiol. 2018, 56, 280–285. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef]
- Sasser, M. Identification of bacteria by gas chromatography of cellular fatty acids. Technol. Note 2001, 101. [Google Scholar]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.-W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]
Figure 1.
Transmission electron microscopy of strain RN2-1T growth on R2A medium plates for 3 days at 30 °C. Bar 2 µm. 8000× indicates 8000 times multiplication.
Figure 1.
Transmission electron microscopy of strain RN2-1T growth on R2A medium plates for 3 days at 30 °C. Bar 2 µm. 8000× indicates 8000 times multiplication.

Figure 2.
Phylogenetic tree reconstructed with the Neighbor-joining method based on 16S rRNA gene sequences of strain RN2-1T and type species of the family Acetobacteraceae. Pseudomonas aeruginosa DSMZ 5071T was used as an outgroup. Numbers at branch nodes represent confidence levels (values > 50% are shown) from 1000 replicate bootstrap samplings. GenBank accession numbers are shown in parentheses. Bar, 0.02 substitutions per nucleotide position.
Figure 2.
Phylogenetic tree reconstructed with the Neighbor-joining method based on 16S rRNA gene sequences of strain RN2-1T and type species of the family Acetobacteraceae. Pseudomonas aeruginosa DSMZ 5071T was used as an outgroup. Numbers at branch nodes represent confidence levels (values > 50% are shown) from 1000 replicate bootstrap samplings. GenBank accession numbers are shown in parentheses. Bar, 0.02 substitutions per nucleotide position.

Figure 3.
The phylogenetic tree was reconstructed using the UBCG (concatenated alignment of 92 core genes) pipeline based on whole-genome sequences of strain RN2-1T and the type species within the family Acetobacteraceae. Pseudomonas aeruginosa DSMZ 5071T was used as an outgroup. GenBank accession numbers are given in parentheses. Bar, 0.20 substitutions per nucleotide position.
Figure 3.
The phylogenetic tree was reconstructed using the UBCG (concatenated alignment of 92 core genes) pipeline based on whole-genome sequences of strain RN2-1T and the type species within the family Acetobacteraceae. Pseudomonas aeruginosa DSMZ 5071T was used as an outgroup. GenBank accession numbers are given in parentheses. Bar, 0.20 substitutions per nucleotide position.

Figure 4.
COG functional classification of proteins in strain RN2-1T genome.

Figure 5.
A graphical circular genome map of strain RN2-1T performed by the CGView server (www.cgview.ca (accessed on 25 March 2023)). From the outside to the center: rings 1 and 2 demonstrate the protein-coding genes (CDS) on both forward and reverse strands, ring 3 represents the GC content plot, and ring 4 represents GC skew.
Figure 5.
A graphical circular genome map of strain RN2-1T performed by the CGView server (www.cgview.ca (accessed on 25 March 2023)). From the outside to the center: rings 1 and 2 demonstrate the protein-coding genes (CDS) on both forward and reverse strands, ring 3 represents the GC content plot, and ring 4 represents GC skew.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Phenotypic characteristics that differentiate the novel genus Limobrevibacterium from its closely related genera Rhodovastum, Acidisphaera, Lichennicoccus, Acidibrevibacterium, and Acidosoma. A detailed description of results from the API ZYM, API 20NE test kits, and carbon sources utilized are provided in Tables S2 and S3. Genera: 1, Limobrevibacterium; 2, Rhodovastum (data are from the present study and from Okamura et al. [5]); 3, Acidisphaera [6]; 4, Lichennicoccus [4]; 5, Acidibrevibacterium [7]; 6, Acidisoma [8]. +, Positive; −, negative. For carbon source utilization: ++, good growth; +, moderate growth; +/−, little or ambiguous growth; −, no growth; nd, no data.
Table 1.
Phenotypic characteristics that differentiate the novel genus Limobrevibacterium from its closely related genera Rhodovastum, Acidisphaera, Lichennicoccus, Acidibrevibacterium, and Acidosoma. A detailed description of results from the API ZYM, API 20NE test kits, and carbon sources utilized are provided in Tables S2 and S3. Genera: 1, Limobrevibacterium; 2, Rhodovastum (data are from the present study and from Okamura et al. [5]); 3, Acidisphaera [6]; 4, Lichennicoccus [4]; 5, Acidibrevibacterium [7]; 6, Acidisoma [8]. +, Positive; −, negative. For carbon source utilization: ++, good growth; +, moderate growth; +/−, little or ambiguous growth; −, no growth; nd, no data.
| Characteristic | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Source of isolation | Forest soil | Paddy soil | Sediment from an acidic hot spring and pyritic mine drainage | The thalli of Cladonia arbuscula and Cladonia stellaris lichens | Acidic mine drainage | Acidic northern wetlands |
| Cell shape | Ovals, Rods | Ovals, rods | Cocci, short rods | Spherical to ovoid | Rods | Coccobacilli |
| Cell size (µm) | 0.8–1.1 × 1.5–3.2 | 2.0–3.0 × 3.0–10 | 0.7–0.9 × 0.9–1.6 | 0.8–1.6 × 0.9–2.26 | 1.3–1.5 × 3.2–3.3 | 0.7–1.5 × 1.4–4.1 |
| Colony color | White, Mucoid | Red brown | Salmon pink | pink or salmon pink | Light yellow | White, cream |
| Motility | − | + | − | − | − | − |
| Anaerobic phototrophy | − | + | − | − | − | − |
| Bacteriochlorophyll a | − | + | + | + | − | − |
| Temperature range for growth (°C) | 20–40 | 20–40 | 20–40 | 5–30 | 20–45 | 2–30 |
| Optimal growth temperature (°C) | 30–35 | 30–35 | 20–35 | 10–15 | 37 | 18–22 |
| pH range for growth | 6–9 | 5.0–8.5 | 3.5–6.0 | 3.0–7.5 | 2.5–5 | 3.7–7.5 |
| pH optimum for growth | 7–8 | 6.0–6.5 | 4.5–5.0 | 4.5–5.5 | 4.0 | 5.0–5.7 |
| Tolerance of >2.5 (%w/v) NaCl | − | − | + | + | − | − |
| Electron donor/carbon source | ||||||
| Acetate | − | ++ | − | − | nd | − |
| Lactate | − | + | + | − | nd | + |
| Mannose | +/− | + | − | + | − | + |
| Sorbitol | +/− | + | + | + | + | + |
| Glycerol | +/− | − | + | − | − | + |
| Asparagine | + | + | − | − | + | + |
| Aspartate | − | + | − | nd | + | − |
| Glutamate | − | ++ | − | − | nd | nd |
| Glutamine | ++ | ++ | − | − | nd | nd |
| Citrate | ++ | − | +/− | − | nd | − |
| Nitrogen fixation | − | + | nd | − | nd | nd |
| Major quinone(s) | Q-10 | Q-10, RQ-10 | Q-10 | Q-10 | Q-10 | Q-10 |
| DNA G+C content (mol%) | 68.7 | 67.8 | 69.1–69.8 | 69.1–69.8 | 65.9 | 60.5–61.9 |
Table 2.
Detailed cellular fatty acid profiles (% of totals) of strain RN2-1T and closely related reference strains. Strains; 1, RN2-1T; 2, Rhodovastum atsumiense KCTC5708; 3, Acidisphaera rubrifaciens JCM10600; 4, Lichennicoccus roseus KCTC72321; 5, Acidibrevibacterium fodinaquatile KCTC62275; 6, Acidisoma tundrae DSM19999. All data are obtained from this study. Fatty acids that revealed <0.5% of the total in all strains are not shown; -, not detected.
Table 2.
Detailed cellular fatty acid profiles (% of totals) of strain RN2-1T and closely related reference strains. Strains; 1, RN2-1T; 2, Rhodovastum atsumiense KCTC5708; 3, Acidisphaera rubrifaciens JCM10600; 4, Lichennicoccus roseus KCTC72321; 5, Acidibrevibacterium fodinaquatile KCTC62275; 6, Acidisoma tundrae DSM19999. All data are obtained from this study. Fatty acids that revealed <0.5% of the total in all strains are not shown; -, not detected.
| Fatty Acid | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Saturated | ||||||
| C14:0 | 1.26 | 2.30 | 1.14 | - | - | - |
| C16:0 | 6.54 | 20.47 | 20.07 | 7.82 | 22.22 | 10.33 |
| C18:0 | 1.02 | 2.17 | 1.31 | 2.71 | - | 4.73 |
| 10-methyl C17:0 | - | 0.68 | - | - | - | - |
| Cyclo-C17:0 | 5.62 | - | - | - | 1.06 | - |
| Unsaturated | ||||||
| C16:1 ω5c | 1.25 | 2.43 | - | - | - | - |
| C18:1 ω5c | - | 0.74 | - | 0.62 | - | - |
| C18:1 ω9c | - | - | - | - | - | 3.33 |
| C20:1 ω7c | - | 0.58 | - | - | - | |
| cyclo C19:0 ω8c | 7.85 | 0.57 | 20.80 | 1.25 | 38.51 | |
| Hydroxy | ||||||
| C16:0 3OH | - | - | 0.68 | 1.27 | - | - |
| C18:0 3OH | 0.68 | 1.56 | 0.81 | 1.64 | - | 9.72 |
| C18:1 2OH | 6.98 | 0.81 | 10.98 | 16.60 | - | 3.62 |
| Summed features * | ||||||
| 2 | - | 3.51 | - | 2.33 | 2.00 | 1.26 |
| 3 | 7.65 | 9.69 | 0.58 | 0.51 | 0.72 | - |
| 8 | 61.14 | 51.84 | 43.64 | 62.50 | 2.97 | 28.50 |
* Summed features represent two or three fatty acids that could not be separated by the Microbial Identification System. Summed feature 2 consisted of C16:1 iso I/C14:0 3OH; summed feature 3 consisted of C16:1 ω7c/C16:1 ω6c summed feature 8 consisted of C18:1 ω7c/C18:1 ω6c.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tuyet, N.L.T.; Kim, J. Limobrevibacterium gyesilva gen. nov., sp. nov., Isolated from Forest Soil. Diversity 2023, 15, 684. https://doi.org/10.3390/d15050684
AMA Style
Tuyet NLT, Kim J. Limobrevibacterium gyesilva gen. nov., sp. nov., Isolated from Forest Soil. Diversity. 2023; 15(5):684. https://doi.org/10.3390/d15050684
Chicago/Turabian StyleTuyet, Nhan Le Thi, and Jaisoo Kim. 2023. "Limobrevibacterium gyesilva gen. nov., sp. nov., Isolated from Forest Soil" Diversity 15, no. 5: 684. https://doi.org/10.3390/d15050684
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.