
The Genus Allochrusa: A Comprehensive Review of Botany, Traditional Uses, Phytochemistry, and Biological Activities
 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
3. Taxonomy and Diversity

{kind=link}
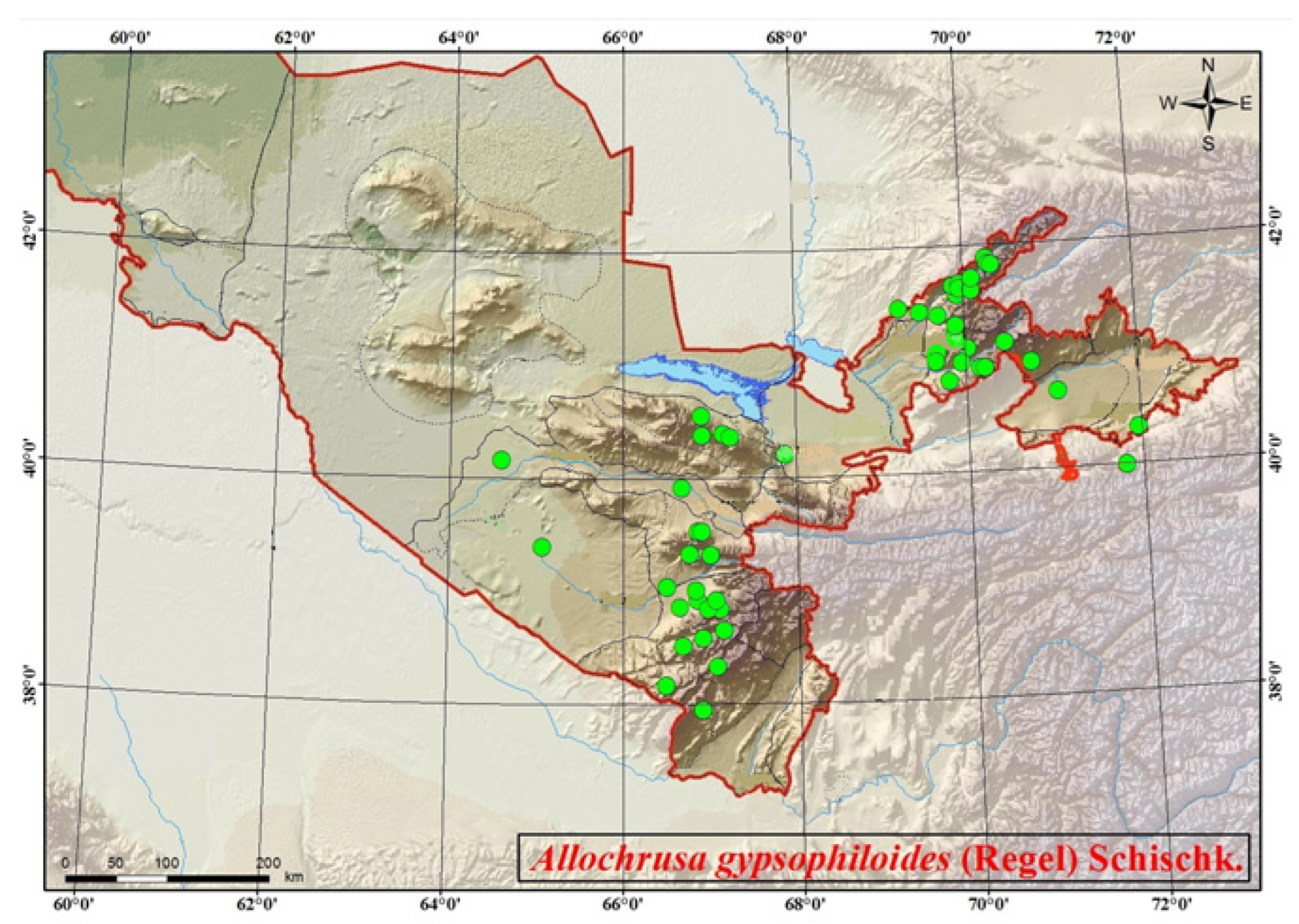{kind=link}
| Species | Distribution | Region | References |
|---|---|---|---|
| A. takhtajanii, A. bungei, A. versicolor | Armenia | Surenavan, Urts Mountains, Transcaucasus | [10]; gbif.org |
| A. versicolor | Azerbaijan | South to north-west Transcaucasus | [10] |
| A. bungei, A. lutea, A. persica, A. tadshikistanica, A. versicolor | Iran | Tabriz, Marand and Jolfa, north-west of Iran | [11] |
| A. gypsophiloides, A. paniculata | Kazakhstan | Karatau, Mount Kazigurt, Aksu-Zhabagly, South Kazakhstan | [17] |
| A. gypsophiloides, A. paniculata | Kyrgyzstan | Chatkal, Talas, and Pskem ranges; Talas Ala-Too, Kyrgiz Ala-Too | [17,18] |
| A. gypsophiloides, A. transhyrcana | Turkmenistan | Amu Darya river | [10,18] |
| A. gypsophiloides, A. tadshikistanica, A. paniculata | Tajikistan | Pamir mountains | [10,21] |
| A. gypsophiloides | Uzbekistan | Chatkal, Hissar, Kurama, Kuhitang ranges (Dzhizak, Kashkadarya, Namangan, Samarkand, Surkhandarya, Tashkent regions) | [17,20] |
| A. paniculata | Andijan, Fergana regions | [20] | |
| A. tadshikistanica | Chatkal, Hissar, Kurama ranges (Kashkadarya, Namangan, Surkhandarya, Tashkent regions) | [22] |
4. Traditional Uses
5. Medicinal Uses
6. Phytochemical Compounds
6.1. Triterpene glycosides
6.2. Ecdysteroids
6.3. Flavonoids
| Plant Sources | Name | Formula | Structure | References | ||
|---|---|---|---|---|---|---|
| Triterpene glycosides | ||||||
 | ||||||
| A. gypsophiloides | Gypsoside (1) | C80H126O44 | R1 = -β-D-GlcUA- |  | 4←1-β-D-Glc -4←1-β-D-Gal 3←1-α-L-Ara | [35] |
| R2 = -α-L-Rha- |  | 4←1-β-D-Fuc-3←1-β-D-Xyl 2←1-β-D-Xyl-3←1-β-D-Xyl | ||||
| R3 = -H | ||||||
| A. gypsophiloides | Acanthophylloside B (2) | С86H136O48 | R1 = -β-D-GlcUA- |  | 4←1-α-L-Ara -4←1-β-D-Gal 2←1-β-D-Gal | [36,37,38,39,41,42,54] |
| R2 = -β-D-Fuc- |  | 4←1-α-L-Rha-4←1-β-D-Xyl-3←1-β-D-Xyl-3←1-β-D-Xyl 2←1-α-D-Qui | ||||
| R3 = -H | ||||||
| A. gypsophiloides | Acanthophylloside C (3) | С92H146O53 | R1 = -β-D-GlcUA- |  | 6←1-β-D-Glc 4←1-α-L-Ara -4←1-β-D-Gal 2←1-β-D-Gal | [36,37,38,39,41,42,54] |
| R2 = -β-D-Fuc- |  | 4←1-α-L-Rha-4←1-β-D-Xyl-3←1-β-D-Xyl -3←1-β-D-Xyl 2←1-α-D-Qui | ||||
| R3 = -H | ||||||
| A. gypsophiloides | Acanthophylloside D (4) | С86H136O49 | R1 = -β-D-GlcUA- |  | 4←1-α-L-Ara -4←1-β-D-Gal 2←1-β-D-Gal | [40,41] |
| R2 = -β-D-Fuc- | 4←1-L-Rha- 4←1-β-D-Xyl- 3←1-β-D-Xyl- 3←1-β-D-Xyl | |||||
| R3 = -OH | ||||||
| A. gypsophiloides | 3-O-[β-D-Galactopyranosyl-(1→2)-[α-L-arabinopyranosyl-(1→3)]-β-D-glucuronopyranosyl] gypsogenin 28-β-D-xylopyranosyl-(1→3)-β-D-xylopyranosyl-(1→3)-α-L-rhamnopyranosyl-(1→2)-[6-deoxy-β-D-glucopyranosyl-(1→4)]-β-D-fucopyranosyl ester (5) | С75H118O39 | R1 = -β-D-GlcUA- |  | 2←1-β-D-Gal 3←1-α-L-Ara | [58] |
| R2 = -β-D-Fuc- |  | 2←1-α-L-Rha-3←1-β-D-Xyl-3←1-β-D-Xyl 4←1-α-D-Qui | ||||
| R3 = -H | ||||||
| A. gypsophiloides | 3-O-[β-D-Galactopyranosyl-(1→2)-[α-L-arabinopyranosyl-(1→3)]-β-D-glucuronopyranosyl]quillaic acid 28-β-D-xylopyranosyl-(1→3)-β-D-xylopyranosyl-(1→3)-α-L-rhamnopyranosyl-(1→2)-[6-deoxy-β-D-glucopyranosyl-(1→4)]-β-D-fucopyranosyl ester (6) | C75H118O40 | R1 = -β-D-GlcUA- |  | 2←1-β-D-Gal 3←1-α-L-Ara | [58] |
| R2 = -β-D-Fuc- |  | 2←1-α-L-Rha-3←1-β-D-Xyl-3←1-β-D-Xyl 4←1-α-D-Qui | ||||
| R3 = -OH | ||||||
| A. paniculata | Paniculatoside C (7) | С54H86O25 |  | [41,59,60] | ||
| R = -β-D-Glc |  | 4←1-β-D-Glc 6←1-β-D-Glc-6←1-β-D-Glc | ||||
| A. gypsophiloides | Gypsogenic acid (8) | C30H46O5 | R = H | [57] | ||
| Ecdysteroids | ||||||
| A. tadshikistanica, A. gypsophiloides | 20-Hydroxyecdysone (9) | C27H44O7 | R = H |  | [21,47] | |
| A. tadshikistanica | Polypodine B (10) | C27H44O8 | R = OH | [21] | ||
| A. gypsophiloides | 3-epi-2-Deoxyecdysone (11) | C27H44O5 |  | [47,65] | ||
| Flavonoids | ||||||
| A. paniculata | Quercetin (12) | C15H10O7 |  | [48] | ||
| A. tadshikistanica | Vicenin (13) | C27H30O15 |  | [21] | ||
| Major volatile compounds | ||||||
| A. gypsophiloides | Pulegone (14) | C10H16O |  | [49] | ||
| A. gypsophiloides | trans-p-Menthan-3-one (15) | C10H18O |  | [49] | ||
| A. gypsophiloides | trans-Verbenol (16) | C10H16O |  | [49] | ||
| A. gypsophiloides | Phytol (17) | C20H40O |  | [49] | ||
| A. gypsophiloides | 2-Isopropyl-5-methylcyclohexanone (18) | C10H18O |  | [49] | ||
| A. gypsophiloides | Isopiperitenon (19) | C10H14O |  | [49] | ||
6.4. Essential Oils
6.5. Fatty Acids
6.6. Polysaccharides, Pectins, and Hemicelluloses
6.7. Other Compounds
7. Biological Activities
7.1. Anti-Inflammatory Activity
7.2. Immunomodulatory Activity
7.3. Adjuvant Activity
7.4. Hemolytic Activity
7.5. Cytotoxic Activity
7.6. Analgesic Activity
7.7. Antifungal Activity
7.8. Antioxidant Activity
7.9. Acute Toxicity
7.10. Other Activities
8. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mabberley, D.J. The Plant-Book: A Portable Dictionary of the Vascular Plants, 2nd ed.; Cambridge University Press: New York, NY, USA, 1997; p. 858. ISBN 0-521-41421-0. [Google Scholar]
- Jia, Z.; Koike, K.; Sahu, N.P.; Nikaido, T. Triterpenoid saponins from Caryophyllaceae family. Stud. Nat. Prod. Chem. 2002, 26, 3–61. [Google Scholar] [CrossRef]
- Jakimiuk, K.; Wink, M.; Tomczyk, M. Flavonoids of the Caryophyllaceae. Phytochem. Rev. 2022, 21, 179–218. [Google Scholar] [CrossRef]
- Alamgir, A.N.M. Therapeutic Use of Medicinal Plants and Their Extracts; Springer International Publishing AG: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Flora Tadzhikskoy SSR. Gvozdichnye. Vol. 3. Orekhovye (Original Name: Флoра Таджикскoй ССР. Гвoздичные. Тoм 3. Орехoвые); Ovchinnikov, P.N., Ed.; Academy of Sciences of SSSR: Leningrad, Russia, 1968; Volume 3, p. 611. [Google Scholar]
- Pirani, A.; Rabeler, R.K. Nomenclatural notes on Acanthophyllum (Caryophylleae, Caryophyllaceae). Phytotaxa 2017, 303, 197. [Google Scholar] [CrossRef]
- Pirani, A.; Zarre, S.; Pfeil, B.E.; Bertrand, Y.J.K.; Assadi, M.; Oxelman, B. Molecular phylogeny of Acanthophyllum (Caryophyllaceae: Caryophylleae), with emphasis on infrageneric classification. Taxon 2014, 63, 592–607. [Google Scholar] [CrossRef]
- Xojimatov, O.K.; Haydarov, X.Q.; Xamraeva, D.T.; Imomova, D.A.; Xujanov, A.N. Atlas of Herbal Plants of Uzbekistan (O‘zbekiston Dorivor O‘simliklar Atlasi); Samarkand State University: Samarqand, Uzbekistan, 2021; p. 219. [Google Scholar]
- Govaerts, R.H.A. World Checklist of Seed Plants; MIM: Deurne, The Netherlands, 1995. [Google Scholar]
- Ghaffari, S.M. Cytogeography and patterns of distribution of the genus Acanthophyllum (Caryophyllaceae). Iran. J. Botany 2021, 27, 164–176. [Google Scholar] [CrossRef]
- Mahmoodi, M.; Falatoury, A.N. Allochrusa lutea (Caryophylloideae, Caryophyllaceae) a new species from Iran. Phytotaxa 2016, 277, 191–198. [Google Scholar] [CrossRef]
- Tamanyan, K. Allochrusa Takhtajanii. In The IUCN Red List of Threatened Species 2014; The International Union for Conservation of Nature: Gland, Switzerland, 2014. [Google Scholar] [CrossRef]
- Gemejiyeva, N.G.; Mursaliyeva, V.K.; Mukhanov, T.M. Assessment of the current state of Allochrusa gypsophiloides (Regel) Schischk. Natural populations in the South-Kazakhstan region. News Natl. Acad. Sci. Repub. Kazakhstan Ser. Biol. Medical. 2016, 1, 22–29. [Google Scholar]
- Aidarbayeva, A.K.; Amantayeva, A.A.; Taneeva, G.T. Allochruzes of South Kazakhstan–perspective saponin plants. Вестник Казахскoгo Нациoнальнoгo Медицинскoгo Университета 2022, 1, 571–573. [Google Scholar]
- Turker, A. Revision of Genus Allochrusa Bunge ex Boiss (Caryophyllaceae) in Turkey. Master’s Thesis, Bozok University, Institute of Science, Yozgat, Turkey, 2018. [Google Scholar]
- Arifkhodzhaev, A.O. Polysaccharides of saponin-bearing plants VII. Study of the polysaccharides of the roots of Allochrusa paniculata. Chem. Nat. Compd. 1995, 31, 455–456. [Google Scholar] [CrossRef]
- Eisenman, S.W.; Zaurov, D.E.; Struwe, L. Medicinal Plants of Central Asia: Uzbekistan and Kyrgyzstan; Springer: New York, NY, USA, 2013; p. 340. [Google Scholar] [CrossRef]
- Khasanov, F. (Ed.) Red Data Book of Republic of Uzbekistan Plants; Chinor ENK: Tashkent, Uzbekistan, 2019; Volume 1, p. 190. [Google Scholar]
- Red Book. Wild Species of the Flora of the USSR in Need of Protection (Красная книга. Дикoрастущие виды флoры СССР, нуждающиеся в oхране); Takhtajyan, A., Ed.; Nauka: Leningrad, Russia, 1975; p. 204. [Google Scholar]
- Vvedensky (Ed.) Flora of Uzbekistan, Vol. 2. (Флoра Узбекистана. Тoм 2); Academy of Sciences of Uzbekskoy SSR: Tashkent, Russia, 1953; p. 414. [Google Scholar]
- Darmogray, S.V.; Filippova, A.S.; Erofeeva, N.S.; Afanasev, А.А.; Darmogray, V.N. Phytochemical study of certain polyphenolic and steroid compounds in the plants of the genus Cerastium L., Arenaria L., Stellaria L., Allochrusa bunge ex Boiss. Caryophyllaceae Juss family. Int. J. Appl. Fundam. Research 2017, 12 Pt 1, 54–58. [Google Scholar]
- Plantarium. Plants and Lichens of Russia and Neighboring Countries: Open Online Galleries and Plant Identification Guide. Available online: https://www.plantarium.ru/lang/en/page/view/item/177.html (accessed on 29 March 2023).
- Mursaliyeva, V.; Kozhebayeva, Z.; Rakhimbayev, I.; Gemejiyeva, N. Qualitative and quantitative analysis of saponins in Allochrusa gypsophiloides, KazNU Bulletin. Biol. Series 2016, 3, 115–123. [Google Scholar]
- Hojimatov, M. Wild Herbal Plants of Tadjikistan (Дикoрастущие лекарственные растения Таджикистана); Tajik Soviet Encyclopedia: Dushanbe, Tajikistan, 1989; p. 368. [Google Scholar]
- Khalmatov, K. Wild Medicinal Plants of Uzbekistan (Дикoрастущие лекарственные растения Узбекистана); Ibn Sino Publihser: Tashkent, Uzbekistan, 1991; p. 205. [Google Scholar]
- Grudzinskaya, L.M.; Gemedzhieva, N.G.; Nelina, N.V.; Karzhaubekova, Z.H. Annotated List of Medicinal Plants in Kazakhstan: A reference Book; House of the USSR Academy of Sciences: Leningrad, Russia, 2014; Volume 99. [Google Scholar]
- Van Wyk, B.E.; Wink, M. Phytomedicines, Herbal Drugs and Poison; Briza: Pretoria, South Africa, 2015; p. 304. ISBN 978-0226204918. [Google Scholar]
- Van Wyk, B.E.; Wink, M. Medicinal Plants of the World; Briza: Pretoria, South Africa, 2017; p. 520. ISBN 978-1-78639-325-8. [Google Scholar]
- Khudaykova, S.; Bogoyavlenskij, V.; Tolmatscheva, V.; Berezin, V.; Salas, P. Immunostimulating activity of saponin from Caryophyllaceae. In Proceedings of the 9th International Conference of Horculture, Lednice, Czech Republic, 3–6 September 2001; Volume 2, pp. 343–347. [Google Scholar]
- Aleksjuk, P.; Moldahanov, E.; Akanova, K.; Anarkulova, J.; Bogojavlenskij, A. Standardization of saponin containing drugs with antiviral activity. Int. J. Appl. Basic Research 2014, 6, 80–81. [Google Scholar]
- Man, S.; Gao, W.; Zhang, Y.; Huang, L.; Lui, C. Chemical study and medical application of saponins as anti-cancer agents. Fitoterapia 2010, 81, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Turmagambetova, A.S.; Alexyuk, P.G.; Bogoyavlenskiy, A.P.; Zaitseva, I.A.; Omirtaeva, E.S.; Alexyuk, M.S.; Sokolova, N.S.; Berezin, V.E. Adjuvant activity of saponins from Kazakhstani plants on the immune responses to subunit influenza vaccine. Arch. Virol. 2017, 162, 3817–3826. [Google Scholar] [CrossRef] [PubMed]
- Mursalıyeva, V.; Imanbayeva, A.; Parkhatova, R. Seed germination of Allochrusa gypsophiloides (Caryophyllaceae), as an endemic species from Central Asia and Kazakhstan. Seed Sci. Technol. 2020, 48, 289–295. [Google Scholar] [CrossRef]
- Mursalıyeva, V.; Imanbayeva, A. Micropropagation of Turkestan Soap Root Allochrusa gypsophiloides–Natural Source of Saponins. Int. J. Second. Metab. 2020, 7, 1–7. [Google Scholar] [CrossRef]
- Amanmuradov, K.; Kondratenko, E.S.; Abubakirov, N.K. The saponin of Acanthophyllum gypsophiloides. Chem. Nat. Compd. 1965, 1, 113. [Google Scholar] [CrossRef]
- Putieva, Z.M.; Mzhel’skaya, L.G.; Kondratenko, E.S.; Abubakirov, N.K. Triterpene glycosides of Acanthophyllum gypsophiloides. Chem. Nat. Compd. 1970, 6, 506–507. [Google Scholar] [CrossRef]
- Putieva, Z.M.; Mzhel’skaya, L.G.; Gorovits, T.T.; Kondratenko, E.S.; Abubakirov, N.K. Triterpene glycosides of Acanthophyllum gypsophiloides. III. Structure of the O-glycosidic carbohydrate chain of acanthophyllosides B and C. Chem. Nat. Compd. 1975, 11, 188–192. [Google Scholar] [CrossRef]
- Putieva, Z.M.; Mzhel’skaya, L.G.; Gorovits, T.T.; Kondratenko, E.S.; Abubakirov, N.K. Triterpene glycosides of Acanthophyllum gypsophiloides IV. The structure of acanthophyllosides B and C. Chem. Nat. Compd. 1975, 11, 756–761. [Google Scholar] [CrossRef]
- Putieva, Z.M.; Mzhel’skaya, L.G.; Gorovits, T.T.; Kondratenko, E.S.; Abubakirov, N.K. Triterpene glycosides of Acanthophyllum gypsophiloides V. D-quinovose in acanthophyllosides B and C. Chem. Nat. Compd. 1977, 13, 679–682. [Google Scholar] [CrossRef]
- Putieva, Z.M.; Gorovich, T.T.; Kondratenko, E.S.; Abubakirov, N.K. Triterpene glycosides of Acanthophllum gypsophiloides. VI. Structure of acanthophylloside D. Chem. Nat. Compd. 1979, 15, 148–151. [Google Scholar] [CrossRef]
- Kondratenko, E.S.; Putieva, Z.M.; Abubakirov, N.K. Triterpene glycosides of plants of the family Caryophyllaceae. Chem. Nat. Compd. 1981, 17, 303–317. [Google Scholar] [CrossRef]
- Aripov, K.N. The chemistry of plant substances in Uzbekistan. Chem. Nat. Compd. 1995, 31, 2–15. [Google Scholar] [CrossRef]
- Arifkhodzhaev, A.O.; Rakhimov, D.A. Polysaccharides of saponin-bearing plants V. Polysaccharides of the epigeal organs of Allochrusa gypsophiloides. Chem. Nat. Compd. 1992, 28, 379. [Google Scholar] [CrossRef]
- Arifkhodzhaev, A.O. Polysaccharides of saponin-bearing plants VIII. Structural investigation of a glucogalactan from the roots of Allochrusa gypsophiloides. Chem. Nat. Compd. 1996, 32, 523–525. [Google Scholar] [CrossRef]
- Arifkhodzhaev, A.O. Polysaccharides of saponin-bearing plants. IX. Structures of oligosaccharides from the glucogalactan of Allochrusa gypsophiloides. Chem. Nat. Compd. 1996, 32, 526–528. [Google Scholar] [CrossRef]
- Arifkhodzhaev, A.O.; Kondratenko, E.S. Polysaccharides of saponin-bearing plants. I. Characteristics of the polysaccharides of Allochrusa gypsophiloides. Chem. Nat. Compd. 1983, 19, 222–223. [Google Scholar] [CrossRef]
- Tuleuov, B.I.; Zavarzin, I.V.; Shashkov, A.S.; Chernoburova, E.I.; Adekenov, S.M. 3α,14α,22R,25-Tetrahydroxy-5β(H)-cholest-7-en-6-one, a phytoecdysteroid from Acanthophyllum gypsophiloides possessing anti-inflammatory and analgesic activities. Russ. Chem. Bull. 2018, 67, 663–666. [Google Scholar] [CrossRef]
- Grudzinskaya, L.; Gemejiyeva, N.; Karzhaubekova, Z.; Nelina, N. Botanical coverage of the leading families of medicinal flora of Kazakhstan. BIO Web Conf. 2021, 31, 00007. [Google Scholar] [CrossRef]
- Mamadalieva, R.Z.; Sharopov, F.S.; Ibragimov, A.A.; Abdullaev, S.V.; Khujaev, V.U. Chemical composition and in vitro biological activities of essential oil from Allochrusa gypsophiloides. Food Ther. Health Care 2021, 3, 10–15. [Google Scholar] [CrossRef]
- Sdykov, T.; Abduazimov, K.A. Lignins of Allochruza paniculata and Glycyrrhiza glabra. Chem. Nat. Compd. 2002, 38, 288–290. [Google Scholar] [CrossRef]
- Hostettmann, K.; Chen, S.; Marston, A.; Stuppner, H. Handbook of Chemical and Biological Plant Analytical Methods; John Wiley & Sons: New York, NY, USA, 2014; p. 1176. ISBN 978-1-119-95275-6. [Google Scholar]
- Du, J.R.; Long, F.Y.; Chen, C. Chapter Six-Research Progress on Natural Triterpenoid Saponins in the Chemoprevention and Chemotherapy of Cancer. Enzymes 2014, 36, 95–130. [Google Scholar] [CrossRef]
- Bottger, S.; Melzig, M.F. Triterpenoid saponins of the Caryophyllaceae and Illecebraceae family. Phytochem. Lett. 2011, 4, 59–68. [Google Scholar] [CrossRef]
- Putieva, Z.M.; Kondratenko, E.S.; Abubakirov, N.K. Formation of gypsogenic acid on the hydrolysis of gypsogenin glycosides. Chem. Nat. Compd. 1979, 15, 206. [Google Scholar] [CrossRef]
- Kuzmin, E.V.; Tugelibaev, G.T.; Sitpaeva, G.T. Plant Population Reconstruction of Allochrusa gypsophiloides Rgl. in Southern Kazakhstan. In A Study of Kazakhstan Flora and Its Protection. Collection of Scientific Articles; Abiev, S., Fursova, N., Veselova, P., Sitpaeva, G., Aidarbayeva, D., Eds.; Aidana LLP: Almaty, Kazakhstan, 2001; pp. 91–194. [Google Scholar]
- Gladyshev, A.; Mishchenko, A. Acanthophyllums of Turkmenistan, Their Biology and Prospects for Economic Use; Science: Ashgabat, Turkmenistan, 1990; ISBN 5-8338-0165-6. [Google Scholar]
- Belous, V.N.; Ryabinin, A.A. Gypsogenic acid from the roots of Acanthophyllum subglabrum and A. gypsophiloides. Chem. Nat. Compd. 1967, 3, 79–80. [Google Scholar] [CrossRef]
- Khatuntseva, E.A.; Men’shov, V.M.; Shashkov, A.S.; Tsvetkov, Y.E.; Stepanenko, R.N.; Vlasenko, R.Y.; Shults, E.E.; Tolstikov, G.A.; Tolstikova, T.G.; Baev, D.S.; et al. Triterpenoid saponins from the roots of Acanthophyllum gypsophiloides Regel. Beilstein. J. Org. Chem. 2012, 8, 763–775. [Google Scholar] [CrossRef]
- Putieva, Z.M.; Kondratenko, E.S.; Abubakirov, N.K. Triterpene glycosides of Acanthophyllum paniculata. Chem. Nat. Compd. 1972, 8, 670. [Google Scholar] [CrossRef]
- Putieva, Z.M.; Kondratenko, E.S.; Abubakirov, N.K. Triterpene glycosides of Acanthophyllum paniculata. II. Structure of paniculatoside C. Chem. Nat. Compd. 1974, 10, 114–115. [Google Scholar] [CrossRef]
- Lafont, R.; Balducci, C.; Dinan, L. Ecdysteroids. Encyclopedia 2021, 1, 1267–1302. [Google Scholar] [CrossRef]
- Dinan, L.; Mamadalieva, N.Z.; Lafont, R. Dietary Phytoecdysteroids. In Handbook of Dietary Phytochemicals; Xiao, J., Sarker, S., Asakawa, Y., Eds.; Springer: Singapore, 2020; pp. 1–54. ISBN 978-981-13-1745-3. [Google Scholar] [CrossRef]
- Zibareva, L.N. Phytoecdysteroids of Caryophyllaceae Juss. Contemp. Probl. Ecology 2009, 2, 476–488. [Google Scholar] [CrossRef]
- Mamadalieva, N.Z.; Lafont, R.; Wink, M. Diversity of secondary metabolites in the genus Silene L. (Caryophyllaceae)–Structures, Distribution, and Biological properties. Diversity 2014, 6, 415–499. [Google Scholar] [CrossRef]
- Temirgaziev, B.S.; Tuleuov, B.I.; Romanova, M.A.; Seidakhmetova, R.B.; Seilkhanov, T.M.; Seilkhanov, O.T.; Salkeeva, L.K.; Adekenov, S.M. Supramolecular complexes of 3-epi-2-deoxyecdysone with cyclodextrins and their anti-inflammatory activity. Russ. J. Gen. Chem. 2019, 89, 424–428. [Google Scholar] [CrossRef]
- Dilworth, L.L.; Riley, C.K.; Stennett, D.K. Chapter 5-Plant Constituents: Carbohydrates, Oils, Resins, Balsams, and Plant Hormones. In Pharmacognosy: Fundamentals, Applications and Strategies; Badal, S., Delgoda, R., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 61–80. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential Oils’ Chemical Characterization and Investigation of Some Biological Activities: A Critical Review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Mode of action and toxicology of plant toxins and poisonous plants. Mitt. Jul. Kühn-Inst. 2009, 421, 93–112. [Google Scholar]
- Makhutova, O.N.; Sushchik, N.N.; Gladyshev, M.I. Fatty Acid-Markers as Foodweb Tracers in Inland Waters. In Encyclopedia of Inland Waters, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2022; Volume 4, pp. 713–726. [Google Scholar] [CrossRef]
- Wink, M. Modes of Action of Herbal Medicines and Plant Secondary Metabolites. Medicines 2015, 2, 251–286. [Google Scholar] [CrossRef]
- Wagle, N.; Nagarjuna, S.; Sharma, H.; Dangi, N.B.; Sapkota, H.P.; Naik, B.S.; Padhaya, R.R. Evaluation of antinociceptive and anti-inflammatory activity of phytosterol present in chloroform extract of Phyllanthus maderaspatensis. Indian J. Physiol. Pharmacol. 2016, 60, 90–95. [Google Scholar]
- Arnold, E.; Benz, T.; Zapp, C.; Wink, M. Inhibition of phospholipase A2α (PLA2α) by medicinal plants in relation to their phenolic content. Molecules 2015, 20, 15033–15048. [Google Scholar] [CrossRef]
- Khushbakatova, Z.A.; Putiyeva, Z.M.; Syrov, V.N.; Kruglikov, B.A.; Tarasenko, T.Y.; Salazhov, Y.L. Adjuvant activity of some triterpenic saponins. Khimiko-Farmatsevticheskii Zhurnal. 1993, 27, 48–52. [Google Scholar]
- Bazhenova, E.D.; Aliev, K.U. Pharmacology of paniculatoside. In Doklady Akademii Nauk UzSSR; Akademiia: Moscow, Russia, 1976; Volume 2, pp. 49–50. [Google Scholar]
- Bomford, R. Saponin and other haemolysins (vitamin A, aliphatic amines, polyene antibiotics) as adjuvants for SRBC in the mouse. Evidence for a role for cholesterol-binding in saponin adjuvanticity. Int. Arch. Allergy. Appl. Immuno. 1980, 63, 170–177. [Google Scholar] [CrossRef]
- Sharopov, F.S.; Mamadalieva, R.Z.; Hussain, H.; Abdullaev, S.V.; Khujaev, V.U. Allochrusa gypsophiloides, an endemic medicinal plant of Central Asia: Phytochemistry and bioactivity. In Proceedings of the 4th International Symposium on Phytochemicals in Medicine and Food, Xi’an, China, 30 November–5 December 2020; p. 380. [Google Scholar]
- Polievtsev, N.P.; Sultanov, M.B. Pharmacology of a saponin extracted from the plant Acanthophyllus gypsophiloides. In Farmakologiia Alkaloidov i Serdechnuikh Glikozidov (Pharmacology of Alkaloids and Cardiac Glycosides); Sultanov, M.B., Ed.; FAN: Tashkent, Uzbekistan, 1971; pp. 240–243. (In Russian) [Google Scholar]
- Mamadaliyeva, R.Z.; Xo’jayev, V.U. Investigation of anthelmintic activity of the plant extracts from Allochrusa gypsophiloides (Allochrusa gypsophiloides o’simligi ekstraktlarining gelmintga qarshi ta’sirini o’rganish). “Problems and Perspectives of Chemistry Goods and Folk Medicine”. In Proceedings of the Materials of the VII-International Scientific-Practical Conference, Andijan, Uzbekistan, 18–19 September 2020. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamadalieva, R.; Khujaev, V.; Šoral, M.; Mamadalieva, N.Z.; Wink, M. The Genus Allochrusa: A Comprehensive Review of Botany, Traditional Uses, Phytochemistry, and Biological Activities. Diversity 2023, 15, 574. https://doi.org/10.3390/d15040574
Mamadalieva R, Khujaev V, Šoral M, Mamadalieva NZ, Wink M. The Genus Allochrusa: A Comprehensive Review of Botany, Traditional Uses, Phytochemistry, and Biological Activities. Diversity. 2023; 15(4):574. https://doi.org/10.3390/d15040574
Chicago/Turabian StyleMamadalieva, Rano, Vahobjon Khujaev, Michal Šoral, Nilufar Z. Mamadalieva, and Michael Wink. 2023. "The Genus Allochrusa: A Comprehensive Review of Botany, Traditional Uses, Phytochemistry, and Biological Activities" Diversity 15, no. 4: 574. https://doi.org/10.3390/d15040574