
Updated Taxonomic Key of European Nycteribiidae (Diptera), with a Host-Parasite Network

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
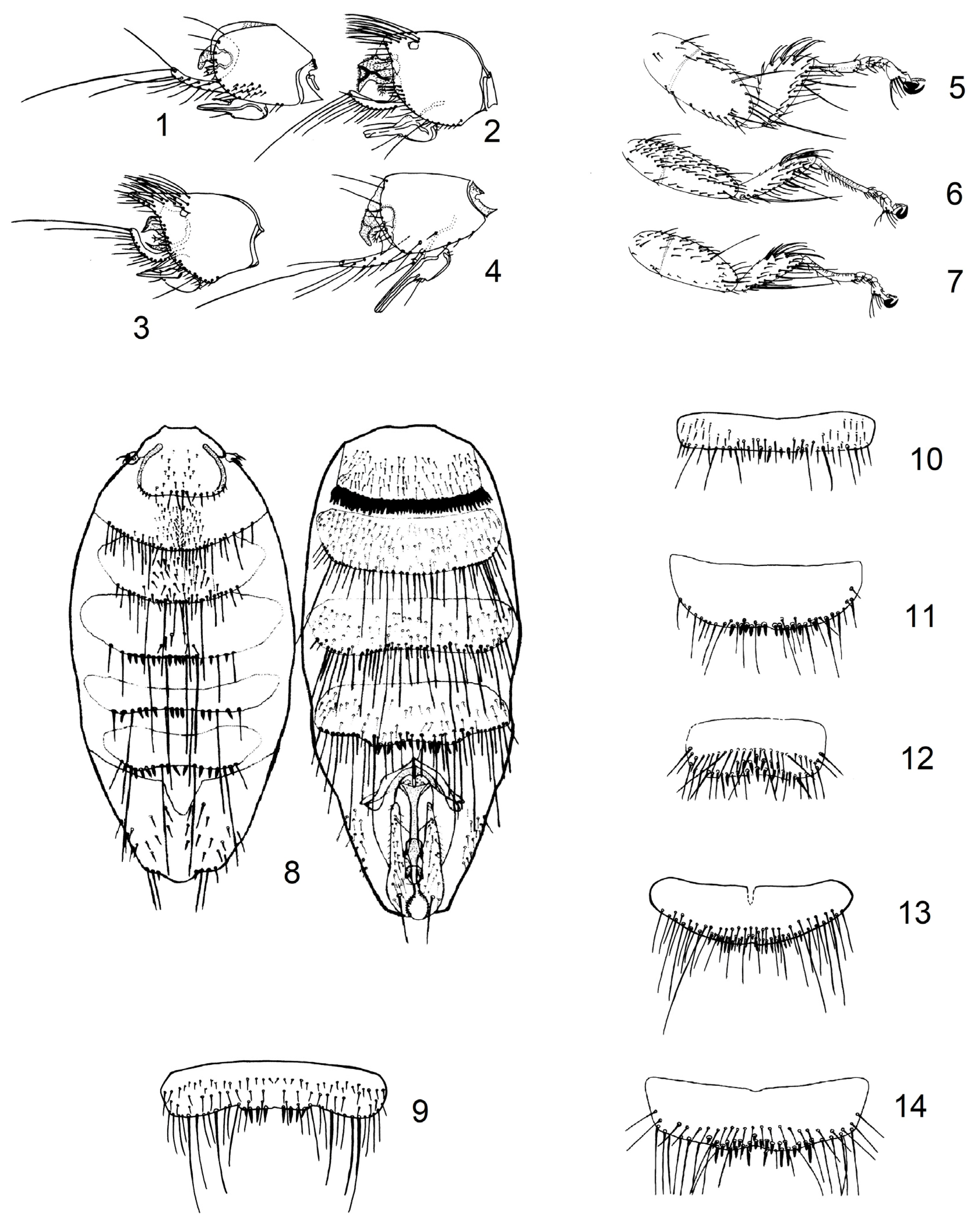
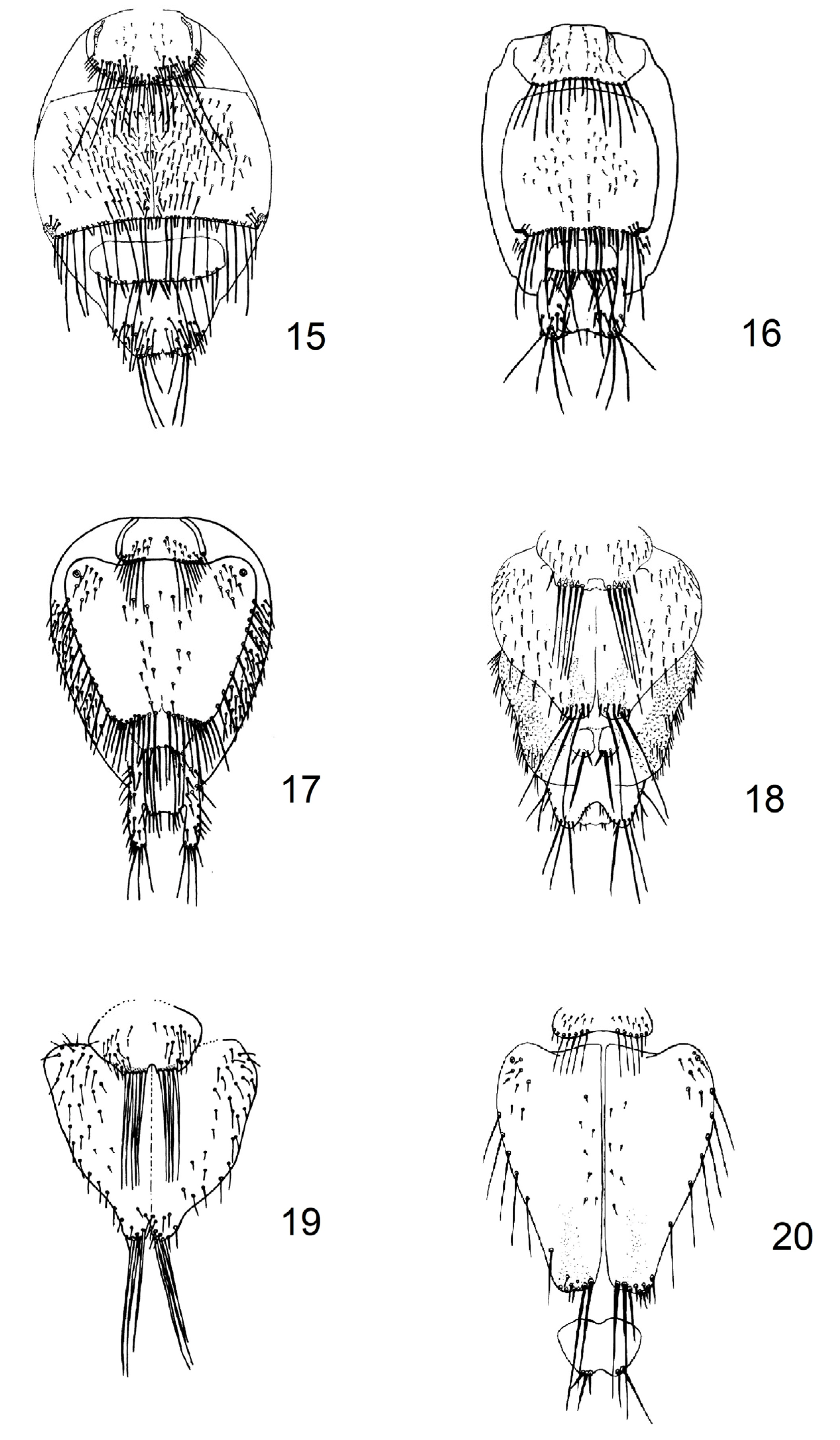
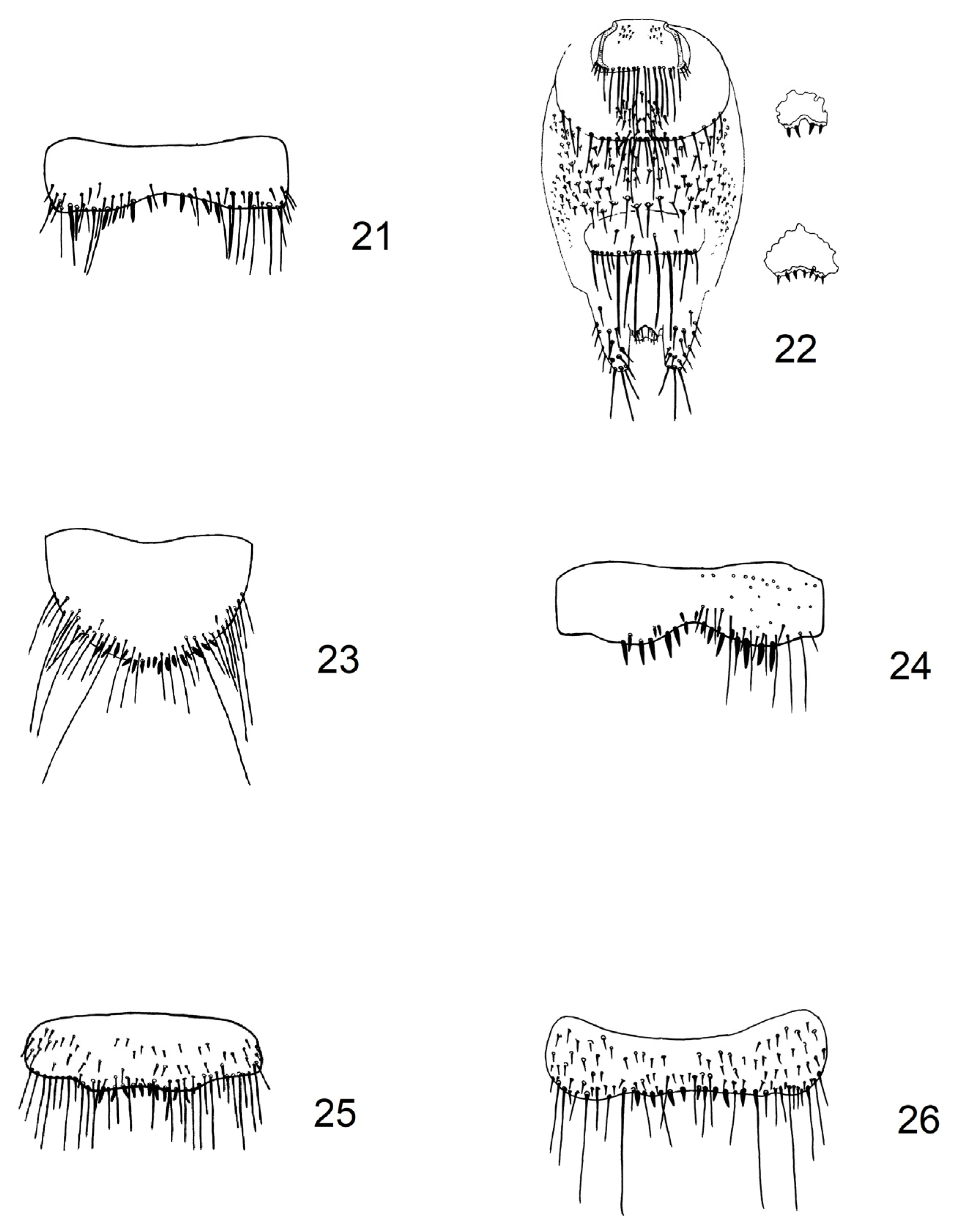
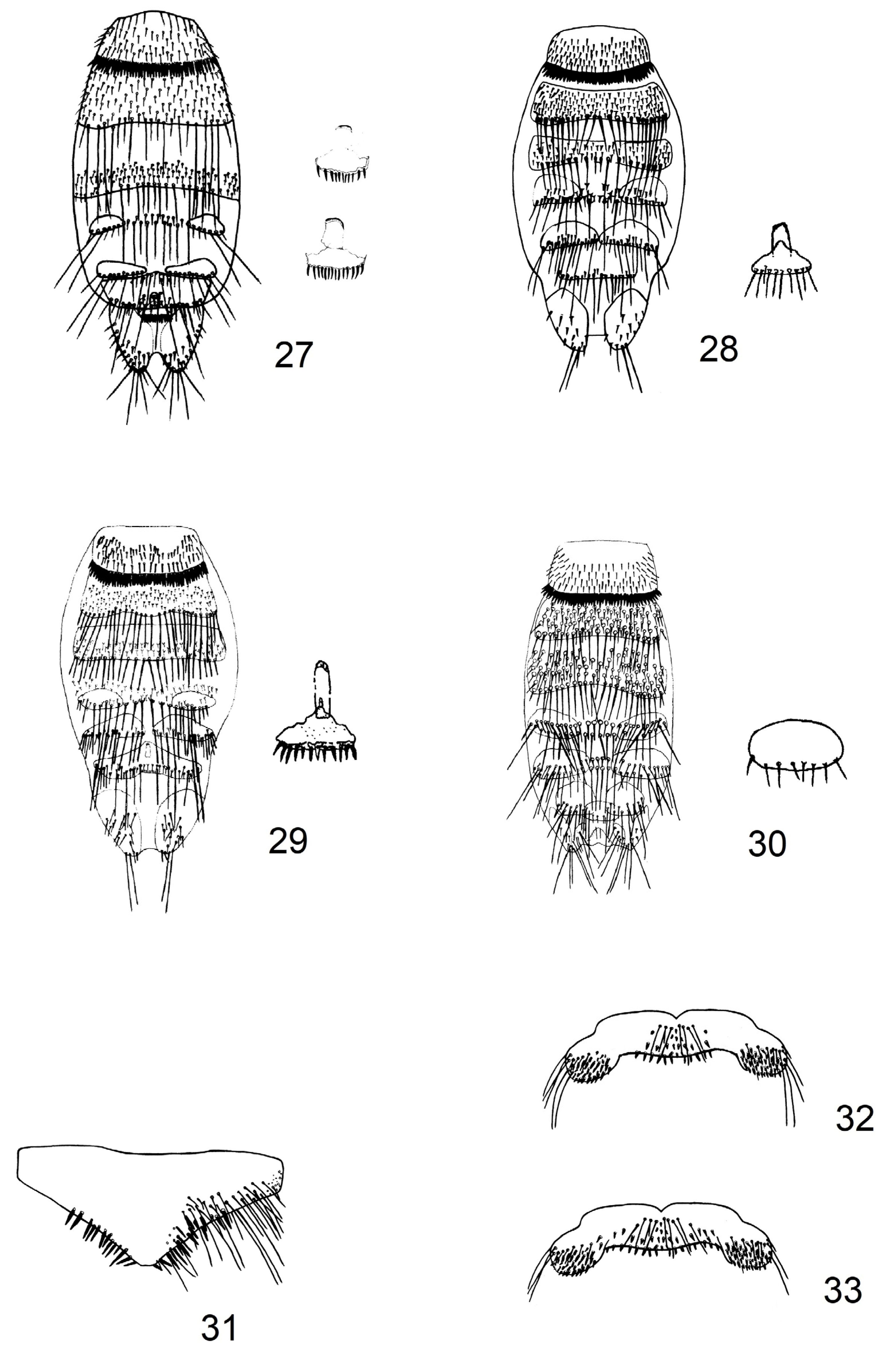
2.1. Updating of Identification Key
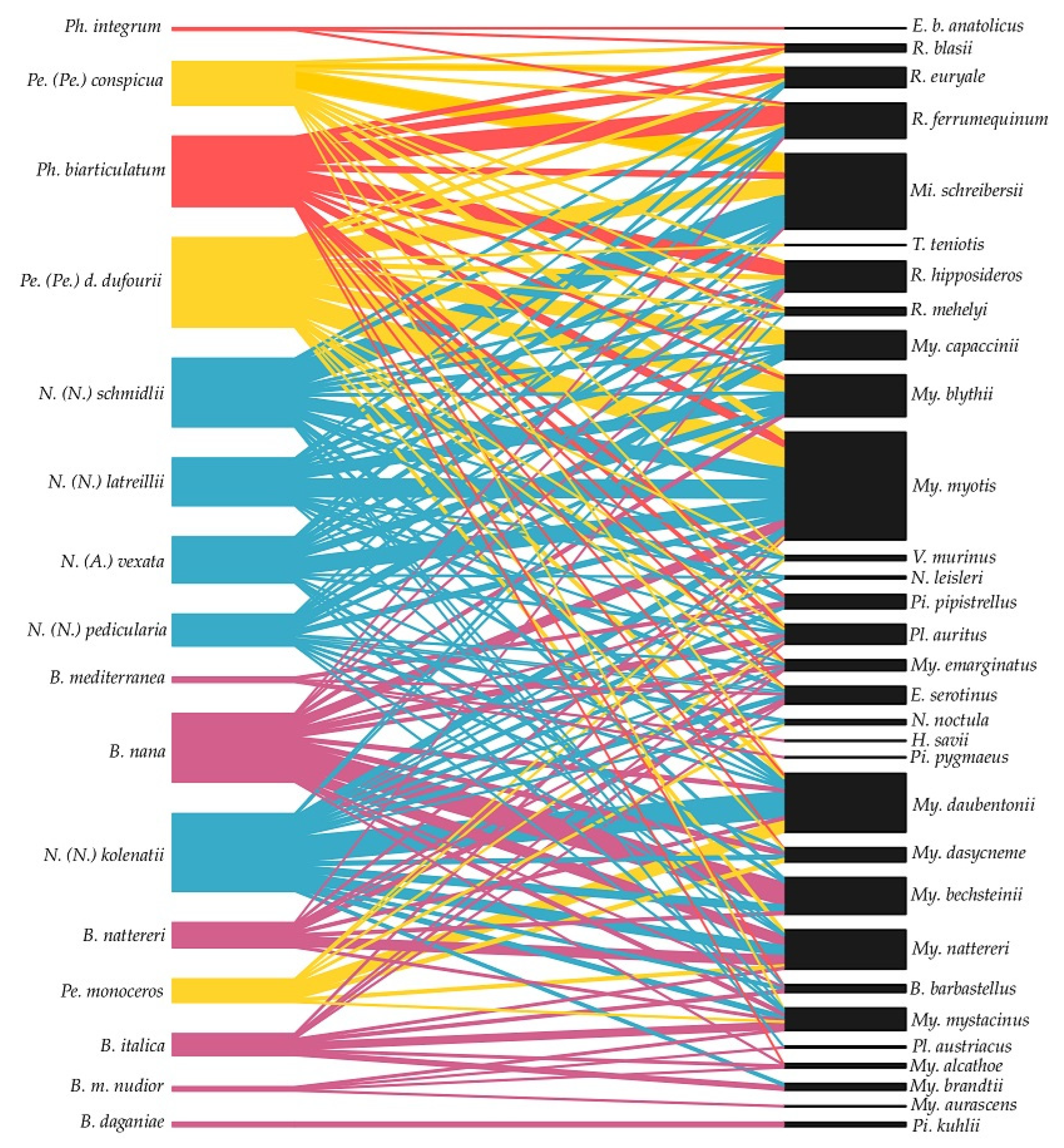
2.2. Host Specificity, Bipartite Networks, and Statistics
3. Results





Bat Fly-Bat Species Associations and the Bipartite Network
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moreno Santillán, D.D.; Lama, T.M.; Gutierrez Guerrero, Y.T.; Brown, A.M.; Donat, P.; Zhao, H.; Rossiter, S.J.; Yohe, L.R.; Potter, J.H.; Teeling, E.C.; et al. Large-scale genome sampling reveals unique immunity and metabolic adaptations in bats. Mol. Ecol. 2021, 30, 6449–6467. [Google Scholar] [CrossRef]
- Kunz, T.H.; Braun de Torrez, E.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Fráncel, L.A.; García-Herrera, L.V.; Losada-Prado, S. Bats and their vital ecosystem services: A global review. Integr. Zool. 2022, 17, 2–23. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Dick, C.W.; Dittmar, K. Parasitic bat flies (Diptera: Streblidae and Nycteribiidae): Host specificity and potential as vectors. In Bats (Chiroptera) as Vectors of Diseases and Parasites; Springer: Berlin/Heidelberg, Germany, 2014; pp. 131–155. [Google Scholar]
- Orlova, M.V.; Klimov, P.B.; Moskvitina, N.S.; Orlov, O.L.; Zhigalin, A.V.; Smirnov, D.G.; Dzhamirzoyev, H.S.; Vekhnik, V.P.; Pavlov, A.V.; Emelyanova, A.A.; et al. New records of bat flies (Diptera: Nycteribiidae), with an updated checklist of the nycteribiids of Russia. Zootaxa 2021, 4927, 410–430. [Google Scholar] [CrossRef]
- Péter, A.; Mihalca, A.D.; Sándor, A.D. Annotated checklist of the bat flies (Diptera: Nycteribiidae) of Romania. Zootaxa 2022, 5120, 111–127. [Google Scholar] [CrossRef]
- Reeves, W.K.; Lloyd, J.E. Louse flies, keds, and bat flies (Hippoboscoidea). In Medical and Veterinary Entomology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 421–438. [Google Scholar]
- Nirmala, X.; Hypša, V.; Žurovec, M. Molecular phylogeny of Calyptratae (Diptera: Brachycera): The evolution of 18S and 16S ribosomal rDNAs in higher dipterans and their use in phylogenetic inference. Insect Mol. Biol. 2001, 10, 475–485. [Google Scholar] [CrossRef]
- Dittmar, K.; Porter, M.L.; Murray, S.; Whiting, M.F. Molecular phylogenetic analysis of nycteribiid and streblid bat flies (Diptera: Brachycera, Calyptratae): Implications for host associations and phylogeographic origins. Mol. Phylogenet. Evol. 2006, 38, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Seo, M.G.; Lee, S.H.; Oem, J.K.; Kim, S.H.; Jeong, H.; Kim, Y.; Jheong, W.H.; Kwon, O.D.; Kwak, D. Relationship among bats, parasitic bat flies, and associated pathogens in Korea. Parasites Vectors 2021, 14, 503. [Google Scholar] [CrossRef] [PubMed]
- Jobling, F. Streblidae from the Belgian Congo, with description of a new genus and three new species. Rev. Zool. Bot. Afr. 1954, 50, 89–115. [Google Scholar]
- Traub, R. The relationship between spines, combs and other skeletal features of fleas (Siphonaptera) and the vestiture, affinities and habits of their hosts. J. Med. Entomol. 1972, 9, 601. [Google Scholar]
- Dick, C.W.; Patterson, B.D. Bat flies: Obligate ectoparasites of bats. In Micromammals and Macroparasites; Morand, S., Krasnov, B.R., Poulin, R., Eds.; Springer: Tokyo, Japan, 2006; pp. 179–194. [Google Scholar]
- Marshall, A.G. The function of combs in ectoparsitic insects. In Fleas; Traub, R., Starcke, H., Eds.; AA Balkema: Rotterdan, The Netherlands, 1980; pp. 79–87. [Google Scholar]
- Marshall, A.G. The Ecology of Ectoparasitic Insects; Academic Press: New York, NY, USA, 1981; 459p. [Google Scholar]
- McAlpine, J.F. (Ed.) Manual of Nearctic Diptera; Research Branch Agriculture: Ottawa, ON, Canada, 1989; Volume 2, 668p.
- Graciolli, G.; Dick, C.W. Checklist of World Nycteribiidae (Diptera: Hippoboscoidea). 2008. Available online: https://www.researchgate.net/publication/322579074_CHECKLIST_OF_WORLD_NYCTERIBIIDAE_DIPTERA_HIPPOBOSCOIDEA (accessed on 27 February 2023).
- Hůrka, K. Fauna Europaea: Nycteribiidae. In Fauna Europaea: Diptera, Brachycera. Pape, T., Ed.; 2011. version 2017.06. Available online: https://fauna-eu.org (accessed on 1 October 2022).
- Sevcik, M.; Benda, P.; Lucan, R.K. Diptera Pupipara from bats of two large eastern Mediterranean islands, Crete and Cyprus. Turk. J. Zool. 2013, 37, 31–37. [Google Scholar]
- Szentiványi, T.A.; Estók, P.; Földvári, M. Checklist of host associations of European bat flies (Diptera: Nycteribiidae, Streblidae). Zootaxa 2016, 4205, 101–126. [Google Scholar] [CrossRef]
- Hagan, H.R. Embryology of the Viviparous Insects; Ronald Press: New York, NY, USA, 1951; 472p. [Google Scholar]
- Ching, L.M.; Marshall, A.G. The breeding biology of the bat-fly Eucampispoda sundaicum Theodor, 1955 (Diptera: Nycteribiidae). Malayan Nat. J. 1968, 21, 171–180. [Google Scholar]
- Marshall, A.G. The life cycle of Basilia hipsida Theodor 1967 (Diptera: Nycteribiidae) in Malaysia. Parasitology 1970, 61, 1–18. [Google Scholar] [CrossRef]
- Foelix, R.F.; Stocker, R.F.; Steinbrecht, R.A. Fine structure of a sensory organ in the arista of Drosophila melanogaster and some other dipterans. Cell. Tissue Res. 1989, 258, 277–287. [Google Scholar] [CrossRef]
- Han, H.J.; Wen, H.L.; Zhou, C.M.; Chen, F.F.; Luo, L.M.; Liu, J.W.; Yu, X.J. Bats as reservoirs of severe emerging infectious diseases. Virus Res. 2015, 205, 1–6. [Google Scholar] [CrossRef]
- De Wit, E.; Van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef]
- Letko, M.; Seifert, S.N.; Olival, K.J.; Plowright, R.K.; Munster, V.J. Bat-borne virus diversity, spillover and emergence. Nat. Rev. Microbiol. 2020, 18, 461–471. [Google Scholar] [CrossRef]
- Butler, C.D. Infectious disease emergence and global change: Thinking systemically in a shrinking world. Infect. Dis. Poverty 2012, 1, 1–7. [Google Scholar] [CrossRef]
- Korytár, Ľ.; Ondrejková, A.; Drážovská, M.; Zemanová, S.; Prokeš, M. Serological survey of lyssaviruses in synanthropic bats and human exposure to bats in Slovakia. Ann. Agric. Environ. Med. AAEM 2022, 29, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Brook, C.E.; Bai, Y.; Dobson, A.P.; Osikowicz, L.M.; Ranaivoson, H.C.; Zhu, Q. Bartonella spp. in fruit bats and blood-feeding ectoparasites in Madagascar. PLoS Negl. Trop. Dis. 2015, 9, 1–9. [Google Scholar] [CrossRef]
- Szentiványi, T.; Christe, P.; Glaizot, O. Bat flies and their microparasites: Current knowledge and distribution. Front. VEt Sci. 2019, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pawęska, J.T.; Jansen van Vuren, P.; Storm, N.; Markotter, W.; Kemp, A. Vector Competence of Eucampsipoda africana (Diptera: Nycteribiidae) for Marburg Virus Transmission in Rousettus aegyptiacus (Chiroptera: Pteropodidae). Viruses 2021, 13, 2226. [Google Scholar] [CrossRef]
- Verrett, T.B.; Webala, P.W.; Patterson, B.D.; Dick, C.W. Remarkably low host specificity in the bat fly Penicillidia fulvida (Diptera: Nycteribiidae) as assessed by mitochondrial COI and nuclear 28S sequence data. Parasites Vectors 2022, 15, 392. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Garcia, K.D.; Sandoval-Ruiz, C.A.; Saldana-Vazquez, R.A.; Schondube, J.E. The effects of seasonality on host-bat fly ecological networks in a temperate mountain cave. Parasitology 2017, 144, 692–697. [Google Scholar] [CrossRef]
- Kelm, D.H.; Toelch, U.; Jones, M.M. Mixed-species groups in bats: Non-random roost associations and roost selection in neotropical understory bats. Front. Zool. 2021, 18, 53. [Google Scholar] [CrossRef]
- Dallas, T.A.; Han, B.A.; Nunn, C.L.; Park, A.W.; Stephens, P.R.; Drake, J.M. Host traits associated with species roles in parasite sharing networks. Oikos 2019, 128, 23–32. [Google Scholar] [CrossRef]
- Sebastián Tello, J.; Stevens, R.D.; Dick, C.W. Patterns of species co-occurrence and density compensation: A test for interspecific competition in bat ectoparasite infracommunities. Oikos 2008, 117, 693–702. [Google Scholar] [CrossRef]
- Durán, A.A.; Saldaña-Vázquez, R.A.; Graciolli, G.; Peinado, L.C. Specialization and Modularity of a Bat Fly Antagonistic Ecological Network in a Dry Tropical Forest in Northern Colombia. Acta Chiropterologica 2018, 20, 503–510. [Google Scholar] [CrossRef]
- Delmas, E.; Besson, M.; Brice, M.H.; Burkle, L.A.; Dalla Riva, G.V.; Fortin, M.J.; Gravel, D.; Guimarães, P.R., Jr.; Hembry, D.H.; Newman, E.A.; et al. Analysing ecological networks of species interactions: Analyzing ecological networks. Biol. Rev. 2019, 94, 16–36. [Google Scholar] [CrossRef] [PubMed]
- Poisot, T.; Bergeron, G.; Cazelles, K.; Dallas, T.; Gravel, D.; MacDonald, A.; Mercier, B.; Violet, C.; Vissault, S.; Chapman, D. Global knowledge gaps in species interaction networks data. J. Biogeogr. 2021, 48, 1552–1563. [Google Scholar] [CrossRef]
- Guimarães, P.R., Jr. The Structure of Ecological Networks Across Levels of Organization. Annu. Rev. Ecol. Evol. Syst. 2022, 51, 433–460. [Google Scholar] [CrossRef]
- Lee, L.; Tan, D.J.X.; Oboňa, J.; Gustafsson, D.R.; Ang, Y.; Meier, R. Hitchhiking into the future on a fly: Toward a better understanding of phoresy and avian louse evolution (Phthiraptera) by screening bird carcasses for phoretic lice on hippoboscid flies (Diptera). Syst. Entomol. 2022, 47, 420–429. [Google Scholar] [CrossRef]
- Dick, C.W.; Esbérard, C.E.L.; Graciolli, G.; Bergallo, H.G.; Gettinger, D. Assessing host specificity of obligate ectoparasites in the absence of dispersal barriers. Parasitol. Res. 2009, 105, 1345–1349. [Google Scholar] [CrossRef]
- Barbier, E.; Bernard, E. From the Atlantic Forest to the borders of Amazonia: Species richness, distribution, and host association of ectoparasitic flies (Diptera: Nycteribiidae and Streblidae) in northeastern Brazil. Parasitol. Res. 2017, 116, 3043–3055. [Google Scholar] [CrossRef]
- Barbier, E.; Graciolli, G.; Bernard, E. Structure and composition of Nycteribiidae and Streblidae flies on bats along an environmental gradient in northeastern Brazil. Can. J. Zool. 2019, 97, 409–418. [Google Scholar] [CrossRef]
- Júnior, L.D.F.; De Araújo, W.S.; Falcão, L.A. Structure of the interaction networks between bats (Mammalia: Chiroptera) and ectoparasite flies (Diptera: Streblidae, Nycteribiidae) on a latitudinal gradient. Acta Chiropterologica 2020, 22, 187–196. [Google Scholar] [CrossRef]
- Lim, Z.X.; Hitch, A.T.; Lee, B.P.H.; Low, D.H.; Neves, E.S.; Borthwick, S.A.; Smith, G.J.; Mendenhall, I.H. Ecology of bat flies in Singapore: A study on the diversity, infestation bias and host specificity (Diptera: Nycteribiidae). Int. J. Parasitol. Parasites Wildl. 2020, 12, 29–33. [Google Scholar] [CrossRef]
- Palheta, L.R.; Urbieta, G.L.; Brasil, L.S.; Dias-Silva, K.; Da Silva, J.B.; Graciolli, G.; Aguiar, L.; Vieira, T.B. The effect of urbanization on bats and communities of bat flies (Diptera: Nycteribiidae and Streblidae) in the Amazon, northern Brazil. Acta Chiropterologica 2020, 22, 403–416. [Google Scholar] [CrossRef]
- Dolabela Falcão, L.A.; Araújo, W.S.; Leite, L.O.; Fagundes, M.; Espírito-Santo, M.M.; Zazá-Borges, M.A.; Vasconcelos, P.; Fernandes, G.W.; Paglia, A. Network Structure of Bat-Ectoparasitic Interactions in Tropical Dry Forests at Two Different Regions in Brazil. Acta Chiropterologica 2022, 24, 239–248. [Google Scholar] [CrossRef]
- Makovetskaya, Y.V.; Larchenka, A.I. To the knowledge of bat-flies fauna (Diptera: Nycteribiidae) of Belarus. Aktual. Probl. Ekol. 2020, 8, 51–52. [Google Scholar]
- Péter, Á.; Mihalca, A.D.; Sándor, A.D. First report of the bat fly species Basilia italica in Romania. Biodivers. Data J. 2021, 9, e57680. [Google Scholar] [CrossRef] [PubMed]
- Hůrka, K. Revision der Nycteribiidae und Streblidae-Nycteriboscinae aus der Dipterensammlung des Zoologischen Museums in Berlin, II. Mit Beschreibung von Basilia (Basilia) mediterranea n.sp. Mitt. Aus Dem Zool. Mus. Berl. 1970, 46, 239–246. [Google Scholar]
- Hůrka, K. Basilia mongolensis nudior subsp. n. nebst Bemerkungen zur Nycteribien-und Streblidenfauna Thrakiens (Diptera: Pupipara). Ann. Des Nat. Mus. Wien. 1972, 4, 709–713. [Google Scholar]
- Falcoz, L. Pupipara (Diptères) (première série). Biospeologica XLIX. Arch. De Zool. Expérimentale Et Générale 1923, 61, 521–552. [Google Scholar]
- Hůrka, K. Čeleď Nycteribiidae–muchulovití. In Krevsajíci Mouchy a Střečci–Diptera; Chvála, M., Ed.; Fauna ČSSR, 22; Academia, Nakladatelství ČSAV Praha: Praha, Czech Republic, 1980; pp. 479–509. [Google Scholar]
- Theodor, O. An Illustrated Catalogue of the Rothschild Collection of Nycteribiidae in the British Museum (Natural History), with Keys and Short Descriptions for the Identification of Subfamilies, Genera, Species and Subspecies; British Museum (Natural History): London, UK, 1967; 506p. [Google Scholar]
- Aellen, V. Etude d’une collection de Nycteribiidae et de Streblidae (Diptera: Pupipara) de la région paléarctique occidentale, particulièrement de la Suisse. Bull. De La Société Neuchâteloise Des Sci. Nat. 1955, 78, 81–104. [Google Scholar]
- Aellen, V. Les nycteribiidés de la Suisse, diptères parasites de chauves-souris. Bull. De La Société Neuchâteloise Des Sci. Nat. 1963, 86, 143–154. [Google Scholar]
- Austen, E.E. Diptera (Flies). In The Victoria History of the County of Devon; Victoria County History: London, UK, 1906; Volume 1, 630p. [Google Scholar]
- Baagøe, H.J. Eptesicus serotinus (Schreber, 1774) Breitflügelfledermaus. In Die Fledermäuse Europas. Ein umfassendes Handbuch zur Biologie, Verbreitung und Bestimmung; Krapp, F., Niethammer, J., Eds.; Aula–Verlag: Wiebelsheim, Germany, 2011; pp. 519–559. [Google Scholar]
- Baagøe, H.J. Myotis bechsteinii (Kuhl, 1818) Bechsteinfledermaus. In Die Fledermäuse Europas. Ein umfassendes Handbuch zur Biologie, Verbreitung und Bestimmung; Krapp, F., Niethammer, J., Eds.; Aula–Verlag: Wiebelsheim, Germany, 2011; pp. 443–471. [Google Scholar]
- Balcells, R.E. Interesantes datos faunísticos y biológicos de la cueva “Sa Guitarreta” de Llucmajoro. Bol. De La Soc. De Hist. Nat. De Balear. 1968, 14, 3–4. [Google Scholar]
- Balcells, R.E. Nuevas citas de murciélagos y nicteríbidos del país vasco-cantábrico. Real. Soc. Española De Hist. Nat. 1968, 66, 17–38. [Google Scholar]
- Bauer, K. Zur Kenntnis der Fledermausfauna Spaniens. Bonn. Zool. Beiträge 1957, 7, 296–319. [Google Scholar]
- Beaucournu, J.C. Ectoparasites des Chiroptères de l’Ouest de la France. 1re partie: Ixodoïdés—Cimicides et Nyctéribiidés. Bull. De La Société Sci. De Bretagne 1961, 36, 315–338. [Google Scholar]
- Beaucournu, J.C. Sur quelques ectoparasites (Siphonaptères, anoploures et nyctéribies) du Portugal. An. Da Esc. Nac. De Saúde Pública E De Med. Trop. 1972, 6, 65–71. [Google Scholar]
- Beaucournu, J.C.; Noblet, J.F. Une nyctéribie (Diptera, Pupipara) nouvelle pour la fauna française: Présence de Basilia mediterranea Hůrka, 1970 en Corse. Ann. De Parasitol. Hum. Et Comparée 1985, 60, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Beaucournu, J.C.; Noblet, J.F. Présence en France continentale de Basilia mediterranea Hůrka, 1970 (Diptera, Nycteribiidae). Bull. De La Société Entomol. De Fr. 1994, 99, 397–400. [Google Scholar] [CrossRef]
- Beaucournu, J.C.; Noblet, J.F. Les Diptères pupipares parasites de chauves-souris dans les Alpes et les Préalpes françaises (Diptera, Streblidae et Nycteribiidae). Bull. De La Société Entomol. De Fr. 1996, 101, 235–240. [Google Scholar] [CrossRef]
- Bequaert, J.G. Exploration du Parc National Albert. Mission, G.F. de Witte (1933–1935). Hippoboscidae and Nycteribiidae. Inst. Des Parcs Natx. Belg. 1953, 79, 89–94. [Google Scholar]
- Czuppon, B. Denevérlegyek (Nycteribiidae) Populáció-Ökológiai Elemzése. Doctoral Thesis, Szent István University, Budapest, Hungary, 2001; 48p. [Google Scholar]
- Czuppon, B.; Molnár, V. Bat fly genus and species new to Hungary (Diptera: Nycteribiidae). Folia Entomol. Hung. 2001, 62, 313–317. [Google Scholar]
- Colyer, C.N.; Hammond, C.O. Flies of the British Isles; Frederick Warne: London, UK, 1951; 383p. [Google Scholar]
- Danko, Š.; Krištín, A.; Krištofík, J. Myotis alcathoe in eastern Slovakia: Occurrence, diet, ectoparasites and notes on its identification in the field. Vespertilio 2010, 13–14, 77–91. [Google Scholar]
- Dietz, M.; Walter, G. Zur Ektoparasitenfauna der Wasserfledermaus (Myotis daubentoni Kuhl, 1819) in Deutschland unter besonderer Berücksichtigung der Flughautmilbe Spinturnix andegavinus Deunff, 1977. Nyctalus (Neue Folge) 1995, 5, 451–468. [Google Scholar]
- Dudich, E. A magyarországi denevérlegyek. Math. És Természettudományi Értesítő 1925, 41, 144–151. [Google Scholar]
- Estrada-Peña, A.; Serra-Cobo, J. The acarinia and nycteribidia zones of Miniopterus schreibersii Kuhl (Mammalia: Chiroptera) in the northeast of Spain. Folia Parasitol. 1991, 38, 345–354. [Google Scholar]
- Falcoz, L. Diptères Pupipares du Muséum National d’histoire naturelle de Paris. Muséum Natl. D’histoire Nat. 1924, 30, 223. [Google Scholar]
- Falcoz, L. Diptères Pupipares. Fauna de France, 14; Fédération Française des Sociétés de Sciences Naturelles: Paris, France, 1926; 64p. [Google Scholar]
- Gil Collado, J. Una nueva especie española de Nycteribia. Eos: Rev. Española De Entomol. 1934, 9, 29–32. [Google Scholar]
- Grulich, I.; Povolný, D. Faunisticko–bionomický nástin muchulovitých (Nycteribiidae) na území ČSR. Zool. A Entomol. Listy 1955, 4, 111–134. [Google Scholar]
- Güttinger, R.; Zahn, A.; Krapp, F.; Schober, W. Myotis myotis (Borkhausen, 1797) Großes Mausohr. In Die Fledermäuse Europas; Ein umfassendes Handbuch zur Biologie, Verbreitung und Bestimmung; Krapp, F., Niethammer, J., Eds.; Aula–Verlag: Wiebelsheim, Germany, 2011; pp. 123–207. [Google Scholar]
- Haitlinger, R. Pasożyty zewnętrzne nietoperzy Dolnego Śląska. II. Nycteribiidae (Diptera). Wiadomości Parazytol. 1978, 24, 467–474. [Google Scholar]
- Haitlinger, R.; Ruprecht, A.L. Parasitic arthropods (Siphonaptera, Diptera, Acari) of bats from western part of the Bialowieża Primeval Forest. Nyctalus (Neue Folge) 1992, 4, 315–319. [Google Scholar]
- Heddergott, M. Erstnachweis der Fledermausfliege Basilia mongolensis nudior Hůrka, 1972 in Deutschland (Diptera: Nycteribiidae). Stud. Dipterol. 2009, 15, 301–304. [Google Scholar]
- Heddergott, M.; Claußen, A. Nachweise von Fledermausfliegen aus dem Nationalpark Hainich in Thüringen (Diptera: Calyptrata: Nycteribiidae). Abh. Ber. Mus. Nat. Gotha 2004, 23, 91–93. [Google Scholar]
- Humphries, D.A. Caves and bats. J. King’s Coll. Nat. Hist. Soc. 1959, 1, 18–21. [Google Scholar]
- Hůrka, K. Beitrag zur Nycteribien–und Streblidenfauna Albaniens nebst Bemerkungen zur Fauna von Bulgarien, Ungarn und UdSSR. Acta Soc. Entomol. Cechoslov. 1962, 59, 156–164. [Google Scholar]
- Hůrka, K. Distribution, bionomy and ecology of the European bat flies with special regard to the Czechoslovak fauna (Diptera, Nycteribidae). Acta Univ. Carolinae. Biol. 1964, 3, 167–231. [Google Scholar]
- Hůrka, K. New taxa and new records of Palearctic Nycteribiidae and Streblidae (Diptera: Pupipara). Věstník Československé Společnosti Zool. 1984, 48, 90–101. [Google Scholar]
- Hutson, A.M. Ectoparasites of British bats. Mammal. Rev. 1972, 1, 143–150. [Google Scholar] [CrossRef]
- Hutson, A.M. Keds, Flat–flies and Bat–flies (Diptera, Hippoboscidae and Nycteribiidae). In Handbooks for the Identification of British Insects; Royal Entomological Society of London: London, UK, 1984; 44p. [Google Scholar]
- Imaz, E.; Aihartza, J.R.; Totorika, M.J. Ectoparasites on bats (Gamasida, Ixodida, Diptera) in Biscay (N Iberian peninsula). Miscel·Lània Zoològica 1999, 22, 21–30. [Google Scholar]
- Jaunbauere, G.; Salmane, I.; Spungis, V. Occurrence of bat ectoparasites in Latvia. Latv. Entomol. 2008, 45, 38–42. [Google Scholar]
- Jobling, B. A revision of the genus Nycteribosca. Parasitology 1934, 26, 64–97. [Google Scholar] [CrossRef]
- Kaňuch, P.; Krištín, A.; Krištofík, J. Phenology, diet, and ectoparasites of Leisler’s bat (Nyctalus leisleri) in the Western Carpathians (Slovakia). Acta Chiropterologica 2005, 7, 249–257. [Google Scholar] [CrossRef]
- Kock, D. Über Nycteribiiden im deutschen Faunengebiet (Ins.: Diptera). Senckenberg. Biol. 1973, 54, 343–352. [Google Scholar]
- Kock, D. Pupipare Dipteren von Säugetieren des nordöstlichen Mittelmeerraumens (Ins.: Diptera). Senckenberg. Biol. 1974, 17, 87–104. [Google Scholar]
- Kock, D. Basilia mongolensis nudior Hůrka 1972 in Österreich (Insecta: Diptera: Nycteribiidae). Senckenberg. Biol. 1984, 65, 43–46. [Google Scholar]
- Kock, D. Fledermaus–Fliegen aus der E–Mediterraneis (Diptera: Nyeteribiidae). Entomol. Z. 1989, 99, 56–58. [Google Scholar]
- Kock, D. Die Fledermauslausfliegen Bayerns (Diptera: Nycteribidae). Entomol. Z. 1999, 109, 444–447. [Google Scholar]
- Kock, D. Bestandsdichte der Wasserfledermaus, Myotis daubentonii Kuhl, 1817 (Mammalia: Chiroptera) und zunehmende Verbreitung ihres Parasiten, Penicillidia monoceros Speiser, 1900 (Diptera: Nycteribiidae) in Deutschland. Myotis 2004, 41–42, 99–107. [Google Scholar]
- Kock, D.; Quetglas, J. The bat flies of the Balearic Islands (Insecta: Diptera: Nycteribiidae). Bolletí De La Soc. D’història Nat. De Les. Balear. 2003, 46, 79–83. [Google Scholar]
- Krištofík, J. Nálezy múch čeľade Nycteribiidae (Diptera) na území SSR. Biol. Bratisl. 1982, 37, 191–197. [Google Scholar]
- Krištofík, J.; Danko, S. Arthropod ectoparasites (Acarina, Heteroptera, Diptera, Siphonaptera) of bats in Slovakia. Vespertilio 2012, 16, 167–189. [Google Scholar]
- Lindecke, O.; Scheffler, I. Ectoparasites of bats in Saxony–Anhalt. Hercynia Neuen Folge 2011, 44, 241–251. [Google Scholar]
- Masson, D. Sur l’infestation de Myotis nattereri (Kuhl, 1818) (Chiroptera, Vespertilionidae) par Basilia nattereri (Kolenati, 1857) (Diptera, Nycteribiidae) dans le Sud–Ouest de la France. Ann. De Parasitol. Hum. Et Comparée 1989, 64, 64–71. [Google Scholar] [CrossRef]
- Noblet, J.F. Els quiròpters del Parc Natural de s‘Albufera de Mallorca. Monogr. De La Soc. D’història Nat. De Les. Balear. 1995, 4, 169–173. [Google Scholar]
- Nowosad, A. Nycteribia kolenatii Theodor et Moscona i Penicillidia monoceros Speiser (Nycteribiidae, Diptera) w Polsce. Pol. Pismo Entomol. 1974, 44, 559–569. [Google Scholar]
- Nowosad, A. Materialy do znajomoéci Nycteribiidae (Diptera, Pupipara) Polski zachodniej i pohiocno-zachodniej. Pol. Pismo Entomol. 1987, 57, 263–276. [Google Scholar]
- Nowosad, A. Stan badañ nad mrokawkowatymi—Nycteribiidae (Diptera, Pupipara) w Polsce, z przeglądem gatunków i stanowisk ich występowania. Wiadomości Entomol. 1990, 9, 77–85. [Google Scholar]
- Nowosad, A.; Batchvarov, G.; Petrov, P. Bat flies (Nycteribiidae, Diptera) of bats collected in Bulgaria. Pol. Pismo Entomol. 1987, 57, 673–694. [Google Scholar]
- Orlova, M.V. Ectoparasite associations of bats from the Urals (Russia). Hystrix Ital. J. Mammal. 2011, 22, 105–110. [Google Scholar]
- Rageau, J.; Mouchet, J. Les arthropodes hématophages de Camargue. Cah. ORSTOM. Série Entomol. Médicale Et Parasitol. 1967, 5, 263–281. [Google Scholar]
- Reckardt, K.; Kerth, G. The reproductive success of the parasitic bat fly Basilia nana (Diptera: Nycteribiidae) is affected by the low roost fidelity of its host, the Bechstein’s bat (Myotis bechsteinii). Parasitol. Res. 2006, 93, 237–243. [Google Scholar] [CrossRef]
- Rupp, D. Zweiflügler aus Bayern XVIQ (Diptera: Nycteribiidae). Entomofauna 1999, 25, 397–400. [Google Scholar]
- Scheffler, I. Untersuchungen zur Ektoparasitenfauna (Siphonaptera: Ischnopsyllidae; Diptera: Nycteribiidae; Heteroptera: Cimicidae) an Fledermäusen (Teil 3). Märkische Entomol. Nachr. 2008, 10, 241–248. [Google Scholar]
- Scheffler, I. Ektoparasiten der Fledermäuse in Sommerquartieren in Brandenburg: Neue Funde seltener Arten. Nyctalus (Neue Folge) 2009, 14, 126–136. [Google Scholar]
- Scheffler, I. Ektoparasiten der Fledermäuse in Winterquartieren in Brandenburg. Märkische Entomol. Nachr. 2010, 12, 119–132. [Google Scholar]
- Scheffler, I. Die Ektoparasiten der Fledermäuse Europas—Teil 1. Nyctalus (Neue Folge) 2011, 16, 246–263. [Google Scholar]
- Scheffler, I. Artenspektren und Wirtsbindung von Ektoparasiten der Fledermäuse aus Nordbulgarien. Bewertung des Zusammenhangs von Körperkondition und Ektoparasitenlast. Nyctalus (Neue Folge) 2011, 16, 119–136. [Google Scholar]
- Scheffler, I. Die Ektoparasiten der Fledermäuse Europas—Teil 2. Nyctalus (Neue Folge) 2012, 17, 104–119. [Google Scholar]
- Scheffler, I.; Bego, F.; Théou, P.; Podany, M.; Pospischil, R.; Hübner, S.; Wittenberg, L. Ektoparasiten der Fledermause in Albanien–Artenspektrum und Wirtsbindung. Nyctalus (Neue Folge) 2013, 18, 84–109. [Google Scholar]
- Scheffler, I.; Hiller, A. Zur Ektoparasitenfauna der Fledermäuse in Niedersachsen: Neue Funde am Iberg bei Bad Grund. Nyctalus (Neue Folge) 2010, 15, 309–317. [Google Scholar]
- Scheffler, I.; Ressler, R. Untersuchungen zur Ektoparasitenfauna (Siphonaptera: Ischnopsyllidae; Diptera: Nycteribiidae) an Fledermäusen in Brandenburg. Märkische Entomol. Nachr. 2005, 7, 123–132. [Google Scholar]
- Scheffler, I.; Ressler, R. Untersuchungen zur Ektoparasitenfauna (Siphonaptera: Ischnopsyllidae; Diptera: Nycteribiidae; Heteroptera: Cimicidae) an Fledermäusen in Brandenburg (Teil 2). Märkische Entomol. Nachr. 2007, 9, 109–119. [Google Scholar]
- Scott, H. Descriptions and records of Nycteribiidae, with a discussion of the genus Basilia. J. Linn. Soc. Lond. Zool. 1936, 39, 479–505. [Google Scholar] [CrossRef]
- Skuratowicz, W. Przyczynek do do znajomości Nycteribiidae (Diptera, Pupipara) Polski. Fragm. Faun. 1962, 10, 215–221. [Google Scholar] [CrossRef]
- Ševčík, M.; Benda, P.; Uhrin, M. First records of the bat fly Phthiridium biarticulatum (Diptera: Nycteribiidae) in Crimea. Vespertilio 2011, 15, 159–160. [Google Scholar]
- Soós, Á. Bábtojó legyek—Muscidae Pupiparae; Magyarország Állatvilága—Fauna hungarie; Akadémia Kiadó: Budapest, Hungary, 1955; 20p. [Google Scholar]
- Szentiványi, T.; Genzoni, E.; Clément, L.; Radonjić, M.; Loce, E.; Théou, P.; Glaizot, O.; Christe, P. Basilia: A new genus to the Albanian bat fly fauna (Diptera: Nycteribiidae). Ecol. Montenegrina 2016, 8, 58–61. [Google Scholar] [CrossRef]
- Theodor, O. On the genus Tripselia and the group of Basilia bathybothyra (Nycteribiidae, Diptera). Parasitology 1956, 46, 353–394. [Google Scholar] [CrossRef] [PubMed]
- Theodor, O.; Moscona, A. On bat parasites in Palestine. I. Nycteribiidae, Streblidae, Hemiptera, Siphonaptera. Parasitology 1954, 44, 157–245. [Google Scholar] [CrossRef]
- Topál, G. Myotis nattereri (Kuhl, 1818) Fransenfledermaus. In Die Fledermäuse Europas. Ein umfassendes Handbuch zur Biologie, Verbreitung und Bestimmung; Krapp, F., Niethammer, J., Eds.; Aula–Verlag: Wiebelsheim, Germany, 2011; pp. 405–442. [Google Scholar]
- Uhrin, M.; Kaňuch, P.; Krištofík, J.; Paule, L. Phenotypic plasticity in the greater mouse–eared bat in extremely different roost conditions. Acta Theriol. 2010, 55, 153–164. [Google Scholar] [CrossRef]
- Walter, G. Nachweise von Nycteribia kolenatii (Diptera, Nycteribiidae) für die Bundesrepublik Deutschland und Berlin (West). Angew. Parasitol. 1987, 28, 177–178. [Google Scholar]
- Walter, G. Überblick zum Vorkommen und zur Biologie von Ektoparasiten (Siphonaptera; Cimicidae; Nycteribiidae; Calliphoridae) bei Fledermäusen in Deutschland. Nyctalus (Neue Folge) 2004, 9, 460–476. [Google Scholar]
- Walter, G.; Benk, A. Zur Ektoparasitenfauna der Fledermäuse (Chiroptera) in Niedersachsen. Angew. Parasitol. 1982, 23, 230–232. [Google Scholar]
- Walter, G.; Ebenau, C. Nachweise von Fledermausfliegen aus Syrien (Diptera: Streblidae, Nycteribiidae). Zool. Middle East. 1997, 14, 115–119. [Google Scholar] [CrossRef]
- Westwood, J.O. On Nycteribia, a genus of wingless insects. Zool. Soc. Lond. 1835, 1, 275–294. [Google Scholar] [CrossRef]
- Müller, J.; Ohlendorf, B. Erstnachweis von Fledermausfliegen (Dipt., Nycteribiidae) aus dem Harz, DDR–Bezirk Magdeburg. Nyctalus (Neue Folge) 1984, 2, 81–84. [Google Scholar]
- Dondini, G.; Vanin, S.; Vergari, S.; Vergari, S. First record of Basilia mediterranea Hůrka, 1970 from Italy (Diptera: Nycteribiidae). Onychium 2017, 13, 139–142. [Google Scholar]
- Haelewaters, D.; Pfliegler, W.P.; Szentiványi, T.; Földvári, M.; Sándor, A.D.; Barti, L.; Pfister, D.H. Parasites of parasites of bats: Laboulbeniales (Fungi: Ascomycota) on bat flies (Diptera: Nycteribiidae) in central Europe. Parasites Vectors 2017, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kemenesi, G.; Tóth, G.E.; Mayora-Neto, M.; Scott, S.; Temperton, N.; Wright, E.; Jakab, F. Isolation of infectious Lloviu virus from Schreiber’s bats in Hungary. Nat. Commun. 2022, 13, 1706. [Google Scholar] [CrossRef] [PubMed]
- McKee, C.D.; Krawczyk, A.I.; Sándor, A.D.; Görföl, T.; Földvári, M.; Földvári, G.; Sprong, H. Host phylogeny, geographic overlap, and roost sharing shape parasite communities in European bats. Front. Ecol. Evol. 2019, 7, 69. [Google Scholar] [CrossRef]
- Pejić, B.; Budinski, I.; Blagojević, J. Ectoparasite Bat Flies (Diptera: Nycteribiidae) of Schreiber’s Bent-Winged Bat and Their Fungus Parasite. In Programme & Abstract Book, Proceedings of the 13th European Multicolloquium of Parasitology, Belgrade, Serbia, 12–16 October 2021. Available online: https://emop2020.org/ (accessed on 27 February 2023).
- Sándor, A.D.; Földvári, M.; Krawczyk, A.I.; Sprong, H.; Corduneanu, A.; Barti, L.; Földvári, G. Eco-epidemiology of novel Bartonella genotypes from parasitic flies of insectivorous bats. Microb. Ecol. 2018, 76, 1076–1088. [Google Scholar] [CrossRef] [PubMed]
- Sándor, A.D.; Péter, Á.; Corduneanu, A.; Barti, L.; Csősz, I.; Kalmár, Z.; Mihalca, A.D. Wide distribution and diversity of malaria-related haemosporidian parasites (Polychromophilus spp.) in bats and their ectoparasites in Eastern Europe. Microorganisms 2021, 9, 230. [Google Scholar] [CrossRef] [PubMed]
- Szentiványi, T.; Estók, P.; Pigeault, R.; Christe, P.; Glaizot, O. Effects of fungal infection on the survival of parasitic bat flies. Parasites Vectors 2020, 13, 1–8. [Google Scholar] [CrossRef]
- Szentiványi, T.; Haelewaters, D.; Pfliegler, W.P.; Clément, L.; Christe, P.; Glaizot, O. Laboulbeniales (Fungi: Ascomycota) infection of bat flies (Diptera: Nycteribiidae) from Miniopterus schreibersii across Europe. Parasites Vectors 2018, 11, 395. [Google Scholar] [CrossRef]
- Szentiványi, T.; Haelewaters, D.; Rádai, Z.; Mizsei, E.; Pfliegler, W.P.; Báthori, F.; Glaizot, O. Climatic effects on the distribution of ant-and bat fly-associated fungal ectoparasites (Ascomycota, Laboulbeniales). Fungal Ecol. 2019, 39, 371–379. [Google Scholar] [CrossRef]
- Szentiványi, T.; Vincze, O.; Estók, P. Density-dependent sex ratio and sex-specific preference for host traits in parasitic bat flies. Parasites Vectors 2017, 10, 405. [Google Scholar] [CrossRef] [PubMed]
- Tendu, A.; Hughes, A.C.; Berthet, N.; Wong, G. Viral hyperparasitism in bat ectoparasites: Implications for pathogen maintenance and transmission. Microorganisms 2022, 10, 1230. [Google Scholar] [CrossRef] [PubMed]
- Theodor, O. Nycteribiidae. In Die Fliegen der Paläarktischen Region; Lindner, E., Ed.; Band 12 [Lieferung 174]. E.; Schweizerbart: Stuttgart, Germany, 1954; 43p. [Google Scholar]
- Caira, J.N.; Jensen, K.; Holsinger, K.I. On a new index of host specificity. In Taxonomie, Écologie et Évolution des Metazoaires Parasites; Combes, C., Jourdane, J., Eds.; Presses Universitaires de Perpignan: Perpignan, France, 2003; pp. 161–201. [Google Scholar]
- Marciniak-Musial, N.; Skoracki, M.; Kosicki, J.Z.; Unsöld, M.; Sikora, B. Host-Parasite Relationships of Quill Mites (Syringophilidae) and Parrots (Psittaciformes). Diversity 2022, 15, 1. [Google Scholar] [CrossRef]
- Gruber, B. Plotweb: Visualize a Bipartite Interaction Matrix (e.g., a foodweb). Available online: https://www.rdocumentation.org/packages/bipartite/versions/2.18/topics/plotweb (accessed on 18 March 2023).
- Strauss, R. plotModuleWeb: plotModuleWeb. Available online: https://www.rdocumentation.org/packages/bipartite/versions/2.18/topics/plotModuleWeb (accessed on 18 March 2023).
- Dormann, F.; Gruber, B.; Fründ, J. Introducing the bipartite package: Analysing ecological networks. R News 2008, 8, 8–11. [Google Scholar]
- May, R.M. Stability and Complexity in Model. Ecosystems, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 1974. [Google Scholar]
- Dormann, C.F. Networklevel: Analysis of Bipartite Webs at the Level of the Entire Network. Available online: https://www.rdocumentation.org/packages/bipartite/versions/2.18/topics/networklevel (accessed on 19 March 2023).
- Blüthgen, N. Why network analysis is often disconnected from community ecology: A critique and an ecologist’s guide. Basic Appl. Ecol. 2010, 11, 185–195. [Google Scholar] [CrossRef]
- Dormann, C.F.; Fründ, J. H2fun: Specialisation of a Bipartite Web. Available online: https://www.rdocumentation.org/packages/bipartite/versions/2.18/topics/H2fun (accessed on 19 March 2023).
- Atmar, W.; Patterson, B.D. The measure of order and disorder in the distribution of species in fragmented habitat. Oecologia 1993, 96, 373–382. [Google Scholar]
- Bascompte, J.; Jordano, P.; Melia, C.J.; Olesen, J.M. The nested assembly of plant–animal mutualistic networks. Proc. Natl. Acad. Sci. USA 2003, 100, 9383–9387. [Google Scholar] [CrossRef]
- Pavlopoulos, G.A.; Kontou, P.I.; Pavlopoulou, A.; Bouyioukos, C.; Markou, E.; Bagos, P.G. Bipartite graphs in systems biology and medicine: A survey of methods and applications. GigaScience 2018, 7, giy014. [Google Scholar] [CrossRef]
- Fortuna, M.A.; Stouffer, D.B.; Olesen, J.M.; Jordano, P.; Mouillot, D.; Krasnov, B.R.; Poulin, R.; Bascompte, J. Nestedness versus modularity in ecological networks: Two sides of the same coin? J. Anim. Ecol. 2010, 79, 811–817. [Google Scholar] [CrossRef]
- Rodriguez-Girones, M.A.; Santamaria, L. A new algorithm to calculate the nestedness temperature of presence-absence matrices. J. Biogeogr. 2006, 33, 924–935. [Google Scholar]
- Newman, M.E.J. The structure and function of complex networks. SIAM Rev. 2003, 45, 167–256. [Google Scholar] [CrossRef]
- Newman, M.E.J.; Girvan, M. Finding and evaluating community structure in networks. Phys. Rev. 2004, 69, 026113. [Google Scholar] [CrossRef] [PubMed]
- Fortunato, S. Community detection in graphs. Phys. Rep. 2010, 486, 75–174. [Google Scholar] [CrossRef]
- Strauss, R.; Dormann, C.; Hegemann, T. Computemodules: Computemodules. Available online: https://www.rdocumentation.org/packages/bipartite/versions/2.18/topics/computeModules (accessed on 19 March 2023).
- Blüthgen, N.; Menzel, F.; Bliithgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 9, 6–9. [Google Scholar]
- Fründ, J.; Dormann, C.F. Dfun: Calculates Standardised Specialisation Index d’ (d prime) for Each Species in the Lower Trophic Level of a Bipartite Network. Available online: https://www.rdocumentation.org/packages/bipartite/versions/2.18/topics/dfun (accessed on 20 March 2023).
- Dick, C.W.; Patterson, B.D. Against all odds: Explaining high host specificity in dispersal-prone parasites. Int. J. Parasitol. 2007, 37, 871–876. [Google Scholar] [CrossRef] [PubMed]
- de Groot, M.D.; Dumolein, I.; Hiller, T.; Sándor, A.D.; Szentiványi, T.; Schilthuizen, M.; Aime, M.C.; Verbeken, A.; Haelewaters, D. On the fly: Tritrophic associations of bats, bat flies, and fungi. J. Fungy 2020, 6, 361. [Google Scholar] [CrossRef]
- Szentiványi, T.; Heintz, A.C.; Markotter, W.; Wassef, J.; Christe, P.; Glaizot, O. Vector-borne protozoan and bacterial pathogen occurrence and diversity in ectoparasites of the Egyptian Rousette bat. Med. VEt Entomol. 2023, 1–6. [Google Scholar] [CrossRef]
- Szentiványi, T.; Markotter, W.; Dietrich, M.; Clément, L.; Ançay, L.; Brun, L.; Genzoni, E.; Kearney, T.; Seamark, E.; Estók, P.; et al. Host conservation through their parasites: Molecular surveillance of vector-borne microorganisms in bats using ectoparasitic bat flies. Conservation des hôtes grâce à leurs parasites: Surveillance moléculaire des microorganismes à transmission vectorielle chez les chauves-souris à l’aide de mouches ectoparasites. Parasite 2020, 27, 72. [Google Scholar]
- Zemanová, S.; Korytár, Ľ.; Tomčová, J.; Prokeš, M.; Drážovská, M.; Myczko, Ł.; Tryjanowski, P.; Nusová, G.; Matysiak, A.; Ondrejková, A. Opportunities and Limitations of Molecular Methods for Studying Bat-Associated Pathogens. Microorganisms 2022, 10, 1875. [Google Scholar] [CrossRef]
- Zeus, V.M.; Puechmaille, S.J.; Kerth, G. Conspecific and heterospecific social groups affect each other’s resource use: A study on roost sharing among bat colonies. Anim. Behav. 2017, 123, 329–338. [Google Scholar]
- Zahn, A.; Rupp, D. Ectoparasite load in European vespertilionid bats. J. Zool. 2004, 262, 383–391. [Google Scholar] [CrossRef]
- Christe, P.; Glaizot, O.; Evanno, G.; Bruyndonckx, N.; Devevey, G.; Yannic, G.; Patthey, P.; Maeder, A.; Vogel, P.; Arlettaz, R. Host sex and ectoparasites choice: Preference for, and higher survival on female hosts. J. Anim. Ecol. 2007, 76, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Sándor, A.D.; Corduneanu, A.; Péter, Á.; Mihalca, A.D.; Barti, L.; Csősz, I.; Szőke, K.; Hornok, S. Bats and ticks: Host selection and seasonality of bat-specialist ticks in eastern Europe. Parasites Vectors 2019, 12, 605. [Google Scholar] [CrossRef] [PubMed]
- Mello, M.A.R.; Marquitti, F.M.D.; Guimarães, P.R., Jr.; Kalko, E.K.V.; Jordano, P.; de Aguiar, M.A.M. The missing part of seed dispersal networks: Structure and robustness of bat-fruit interactions. PLoS ONE 2011, 6, e17395. [Google Scholar] [CrossRef]
- Krasnov, B.R.; Fortuna, M.A.; Mouillot, D.; Khokhlova, I.S.; Shenbrot, G.I.; Poulin, R. Phylogenetic signal in module composition and species connectivity in compartmentalized host–parasite networks. Am. Nat. 2012, 179, 501–511. [Google Scholar] [CrossRef]
- Bellay, S.; de Oliveira, E.F.; Almeida-Neto, M.; Lima Junior, D.P.; Takemoto, R.M.; Luque, J.L. Developmental stage of parasites influences the structure of fish–parasite networks. PLoS ONE 2013, 8, e75710. [Google Scholar] [CrossRef]
- Brito, S.V.; Corso, G.; Almeida, A.M.; Ferreira, F.S.; Almeida, W.O.; Anjos, L.A.; Mesquita, D.O.; Vasconcellos, A. Phylogeny and micro-habitats utilized by lizards determine the composition of their endoparasites in the semiarid Caatinga of Northeast Brazil. Parasitol. Res. 2014, 113, 3963–3972. [Google Scholar] [CrossRef]
- Obame-Nkoghe, J.; Rahola, N.; Bourgarel, M.; Yangari, P.; Prugnolle, F.; Maganga, G.D.; Leroy, E.M.; Fontenille, D.; Ayala, D.; Paupy, C. Bat flies (Diptera: Nycteribiidae and Streblidae) infesting cave-dwelling bats in Gabon: Diversity, dynamics and potential role in Polychromophilus melanipherus transmission. Parasites Vectors 2016, 9, 1–12. [Google Scholar] [CrossRef]
- Poon, E.S.K.; Chen, G.; Tsang, H.Y.; Shek, C.T.; Tsui, W.C.; Zhao, H.; Guénard, B.; Sin, S.Y.W. Species richness of bat flies and their associations with host bats in a subtropical East Asian region. Parasites Vectors 2023, 16, 1–15. [Google Scholar] [CrossRef]
- Spitzenberger, F.; Weiss, E. Changes in roost occupancy and abundance in atticdwelling bats during decreasing roost availability in Burgenland, Austria. Vespertilio 2012, 16, 279–288. [Google Scholar]
- Bartonička, T.; Jedlička, P. First record of Schreiber’s bat, Miniopterus schreibersii (Kuhl, 1817), in the Czech Republic. Lynx (Praha) 2011, 42, 83–89. [Google Scholar]
- Pejić, B.; Budinski, I.; van Schaik, J.; Blagojević, J. Sharing roosts but not ectoparasites: High host-specificity in bat flies and wing mites of Miniopterus schreibersii and Rhinolophus ferrumequinum (Mammalia: Chiroptera). Curr. Zool. 2022, 68, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Morse, S.F.; Bush, S.E.; Patterson, B.D.; Dick, C.W.; Gruwell, M.E.; Dittmar, K. Evolution, multiple acquisition, and localization of endosymbionts in bat flies (Diptera: Hippoboscoidea: Streblidae and Nycteribiidae). Appl. Environ. Microbiol. 2013, 79, 2952–2961. [Google Scholar] [CrossRef] [PubMed]
- Urushadze, L.; Bai, Y.; Osikowicz, L.; McKee, C.; Sidamonidze, K.; Putkaradze, D.; Imnadze, P.; Kandaurov, A.; Kuzmin, I.; Kosoy, M. Prevalence, diversity, and host associations of Bartonella strains in bats from Georgia (Caucasus). PLoS Negl. Trop. Dis. 2017, 11, e0005428. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bat Fly Species | No. of Hosts spp. Published | d’ | Specificity |
|---|---|---|---|
| Basilia daganiae | 1 | 1.00 | monoxenous |
| Basilia italica | 9 | 0.50 | oligoxenous |
| Basilia mediterranea | 4 | 0.67 | stenoxenous |
| Basilia mongolensis nudior | 4 | 0.65 | stenoxenous |
| Basilia nana | 15 | 0.32 | oligoxenous |
| Basilia nattereri | 9 | 0.30 | oligoxenous |
| Nycteribia (Acrocholidia) vexata | 13 | 0.14 | oligoxenous |
| Nycteribia (Nycteribia) kolenatii | 17 | 0.30 | oligoxenous |
| Nycteribia (Nycteribia) latreillii | 14 | 0.18 | oligoxenous |
| Nycteribia (Nycteribia) pedicularia | 14 | 0.20 | oligoxenous |
| Nycteribia (Nycteribia) schmidlii | 16 | 0.18 | oligoxenous |
| Penicillidia (Penicillidia) conspicua | 13 | 0.24 | oligoxenous |
| Penicillidia (Penicillidia) dufourii dufourii | 17 | 0.19 | oligoxenous |
| Penicillidia (Penicillidia) monoceros | 8 | 0.41 | oligoxenous |
| Phthiridium biarticulatum | 16 | 0.35 | oligoxenous |
| Phthiridium integrum | 3 | 0.56 | stenoxenous |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mlynárová, L.; Korytár, Ľ.; Manko, P.; Ondrejková, A.; Prokeš, M.; Smoľák, R.; Oboňa, J. Updated Taxonomic Key of European Nycteribiidae (Diptera), with a Host-Parasite Network. Diversity 2023, 15, 573. https://doi.org/10.3390/d15040573
Mlynárová L, Korytár Ľ, Manko P, Ondrejková A, Prokeš M, Smoľák R, Oboňa J. Updated Taxonomic Key of European Nycteribiidae (Diptera), with a Host-Parasite Network. Diversity. 2023; 15(4):573. https://doi.org/10.3390/d15040573
Chicago/Turabian StyleMlynárová, Laura, Ľuboš Korytár, Peter Manko, Anna Ondrejková, Marián Prokeš, Radoslav Smoľák, and Jozef Oboňa. 2023. "Updated Taxonomic Key of European Nycteribiidae (Diptera), with a Host-Parasite Network" Diversity 15, no. 4: 573. https://doi.org/10.3390/d15040573