


Turnover and Natal Dispersal in the Finnish Golden Eagle (Aquila chrysaetos) Population

Abstract
:
1. Introduction
2. Materials and Methods
2.1. The Study Species
2.2. Sampling and Laboratory Analyses
2.3. Genotyping and Sequencing Quality
2.4. Data Analysis
2.4.1. Genetic Diversity and Population Genetic Structure
2.4.2. Turnover
2.4.3. Natal Dispersal
3. Results
3.1. Genotyping Quality
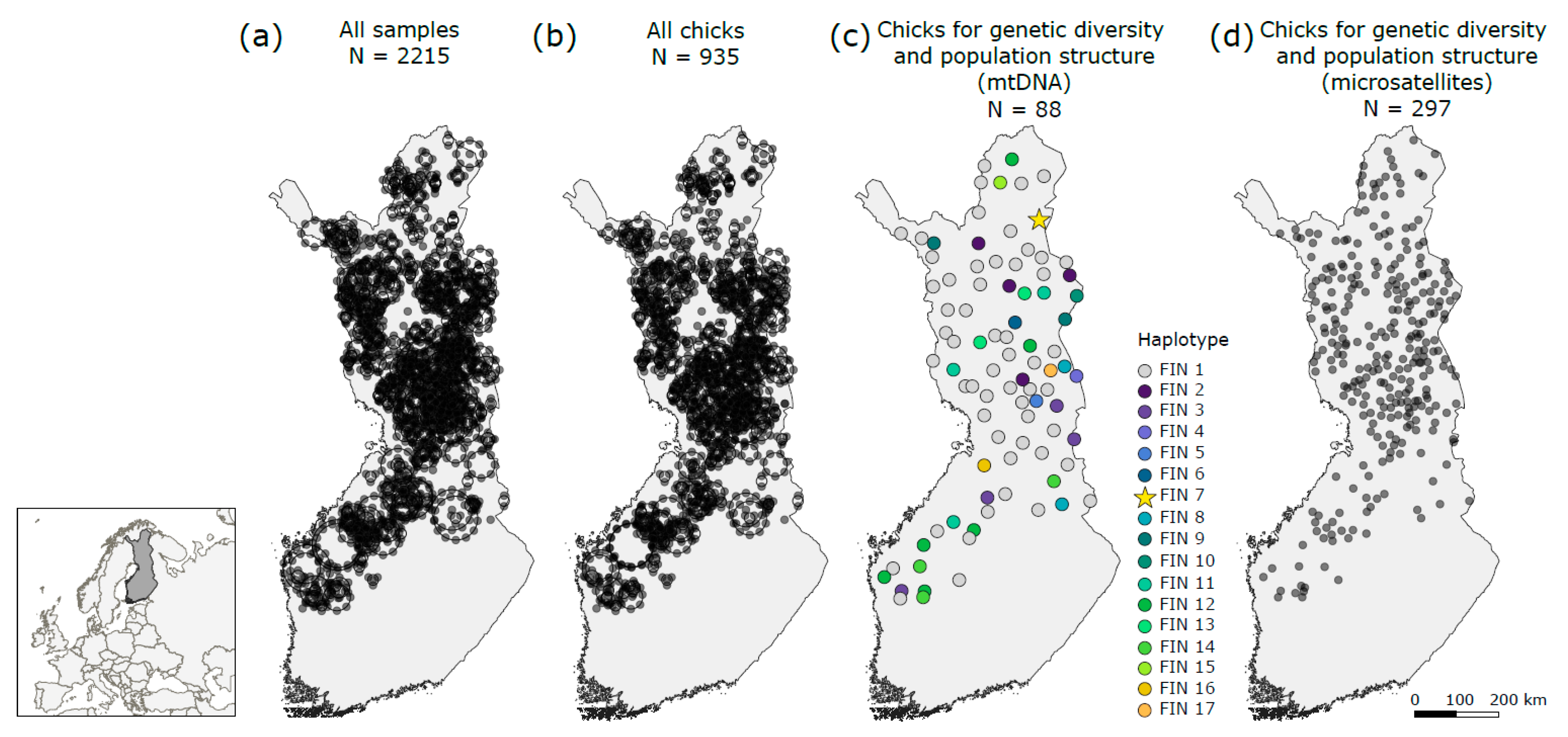
3.2. Genetic Diversity and Population Genetic Structure
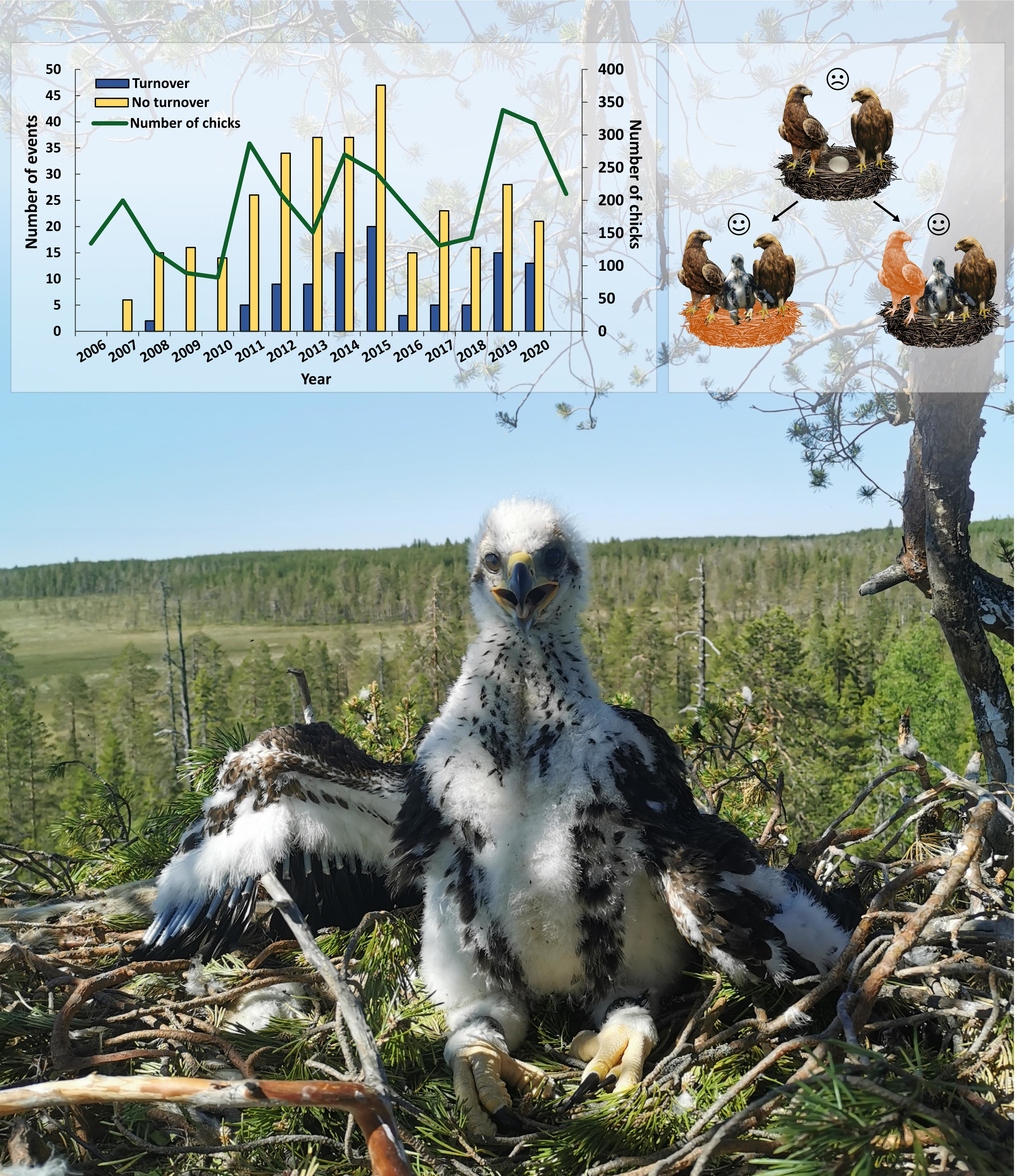
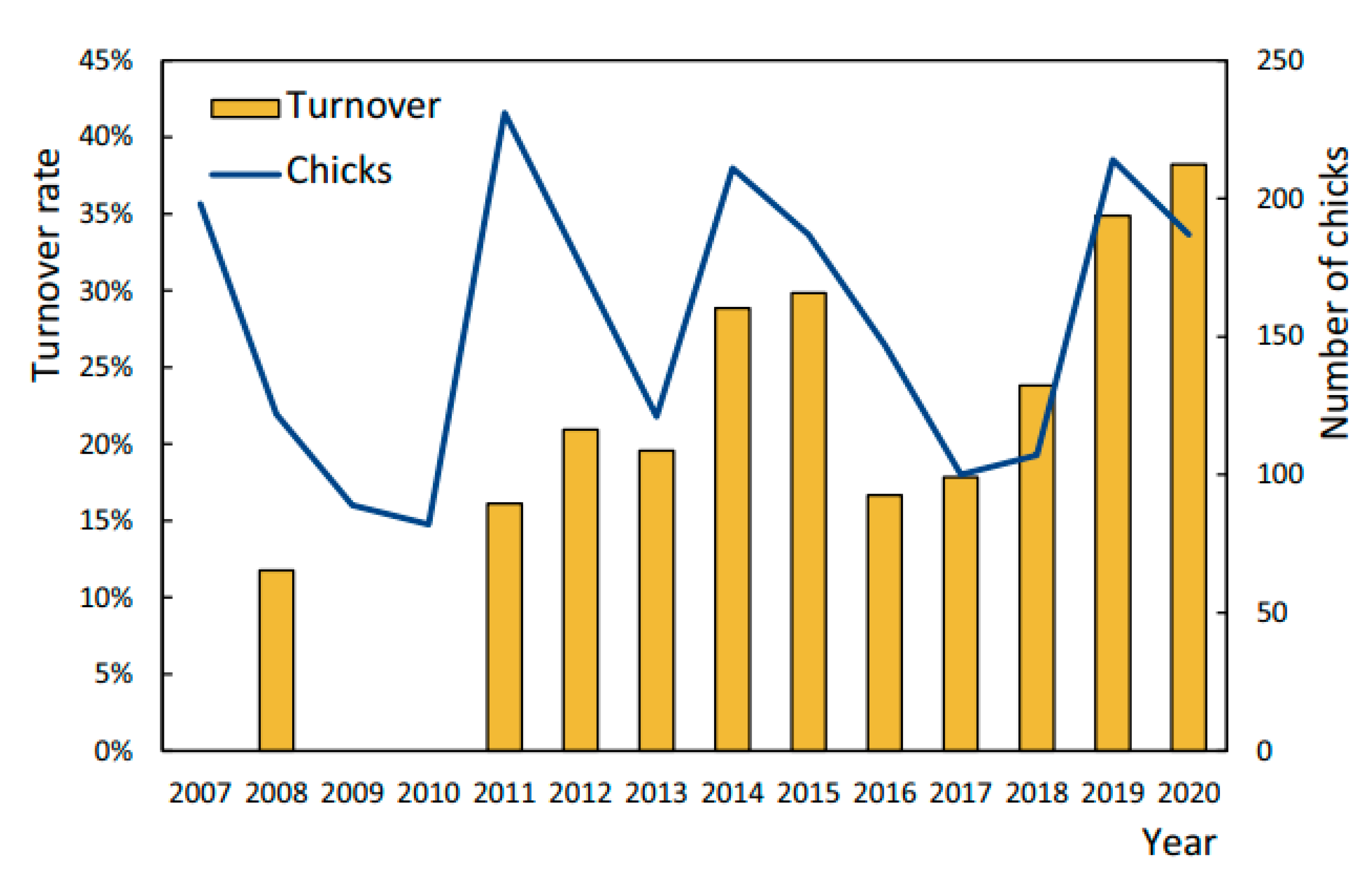
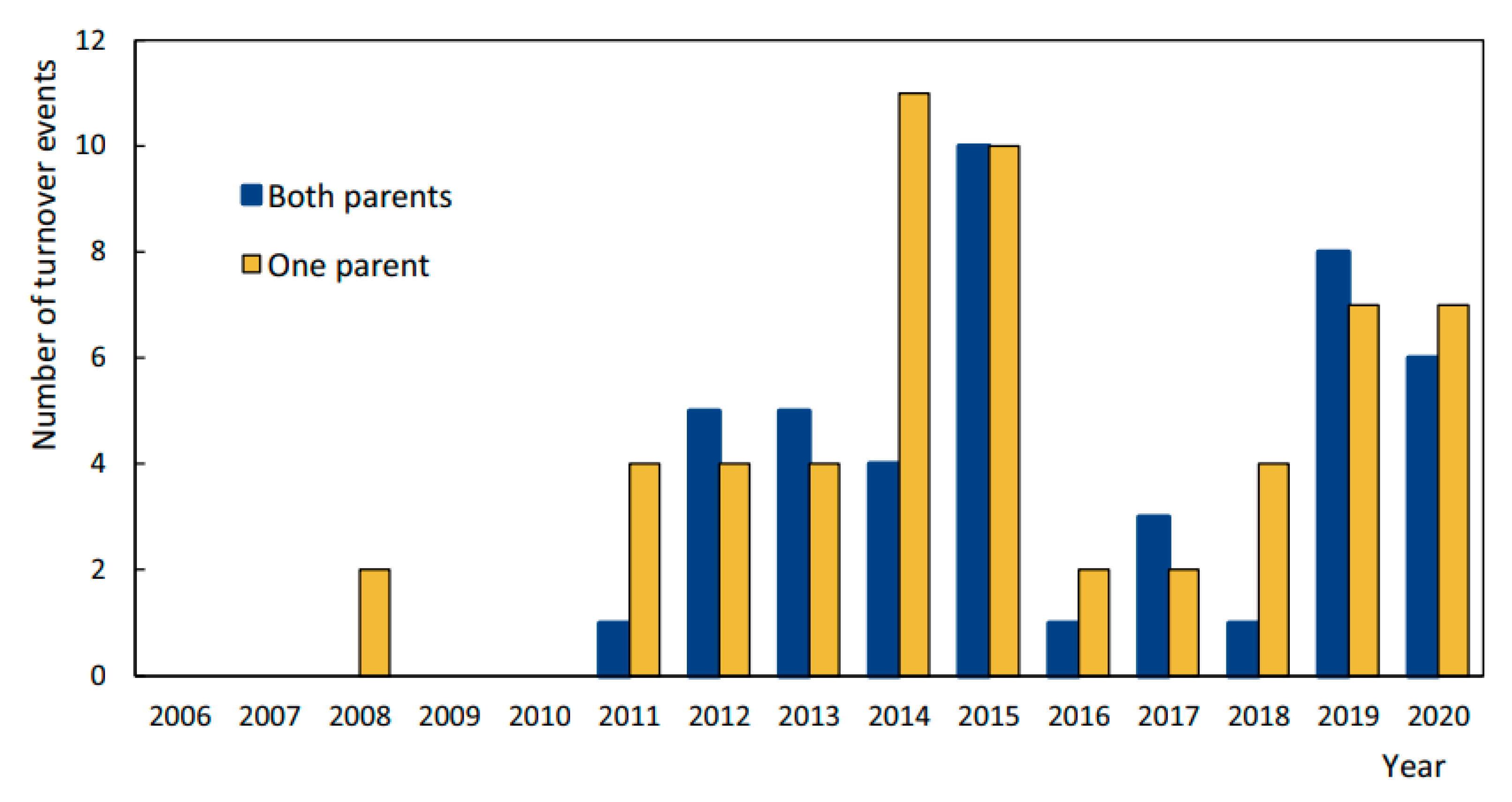
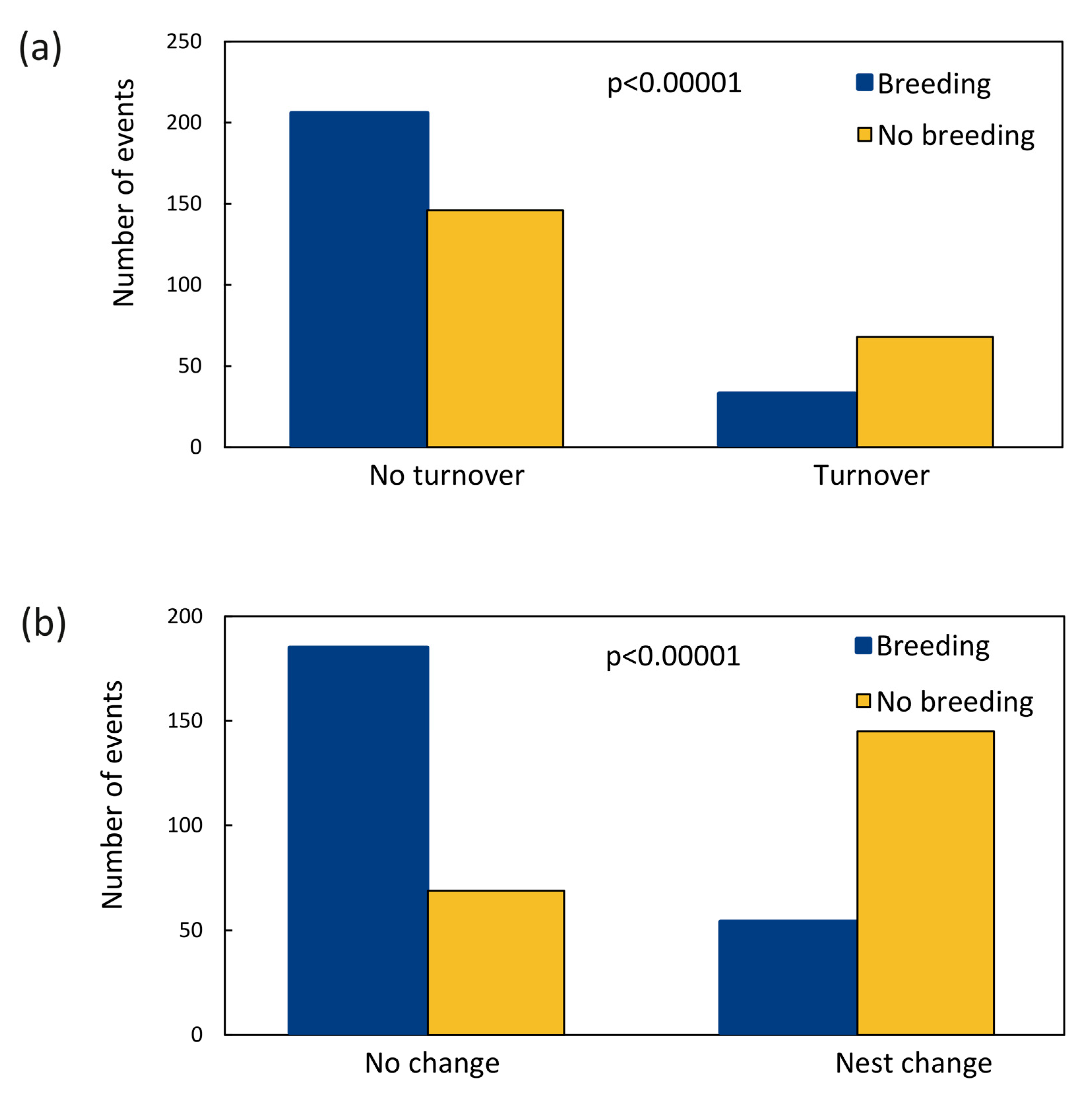
3.3. Turnover
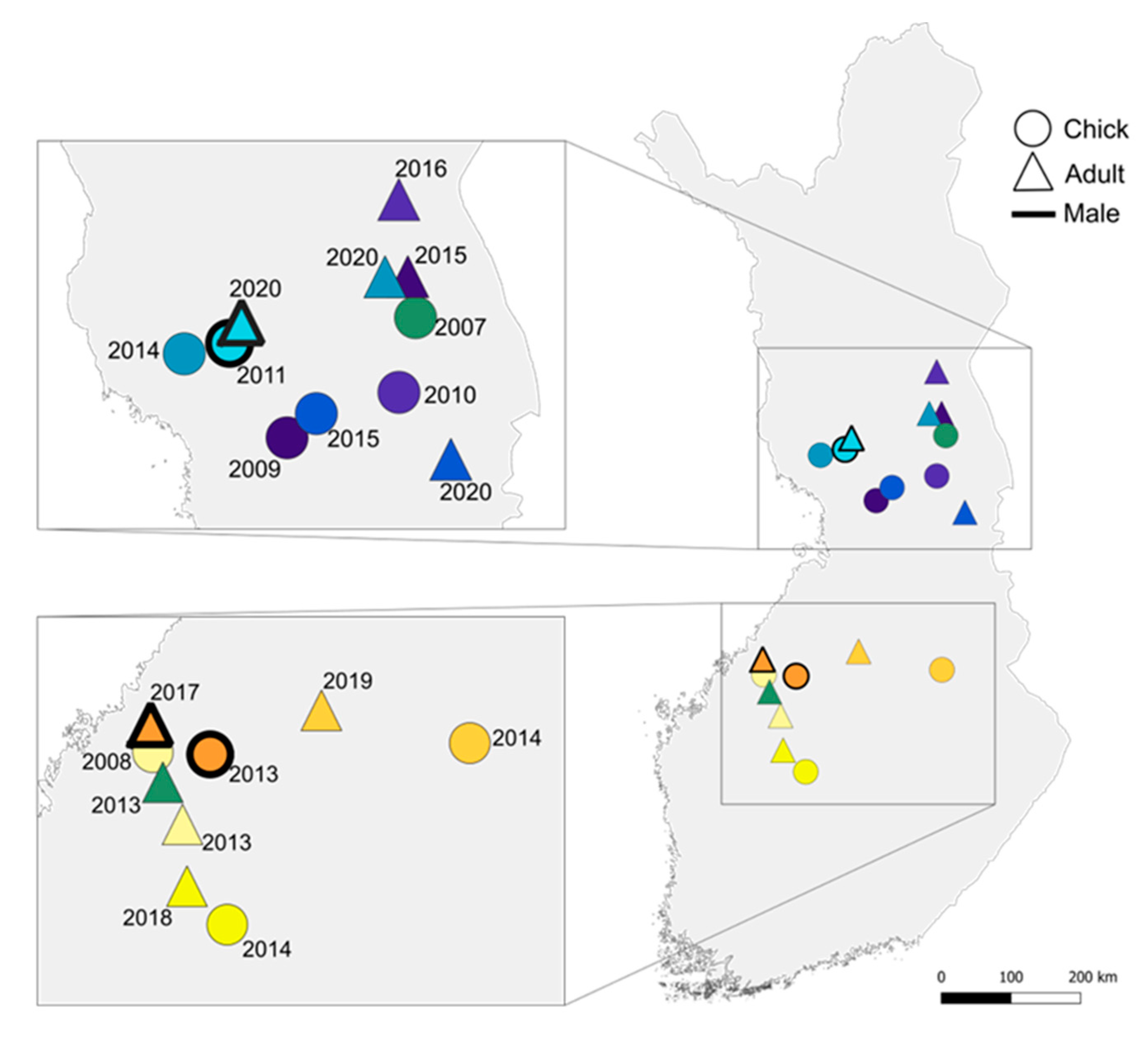
3.4. Natal Dispersal
4. Discussion
4.1. Genetic Diversity and Population Genetic Structure
4.2. Turnover Rate
4.3. Turnover and Nest Change as a Result of Unsuccessful Breeding
4.4. Natal Dispersal
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Greenwood, P.J. Mating systems, philopatry and dispersal in birds and mammals. Anim. Behav. 1980, 28, 1140–1162. [Google Scholar] [CrossRef]
- Linkhart, B.D.; Reynolds, R.T. Return rate, fidelity, and dispersal in a breeding population of Flammulated Owls (Otus flammeolus). Auk 2007, 124, 264–275. [Google Scholar] [CrossRef]
- Cilimburg, A.B.; Lindberg, M.S.; Tewksburry, J.J.; Hejl, S.J. Effects of dispersal on survival probability of adult Yellow Warblers (Dendroica petechia). Auk 2002, 119, 778–789. [Google Scholar] [CrossRef]
- Greenwood, P.J.; Harvey, P.H. The natal and breeding dispersal of birds. Annu. Rev. Ecol. Syst. 1982, 13, 1–21. [Google Scholar] [CrossRef]
- Paradis, E.; Baillie, S.R.; Sutherland, W.J.; Gregory, R.D. Patterns of natal and breeding dispersal in birds. J. Anim. Ecol. 1998, 67, 518–536. [Google Scholar] [CrossRef]
- Briggs, C.W.; Collopy, M.W.; Woodbridge, B. Correlates and fitness consequences of natal dispersal in Swainson’s Hawks. Condor 2012, 114, 764–770. [Google Scholar] [CrossRef]
- Clarke, A.L.; Sæther, B.-E.; Røskaft, E. Sex Biases in Avian Dispersal: A Reappraisal. Oikos 1997, 79, 429–438. [Google Scholar] [CrossRef]
- Pasinelli, G.; Schiegg, K.; Walters, J.R. Genetic and environmental influences on natal dispersal distance in a resident bird species. Am. Nat. 2004, 164, 660–669. [Google Scholar] [CrossRef]
- Murphy, R.K.; Stahlecker, D.W.; Millsap, B.A.; Jacobson, K.V.; Johnson, A.; Smith, C.S.; Tator, K.J.; Kruse, K.L. Natal dispersal distance of golden eagles in the southwestern United States. J. Fish Wildl. Manag. 2019, 10, 213–218. [Google Scholar] [CrossRef]
- Doligez, B.; Pärt, T. Estimating fitness consequences of dispersal: A road to “know-where”? Non-random dispersal and the underestimation of dispersers’ fitness. J. Anim. Ecol. 2008, 77, 1199–1211. [Google Scholar] [CrossRef]
- Scheiner, S.M. Bet-hedging as a complex interaction among developmental instability, environmental heterogeneity, dispersal, and life-history strategy. Ecol. Evol. 2014, 4, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Ye, X.; Zhong, L.; Li, X.; Li, M.; Wang, H.; Yu, X. Effects of breeding success, age and sex on breeding dispersal of a reintroduced population of the Crested Ibis (Nipponia nippon) in Ningshan County, China. Avian Res. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Riding, C.S.; Belthoff, J.R. Breeding Dispersal by Burrowing Owls (Athene cunicularia) in Idaho. J. Raptor Res. 2018, 52, 143–157. [Google Scholar] [CrossRef]
- Kochert, M.N.; Steenhof, K. Frequency of nest use by Golden Eagles in southwestern Idaho. J. Raptor Res. 2012, 46, 239–247. [Google Scholar] [CrossRef]
- Ponnikas, S.; Ollila, T.; Kvist, L. Turnover and post-bottleneck genetic structure in a recovering population of Peregrine Falcons Falco peregrinus. Ibis 2017, 159, 311–323. [Google Scholar] [CrossRef]
- Harts, A.M.F.; Jaatinen, K.; Kokko, H. Evolution of natal and breeding dispersal: When is a territory an asset worth protecting? Behav. Ecol. 2016, 27, 287–294. [Google Scholar] [CrossRef]
- Otterbeck, A.; Lindén, A.; Gunko, R.; Ylinen, E.; Byholm, P. Female breeding dispersal to higher quality habitats in a philopatric top predator. J. Ornithol. 2022, 163, 3–92. [Google Scholar] [CrossRef]
- Enfjäll, K.; Leimar, O. The evolution of dispersal—The importance of information about population density and habitat characteristics. Oikos 2009, 118, 291–299. [Google Scholar] [CrossRef]
- Vasko, V.; Laaksonen, T.; Valkama, J.; Korpimäki, E. Breeding dispersal of Eurasian kestrels Falco tinnunculus under temporally fluctuating food abundance. J. Avian Biol. 2011, 42, 552–563. [Google Scholar] [CrossRef]
- Booms, T.L.; Talbot, S.L.; Sage, G.K.; McCaffery, B.J.; McCracken, K.G.; Schempf, P.F. Nest-site fidelity and dispersal of Gyrfalcons estimated by noninvasive genetic sampling. Condor 2011, 113, 768–778. [Google Scholar] [CrossRef]
- Vili, N.; Szabó, K.; Kovács, S.; Kabai, P.; Kalmár, L.; Horváth, M. High turnover rate revealed by non-invasive genetic analyses in an expanding Eastern Imperial Eagle population. Acta Zool. Acad. Sci. Hung. 2013, 59, 279–295. [Google Scholar]
- Watson, J. The Golden Eagle, 2nd ed.; T & AD Poyser: London, UK, 2010. [Google Scholar]
- Selås, V.; Kleven, O.; Steen, O.F. Female turnover rate differs between two Northern Goshawk Accipiter gentilis nesting areas, as revealed by DNA analysis of moulted feathers. Ibis 2017, 159, 554–566. [Google Scholar] [CrossRef]
- Bush, K.L.; Vinsky, M.D.; Aldridge, C.L.; Paszkowski, C.A. A comparison of sample types varying in invasiveness for use in DNA sex determination in an endangered population of Greater Sage-Grouse (Centrocercus uropihasianus). Conserv. Genet. 2005, 6, 867–870. [Google Scholar] [CrossRef]
- Hyvärinen, E.; Juslén, A.K.; Kemppainen, E.; Uddström, A.; Liukko, U.-M. Suomen Lajien Uhanalaisuus 2019—Punainen Kirja: The 2019 Red List of Finnish Species; Ympäristöministeriö & Suomen Ympäristökeskus: Helsinki, Finland, 2019. [Google Scholar]
- BirdLife International Species Factsheet: Aquila chrysaetos. Available online: http://datazone.birdlife.org/species/factsheet/golden-eagle-aquila-chrysaetos (accessed on 6 March 2023).
- Below, A. Suojelualueverkoston merkitys eräille nisäkäs-ja lintulajeille. In Metsähallituksen Luonnonsuojelujulkaisuja, Sarja A, Nro 121; Metsähallitus: Vantaa, Finland, 2000. (In Finnish) [Google Scholar]
- Ollila, T.; Koskimies, P. Maakotkan ja muuttohaukan suojelutaso Suomessa. Linnut Vuosik. 2008, 2007, 8–17, (In Finnish with English Summary). [Google Scholar]
- Ollila, T. Raportti Maakotkan, Muuttohaukan, Tunturihaukan Sekä Oulun Ja Lapin Läänien Merikotkien Pesinnöistä Vuonna 2021; Metsähallitus: Vantaa, Finland, 2021. (In Finnish) [Google Scholar]
- Ollila, T. The monitoring project of the Golden Eagle Aquila chrysaetos in Finland. In Status of Raptor Populations in Eastern Fennoscandia, Proceedings of the Workshop, Kostomuksha, Karelia, Russia, 8–10 November 2005; Karelian Research Centre of the Russian Academy of Science Finnish-Russian Working Group on Nature Conservation: Petrozavodosk, Russia, 2005; pp. 114–116. [Google Scholar]
- Ollila, T. Raportti Maakotkan, Muuttohaukan, Tunturihaukan Sekä Oulun ja Lapin Läänien Merikotkien Pesinnöistä Vuonna 2019; Metsähallitus: Vantaa, Finland, 2019. (In Finnish) [Google Scholar]
- Ollila, T. Raportti Maakotkan, Muuttohaukan, Tunturihaukan Sekä Oulun ja Lapin Läänien Merikotkien Pesinnöistä Vuonna 2020; Metsähallitus: Vantaa, Finland, 2020. (In Finnish) [Google Scholar]
- Ollila, T. Suomen maakotkat vuonna 2018—Onko pesintämenestys huonontumassa? Linnut Vuosik. 2019, 2018, 104–109, (In Finnish with English Summary). [Google Scholar]
- Honkala, J.; Viikki, C. Europe’s Oldest Golden Eagle is Finnish; Life Science News. University of Helsinki. 2019. Available online: https://www.helsinki.fi/en/news/life-science-news/europes-oldest-golden-eagle-is-finnish (accessed on 14 March 2023).
- Tjernberg, M. Kungsörnens status och ekologi i Sverige 2006 samt tänkbara prognoser för artens urveckling. In Rapport till Rovdjursutredningen—ArtDatabanken; Swedish Agricultural University: Uppsala, Sweden, 2006. (In Swedish) [Google Scholar]
- Brown, L.; Amadon, D. Eagles, Hawks, and Falcons of the World; Country Life Books: London, UK, 1968; Volume 2. [Google Scholar]
- Slater, S.; Keller, K.; Knight, R. Interannual Golden Eagle (Aquila chrysaetos) Nest-Use Patterns in Central Utah: Implications for Long-Term Nest Protection. J. Raptor Res. 2017, 51, 129–135. [Google Scholar] [CrossRef]
- Millsap, B.A.; Grubb, T.G.; Murphy, R.K.; Swem, T.; Watson, J.W. Conservation significance of alternative nests of Golden Eagles. Glob. Ecol. Conserv. 2015, 3, 234–241. [Google Scholar] [CrossRef]
- Bielikova, M.; Ficek, A.; Valkova, D.; Turna, J. Multiplex PCR amplification of 13 microsatellite loci for Aquila chrysaetos in forensic applications. Biologia 2010, 65, 1081–1088. [Google Scholar] [CrossRef]
- Nebel, C.; Gamauf, A.; Haring, E.; Segelbacher, G.; Villers, A.; Zachos, F.E. Mitochondrial DNA analysis reveals Holarctic homogeneity and a distinct Mediterranean lineage in the Golden Eagle (Aquila chrysaetos). Biol. J. Linn. Soc. 2015, 116, 328–340. [Google Scholar] [CrossRef]
- Sonsthagen, S.A.; Coonan, T.J.; Latta, B.C.; Sage, G.K.; Talbot, S.L. Genetic diversity of a newly established population of Golden Eagles on the Channel Islands, California. Biol. Conserv. 2012, 146, 116–122. [Google Scholar] [CrossRef]
- Griffiths, R.; Double, M.C.; Orr, K.; Dawson, R.J. A DNA test to sex most birds. Mol. Ecol. 1998, 7, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Sudo-Yamaji, A.; Abe, M.; Murase, T.; Tsubota, T. Sex identification by alternative polymerase chain reaction methods in Falconiformes. Zool. Sci. 2003, 20, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Honka, J.; Merikanto, K. Microsat_Errcalc. 2020. Available online: https://github.com/karimerikanto/microsat_errcalc (accessed on 25 April 2022).
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program Cervus accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- van Oosterhout, C.; Hutchinson, B.; Wills, D.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Goudet, J.J. FSTAT (Version 1.2): A Computer Program to Calculate F-Statistics. J. Hered. 1995, 86, 485–486. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver. 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Res. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. Heredity 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Li, Y.L.; Liu, J.X. StructureSelector: A web-based software to select and visualize the optimal number of clusters using multiple methods. Mol. Ecol. Resour. 2018, 18, 176–177. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software Structure: A Simulation Study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Puechmaille, S.J. The Program Structure Does Not Reliably Recover the Correct Population Structure When Sampling Is Uneven: Subsampling and New Estimators Alleviate the Problem. Mol. Ecol. Resour. 2016, 16, 608–627. [Google Scholar] [CrossRef] [PubMed]
- Francis, R.M. POPHELPER: An R package and web app to analyse and visualise population structure. Mol. Ecol. Resour. 2016, 17, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Bandelt, H.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Inkscape Project. Inkscape. Available online: https://inkscape.org (accessed on 25 February 2023).
- RStudio Team. RStudio: Integrated Development for R. 2022. Available online: http://www.rstudio.com/ (accessed on 10 May 2022).
- Paradis, E. Pegas: An R package for population genetics with an integrated–modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. Ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Wagner, A.P.; Taper, M.L. ML-Relate: A computer program for maximum likelihood estimation of relatedness and relationship. Mol. Ecol. Notes 2006, 6, 576–579. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2023. Available online: http://qgis.osgeo.org (accessed on 14 March 2023).
- Nebel, C.; Gamauf, A.; Haring, E.; Segelbacher, G.; Väli, Ü.; Villers, A.; Zachos, F.E. New insights into population structure of the European Golden Eagle (Aquila chrysaetos) revealed by microsatellite analysis. Biol. J. Linn. Soc. 2019, 128, 611–631. [Google Scholar] [CrossRef]
- Craig, E.H.; Adams, J.R.; Waits, L.P.; Fuller, M.R.; Whittington, D.M. Nuclear and Mitochondrial DNA Analyses of Golden Eagles (Aquila chrysaetos canadensis) from Three Areas in Western North America; Initial Results and Conservation Implications. PLoS ONE 2016, 11, 164248. [Google Scholar] [CrossRef] [PubMed]
- Bourke, B.P.; Frantz, A.C.; Lavers, C.P.; Davison, A.; Dawson, D.A.; Burke, T.A. Genetic Signatures of Population Change in the British Golden Eagle (Aquila chrysaetos). Conserv. Genet. 2010, 11, 1837–1846. [Google Scholar] [CrossRef]
- Ogden, R.; Heap, E.; McEwing, R.; Tingay, R.; Whitfield, D.P. Population Structure and Dispersal Patterns in Scottish Golden Eagles Aquila chrysaetos Revealed by Molecular Genetic Analysis of Territorial Birds. Ibis 2015, 157, 837–848. [Google Scholar] [CrossRef]
- Sato, Y.; Ogden, R.; Komatsu, M.; Maeda, T.; Inoue-Murayama, M. Integration of Wild and Captive Genetic Management Approaches to Support Conservation of the Endangered Japanese Golden Eagle. Biol. Conserv. 2017, 213, 175–184. [Google Scholar] [CrossRef]
- Judkins, M.E.; van den Bussche, R.A. Holarctic Phylogeography of Golden Eagles (Aquila chrysaetos) and Evaluation of Alternative North American Management Approaches. Biol. J. Linn. Soc. 2018, 123, 471–482. [Google Scholar] [CrossRef]
- Klug, H. Why monogamy? A review of potential ultimate drivers. Front. Ecol. Evol. 2018, 6, 30. [Google Scholar] [CrossRef]
- Harvey, P.H.; Greenwood, P.J.; Campbell, B.; Stenning, M.J. Breeding Dispersal of the Pied Flycatcher (Ficedula hypoleuca). J. Anim. Ecol. 1984, 53, 727–736. [Google Scholar] [CrossRef]
- Divoky, G.J.; Horton, M. Breeding and Natal Dispersal, Nest Habitat Loss and Implications for Marbled Murrelet Populations. In Ecology and Conservation of the Marbled Murrelet; USDA Forest Service General Technical Report PSW-GTR-152; Ralph, C.J., Hunt, G.L., Jr., Raphael, M.G.E., Piatt, J.F., Eds.; USDA Forest Service: Albany, CA, USA, 1995; pp. 83–88. [Google Scholar]
- Suvantola, L. The Golden Eagle Compensation Scheme in Finland as an Example of Incentive Measures: Potential for Conflict Management? In Human-Wildlife Conflicts in Europe: Fisheries and Fish-Eating Vertebrates as a Model Case; Klenke, R.A., Ring, I., Kranz, A., Jepsen, N., Rauschmayer, F., Henle, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 201–214. [Google Scholar]
- Gowaty, P.A.; Karlin, A.A. Multiple maternity and paternity in single broods of apparently monogamous Eastern Bluebirds (Sialia sialis). Behav. Ecol. Sociobiol. 1984, 15, 91–95. [Google Scholar] [CrossRef]
- Blakesley, J.A.; Anderson, D.R.; Noon, B.R. Breeding dispersal in the California Spotted Owl. Condor 2006, 108, 71–81. [Google Scholar] [CrossRef]
- Blomqvist, D.; Andersson, M.; Küpper, C.; Cuthill, I.C.; Kis, J.; Lanctot, R.B.; Brett, K.; Sandercock, B.K.; Székely, T.; Wallander, J.; et al. Genetic similarity between mates and extra-pair parentage in three species of shorebirds. Nature 2002, 419, 613–615. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Liu, F.; Fan, Y.; Jia, B.; Li, J. Extra-pair paternity in two passerine species living in an extreme-arid desert region. Wilson J. Ornithol. 2019, 131, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Krams, I.A.; Mennerat, A.; Krama, T.; Krams, R.; Joers, P.; Elferts, D.; Luoto, E.; Rantala, M.J.; Eliassen, S. Extra-pair paternity explains cooperation in a bird species. Proc. Natl. Acad. Sci. USA 2022, 119, e2112004119. [Google Scholar] [CrossRef] [PubMed]
- Laistal, O. Im Tal der Königsadler: Mit den Zugvögeln zum Polarkreis; Landbuck-Verklag GmbH: Hannover, Germany, 1966. (In German) [Google Scholar]
- Dennis, R. Probable polygyny by Golden Eagle. Br. Birds 1983, 76, 310–311. [Google Scholar]
- Bergo, G. Trios in the Golden Eagle Aquila chrysaetos (L.). Fauna Norv. Ser. C Cindus 1988, 11, 40–44. [Google Scholar]
- Whitfield, D.P.; Fielding, A.H.; McLeod, D.R.A.; Haworth, P.F.; Watson, J. A conservation framework for the Golden Eagle in Scotland: Refining condition targets and assessment of constraint influences. Biol. Conserv. 2006, 130, 465–480. [Google Scholar] [CrossRef]
- Culina, A.; Radersma, R.; Sheldon, B.C. Trading up: The fitness consequences of divorce in monogamous birds. Biol. Rev. 2015, 90, 1015–1034. [Google Scholar] [CrossRef]
- Boeker, E.L.; Ray, T.D. Golden Eagle population studies in the Southwest. Condor 1971, 73, 463–467. [Google Scholar] [CrossRef]
- Millon, A.; Arroyo, B.E.; Bretagnolle, V. Variable but predictable prey availability affects predator breeding success: Natural versus experimental evidence. J. Zool. 2008, 275, 349–358. [Google Scholar] [CrossRef]
- Steenhof, K.; Kochert, M.N.; Mcdonald, T.L. Interactive effects of prey and weather on Golden Eagle reproduction. J. Anim. Ecol. 1997, 66, 350. [Google Scholar] [CrossRef]
- Kaisanlahti-Jokimäki, M.L.; Jokimäki, J.; Huhta, E.; Ukkola, M.; Helle, P.; Ollila, T. Territory occupancy and breeding success of the Golden Eagle (Aquila chrysaetos) around tourist destinations in northern Finland. Ornis Fenn. 2008, 85, 2–12. [Google Scholar]
- Watson, J.; Rae, S.R.; Stillman, R. Nesting density and breeding success of Golden Eagles in relation to food supply in Scotland. J. Anim. Ecol. 1992, 61, 543–550. [Google Scholar] [CrossRef]
- Helle, P.; Ikonen, K.; Kantola, A. Wildlife monitoring in Finland: Online information for game administration, hunters, and the wider public. Can. J. For. Res. 2016, 46, 1491–1496. [Google Scholar] [CrossRef]
- Martínez-Cruz, B.; David, V.A.; Godoy, J.A.; Negro, J.J.; O’Brien, S.J.; Johnson, W.E. Eighteen polymorphic microsatellite markers for the highly endangered Spanish imperial eagle (Aquila adalberti) and related species. Mol. Ecol. Notes 2002, 2, 323–326. [Google Scholar] [CrossRef]
- Nesje, M.; Røed, K.H. Microsatellite DNA markers from the gyrfalcon (Falco rusticolus) and their use in other raptor species. Mol. Ecol. 2000, 9, 1438–1440. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Chicks | Nests | Territories |
|---|---|---|---|
| 2006 | 25 | 24 | 24 |
| 2007 | 109 | 84 | 84 |
| 2008 | 58 | 53 | 53 |
| 2009 | 39 | 35 | 35 |
| 2010 | 30 | 29 | 29 |
| 2011 | 75 | 61 | 61 |
| 2012 | 80 | 70 | 70 |
| 2013 | 81 | 72 | 72 |
| 2014 | 103 | 86 | 86 |
| 2015 | 101 | 91 | 91 |
| 2016 | 36 | 33 | 33 |
| 2017 | 42 | 39 | 39 |
| 2018 | 35 | 30 | 30 |
| 2019 | 65 | 58 | 58 |
| 2020 | 56 | 47 | 47 |
| Total | 935 | 455 | 297 |
| Year | N | HO | HE | A | AR | FIS |
|---|---|---|---|---|---|---|
| 2006 | 9 | 0.586 | 0.584 | 4.27 | 3.95 | 0.032 |
| 2007 | 30 | 0.593 | 0.593 | 5.82 | 4.04 | 0.018 |
| 2008 | 16 | 0.552 | 0.551 | 4.64 | 3.57 | 0.061 |
| 2009 | 13 | 0.618 | 0.618 | 4.73 | 3.94 | 0.009 |
| 2010 | 8 | 0.574 | 0.573 | 3.73 | 3.63 | 0.007 |
| 2011 | 21 | 0.580 | 0.579 | 5.09 | 3.79 | 0.055 |
| 2012 | 28 | 0.581 | 0.580 | 5.73 | 3.80 | 0.034 |
| 2013 | 31 | 0.594 | 0.592 | 5.18 | 3.76 | −0.045 |
| 2014 | 34 | 0.551 | 0.551 | 5.45 | 3.63 | 0.053 |
| 2015 | 30 | 0.603 | 0.603 | 5.45 | 3.92 | 0.015 |
| 2016 | 17 | 0.615 | 0.613 | 5.55 | 4.09 | 0.096 |
| 2017 | 13 | 0.609 | 0.707 | 4.64 | 3.96 | 0.089 |
| 2018 | 12 | 0.569 | 0.569 | 4.64 | 3.88 | 0.008 |
| 2019 | 25 | 0.583 | 0.582 | 5.73 | 3.86 | 0.046 |
| 2020 | 11 | 0.633 | 0.631 | 4.73 | 4.16 | 0.063 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kylmänen, A.; Karabanina, E.; Ollila, T.; Ponnikas, S.; Kvist, L. Turnover and Natal Dispersal in the Finnish Golden Eagle (Aquila chrysaetos) Population. Diversity 2023, 15, 567. https://doi.org/10.3390/d15040567
Kylmänen A, Karabanina E, Ollila T, Ponnikas S, Kvist L. Turnover and Natal Dispersal in the Finnish Golden Eagle (Aquila chrysaetos) Population. Diversity. 2023; 15(4):567. https://doi.org/10.3390/d15040567
Chicago/Turabian StyleKylmänen, Aure, Ekaterina Karabanina, Tuomo Ollila, Suvi Ponnikas, and Laura Kvist. 2023. "Turnover and Natal Dispersal in the Finnish Golden Eagle (Aquila chrysaetos) Population" Diversity 15, no. 4: 567. https://doi.org/10.3390/d15040567