
Use of Microbially Treated Olive Mill Wastewaters as Soil Organic Amendments; Their Short-Term Effects on the Soil Nematode Community
 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Treatments
2.1.1. Initial Preparation of Olive Mill Wastewaters
2.1.2. Selection of Fungal Strains
2.1.3. Chlorella vulgaris
2.1.4. OMW Fermentation by Pleurotus ostreatus AMRL 133 and Chlorella vulgaris in Bioreactors
2.2. Pot Experiment
2.3. Estimation of OMW and Soil Physicochemical Properties
2.4. Nematode Extraction, Identification, and Estimation of Nematode Functional Indices
2.5. Data Analysis
2.5.1. Statistical Analysis
2.5.2. Network Analysis
3. Results
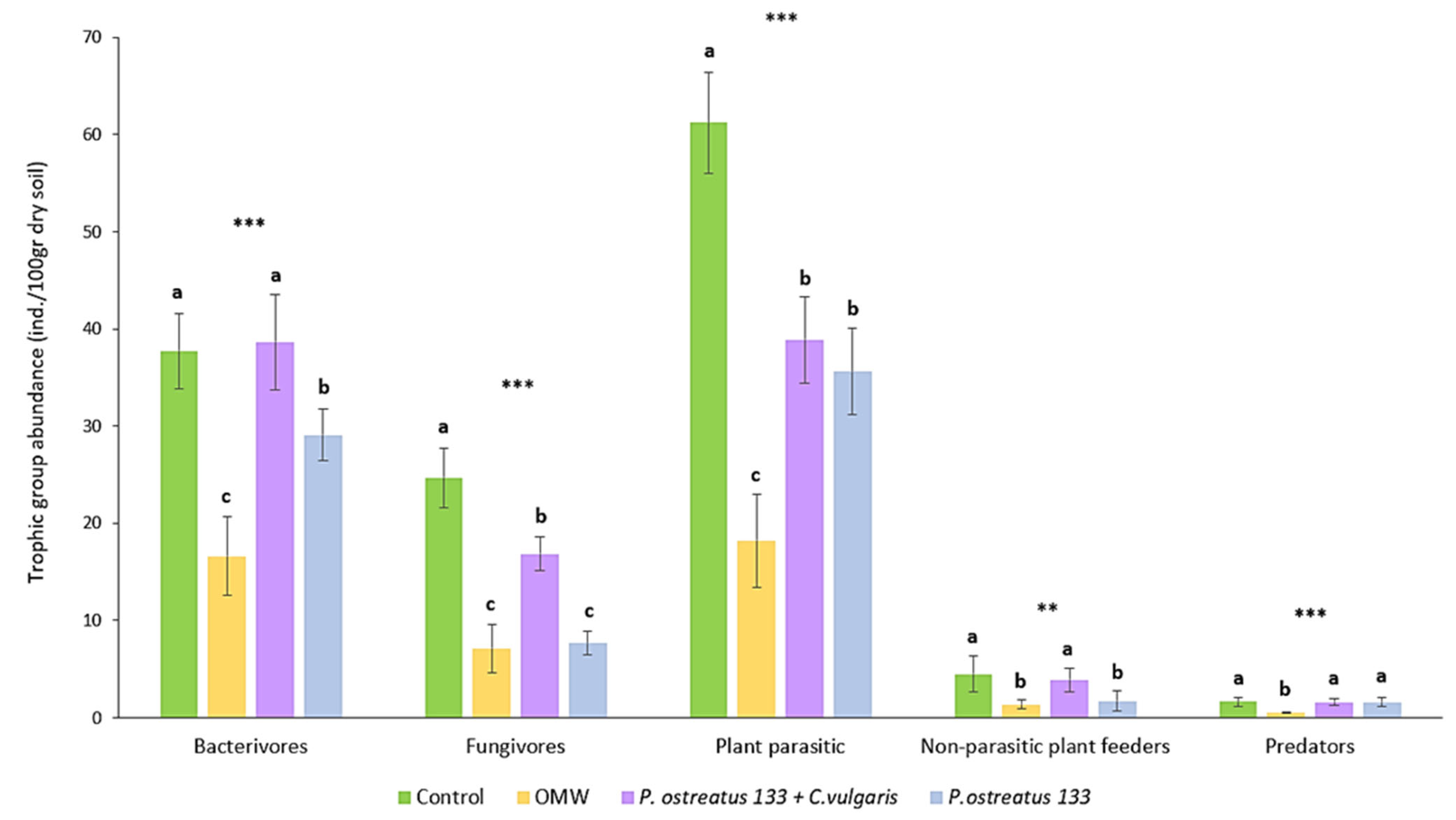
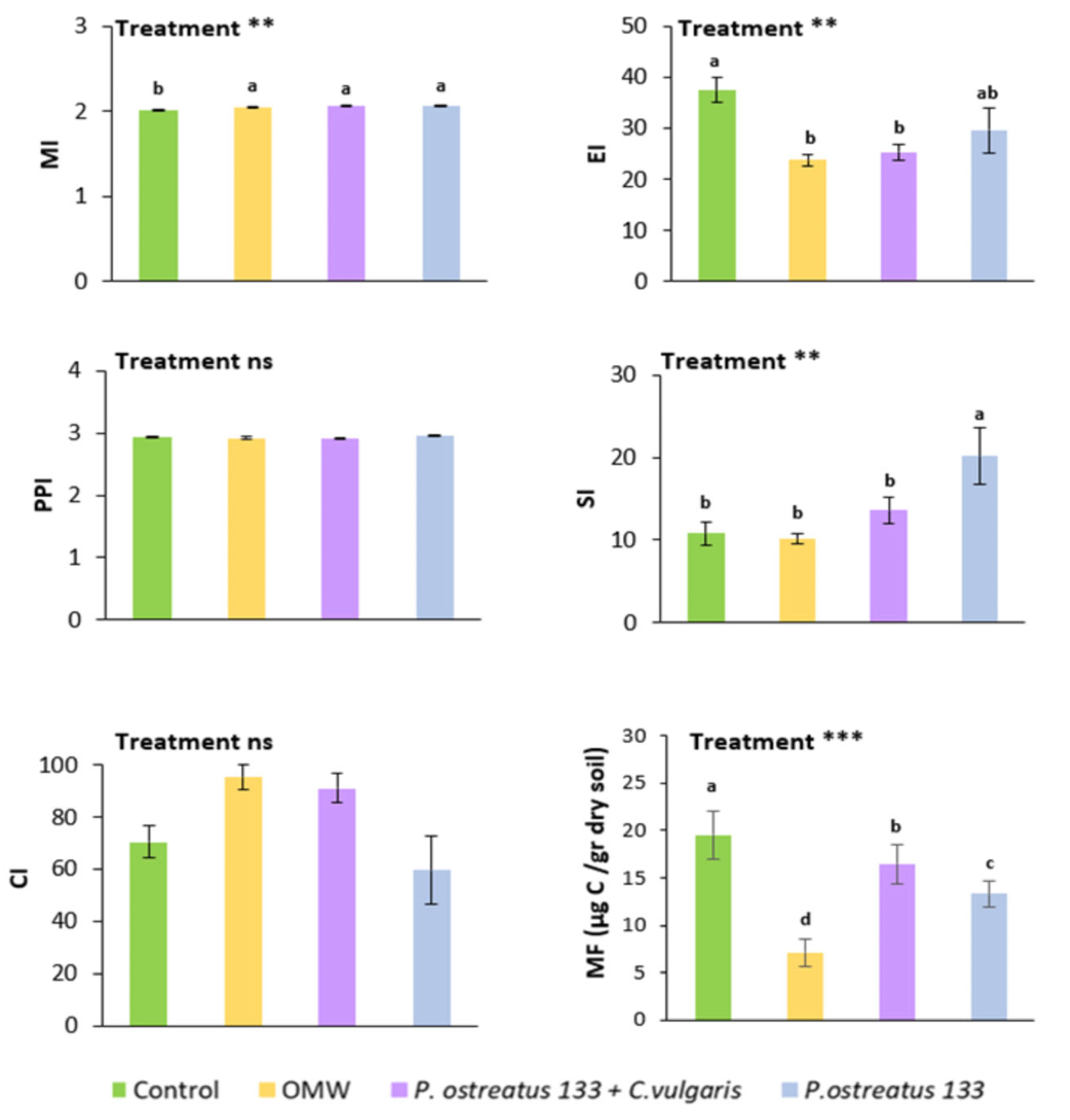
3.1. Trophic and Functional Structure of the Soil Nematode Community
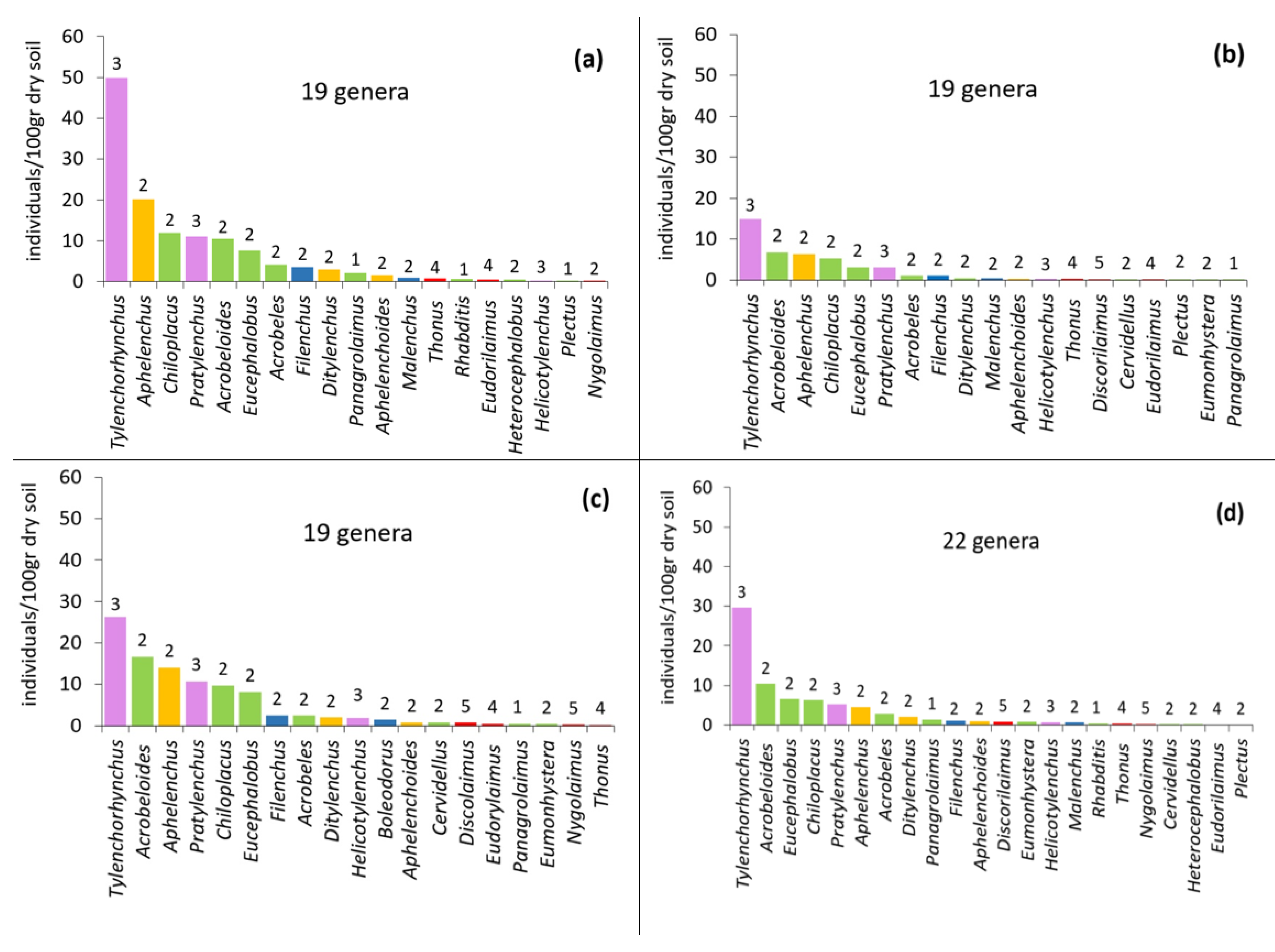
3.2. Community Composition
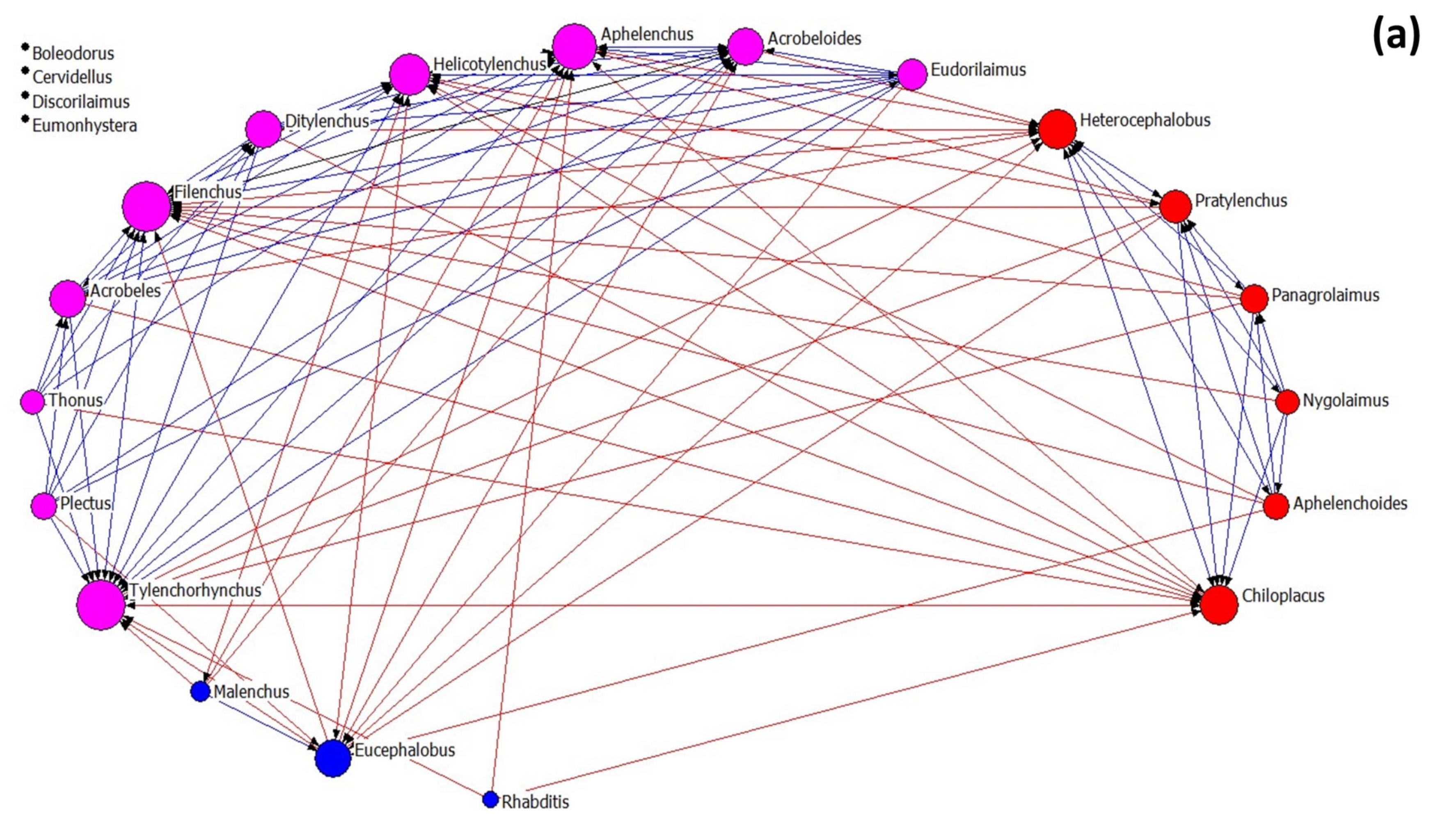
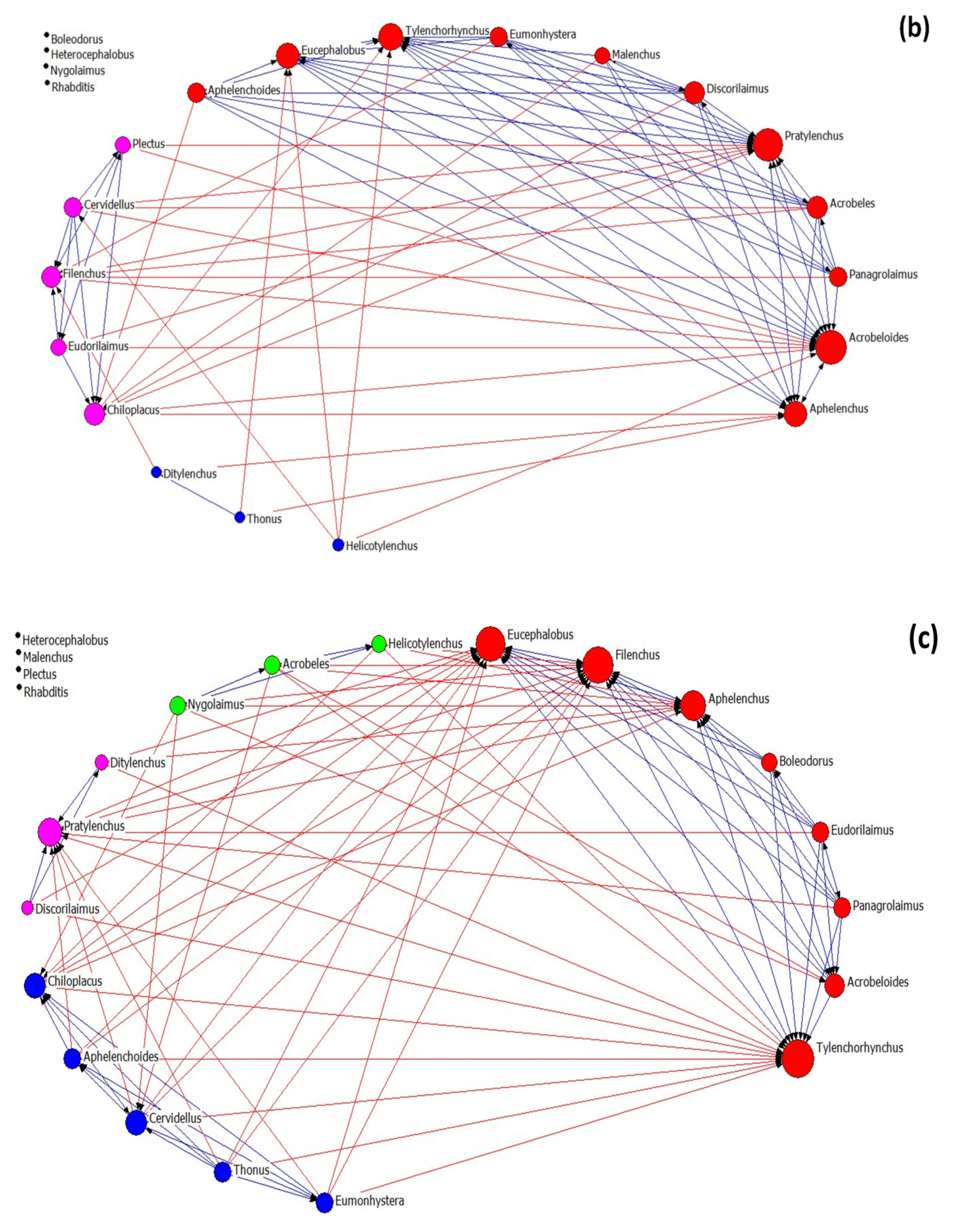
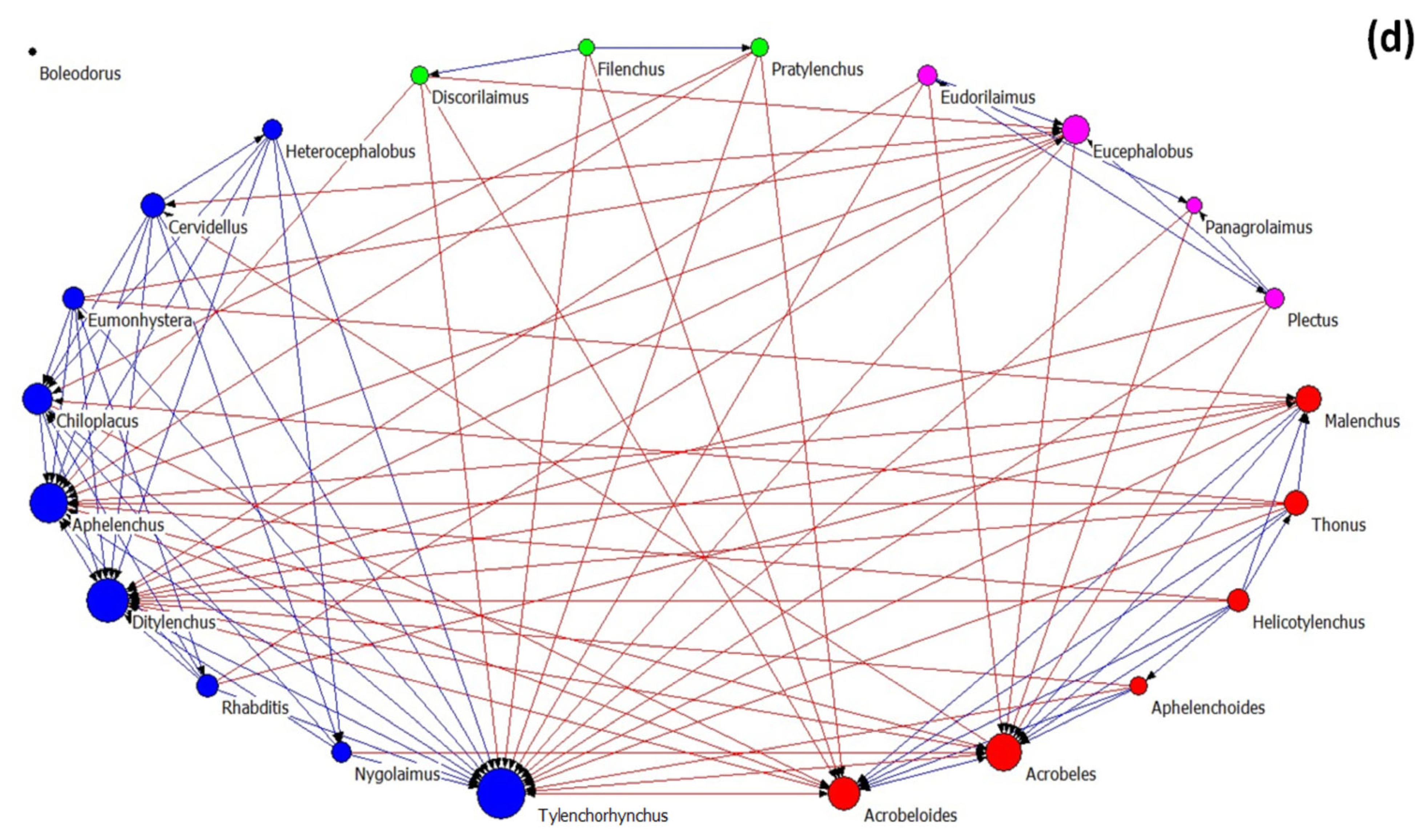
3.3. Nematode Networks
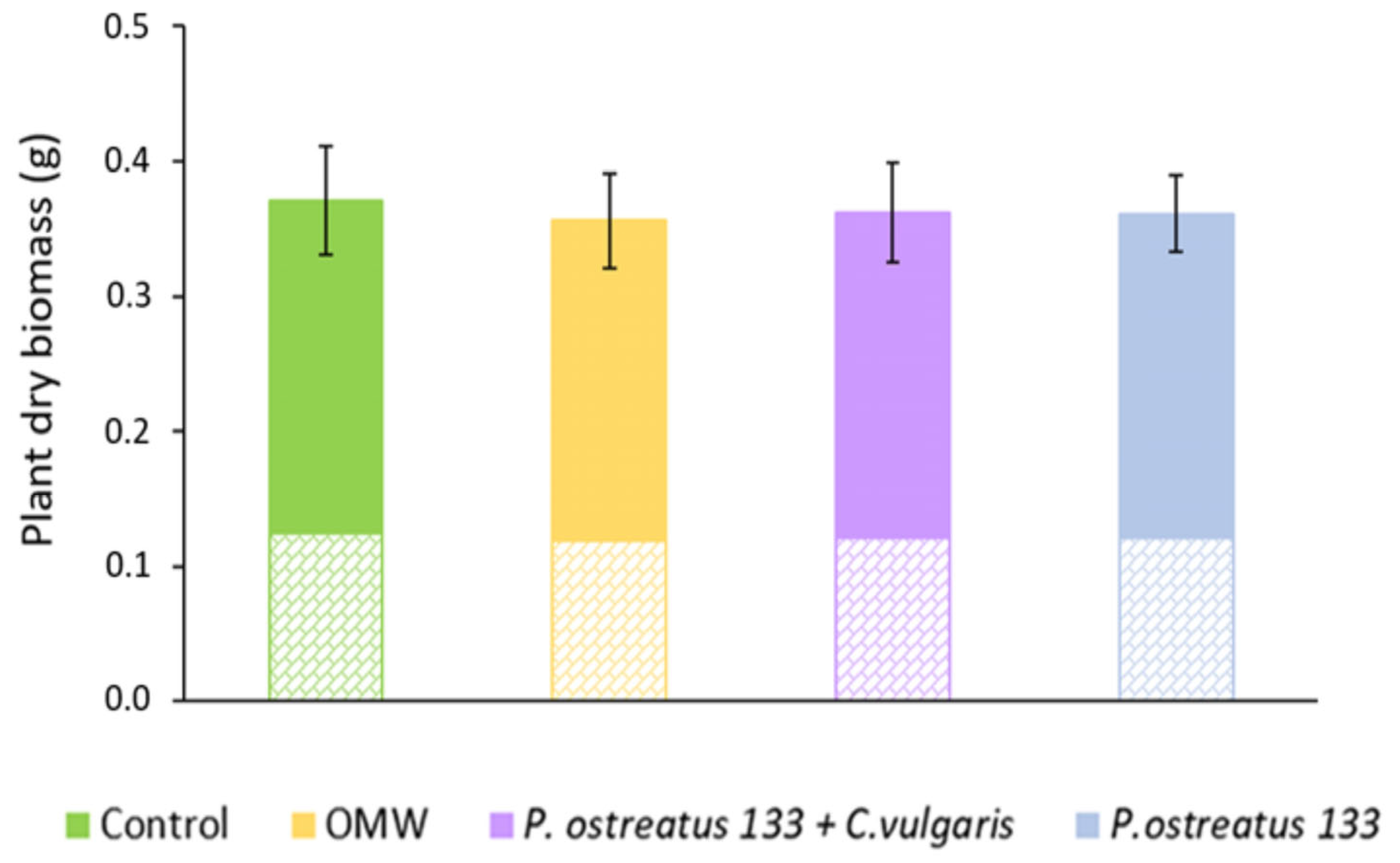
3.4. Growth of Lettuce Plants
4. Discussion
4.1. Nematode Community
4.2. Plants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gholamzadeh, N.; Peyravi, M.; Jahanshahi, M. Study on Olive Mill Wastewater Treatment: Nanotechnology Impact. Nanotechnol. J. Water Environ. Nanotechnol. 2016, 1, 145–161. [Google Scholar] [CrossRef]
- Lesage-Meessen, L.; Navarro, D.; Maunier, S.; Sigoillot, J.C.; Lorquin, J.; Delattre, M.; Simon, J.L.; Asther, M.; Labat, M. Simple Phenolic Content in Olive Oil Residues as a Function of Extraction Systems. Food Chem. 2001, 75, 501–507. [Google Scholar] [CrossRef]
- Azbar, N.; Bayram, A.; Filibeli, A.; Muezzinoglu, A.; Sengul, F.; Ozer, A. A Review of Waste Management Options in Olive Oil Production. Crit. Rev. Environ. Sci. Technol. 2004, 34, 209–247. [Google Scholar] [CrossRef]
- Hamdi, M. Future Prospects and Constraints of Olive Mill Wastewaters Use and Treatment: A Review. Bioprocess Eng. 1993, 8, 209–214. [Google Scholar] [CrossRef]
- Dermeche, S.; Nadour, M.; Larroche, C.; Moulti-Mati, F.; Michaud, P. Olive Mill Wastes: Biochemical Characterizations and Valorization Strategies. Process Biochem. 2013, 48, 1532–1552. [Google Scholar] [CrossRef]
- Mekki, A.; Dhouib, A.; Sayadi, S. Changes in Microbial and Soil Properties Following Amendment with Treated and Untreated Olive Mill Wastewater. Microbiol. Res. 2006, 161, 93–101. [Google Scholar] [CrossRef]
- Omer, A.M. Production of Organic Biofertilizer from Olive Mill Waste Water. Aust. J. Basic Appl. Sci. 2012, 6, 654–663. [Google Scholar]
- Paredes, C.; Cegarra, J.; Roig, A.; Sánchez-Monedero, M.A.; Bernal, M.P. Characterization of Olive Mill Wastewater (Alpechin) and Its Sludge for Agricultural Purposes. Bioresour. Technol. 1999, 67, 111–115. [Google Scholar] [CrossRef]
- Mahmoud, M.; Janssen, M.; Haboub, N.; Nassour, A.; Lennartz, B. Impact of Olive Mill Wastewater Application on Flow and Transport Properties in Soils. Soil Tillage Res. 2010, 107, 36–41. [Google Scholar] [CrossRef]
- Sierra, J.; Martí, E.; Garau, M.A.; Cruañas, R. Effects of the Agronomic Use of Olive Oil Mill Wastewater: Field Experiment. Sci. Total Environ. 2007, 378, 90–94. [Google Scholar] [CrossRef]
- Barbera, A.C.; Maucieri, C.; Cavallaro, V.; Ioppolo, A.; Spagna, G. Effects of Spreading Olive Mill Wastewater on Soil Properties and Crops, a Review. Agric. Water Manag. 2013, 119, 43–53. [Google Scholar] [CrossRef]
- Cayuela, M.L.; Millner, P.D.; Meyer, S.L.F.; Roig, A. Potential of Olive Mill Waste and Compost as Biobased Pesticides against Weeds, Fungi, and Nematodes. Sci. Total Environ. 2008, 399, 11–18. [Google Scholar] [CrossRef]
- Brozzoli, V.; Crognale, S.; Sampedro, I.; Federici, F.; Annibale, A.D.; Petruccioli, M. Bioresource Technology Assessment of Olive-Mill Wastewater as a Growth Medium for Lipase Production by Candida cylindracea in Bench-Top Reactor. Bioresour. Technol. 2009, 100, 3395–3402. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Oliveira, F.; Dantas, D.; Gonçalves, C.; Belo, I. Lipase Production by Aspergillus ibericus Using Olive Mill Wastewater. Bioprocess Biosyst. Eng. 2013, 36, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Díaz, A.I.; Laca, A.; Díaz, M. Fungal Treatment of an Effluent from Sewage Sludge Digestion to Remove Recalcitrant Organic Matter. Biochem. Eng. J. 2021, 172, 108056. [Google Scholar] [CrossRef]
- Blánquez, P.; Caminal, G.; Sarrà, M.; Vicent, M.T.; Gabarrell, X. Olive Oil Mill Waste Waters Decoloration and Detoxification in a Bioreactor by the White Rot Fungus Phanerochaete flavido-alba. Biotechnol. Prog. 2002, 18, 660–662. [Google Scholar] [CrossRef]
- Aggelis, G.; Iconomou, D.; Christou, M.; Bokas, D.; Kotzailias, S.; Christou, G.; Tsagou, V.; Papanikolaou, S. Phenolic Removal in a Model Olive Oil Mill Wastewater Using Pleurotus ostreatus in Bioreactor Cultures and Biological Evaluation of the Process. Water Res. 2003, 37, 3897–3904. [Google Scholar] [CrossRef]
- Ntougias, S.; Baldrian, P.; Ehaliotis, C.; Nerud, F.; Antoniou, T.; Merhautová, V.; Zervakis, G.I. Biodegradation and Detoxification of Olive Mill Wastewater by Selected Strains of the Mushroom Genera Ganoderma and Pleurotus. Chemosphere 2012, 88, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Olivier, G.; Russo, M.E.; Giardina, P.; Marzocchella, A.; Sannia, G.; Salatino, P. Strategies for Dephenolization of Raw Olive Mill Wastewater by Means of Pleurotus ostreatus. J. Ind. Microbiol. Biotechnol. 2012, 39, 719–729. [Google Scholar] [CrossRef]
- Diamantis, I.; Melanouri, E.M.; Dedousi, M.; Panagopoulou, I.; Papanikolaou, S.; Stoforos, N.G.; Diamantopoulou, P. Sustainable and Eco-Friendly Conversions of Olive Mill Wastewater-Based Media by Pleurotus pulmonarius Cultures. Fermentation 2022, 8, 129. [Google Scholar] [CrossRef]
- Oswald, W.; Gotaas, H. Photosynthesis in Sewage Treatment. Trans. Am. Soc. Civ. Eng. 1957, 122, 73–105. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Y.; Chen, P.; Min, M.; Zhou, W.; Martinez, B.; Zhu, J.; Ruan, R. Bioresource Technology Characterization of a Microalga Chlorella sp. Well Adapted to Highly Concentrated Municipal Wastewater for Nutrient Removal and Biodiesel Production. Bioresour. Technol. 2011, 102, 5138–5144. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H.; Venette, R.C.; Lau, S.S. Dynamics of Nematode Communities in Tomatoes Grown in Conventional and Organic Farming Systems, and Their Impact on Soil Fertility. Appl. Soil Ecol. 1996, 3, 161–175. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T. Nematode Indicators of Organic Enrichment. J. Nematol. 2006, 38, 3–12. [Google Scholar] [PubMed]
- Vouyoukalou, E.; Stefanoudaki, E. Nematicidal activity of wastewater from olive oil mills. Nematol. Mediterr. 1998, 26, 157–160. [Google Scholar]
- Rutkowska, A.; Piku, D. Effect of crop rotation and nitrogen fertilization on the quality and quantity of soil organic matter. In Soil Processes and Current Trends in Quality Assessment; IntechOpen: London, UK, 2013; Volume 25, pp. 249–257. [Google Scholar] [CrossRef] [Green Version]
- Valta, K.; Aggeli, E.; Papadaskalopoulou, C.; Panaretou, V.; Sotiropoulos, A.; Malamis, D.; Moustakas, K.; Haralambous, K.J. Adding Value to Olive Oil Production through Waste and Wastewater Treatment and Valorisation: The Case of Greece. Waste Biomass Valorization 2015, 6, 913–925. [Google Scholar] [CrossRef]
- Diamantopoulou, P.; Gardeli, C.; Papanikolaou, S. Impact of Olive Mill Wastewaters on the Physiological Behavior of a Wild-Type New Ganoderma resinaceum Isolate. Environ. Sci. Pollut. Res. 2021, 28, 20570–20585. [Google Scholar] [CrossRef]
- Randrianarison, G.; Ashraf, M.A. Microalgae Plant (Chlorella sp.) for Wastewater Treatment and Energy Production. Ekoloji 2018, 27, 1455–1465. [Google Scholar]
- Lindner, A.V.; Pleissner, D. Removal of Phenolic Compounds from Olive Mill Wastewater by Microalgae Grown under Dark and Light Conditions. Waste Biomass Valorization 2021, 13, 525–534. [Google Scholar] [CrossRef]
- Troelstra, S.R.; Wagenaar, R.; Smant, W.; Peters, B.A.M. Interpretation of Bioassays in the Study of Interactions between Soil Organisms and Plants: Involvement of Nutrient Factors. New Phytol. 2001, 150, 697–706. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Box, J.D. Investigation of the Folin-Ciocalteau Phenol Reagent for the Determination of Polyphenolic Substances in Natural Waters. Water Res. 1983, 17, 511–525. [Google Scholar] [CrossRef]
- Sayadi, S.; Ellouz, R. Roles of Lignin Peroxidase and Manganese Peroxidase from Phanerochaete chrysosporium in the Decolorization of Olive Mill Wastewaters. Appl. Environ. Microbiol. 1995, 61, 1098–1103. [Google Scholar] [CrossRef] [Green Version]
- Kelling, P.S. Some experiments in the low-temperature removal of carbonaceous material from clay. Clay Min. Bull. 1962, 28, 155–158. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. Hydrometer method improved for making particle and size analysis of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- ISO 11465; Soil Quality: Determination of Dry Matter and Water Content on a Mass Basis: Gravimetric Method. International Organization for Standardization: Geneva, Switzerland, 1993.
- S’Jacob, J.J.; van Bezooijen, J. A Manual for Practical Work in Nematology; Department of Nematology, Wageningen Agricultural University: Wageningen, The Netherlands, 1984. [Google Scholar]
- Bongers, T. De Nematoden van Nederland; Koninklijke Nederlandse Natuurhistorische Vereniging: Utrecht, The Netherlands, 1994. [Google Scholar]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding Habits in Soil Nematode Families and Genera—An Outline for Soil Ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Bongers, T. The Maturity Index: An Ecological Measure of Environmental Disturbance Based on Nematode Species Composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T.; De Goede, R.G.M. A Framework for Soil Food Web Diagnostics: Extension of the Nematode Faunal Analysis Concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Bongers, T.; Ferris, H. Nematode Community Structure as a Bioindicator in Environmental Monitoring. Trends Ecol. Evol. 1999, 14, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H. Form and Function: Metabolic Footprints of Nematodes in the Soil Food Web. Eur. J. Soil Biol. 2010, 46, 97–104. [Google Scholar] [CrossRef]
- Sieriebriennikov, B.; Ferris, H.; de Goede, R.G.M. NINJA: An Automated Calculation System for Nematode-Based Biological Monitoring. Eur. J. Soil Biol. 2014, 61, 90–93. [Google Scholar] [CrossRef]
- Butts, C.T. A Relational Event Framework for Social Action. Sociol. Methodol. 2008, 38, 155–200. [Google Scholar] [CrossRef]
- Macarthur, R.; Levins, R. The limiting similarity convergence and divergence of coexisting species. Am. Nat. 1967, 101, 377–385. [Google Scholar] [CrossRef]
- Stamou, G.P.; Argyropoulou, M.D.; Rodriguez-Polo, I.; Boutsis, G.; Kapagianni, P.; Papatheodorou, E.M. A Case Study of Nematode Communities’ Dynamics along Successional Paths in the Reclaimed Landfill. Diversity 2020, 12, 274. [Google Scholar] [CrossRef]
- Borgatti, S.P.; Everett, G.; Freeman, L.C. UCINET 5.0, Version 1.00; Computer Manual; Analytech Technologies: Natick, MA, USA, 1999.
- Huisman, M.; Van Duijn, M.A.J. Software for social network analysis. In Models and Methods in Social Network Analysis; Carrington, P.J., Scott, J., Wasserman, S., Eds.; Cambridge University Press: New York, NY, USA, 2005; pp. 270–316. [Google Scholar]
- O’Malley, A.J.; Marsden, P.V. The Analysis of Social Networks. Health Serv. Outcomes Res. Methodol. 2008, 8, 222–269. [Google Scholar] [CrossRef] [Green Version]
- Rampelotto, P.H.; Barboza, A.D.M.; Pereira, A.B.; Triplett, E.W.; Schaefer, C.E.G.R.; Camargo, F.A.D.O.; Roesch, L.F.W. Distribution and Interaction Patterns of Bacterial Communities in an Ornithogenic Soil of Seymour Island, Antarctica. Microb. Ecol. 2015, 69, 684–694. [Google Scholar] [CrossRef]
- Humphries, M.D.; Gurney, K. Network “Small-World-Ness”: A Quantitative Method for Determining Canonical Network Equivalence. PLoS ONE 2008, 3, e0002051. [Google Scholar] [CrossRef]
- Nakai, S.; Inoue, Y.; Hosomi, M. Algal Growth Inhibition Effects and Inducement Modes by Plant-Producing Phenols. Water Res. 2001, 35, 1855–1859. [Google Scholar] [CrossRef]
- Chitwood, D.J. Phytochemical Based Strategies for Nematode Control. Annu. Rev. Phytopathol. 2002, 40, 221–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Kabana, R.; Morgan-Jones, G.; Chet, I. Biological control of nematodes: Soil amendments and microbial antagonists. Plant Soil 1987, 100, 237–247. [Google Scholar] [CrossRef]
- Akhtar, M.; Malik, A. Roles of organic soil amendments and soil organisms in the biological control of plant-parasitic nematodes: A review. Biores. Technol. 2000, 74, 35–47. [Google Scholar] [CrossRef]
- Kwok, O.C.H.; Plattner, R.; Weisleder, D.; Wicklow, D.T. A Nematicidal Toxin from Pleurotus ostreatus NRRL 3526. J. Chem. Ecol. 1992, 18, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Nordbring-Hertz, B.; Jansson, H.-B.; Friman, E.; Persson, Y.; Dackman, C.; Hard, T.; Poloczek, E.; Feldmann, R. Nematophagous Fungi; Film no. V 1851; Institut für den Wissenschaftlichen Film: Göttingen, Germany, 1995. [Google Scholar]
- Satou, T.; Kaneko, K.; Li, W.; Koike, K. The Toxin Produced by Pleurotus ostreatus Reduces the Head Size of Nematodes. Biol. Pharm. Bull. 2008, 31, 574–576. [Google Scholar] [CrossRef] [Green Version]
- Genier, H.L.A.; Soares, F.E.F.; Queiroz, J.H.; Gouveia, A.S.; Araújo, J.V.; Braga, F.R.; Pinheiro, I.R.; Kasuya, M.C.M. Activity of the fungus Pleurotus ostreatus and of its proteases on Panagrellus sp. larvae. Afr. J. Biotechnol. 2015, 14, 1496–1503. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Zhang, K.; Xu, J.; Dong, J.; Liu, Y. Nematicidal Substances from Fungi. Recent Pat. Biotechnol. 2008, 1, 212–233. [Google Scholar] [CrossRef]
- Samsam-Shariat, H.; Farid, H.; Kavianpour, M. A Study of the Anthelmintic Activity of Aqueous Extract of Pleurotus eryngii on Syphacia obvelata and Hymenolepis nana. J. Sci. Islam. Repub. Iran 1994, 5, 19–22. [Google Scholar]
- Palizi, P.; Goltapeh, E.M.; Pourjam, E.; Safaie, N. Potential of Oyster Mushrooms for the Biocontrol of Sugar Beet Nematode (Heterodera schachtii). J. Plant Prot. Res. 2009, 49, 27–33. [Google Scholar] [CrossRef]
- Khan, A.; Iqbal, M.; Hussain, S.; Pakhtunkhwa, K.; Pakhtunkhwa, K. Organic Control of Phytonematodes with Pleurotus Species. J. Nematol. 2014, 32, 155–161. [Google Scholar]
- Okada, H.; Harada, H.; Kadota, I. Fungal-feeding habits of six nematode isolates in the genus Filenchus. Soil Biol. Biochem. 2005, 37, 1113–1120. [Google Scholar] [CrossRef]
- Monokrousos, N.; Charalampidis, G.; Boutsis, G.; Sousanidou, V.; Papatheodorou, E.M.; Argyropoulou, M.D. Plant-Induced Differentiation of Soil Variables and Nematode Community Structure in a Mediterranean Serpentine Ecosystem. Soil Res. 2014, 52, 593–603. [Google Scholar] [CrossRef]
- Kapagianni, P.D.; Boutsis, G.; Argyropoulou, M.D.; Papatheodorou, E.M.; Stamou, G.P. The network of interactions among soil quality variables and nematodes: Short-term responses to disturbances induced by chemical and organic disinfection. Appl. Soil Ecol. 2010, 44, 67–74. [Google Scholar] [CrossRef]
- Ibrahim, A.; Usman, A.R.A.; Al-Wabel, M.I.; Nadeem, M.; Ok, Y.S.; Al-Omran, A. Effects of Conocarpus Biochar on Hydraulic Properties of Calcareous Sandy Soil: Influence of Particle Size and Application Depth. Arch. Agron. Soil Sci. 2017, 63, 185–197. [Google Scholar] [CrossRef]
- Van Diepeningen, A.D.; De Vos, O.J.; Korthals, G.W.; Van Bruggen, A.H.C. Effects of Organic versus Conventional Management on Chemical and Biological Parameters in Agricultural Soils. Appl. Soil Ecol. 2006, 31, 120–135. [Google Scholar] [CrossRef]
- Prasad, J.S.; Rao, Y.S. Influence of edaphic factors on the build-up of the root lesion nematode, Pratylenchus indicus Das, 1960 in rice. I. Effect of type, texture, porosity, and moisture of the soil. Eur. J. Soil Biol. 1980, 17, 173–179. [Google Scholar]
- Stamou, G.P.; Argyropoulou, M.D.; Tsiafouli, M.A.; Monokrousos, N.; Sgardelis, S.P.; Papatheodorou, E.M. The Study of Secondary Successional Patterns in Soil Using Network Analysis: The Case of Conversion from Conventional to Organic Farming. Pedobiologia 2011, 54, 253–259. [Google Scholar] [CrossRef]
- Stamou, G.P.; Monokrousos, N.; Gwynn-Jones, D.; Whitworth, D.E.; Papatheodorou, E.M. A Polyphasic Approach for Assessing Ecosystem Connectivity Demonstrates That Perturbation Remodels Network Architecture in Soil Microcosms. Microb. Ecol. 2019, 78, 949–960. [Google Scholar] [CrossRef] [Green Version]
- Peng, G.S.; Tan, S.Y.; Wu, J.; Holme, P. Trade-Offs between Robustness and Small-World Effect in Complex Networks. Sci. Rep. 2016, 6, 37317. [Google Scholar] [CrossRef] [Green Version]
- Rusan, M.J.M.; Albalasmeh, A.A.; Zuraiqi, S.; Bashabsheh, M. Evaluation of Phytotoxicity Effect of Olive Mill Wastewater Treated by Different Technologies on Seed Germination of Barley (Hordeum vulgare L.). Environ. Sci. Pollut. Res. 2015, 22, 9127–9135. [Google Scholar] [CrossRef]
- Black, C.A. Soil–Plant Relationships, 2nd ed.; Wiley: New York, NY, USA, 1968; pp. 230–238+372–404. [Google Scholar]
- Kurtz, M.P.; Dag, A.; Zipori, I.; Laor, Y.; Buchmann, C.; Saadi, I.; Medina, S.; Raviv, M.; Zchori-Fein, E.; Schaumann, G.E.; et al. Toward Balancing the Pros and Cons of Spreading Olive Mill Wastewater in Irrigated Olive Orchards. Processes 2021, 9, 780. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Non-Treated OMW | OMW Treated with P. ostreatus | OMW Treated with P. ostreatus + C. vulgaris |
|---|---|---|---|
| pH | 6.00 ± 0.00 | 6.00 ± 0.01 | 6.00 ± 0.02 |
| Electrical conductivity, mS/cm | 12.25 ± 0.03 | 3.74 ± 0.04 | 4.57 ± 0.02 |
| Total sugars (g/L) | 3.405 ± 0.05 | 0.62 ± 0.03 | 0.65 ± 0.01 |
| Total phenol content (TPC), g/L | 1.278 ± 0.04 | 0.31 ± 0.02 | 0.52 ± 0.04 |
| C-organic (%) | 33.26 ± 0.06 | 33.24 ± 0.02 | 32.70 ± 0.02 |
| Total N (%) | 0.82 ± 0.01 | 0.86 ± 0.01 | 0.83 ± 0.03 |
| C/N | 40.56 ± 0.03 | 38.65 ± 0.02 | 39.39 ± 0.01 |
| Soil after Treatments | |||||
|---|---|---|---|---|---|
| Properties | Soil before Treatments | Control | Non-Treated OMW | OMW Treated with P. ostreatus AMRL 133 + C. vulgaris | OMW Treated with P. ostreatus AMRL 133 |
| pH | 7.3 ± 0.01 | 7.680 ± 0.01 | 7.330 ± 0.01 | 7.553 ± 0.01 | 7.420 ±0.01 |
| C-organic (%) | 1.69 ± 0.04 | 1.827 ± 0.02 | 2.066 ± 0.01 | 2.049 ± 0.01 | 2.051 ± 0.03 |
| Total N (%) | 0.14 ± 0. 01 | 0.13 ± 0.00 | 0.14 ± 0.04 | 0.13 ± 0.00 | 0.14 ± 0.03 |
| C/N | 12.07 ± 0.02 | 14.05 | 14.76 | 15.76 | 14.65 |
| Slit (%) | 23 ± 0.02 | ||||
| Clay (%) | 12 ± 0.01 | ||||
| Sand (%) | 65 ± 0.04 | ||||
| Metrics | Control | OMW | P. ostreatus AMRL 133 + C.vulgaris | P. ostreatus AMRL 133 |
|---|---|---|---|---|
| No. of ties | 120 | 91 | 105 | 106 |
| Degree centralization | 0.390 | 0.442 | 0.461 | 0.580 |
| Density | 0.237 | 0.180 | 0.208 | 0.209 |
| Fragmentation | 0.431 | 0.725 | 0.626 | 0.587 |
| Average distance | 1.771 | 1.410 | 1.640 | 1.756 |
| Compactness | 0.386 | 0.224 | 0.278 | 0.294 |
| Small-worldness | 3.175 | 7.130 | 4.893 | 4.094 |
| Modularity | 0.086 | 0.090 | 0.101 | 0.193 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimou, M.D.; Monokrousos, N.; Katapodis, P.; Diamantopoulou, P.A.; Argyropoulou, M.D.; Papatheodorou, E.M. Use of Microbially Treated Olive Mill Wastewaters as Soil Organic Amendments; Their Short-Term Effects on the Soil Nematode Community. Diversity 2023, 15, 497. https://doi.org/10.3390/d15040497
Dimou MD, Monokrousos N, Katapodis P, Diamantopoulou PA, Argyropoulou MD, Papatheodorou EM. Use of Microbially Treated Olive Mill Wastewaters as Soil Organic Amendments; Their Short-Term Effects on the Soil Nematode Community. Diversity. 2023; 15(4):497. https://doi.org/10.3390/d15040497
Chicago/Turabian StyleDimou, Maria D., Nikolaos Monokrousos, Petros Katapodis, Panagiota A. Diamantopoulou, Maria D. Argyropoulou, and Efimia M. Papatheodorou. 2023. "Use of Microbially Treated Olive Mill Wastewaters as Soil Organic Amendments; Their Short-Term Effects on the Soil Nematode Community" Diversity 15, no. 4: 497. https://doi.org/10.3390/d15040497