
Climatic Niche Differentiation between the Invasive Hornet Vespa velutina nigrithorax and Two Native Hornets in Europe, Vespa crabro and Vespa orientalis

Abstract
:1. Introduction
2. Materials and Methods
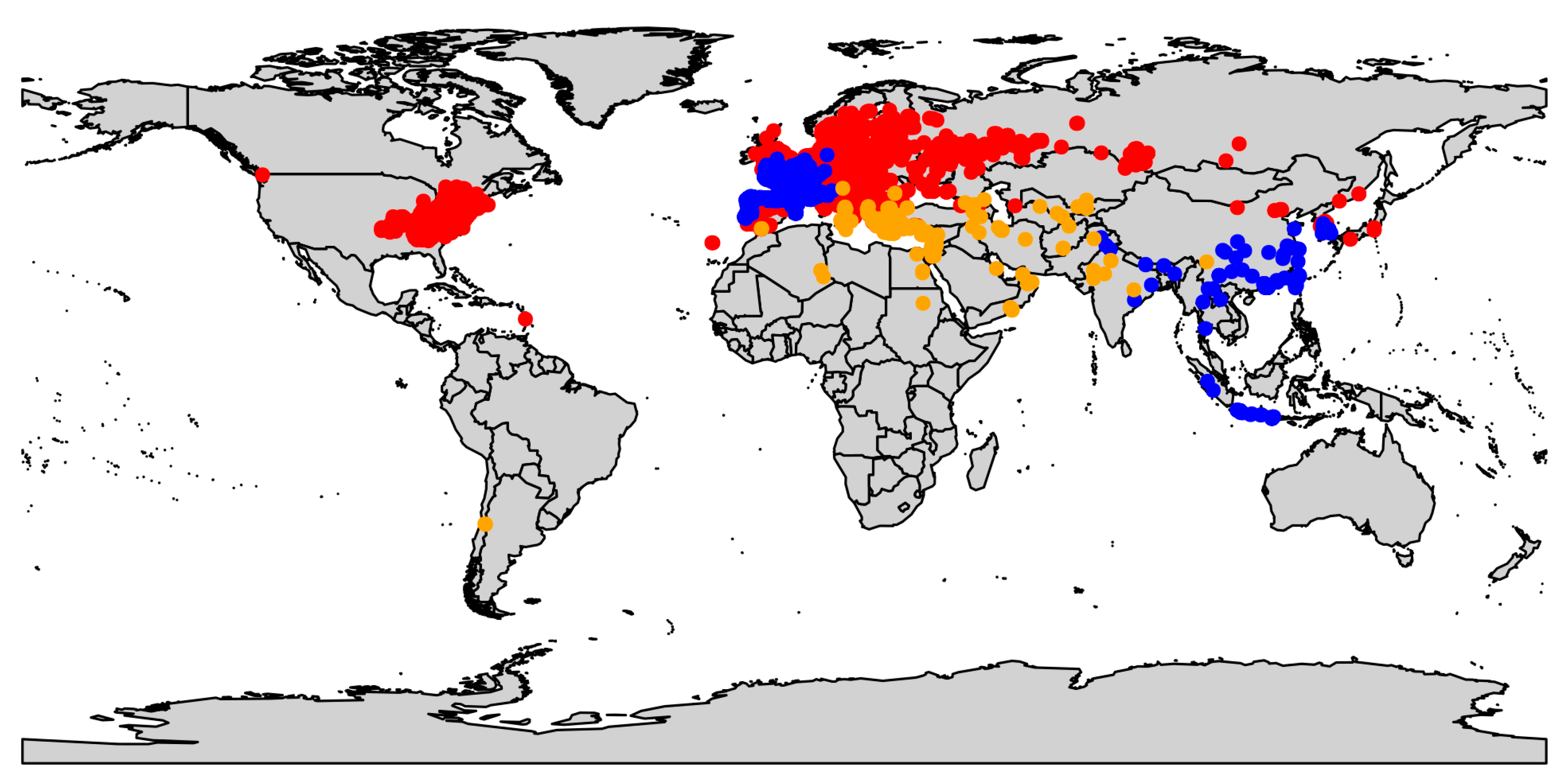
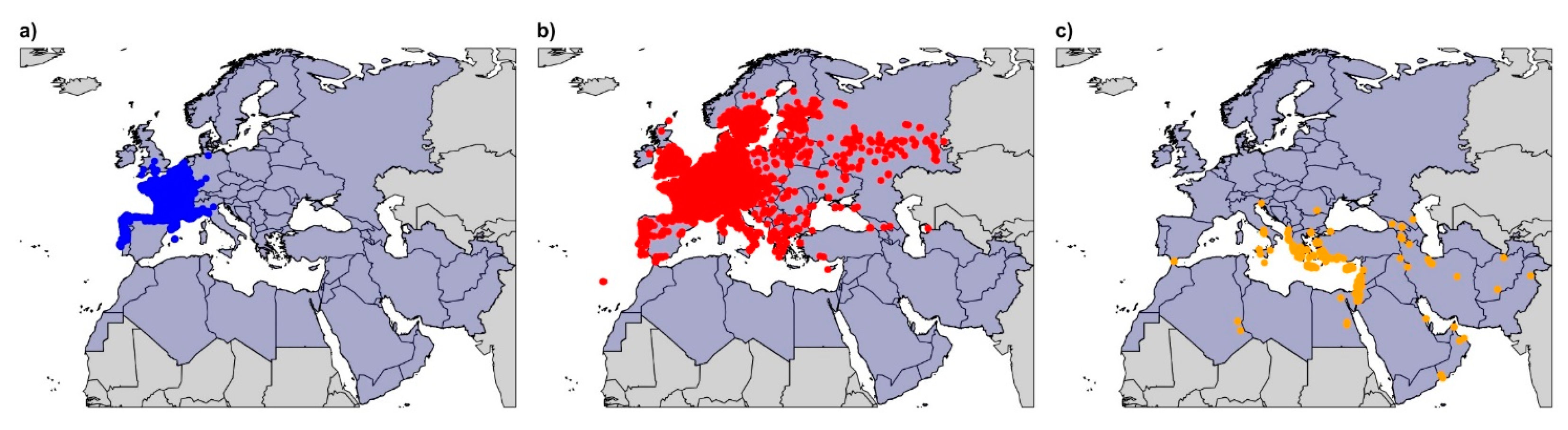
2.1. Data Acquisition and Refinement
2.2. Climatic Niche Analysis for V. v. nigrithorax, V. crabro and V. orientalis at the Study Area Level
2.3. Climatic Niche Analysis for V. v. nigrithorax and V. crabro in the Areas of Co-Occurrence
3. Results
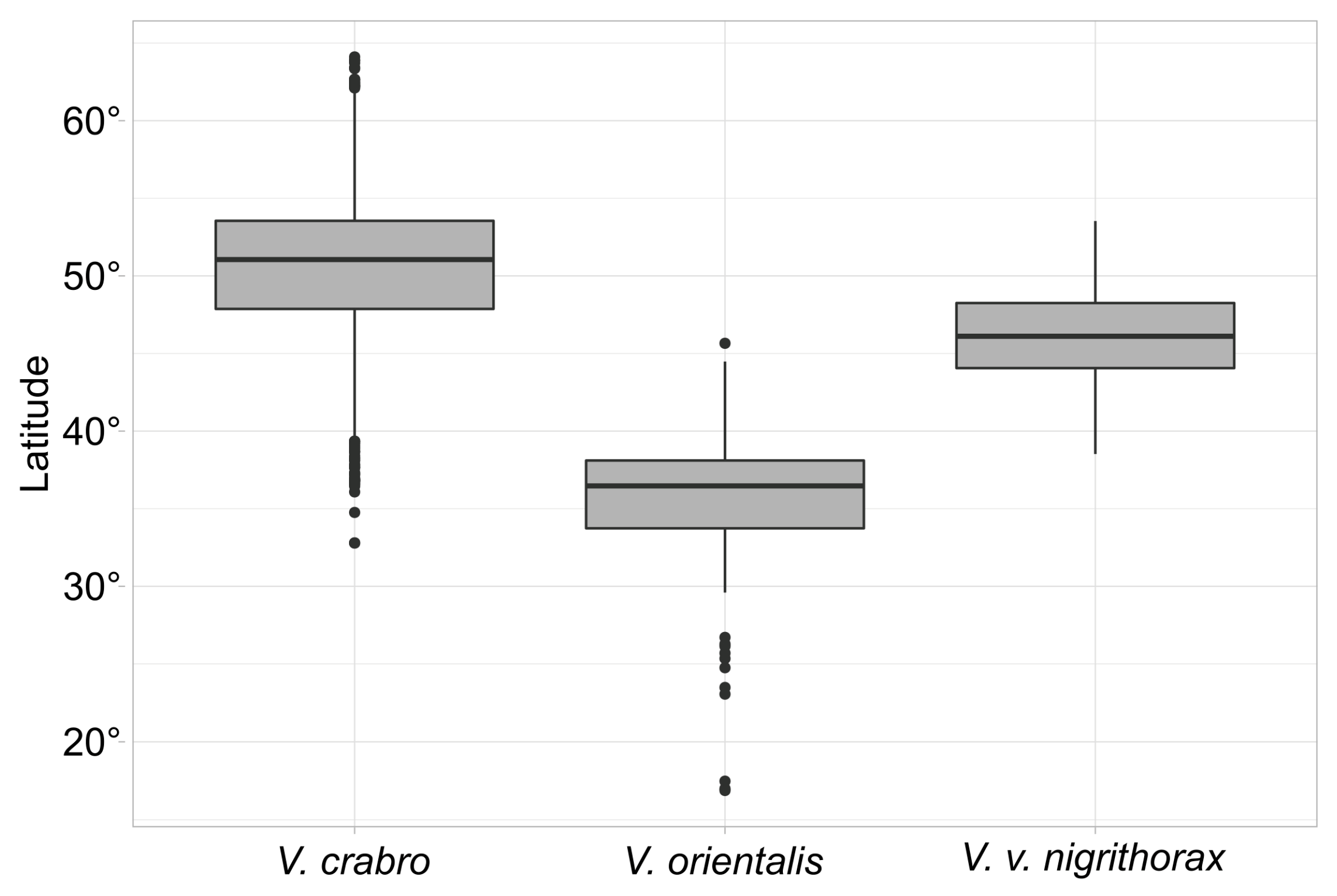
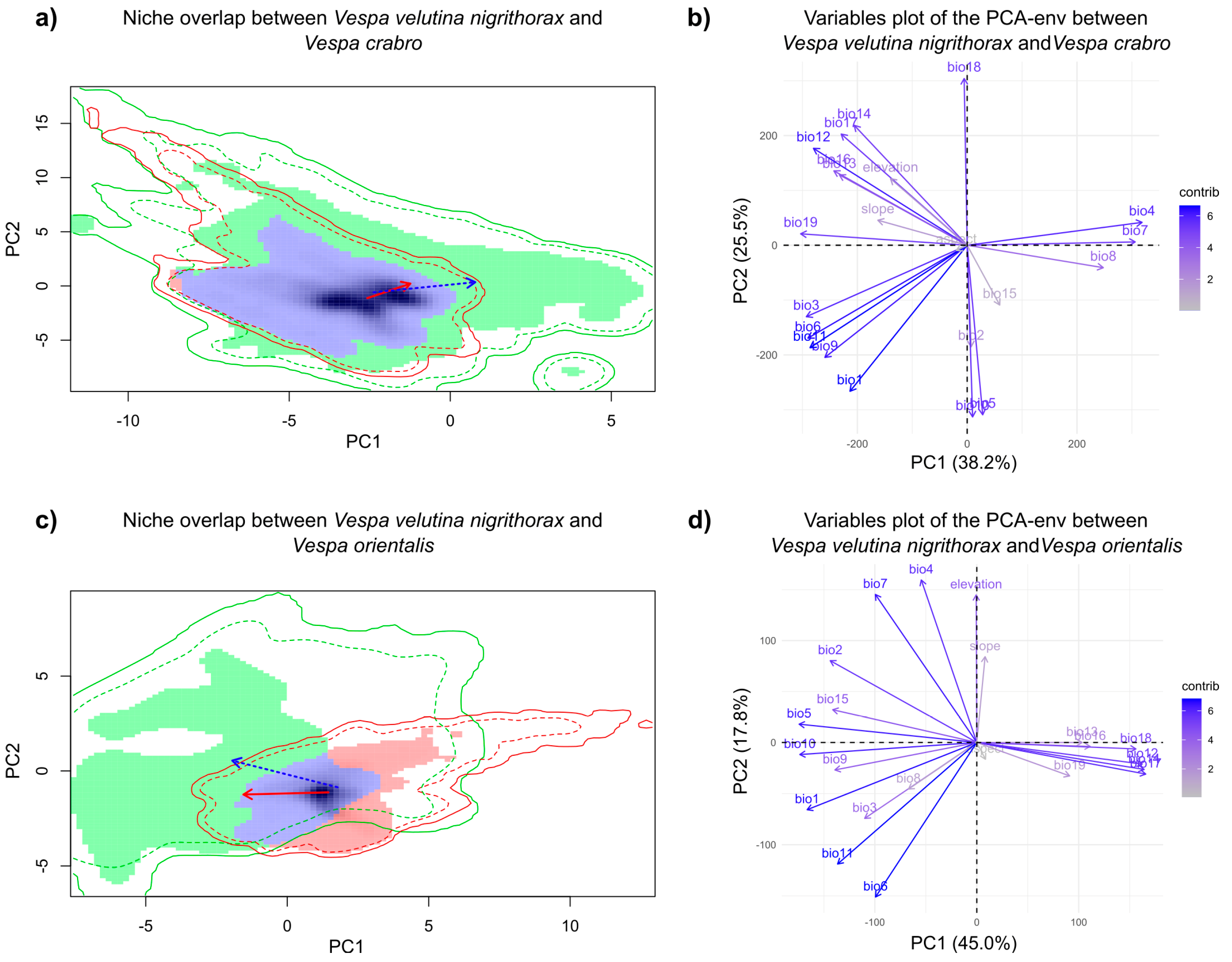
3.1. Climatic Niche Characterisation and Overlapping at the Study Area Level
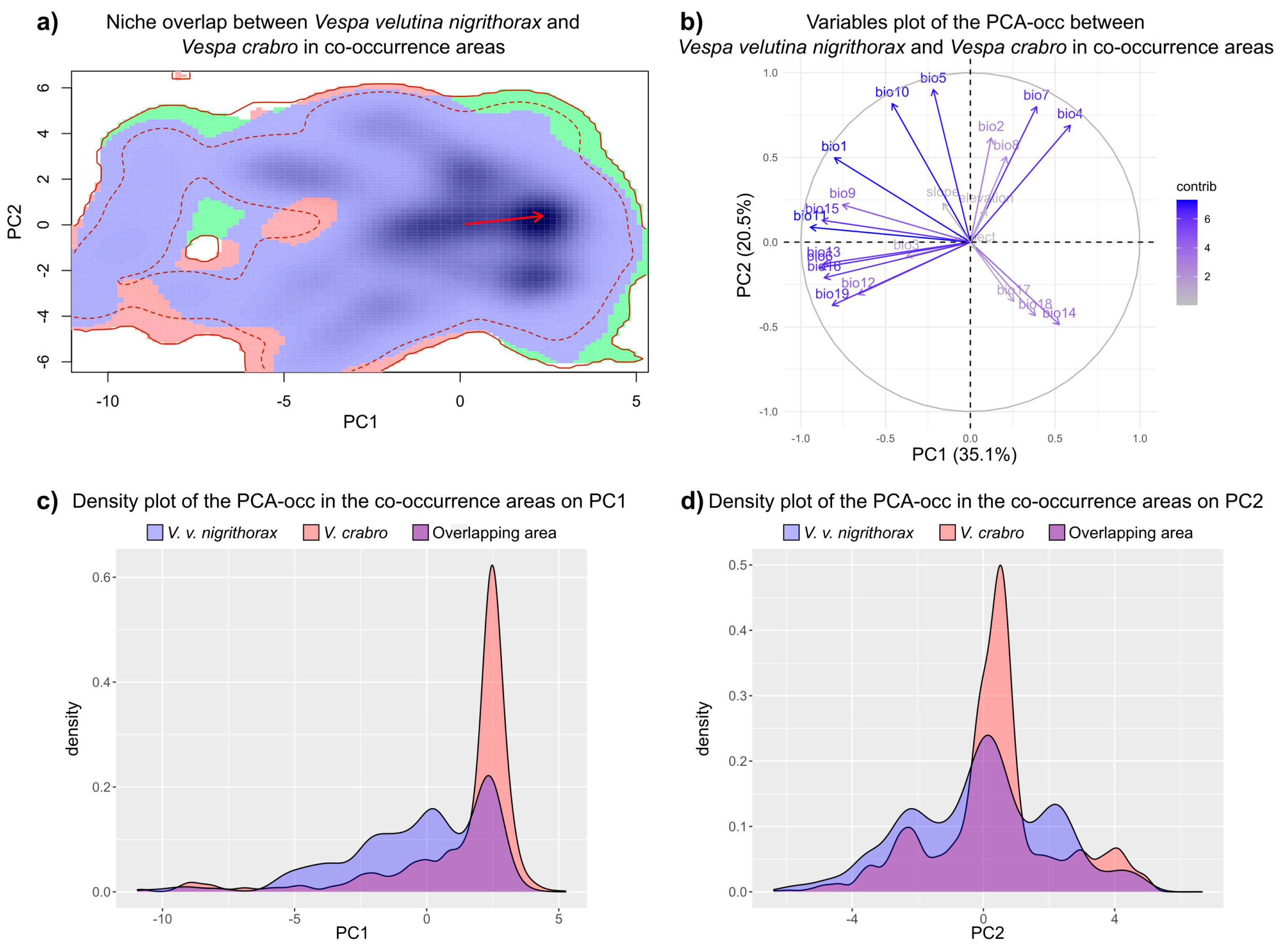
3.2. Climatic Niche Analysis in the Areas of Co-Occurrence of V. v. nigrithorax and V. crabro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Archer, M. Vespine Wasps of the World: Behaviour, Ecology & Taxonomy of the Vespinae; Siri Scientific Press Monograph Series 4; Siri Scientific Press: Rochdale, UK, 2012. [Google Scholar]
- Perrard, A.; Arca, M.; Rome, Q.; Muller, F.; Tan, J.; Bista, S.; Nugroho, H.; Baudoin, R.; Baylac, M.; Silvain, J.F.; et al. Geographic Variation of Melanisation Patterns in a Hornet Species: Genetic Differences, Climatic Pressures or Aposematic Constraints? PLoS ONE 2014, 9, e94162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arca, M.; Mougel, F.; Guillemaud, T.; Dupas, S.; Rome, Q.; Perrard, A.; Muller, F.; Fossoud, A.; Capdevielle-Dulac, C.; Torres-Leguizamon, M.; et al. Reconstructing the Invasion and the Demographic History of the Yellow-Legged Hornet, Vespa velutina, in Europe. Biol. Invasions 2015, 17, 2357–2371. [Google Scholar] [CrossRef]
- Haxaire, J.; Tamisier, J.P.; Bouguet, J.P. Vespa velutina Lepeletier, 1836, Une Redoutable Nouveauté Pour La Faune de France (Hym., Vespidae). Bull. Soc. Entomol. Fr. 2006, 111, 194. [Google Scholar] [CrossRef]
- Choi, M.B.; Martin, S.J.; Lee, J.W. Distribution, Spread, and Impact of the Invasive Hornet Vespa velutina in South Korea. J. Asia Pac. Entomol. 2012, 15, 473–477. [Google Scholar] [CrossRef]
- Ueno, T. Establishment of the Invasive Hornet Vespa velutina (Hymenoptera: Vespidae) in Japan. Int. J. Chem. Environ. Biol. Sci. 2014, 2, 220–222. [Google Scholar]
- Lioy, S.; Bergamino, C.; Porporato, M. The Invasive Hornet Vespa velutina: Distribution, Impacts and Management Options. CABI Rev. 2022, 17, 30. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Essl, F.; Evans, T.; Hulme, P.E.; Jeschke, J.M.; Kühn, I.; Kumschick, S.; Marková, Z.; Mrugała, A.; Nentwig, W.; et al. A Unified Classification of Alien Species Based on the Magnitude of Their Environmental Impacts. PLoS Biol. 2014, 12, e1001850. [Google Scholar] [CrossRef] [Green Version]
- Bacher, S.; Blackburn, T.M.; Essl, F.; Genovesi, P.; Heikkilä, J.; Jeschke, J.M.; Jones, G.; Keller, R.; Kenis, M.; Kueffer, C.; et al. Socio-Economic Impact Classification of Alien Taxa (SEICAT). Methods Ecol. Evol. 2018, 9, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Monceau, K.; Bonnard, O.; Thiéry, D. Vespa velutina: A New Invasive Predator of Honeybees in Europe. J. Pest Sci. 2014, 87, 1–16. [Google Scholar] [CrossRef]
- Laurino, D.; Lioy, S.; Carisio, L.; Manino, A.; Porporato, M. Vespa velutina: An Alien Driver of Honey Bee Colony Losses. Diversity 2020, 12, 5. [Google Scholar] [CrossRef] [Green Version]
- Rome, Q.; Perrard, A.; Muller, F.; Fontaine, C.; Quilès, A.; Zuccon, D.; Villemant, C. Not Just Honeybees: Predatory Habits of Vespa velutina (Hymenoptera: Vespidae) in France. Ann. Soc. Entomol. Fr. 2021, 57, 1–11. [Google Scholar] [CrossRef]
- Requier, F.; Rome, Q.; Chiron, G.; Decante, D.; Marion, S.; Menard, M.; Muller, F.; Villemant, C.; Henry, M. Predation of the Invasive Asian Hornet Affects Foraging Activity and Survival Probability of Honey Bees in Western Europe. J. Pest Sci. 2019, 92, 567–578. [Google Scholar] [CrossRef]
- Rojas-Nossa, S.V.; Calviño-Cancela, M. The Invasive Hornet Vespa velutina Affects Pollination of a Wild Plant through Changes in Abundance and Behaviour of Floral Visitors. Biol. Invasions 2020, 22, 2609–2618. [Google Scholar] [CrossRef]
- Ikegami, M.; Tsujii, K.; Ishizuka, A.; Nakagawa, N.; Kishi, S.; Sakamoto, Y.; Sakamoto, H.; Goka, K. Environments, Spatial Structures, and Species Competitions: Determining the Impact of Yellow-Legged Hornets, Vespa velutina, on Native Wasps and Bees on Tsushima Island, Japan. Biol. Invasions 2020, 22, 3131–3143. [Google Scholar] [CrossRef]
- Kwon, O.; Choi, M.B. Interspecific Hierarchies from Aggressiveness and Body Size among the Invasive Alien Hornet, Vespa velutina nigrithorax, and Five Native Hornets in South Korea. PLoS ONE 2020, 15, e0226934. [Google Scholar] [CrossRef]
- Carisio, L.; Cerri, J.; Lioy, S.; Bianchi, E.; Bertolino, S.; Porporato, M. Impacts of the Invasive Hornet Vespa velutina on Native Wasp Species: A First Effort to Understand Population-Level Effects in an Invaded Area of Europe. J. Insect Conserv. 2022, 26, 663–671. [Google Scholar] [CrossRef]
- Cini, A.; Cappa, F.; Petrocelli, I.; Pepiciello, I.; Bortolotti, L.; Cervo, R. Competition between the Native and the Introduced Hornets Vespa crabro and Vespa velutina: A Comparison of Potentially Relevant Life-History Traits. Ecol. Entomol. 2018, 43, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Monceau, K.; Moreau, J.; Poidatz, J.; Bonnard, O.; Thiéry, D. Behavioral Syndrome in a Native and an Invasive Hymenoptera Species. Insect Sci. 2015, 22, 541–548. [Google Scholar] [CrossRef]
- Monceau, K.; Maher, N.; Bonnard, O.; Thiéry, D. Evaluation of Competition between a Native and an Invasive Hornet Species: Do Seasonal Phenologies Overlap? Bull. Entomol. Res. 2015, 105, 462–469. [Google Scholar] [CrossRef] [Green Version]
- Poidatz, J.; Bressac, C.; Bonnard, O.; Thiéry, D. Comparison of Reproductive Traits of Foundresses in a Native and an Invasive Hornet in Europe. J. Insect Physiol. 2018, 109, 93–99. [Google Scholar] [CrossRef]
- Ricciardi, A.; Hoopes, M.F.; Marchetti, M.P.; Lockwood, J.L. Progress toward Understanding the Ecological Impacts of Nonnative Species. Ecol. Monogr. 2013, 83, 263–282. [Google Scholar] [CrossRef] [Green Version]
- Villemant, C.; Barbet-Massin, M.; Perrard, A.; Muller, F.; Gargominy, O.; Jiguet, F.; Rome, Q. Predicting the Invasion Risk by the Alien Bee-Hawking Yellow-Legged Hornet Vespa velutina nigrithorax across Europe and Other Continents with Niche Models. Biol. Conserv. 2011, 144, 2142–2150. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Salles, J.M.; Courchamp, F. The Economic Cost of Control of the Invasive Yellow-Legged Asian Hornet. NeoBiota 2020, 55, 11–25. [Google Scholar] [CrossRef] [Green Version]
- GBIF Occurrence Download for Vespa velutina. Available online: http://doi.org/10.15468/dl.vhqe7h (accessed on 19 February 2021).
- GBIF Occurrence Download for Vespa crabro. Available online: http://doi.org/10.15468/dl.49jr3y (accessed on 19 February 2021).
- GBIF Occurrence Download for Vespa orientalis. Available online: http://doi.org/10.15468/dl.323rb5 (accessed on 19 February 2021).
- Leza, M.; Miranda, M.Á.; Colomar, V. First Detection of Vespa velutina nigrithorax (Hymenoptera: Vespidae) in the Balearic Islands (Western Mediterranean): A Challenging Study Case. Biol. Invasions 2018, 20, 1643–1649. [Google Scholar] [CrossRef]
- Husemann, M.; Sterr, A.; Mack, S.; Abraham, R. The Northernmost Record of the Asian Hornet Vespa velutina nigrithorax (Hymenoptera, Vespidae). Evol. Syst. 2020, 4, 1–4. [Google Scholar] [CrossRef]
- Poidatz, J.; Monceau, K.; Bonnard, O.; Thiéry, D. Activity Rhythm and Action Range of Workers of the Invasive Hornet Predator of Honeybees Vespa velutina, Measured by Radio Frequency Identification Tags. Ecol. Evol. 2018, 8, 7588–7598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lioy, S.; Laurino, D.; Maggiora, R.; Milanesio, D.; Saccani, M.; Mazzoglio, P.J.; Manino, A.; Porporato, M. Tracking the Invasive Hornet Vespa velutina in Complex Environments by Means of a Harmonic Radar. Sci. Rep. 2021, 11, 12143. [Google Scholar] [CrossRef]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. SpThin: An R Package for Spatial Thinning of Species Occurrence Records for Use in Ecological Niche Models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Silva, D.P.; Vilela, B.; Buzatto, B.A.; Moczek, A.P.; Hortal, J. Contextualized Niche Shifts upon Independent Invasions by the Dung Beetle Onthophagus taurus. Biol. Invasions 2016, 18, 3137–3148. [Google Scholar] [CrossRef] [Green Version]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.J.; Randin, C.; Zimmermann, N.E.; et al. Measuring Ecological Niche Overlap from Occurrence and Spatial Environmental Data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Warren, D.L.; Beaumont, L.J.; Dinnage, R.; Baumgartner, J.B. New Methods for Measuring ENM Breadth and Overlap in Environmental Space. Ecography 2019, 42, 444–446. [Google Scholar] [CrossRef]
- Thioulouse, J.; Dray, S.; Dufour, A.-B.; Siberchicot, A.; Jombart, T.; Pavoine, S. Multivariate Analysis of Ecological Data with Ade4; Springer: New York, NY, USA, 2018; ISBN 978-1-4939-8848-8. [Google Scholar]
- Di Cola, V.D.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; D’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A.; et al. Ecospat: An R Package to Support Spatial Analyses and Modeling of Species Niches and Distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Schoener, T.W. Nonsynchronous Spatial Overlap of Lizards in Patchy Habitats. Ecology 1970, 51, 408–418. [Google Scholar] [CrossRef] [Green Version]
- Verdasca, M.J.; Carvalheiro, L.; Aguirre Gutierrez, J.; Granadeiro, J.P.; Rome, Q.; Puechmaille, S.J.; Rebelo, R.; Rebelo, H. Contrasting Patterns from Two Invasion Fronts Suggest a Niche Shift of an Invasive Predator of Native Bees. PeerJ 2022, 10, e13269. [Google Scholar] [CrossRef] [PubMed]
- Burton, O.J.; Phillips, B.L.; Travis, J.M.J. Trade-Offs and the Evolution of Life-Histories during Range Expansion. Ecol. Lett. 2010, 13, 1210–1220. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.; Volov, M.; Bodner, L.; Bouchebti, S.; Levin, E. Body Size, Metabolic Rate and Diapause in the Oriental Hornet (Vespa orientalis), in Two Extreme Climatic Regions. Ecol. Entomol. 2022, 47, 1022–1031. [Google Scholar] [CrossRef]
- Steen, V.A.; Tingley, M.W.; Paton, P.W.C.; Elphick, C.S. Spatial Thinning and Class Balancing: Key Choices Lead to Variation in the Performance of Species Distribution Models with Citizen Science Data. Methods Ecol. Evol. 2021, 12, 216–226. [Google Scholar] [CrossRef]
- Bates, O.K.; Ollier, S.; Bertelsmeier, C. Smaller Climatic Niche Shifts in Invasive than Non-Invasive Alien Ant Species. Nat. Commun. 2020, 11, 5213. [Google Scholar] [CrossRef] [PubMed]
- Polidori, C.; Sánchez-Fernández, D. Environmental Niche and Global Potential Distribution of the Giant Resin Bee Megachile sculpturalis, a Rapidly Spreading Invasive Pollinator. Glol. Ecol. Conserv. 2020, 24, e01365. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Rome, Q.; Villemant, C.; Courchamp, F. Can Species Distribution Models Really Predict the Expansion of Invasive Species? PLoS ONE 2018, 13, e0193085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandler, M.; See, L.; Copas, K.; Bonde, A.M.Z.; López, B.C.; Danielsen, F.; Legind, J.K.; Masinde, S.; Miller-Rushing, A.J.; Newman, G.; et al. Contribution of Citizen Science towards International Biodiversity Monitoring. Biol. Conserv. 2017, 213, 280–294. [Google Scholar] [CrossRef] [Green Version]
- Pocock, M.J.O.; Roy, H.E.; August, T.; Kuria, A.; Barasa, F.; Bett, J.; Githiru, M.; Kairo, J.; Kimani, J.; Kinuthia, W.; et al. Developing the Global Potential of Citizen Science: Assessing Opportunities That Benefit People, Society and the Environment in East Africa. J. Appl. Ecol. 2019, 56, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Vohland, K.; Göbel, C.; Balázs, B.; Butkevičienė, E.; Daskolia, M.; Duží, B.; Hecker, S.; Manzoni, M.; Schade, S. Citizen Science in Europe. In The Science of Citizen Science; Vohland, K., Land-Zandstra, A., Ceccaroni, L., Lemmens, R., Perelló, J., Ponti, M., Samson, R., Wagenknecht, K., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 35–53. ISBN 978-3-030-58277-7. [Google Scholar]
- Barbet-Massin, M.; Rome, Q.; Muller, F.; Perrard, A.; Villemant, C.; Jiguet, F. Climate Change Increases the Risk of Invasion by the Yellow-Legged Hornet. Biol. Conserv. 2013, 157, 4–10. [Google Scholar] [CrossRef]
- Bressi, N.; Colla, A.; Tomasin, G. Orientali Verso Nord: Insediamento Di Una Popolazione Urbana Di Calabrone Orientale (Vespa orientalis Linnaeus, 1771) a Trieste, NE Italy (Hymenoptera, Vespidae). Atti. Mus. Civ. St. Nat. Trieste 2019, 60, 273–275. [Google Scholar]
- Rome, Q.; Muller, F.J.; Touret-Alby, A.; Darrouzet, E.; Perrard, A.; Villemant, C. Caste Differentiation and Seasonal Changes in Vespa velutina (Hym.: Vespidae) Colonies in Its Introduced Range. J. Appl. Entomol. 2015, 139, 771–782. [Google Scholar] [CrossRef]
- Monceau, K.; Arca, M.; Leprêtre, L.; Mougel, F.; Bonnard, O.; Silvain, J.F.; Maher, N.; Arnold, G.; Thiéry, D. Native Prey and Invasive Predator Patterns of Foraging Activity: The Case of the Yellow-Legged Hornet Predation at European Honeybee Hives. PLoS ONE 2013, 8, e66492. [Google Scholar] [CrossRef] [Green Version]
- Bhatnagar, S.; Sharma, N.; Suman, R.K.; Sankhla, M. Seasonal Abundance of Oriental Hornet in Apis mellifera L. Colonies. J. Entomol. Zool. Stud. 2021, 9, 1150–1153. [Google Scholar]
- Rodríguez-Flores, M.S.; Seijo-Rodríguez, A.; Escuredo, O.; Seijo, M. Spreading of Vespa velutina in Northwestern Spain: Influence of Elevation and Meteorological Factors and Effect of Bait Trapping on Target and Non-Target Living Organisms. J. Pest Sci. 2019, 92, 557–565. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Study Area | Area of Co-Occurrence | ||||
|---|---|---|---|---|---|---|
| Overlapping VVN-VC | Overlapping VVN-VO | Overlapping VVN-VC | ||||
| % | SE | % | SE | % | SE | |
| Bio 01—Annual Mean Temperature (°C) | 0.27 * | 0.01 | 0.10 * | 0.02 | 0.54 * | 0.02 |
| Bio 02—Mean Diurnal Range (°C) | 0.48 * | 0.01 | 0.50 * | 0.03 | 0.52 | 0.02 |
| Bio 03—Isothermality (%) | 0.28 * | 0.01 | 0.37 * | 0.03 | 0.65 * | 0.02 |
| Bio 04—Temperature Seasonality (°C) | 0.36 * | 0.02 | 0.53 * | 0.04 | 0.48 * | 0.02 |
| Bio 05—Max Temperature of Warmest Month (°C) | 0.42 * | 0.01 | 0.08 * | 0.01 | 0.56 | 0.02 |
| Bio 06—Min Temperature of Coldest Month (°C) | 0.36 * | 0.01 | 0.14 * | 0.02 | 0.46 * | 0.01 |
| Bio 07—Temperature Annual Range (°C) | 0.64 * | 0.01 | 0.59 * | 0.04 | 0.53 * | 0.02 |
| Bio 08—Mean Temperature of Wettest Quarter (°C) | 0.49 * | 0.01 | 0.39 * | 0.03 | 0.51 * | 0.01 |
| Bio 09—Mean Temperature of Driest Quarter (°C) | 0.32 * | 0.01 | 0.08 * | 0.01 | 0.50 * | 0.02 |
| Bio 10—Mean Temperature of Warmest Quarter (°C) | 0.40 * | 0.01 | 0.06 * | 0.01 | 0.54 * | 0.02 |
| Bio 11—Mean Temperature of Coldest Quarter (°C) | 0.33 * | 0.01 | 0.13 * | 0.02 | 0.42 * | 0.01 |
| Bio 12—Annual Precipitation (mm) | 0.64 * | 0.01 | 0.43 * | 0.03 | 0.66 * | 0.02 |
| Bio 13—Precipitation of Wettest Month (mm) | 0.72 * | 0.02 | 0.23 * | 0.02 | 0.64 * | 0.02 |
| Bio 14—Precipitation of Driest Month (mm) | 0.54 * | 0.01 | 0.06 * | 0.01 | 0.81 * | 0.02 |
| Bio 15—Precipitation Seasonality (mm) | 0.63 * | 0.02 | 0.06 * | 0.01 | 0.56 * | 0.02 |
| Bio 16—Precipitation of Wettest Quarter (mm) | 0.80 * | 0.02 | 0.25 * | 0.02 | 0.59 * | 0.02 |
| Bio 17—Precipitation of Driest Quarter (mm) | 0.55 * | 0.01 | 0.04 * | 0.01 | 0.66 * | 0.02 |
| Bio 18—Precipitation of Warmest Quarter (mm) | 0.54 * | 0.01 | 0.06 * | 0.01 | 0.65 * | 0.02 |
| Bio 19—Precipitation of Coldest Quarter (mm) | 0.50 * | 0.01 | 0.26 * | 0.02 | 0.55 * | 0.02 |
| Elevation (m a.s.l.) | 0.82 * | 0.02 | 0.57 | 0.04 | 0.72 * | 0.02 |
| Aspect (degrees) | 0.82 * | 0.02 | 0.76 | 0.04 | 0.91 | 0.02 |
| Slope (degrees) | 0.87 * | 0.02 | 0.27 * | 0.02 | 0.82 * | 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lioy, S.; Carisio, L.; Manino, A.; Porporato, M. Climatic Niche Differentiation between the Invasive Hornet Vespa velutina nigrithorax and Two Native Hornets in Europe, Vespa crabro and Vespa orientalis. Diversity 2023, 15, 495. https://doi.org/10.3390/d15040495
Lioy S, Carisio L, Manino A, Porporato M. Climatic Niche Differentiation between the Invasive Hornet Vespa velutina nigrithorax and Two Native Hornets in Europe, Vespa crabro and Vespa orientalis. Diversity. 2023; 15(4):495. https://doi.org/10.3390/d15040495
Chicago/Turabian StyleLioy, Simone, Luca Carisio, Aulo Manino, and Marco Porporato. 2023. "Climatic Niche Differentiation between the Invasive Hornet Vespa velutina nigrithorax and Two Native Hornets in Europe, Vespa crabro and Vespa orientalis" Diversity 15, no. 4: 495. https://doi.org/10.3390/d15040495