

The Factors Influencing Wildlife to Use Existing Bridges and Culverts in Giant Panda National Park
by
 , ,
, ,
Song Lu
1,2,3 ,
,
Ying Yue
2,
Yihong Wang
2,
Dawei Zhang
1,
Biao Yang
4,
Zhen Yu
5,
Honghui Lin
1,* and
Qiang Dai
2,* 1
Ministry of Education Key Laboratory of Bio-Resource and Eco-Environment, College of Life Sciences, Sichuan University, Chengdu 610065, China
2
Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610042, China
3
State Key Laboratory of Hydraulics and Mountain River Engineering, Sichuan University, Chengdu 610065, China
4
Key Laboratory of Southwest China Wildlife Resources Conservation (Ministry of Education), China West Normal University, Nanchong 637002, China
5
Fulinyuan Forestry Development Limited Company, Pingwu County, Mianyang 622550, China
*
Authors to whom correspondence should be addressed.
Diversity 2023, 15(4), 487; https://doi.org/10.3390/d15040487
Submission received: 28 January 2023
/
Revised: 19 March 2023
/
Accepted: 23 March 2023
/
Published: 25 March 2023
(This article belongs to the Special Issue Ecology, Conservation and Restoration of Threatened Animal)
Abstract
:Roads, acting as barriers, hamper wildlife movements and disrupt habitat connectivity. Bridges and culverts are common structures on roads, and some of them can function to allow wildlife passage. This study investigated the effects of traffic, the surrounding landscape, human disturbance, and bridge and culvert structures on the utilization of bridges and culverts as dedicated passages by wildlife, using motion-activated infrared camera traps along a 64 km road in Giant Panda National Park, Sichuan, China. The results show that both species richness and counts of wildlife recorded at the bridge and culvert were significantly lower than those observed at sites distant from roads. No large-sized wildlife was recorded at the bridges and culverts. Human activities and traffic volume significantly and negatively affect medium-sized wildlife utilization of bridges and culverts. We conclude that bridges and culverts serve as wildlife crossings, but their efficacy is weak. This emphasizes the necessity of retrofitting bridges and culverts via mitigation facilities such as noise and light barriers, and vegetation restoration on both sides of the roads in Giant Panda National Park.
1. Introduction
The construction of linear transportation infrastructure (such as roads, railways, etc.), regarded as one of the gravest threats to terrestrial ecosystems [1,2,3], is increasing rapidly worldwide [4,5]. Studies have demonstrated that roads negatively impact wildlife on multiple levels [6,7,8]. Roads cause habitat degradation and loss not only through their physical occupation but also through their noise, light pollution, etc. [9,10]. Many studies have shown that roads also lead to wildlife habitat fragmentation and population isolation [11,12]. Braschler’s research showed that roads influence the dispersal and the population dynamics of small mammals, amphibians, reptiles, and invertebrates [13]. A more visible consequence is roadkill resulting from the collision of wildlife with vehicles, carnivores, small mammals, amphibians, birds, insects, etc., may be killed on the road [14,15]. Wildlife crossing structures can reduce roads’ negative impacts [16,17] by concatenating habitats, mitigating the barrier effects of roads, and reducing roadkill [18,19]. Over the past 50 years, numerous dedicated wildlife overpasses and underpasses have been built worldwide, and the number has steadily increased over time [20]. However, dedicated passages for wildlife are expensive to build and maintain [21]. Given the length of roads in various types of wildlife habitats, the current number of dedicated passages for wildlife is far from sufficient. On the other hand, bridges and culverts (BC), which are ubiquitous in the road system, have been demonstrated to serve as wildlife crossing structures, even though they were not designed initially for wildlife crossings [22,23,24].
Many studies have shown that road infrastructure, such as bridges and culverts, can serve as wildlife passages and decrease roadkill. For instance, in southern Portugal, carnivores were observed using existing bridges and culverts [25]. Ascensão’s research found that drainage culverts resulted in a lower incidence of road mortality among small mammals, indicating that such structures may function as wildlife passages [21].
However, existing bridges and culverts are often unsuitable for wildlife to pass through and are less effective than dedicated wildlife passages in mitigating the road barrier effect [25,26]. The size and surrounding environment of bridges and culverts can have a significant impact on their usage by wildlife [27,28,29]. Clevenger also found that road traffic volume, noise level and road width are the main factors affecting the use of BC by wildlife, and he believed that BC may be critical for maintaining habitat connectivity, making them potentially valuable protection tools, and worthy of analysis [22]. Therefore, understanding the driving factors affecting the efficiency of wildlife utilization of BC provides essential guidance for BC reconstruction to promote wildlife connectivity. Accurately determining the driving factors is essential for ensuring the ecological efficiency of BC while reducing construction costs [17].
Giant Panda National Park (GPNP), which is inhabited by many endangered species and crossed by extensive roads, is located in one of the world’s biodiversity hotspots [30,31]. Studies have revealed that roads in GPNP cause the rapid and permanent alteration and fragmentation of these naturally productive habitats, leading to extinction crises in many species that cannot adapt to such radical changes, including giant pandas (Ailuropoda melanoleuca) [32]. Between 2000 and 2010, there was a significant increase in road disturbances to the giant panda habitat [33], and this trend is expected to continue in the coming years as the region’s economy continues to grow. It has been demonstrated that roads increase accessibility within the giant panda’s habitat, reducing nearby forest coverage and habitat suitability, which in turn leads to a decrease in the presence of giant pandas in the 5 km around roads [32]. Qiu found that controlling the impacts of infrastructure, such as roads, can aid in the restoration of large areas of giant panda habitat [34]. Therefore, mitigating the negative ecological effects caused by roads is critical to conserving native biodiversity in GPNP [34]. To address the problem, managers have planned panda ecological corridors with wildlife crossing structures in some key areas, but the construction has yet to be carried out [35]. Still, widespread existing BC, providing passage for transportation over running waterways such mountain runoff, could mitigate the barrier effects of roads on wildlife habitats before these corridors are established. On the other hand, wildlife crossings in the GPNP, even if they are built, will be limited in number and distribution and should be planned preferentially for the giant panda. Therefore, BC can still be incorporated as a mitigation technology to complement the role of wildlife corridors and protect other species.
To assess the utilization of road BC by wildlife in GPNP and understand the driving factors, we employed long-term monitoring to measure activity patterns for wildlife, using camera traps in the north of GPNP, Sichuan. We further analyzed the quantified factors influencing the utilization of the BC by wildlife from the perspectives of landscape, human activities, traffic volume, and the physical dimension of the BC. The results may provide valuable information for the ecological modification of existing roads and mitigation measures for the construction of new roads in GPNP.
2. Materials and Methods
2.1. Study Area
We conducted our study in Pingwu County, north of GPNP, Sichuan, China (Figure 1). This region is located on the eastern edge of the Tibetan Plateau, where the terrain is mainly sharp mountainous. The vegetation composition is characteristic of primordial forests with a mosaic of spruce (Picea asperata Mast), fir (Abies fabri (Mast.) Craib), nanmu (Phoebe zhennan S.Lee et F.N.Wei), and alpine meadows. The region has rich biodiversity and is inhabited by wildlife such as giant panda, Sichuan snub-nosed monkey (Rhinopithecus roxellana), and takin (Budorcas taxicolor). The road we focused on is a paved, busy tourist route 64 km long. Research has suggested that the road has caused habitat fragmentation for wildlife in this region [31].
2.2. Camera Trap Monitoring
To quantify the utilization of BC by wildlife, 15 camera traps were deployed at bridges and culverts from October 2019 to October 2021. Those bridges and culverts provide passage for mountain runoff. Bridges provide open space (width × height/length) in contrast to culverts. Thirty camera traps were placed on wildlife trails in mountain forests away (4.5 ± 1.7 km) from the roads in the study area to investigate the background richness and abundance of wildlife in the region. The camera traps recorded three images and a 10-s video after triggering. All camera traps operated 24 h a day, and the cameras were visited once every month to replace SD cards and batteries. Successive photo-captures (>30 min apart) of the same species were considered as an independent event [36,37].
2.3. Traffic Volume
The overpassing traffic volume was measured by 10 long-term outdoor audio recorders deployed close to the roads. The recorders continuously worked for at least 11 days, The data were downloaded, and the recorders were recharged every month. Therefore, we derived 11 days of data at each traffic monitoring site per month. The traffic was counted by recognizing vehicle sounds manually using the recorders. We ensured that there was no fork in the road between the camera trap and the nearest recorders. The traffic count for each camera trap was assigned using the nearest recorders.
2.4. Land Cover
For land cover within a 50 m radius of the camera sites, we manually mapped them on high-resolution satellite images (resolution of about 1 m or 0.5 m) from Google Earth map by visual interpretation. Land cover data for 2019, 2020, and 2021, including the distribution of scrub and woodland with a radius of 1 km at camera sites, were obtained by interpreting Landsat 8 30-m resolution satellite images in the study area [38]. After cloud removal, we only selected satellite images from May to October because vegetation was most conspicuous during this period. We chose the Random Forest algorithm as a classifier, given the superior performance of its multi-dimensional features [39]. All training and validation samples were collected based on the manual visual interpretation of high-resolution images from Google Earth. The training and validation samples were derived at a ratio of 7:3, and finally, we had 422 training polygons and 181 validation points. The overall accuracy was 83.71%, and the Kappa coefficient was 0.80. The accuracy of all classifications was above 80%.
2.5. Statistical Analysis
We fitted a generalized linear model (GLM) in a Bayesian framework to compare the differences in wildlife richness between the BC and the sites distant from roads. GLM with Gaussian distribution was used. To correct for differences in camera working days in the models, the working day of each camera was offset to account for the impact of inconsistent trapping efforts [40,41,42].
To compare the difference in the count of independent events for wildlife recorded between the BC and the areas far from roads, we employed generalized linear mixed models (GLMMs) in a Bayesian framework for each species, with an interaction term of species and BC/Control. Since the counts of independent events for wildlife often contain too many zeros and are overdispersed, we applied zero-inflation GLMM with a negative binomial distribution (ZINB) and logit link [43].
To investigate the factors affecting BC’s utilization by wildlife, we employed GLMM in a Bayesian framework across all BC in the region. Only the data from BC sites were used in this analysis. We divided the factors potentially affecting wildlife use of BC into those related to human disturbance and vegetation coverage. We divided the factors potentially affecting wildlife use of BC into those related to human disturbance and vegetation coverage. We note that the distance from water sources and BC material have also been included as variables in other studies. However, in our study, the BC were all designed for drainage, all had water flowing through them, and the materials were all of cement origin, so we believe those variables were unnecessary in our models. The counts of independent events for medium-sized mammals per BC were used as the dependent variable; BC openness, coverage of scrub and woodland within 50 m, coverage of scrub and woodland within 1 km, mean traffic volume, counts of independent human events, and domestic animal events were taken as the independent variables [6,44]. Mammals weighing between 1.0 and 15.0 kg were considered medium-sized according to Gehrt’s and William’s research [45,46].
Zero-inflated formula:
Conditional Formula:
where Cover50 and Cover1 km indicated coverage of scrub and woodland within 50 m and 1 km, respectively. We fitted the GLMM with a zero-inflation Poisson (ZIP) distribution, accounting for the overdispersion in count data. The coverage of scrub and woodland within 1 km was also used as the zero-inflation term. With a threshold of 20 vehicles per hour, traffic volumes were classified into “high” and “low” levels. The independent events of humans and domestic animal identification were used after logarithmic transformation. The coverage of scrub and woodland was transformed using arcsin transformation before analysis. As with ZINB, the working day of each camera was set as an offset, and the ID of the camera site was set as the random effect. At this stage, the independent events of the five most typically recorded medium-sized mammals were pooled for analysis, since the count of events was too low for each respective species when the model included multiple factors.
Variance inflation factor (VIF) was used to test the multicollinearity in models. Any predictor variable that has a VIF more than 5 indicates a potentially severe correlation with at least one other predictor variable and should be removed from the model [47]. No significant collinearity was detected for any of the variables in our models (Table S1).
We first built a global model with full variables, and then removed nonsignificant variables [48]. All model fits were assessed using widely applicable information criteria (WAIC) and leave one out cross-validation (LOOIC). We estimated fixed effects (means and 95% CI) from the posterior distributions for each predictor. If its 95% confidence interval did not contain zero, we considered the effect of the variable to be significantly relevant [49].
For all models in the Bayesian framework, we simultaneously used four Markov chain Monte Carlo (MCMC) sampling to obtain draws from posterior distributions, each with 3000 iterations and 1000 warm-up iterations and achieved convergence on all four chains (all coefficient estimates: rhat = 1.00). GLM and GLMM were built using “brms” packages [50].
To further test whether the daily wildlife activity pattern at the BC was influenced by human activity and traffic, we compared the activity patterns of medium-sized mammals at the BC with those of the same species at sites far from roads. In the R package “overlap”, the estimator suggested in the package (Dhat4 is used when the small sample number is greater than 50) was used to calculate the overlap coefficient, which is used to evaluate the transformation of animal activity patterns. The overlap coefficient ranges from 0 (no overlap) to 1 (complete overlap). To account for the successive changes of the sun’s position throughout the year, we used the “sunTime” function of “Overlap” to remaps clock times to sun times (times to sunrise or sunset) [51]. Meanwhile, bootstrap simulation was undertaken 10,000 times with a 95% confidence interval. When the upper bound of the overlap coefficient within the 95% confidence interval was less than 0.90, we inferred that there were significant alterations in the activity patterns of wild animals between the two sites [52].
3. Results
3.1. Richness and Abundance
All 45 cameras have a total sample effort of 10,961 camera-days at the BC sites and 5020 camera-days at sites far from roads. We captured 137,496 images, including 4159 images and 1391 independent events of wildlife. Cameras at the BC sites recorded only small and medium-size wildlife, while large mammals such as Tufted Deer (Elaphodus cephalophus) and Reeves’s Muntjac (Muntiacus reevesi) were only recorded at the at sites far from roads. (Tables S2 and S3). Five species of Galliformes were recorded at the sites far from roads, while only one species of them was recorded at BC sites. Masked Palm Civet, Yellow Throated Marten, Leopard Cat (Prionailurus bengalensis), Hog Badger (Arctonyx collaris), and Malayan Porcupine were the top five species recorded at BC sites (counts of image > 72%). Meanwhile, we captured 3683 images of human activities and 47,715 images of domestic animals at the BC sites, resulting in 512 and 3310 independent events, respectively.
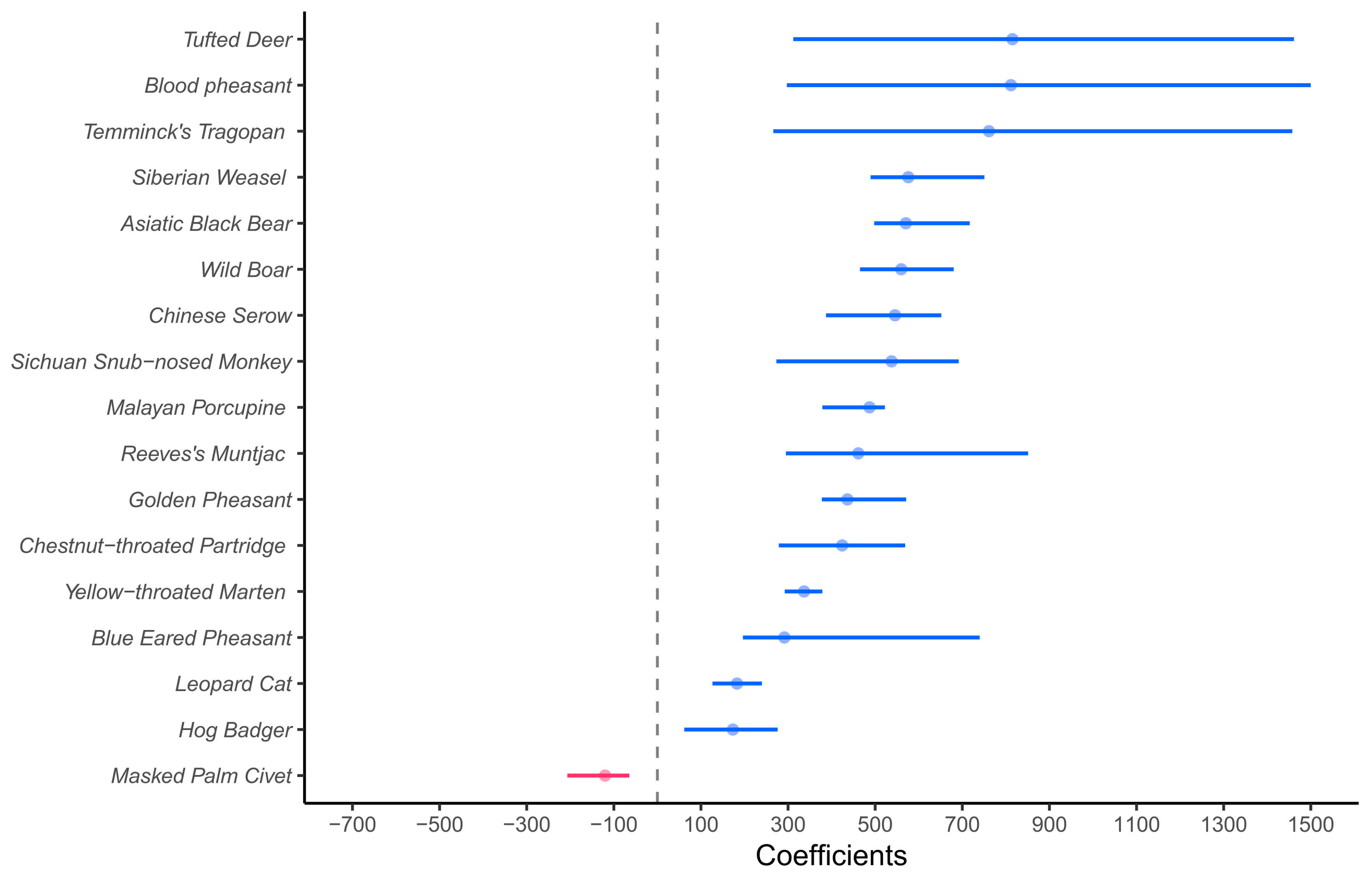
The GLM results showed that, unsurprisingly, the richness of wildlife recorded at sites far from roads was significantly higher than at the BC sites (Table 1). Meanwhile, the results of the GLMM showed that the counts of independent events for each species at sites far from roads were significantly higher than those at the BC sites, except for Masked Palm Civet, Yellow Throat Marten, and Hog Badger (Figure 2).
3.2. Factors Affecting Wildlife Visiting
For the GLMMs investigating the factors that affect the wildlife utilization of BC, the reduced-predictor model was chosen for its lower LOOIC and WAIC compared to the global model (Table 2). Human activities and traffic volume significantly negatively affect the counts of independent events of wildlife at BCs. In contrast, domestic animals significantly positively affected independent events of wildlife. Surprisingly, the coefficient for the coverage of woodland and scrub areas within 1 km radius of the camera sites with zero inflation was positive. This indicates that wildlife was more likely to use BCs with sparse woodland and scrub in the surrounding area.
3.3. Daily Pattern of Visiting
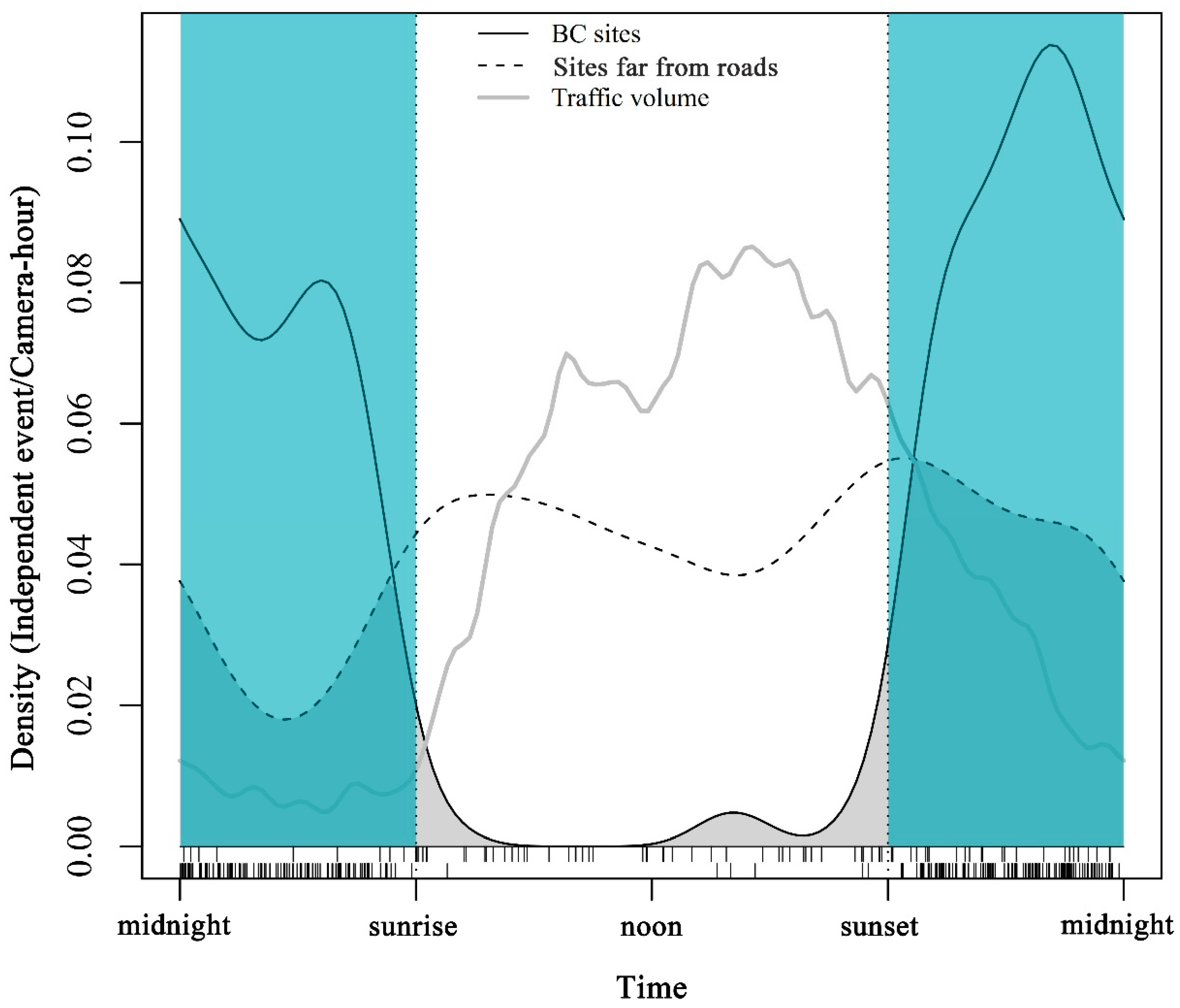
Traffic volumes were dense during the day, especially from 07:00 to 22:00, accounting for 92.09% of the total traffic volume. Overlap analysis shows that the visiting patterns of wildlife at the BC sites were significantly different from those at sites far from roads (overlap estimate = 0.47, 95% confidence interval: 0.37–0.56) (Figure 3).
4. Discussion
Many studies have highlighted that bridges and culverts could help wildlife cross the road and mitigate road effects of fragmentation [25,27]. A survey at Banff Park, Canada, showed that culverts could mitigate the potentially harmful effects of busy transport corridors by providing a vital habitat linkage for many small- and medium-sized mammals [22]. Braschler also found that set-aside railway bridges were able to serve as corridors for dispersal and population communication among small mammals, amphibians, reptiles, and invertebrates [13]. Our study found that bridges and culverts can serve as wildlife passageways. However, despite this region’s rich and abundant wildlife, only a few medium-sized mammals and no large mammals were recorded at the bridge and culvert. It was caused by a combination of at least two negative effects: one was the anthropogenic disturbances introduced by the road, and the second was the unsuitable location or structure of the BC. Many studies have highlighted that bridges and culverts could help wildlife to cross the road and mitigate road effects of fragmentation [25,27]. Nevertheless, it was suggested that bridges and culverts should be modified to address wildlife requirements before they can serve as wildlife crossings [23]. Brunen’s research recommends that the construction of drainage culverts in combination with fences will effectively reduce wildlife mortality [23]. Kenneth and Clevenger’s also suggested reducing road noise and creating appropriate culvert sizes to promote the use of culverts by mammals [22,53]. A substantial network of roads totaling 1192 km is located within the giant panda habitat in Sichuan Province [35]. This region is also inhabited by abundant wildlife, including giant pandas and other threatened species. Numerous bridges and culverts have been constructed along the roads for drainage, since a mountainous terrain and ample rainfall characterize this region. However, none of these infrastructures have undergone ecological modifications. Our results highlight the urgent need for the ecological retrofitting BCs in the GPNP.
The results show that, at BCs, wildlife visits were more common at night. Meanwhile, the wildlife was recorded more during the day than at night at sites distant from roads. The traffic density was low at night. This implies that the high volume of traffic could hinder wildlife from BCs. However, it is still possible that wildlife visited BC at night because wildlife tend to leave the forest and travel to roads at night. Studies found that noise and light from passing vehicles can reduce wildlife’s utilization of road crossing facilities [54]. Nevertheless, for dedicated wildlife crossings, the negative effects of traffic can be mitigated by noise barriers, light shielding, and other traffic mitigation measures [55,56]. Research has shown that traffic levels as high as 17,970 vehicles/day did not significantly impact the utilization of dedicated wildlife underpasses in Banff, Canada [55]. Meanwhile, the traffic volumes in our study were less than 114 vehicles/hour at the peak hour of the day. This highlighted the significance of disturbance mitigation measures for those bridges and culverts [57].
Our results imply facilitation on the part of from domestic animals but interference on the part of human activities in relation to wildlife using bridges and culverts. The negative impact of human activity was not surprising, since most wildlife avoids not only the presence of people, but also places where human activity has just occurred [58,59,60]. In this study, the effect of domestic animals can be regarded as not associated with human activity. Domestic animals at the bridge and culvert, including cattle, chickens, sheep, horses, etc., were free-range with little human management. The impact of domestic animals varies among studies. Some studies found that wildlife avoids livestock, but many suggested that livestock has little impact on wildlife, and can even attract wildlife [61,62].
Medium mammals showed no significant preference for the openness of bridges and culverts in our study. The effects of passages’ openness are still unclear. Most studies suggest that wildlife prefers bridges and culverts with a higher openness for an open field of view [6,25]. Nevertheless, some studies found that wildlife prefers culverts with less openness [63,64].
Surprisingly, wildlife was more likely to cross the roads using bridges and culverts with sparse surrounding forest and scrub compared to those with dense surrounding forest and scrub. Studies have shown that surrounding vegetation cover facilitates the passage of wildlife [65,66,67], yet our results found an opposite trend. It is possible that because of this, wildlife will move quickly in areas with little vegetation, and are not willing to get close to the heavily disturbed road sections when they have nearby forests and scrubs as shelters.
The sites where wildlife crosses roads are clustered in space and characterized by attributes associated with the landscape and road [68,69]. Gu’s research has shown that roads surrounded by abundant suitable habitat are more likely to experience high levels of road mortality [69]. Possibly, the relatively low use of BC by wildlife is because these structures were built in inappropriate places along the road. The locations of bridges and culverts are selected mostly according to hydrology and topography considerations, not to the needs of wildlife. Surveying along the road, including assessing roadkill and wildlife distribution, can help reveal the spatial distribution patterns of wildlife crossing the road and identify sites where potential wildlife crossing structures need to be built.
Overall, existing bridges and culverts provide means for wildlife to cross roads, yet the number of species using them was only a tiny fraction of species present in this region. Although the mechanism leading to the problem has not been fully investigated in this study, our research suggested that these structures require alterations to accommodate wildlife crossing, as they were not originally designed for wildlife. Wildlife is rich in this region, but roads have caused severe habitat fragmentation [70]. The movement of wildlife across roads is expected to be harder in the future, with the busier traffic bought about by the development of tourism and the economy [71]. A highway in this region is under construction to connect Jiuzhaigou Valley, a renowned tourist resort, to the existing highway network. This highlights the urgency of planning dedicated wildlife crossings in this region. A detailed survey of wildlife distribution and activity rhythms along the roads will help identify future sites for wildlife corridors. Mitigation measures, such as noise and light barriers, will be helpful for wildlife at existing bridges and culverts. Besides this, barrier wall–culvert systems, which have been proven to increase the number of species and counts of individuals using BCs [54,71], should be adopted to reduce road deaths.
Supplementary Materials
The following supporting information is available at https://www.mdpi.com/article/10.3390/d15040487/s1. Table Sl: Variance inflation factors (VIF) for variables fit in ZIP GLMM. Table S2: Total independent camera detections at sites of bridges and culverts. Table S3: Total independent camera detections at sites far from roads.
Author Contributions
Methodology and writing—original draft, S.L.; software and formal analysis, Y.Y. and Y.W.; supervision, D.Z., B.Y., Z.Y. and H.L.; project administration and writing—original draft, Q.D. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by the National Natural Science Foundation of China (Grant numbers: 32070520) and Global Environment Facility (GEF) China Protected Land Management Reform Planning Project, China National Park System and Mechanism Innovation Project (C-PAR1).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Raw data are available upon request from the corresponding author.
Acknowledgments
We thank Hanlin Wang (MSc) for the adaptation of the manuscript format.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Trombulak, S.C.; Frissell, C.A. Review of Ecological Effects of Roads on Terrestrial and Aquatic Communities. Conserv. Biol. 2000, 14, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Forman, R.T.T. Estimate of the Area Affected Ecologically by the Road System in the United States. Conserv. Biol. 2000, 14, 31–35. [Google Scholar] [CrossRef]
- Forman, R.T.; Alexander, L.E.J. Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef] [Green Version]
- Laurance, W. If you can’t build well, then build nothing at all. Nature 2018, 563, 295. [Google Scholar] [CrossRef] [Green Version]
- Russo, L.F.; Barrientos, R.; Fabrizio, M.; Di Febbraro, M.; Loy, A. Prioritizing road-kill mitigation areas: A spatially explicit national-scale model for an elusive carnivore. Divers. Distrib. 2020, 26, 1093–1103. [Google Scholar] [CrossRef]
- Clevenger, A.P.; Waltho, N. Factors Influencing the Effectiveness of Wildlife Underpasses in Banff National Park, Alberta, Canada. Conserv. Biol. 2000, 14, 47–56. [Google Scholar] [CrossRef]
- Coffin, A.W. From roadkill to road ecology: A review of the ecological effects of roads. J. Transp. Geogr. 2007, 15, 396–406. [Google Scholar] [CrossRef]
- Wilcox, B.A.; Murphy, D.D. Conservation Strategy: The Effects of Fragmentation on Extinction. Am. Nat. 1985, 125, 879–887. [Google Scholar] [CrossRef]
- Madadi, H.; Moradi, H.; Soffianian, A.; Salmanmahiny, A.; Senn, J.; Geneletti, D. Degradation of natural habitats by roads: Comparing land-take and noise effect zone. Environ. Impact Assess. Rev. 2017, 65, 147–155. [Google Scholar] [CrossRef]
- Geneletti, D.; Dawa, D. Environmental impact assessment of mountain tourism in developing regions: A study in Ladakh, Indian Himalaya. Environ. Impact Assess. Rev. 2009, 29, 229–242. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e15000522015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceia-Hasse, A.; Navarro, L.M.; Borda-de-Água, L.; Pereira, H.M. Population persistence in landscapes fragmented by roads: Disentangling isolation, mortality, and the effect of dispersal. Ecol. Model. 2018, 375, 45–53. [Google Scholar] [CrossRef]
- Braschler, B.; Dolt, C.; Baur, B. The Function of A Set-Aside Railway Bridge in Connecting Urban Habitats for Animals: A Case Study. Sustainability 2020, 12, 1194. [Google Scholar] [CrossRef] [Green Version]
- Ceia-Hasse, A.; Borda-de-Água, L.; Grilo, C.; Pereira, H.M. Global exposure of carnivores to roads. Glob. Ecol. Biogeogr. 2017, 26, 592–600. [Google Scholar] [CrossRef] [Green Version]
- Rytwinski, T.; Fahrig, L. Do species life history traits explain population responses to roads? A meta-analysis. Biol. Conserv. 2012, 147, 87–98. [Google Scholar] [CrossRef]
- Corlatti, L.; Hackländer, K.; Frey-Roos, F. Ability of Wildlife Overpasses to Provide Connectivity and Prevent Genetic Isolation. Conserv. Biol. 2009, 23, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Glista, D.J.; DeVault, T.L.; DeWoody, J.A. A review of mitigation measures for reducing wildlife mortality on roadways. Landsc. Urban Plan. 2009, 91, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sawaya, M.A.; Clevenger, A.P.; Kalinowski, S.T. Demographic Connectivity for Ursid Populations at Wildlife Crossing Structures in Banff National Park. Conserv. Biol. 2013, 27, 721–730. [Google Scholar] [CrossRef]
- van der Grift, E.A.; van der Ree, R.; Fahrig, L.; Findlay, S.; Houlahan, J.; Jaeger, J.A.G.; Klar, N.; Madriñan, L.F.; Olson, L. Evaluating the effectiveness of road mitigation measures. Biodivers. Conserv. 2013, 22, 425–448. [Google Scholar] [CrossRef] [Green Version]
- Seidler, R.G.; Green, D.S.; Beckmann, J.P. Highways, crossing structures and risk: Behaviors of Greater Yellowstone pronghorn elucidate efficacy of road mitigation. Glob. Ecol. Conserv. 2018, 15, e00416. [Google Scholar] [CrossRef]
- Ascensão, F.; Mira, A. Factors affecting culvert use by vertebrates along two stretches of road in southern Portugal. Ecol. Res. 2007, 22, 57–66. [Google Scholar] [CrossRef]
- Clevenger, A.P.; Chruszcz, B.; Gunson, K. Drainage culverts as habitat linkages and factors affecting passage by mammals. J. Appl. Ecol. 2001, 38, 1340–1349. [Google Scholar] [CrossRef]
- Brunen, B.; Daguet, C.; Jaeger, J.A.G. What attributes are relevant for drainage culverts to serve as efficient road crossing structures for mammals? J. Environ. Manag. 2020, 268, 110423. [Google Scholar] [CrossRef]
- Ng, S.J.; Dole, J.W.; Sauvajot, R.M.; Riley, S.P.D.; Valone, T.J. Use of highway undercrossings by wildlife in southern California. Biol. Conserv. 2004, 115, 499–507. [Google Scholar] [CrossRef]
- Grilo, C.; Bissonette, J.A.; Santos-Reis, M. Response of carnivores to existing highway culverts and underpasses: Implications for road planning and mitigation. Biodivers. Conserv. 2008, 17, 1685–1699. [Google Scholar] [CrossRef]
- Villalva, P.; Reto, D.; Santos-Reis, M.; Revilla, E.; Grilo, C. Do dry ledges reduce the barrier effect of roads? Ecol. Eng. 2013, 57, 143–148. [Google Scholar] [CrossRef]
- Wang, Y.; Guan, L.; Chen, J.; Kong, Y. Influences on mammals frequency of use of small bridges and culverts along the Qinghai–Tibet railway, China. Ecol. Res. 2018, 33, 879–887. [Google Scholar] [CrossRef]
- Bhardwaj, M.; Olsson, M.; Seiler, A. Ungulate use of non-wildlife underpasses. J. Environ. Manag. 2020, 273, 111095. [Google Scholar] [CrossRef]
- Clevenger, A.; Barrueto, M. Trans-Canada Highway Wildlife and Monitoring Research, Final Report. Part B: Research; Western Transportation Institute: Bozeman, MT, USA, 2014. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J.J.N. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Xu, W.; Viña, A.; Kong, L.; Pimm, S.L.; Zhang, J.; Yang, W.; Xiao, Y.; Zhang, L.; Chen, X.; Liu, J.; et al. Reassessing the conservation status of the giant panda using remote sensing. Nat. Ecol. Evol. 2017, 1, 1635–1638. [Google Scholar] [CrossRef]
- He, K.; Dai, Q.; Gu, X.; Zhang, Z.; Zhou, J.; Qi, D.; Gu, X.; Yang, X.; Zhang, W.; Yang, B.; et al. Effects of roads on giant panda distribution: A mountain range scale evaluation. Sci. Rep. 2019, 9, 1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Swaisgood, R.R.; Dai, Q.; Yang, Z.; Yuan, S.; Owen, M.A.; Pilfold, N.W.; Yang, X.; Gu, X.; Zhou, H.; et al. Giant panda distributional and habitat-use shifts in a changing landscape. Conserv. Lett. 2018, 11, e12575. [Google Scholar] [CrossRef]
- Qiu, L.; Han, H.; Zhou, H.; Hong, M.; Zhang, Z.; Yang, X.; Gu, X.; Zhang, W.; Wei, W.; Dai, Q. Disturbance control can effectively restore the habitat of the giant panda (Ailuropoda melanoleuca). Biol. Conserv. 2019, 238, 108233. [Google Scholar] [CrossRef]
- Administration, S.F. The Fourth National Giant Panda Survey. State Forestry Administration of China Press: Beijing, China, 2015. [Google Scholar]
- Tanwar, K.S.; Sadhu, A.; Jhala, Y.V. Camera trap placement for evaluating species richness, abundance, and activity. Sci. Rep. 2021, 11, 23050. [Google Scholar] [CrossRef]
- Burton, A.C.; Neilson, E.; Moreira, D.; Ladle, A.; Steenweg, R.; Fisher, J.T.; Bayne, E.; Boutin, S. REVIEW: Wildlife camera trapping: A review and recommendations for linking surveys to ecological processes. J. Appl. Ecol. 2015, 52, 675–685. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote. Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Huang, H.; Chen, Y.; Clinton, N.; Wang, J.; Wang, X.; Liu, C.; Gong, P.; Yang, J.; Bai, Y.; Zheng, Y.; et al. Mapping major land cover dynamics in Beijing using all Landsat images in Google Earth Engine. Remote Sens. Environ. 2017, 202, 166–176. [Google Scholar] [CrossRef]
- Lovell, C.; Li, S.; Turner, J.; Carbone, C. The effect of habitat and human disturbance on the spatiotemporal activity of two urban carnivores: The results of an intensive camera trap study. Ecol. Evol. 2022, 12, e8746. [Google Scholar] [CrossRef]
- Greenville, A.C.; Wardle, G.M.; Dickman, C.R. Desert mammal populations are limited by introduced predators rather than future climate change. R. Soc. Open Sci. 2017, 4, 170384. [Google Scholar] [CrossRef] [Green Version]
- Etana, B.; Atickem, A.; Tsegaye, D.; Bekele, A.; De Beenhouwer, M.; Hundera, K.; Lens, L.; Fashing, P.J.; Stenseth, N.C. Traditional shade coffee forest systems act as refuges for medium- and large-sized mammals as natural forest dwindles in Ethiopia. Biol. Conserv. 2021, 260, 109219. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R.; Springer: New York, NY, USA, 2009; Volume 574. [Google Scholar]
- Reed, D.; Ward, A. Efficacity of methods advocated to reduce deer vehicle accidents: Research and rationale in the USA. In Proceedings of the Highway and Wildlife Relationships, Symposium, Strasbourg, France, 5–7 June 1985. [Google Scholar]
- Gehrt, S.D.; Prange, S.J.B.E. Interference competition between coyotes and raccoons: A test of the mesopredator release hypothesis. Behav. Ecol. 2007, 18, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Ripple, W.J.; Estes, J.A.; Beschta, R.L.; Wilmers, C.C.; Ritchie, E.G.; Hebblewhite, M.; Berger, J.; Elmhagen, B.; Letnic, M.; Nelson, M.P. Status and ecological effects of the world’s largest carnivores. Science 2014, 343, 1241484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansfield, E.R.; Helms, B.P. Detecting Multicollinearity. Am. Stat. 1982, 36, 158–160. [Google Scholar] [CrossRef]
- Barrow, L.N.; McNew, S.M.; Mitchell, N.; Galen, S.C.; Lutz, H.L.; Skeen, H.; Valqui, T.; Weckstein, J.D.; Witt, C.C. Deeply conserved susceptibility in a multi-host, multi-parasite system. Ecol. Lett. 2019, 22, 987–998. [Google Scholar] [CrossRef] [PubMed]
- Golivets, M.; Woodall, C.W.; Wallin, K.F. Functional form and interactions of the drivers of understory non-native plant invasions in northern US forests. J. Appl. Ecol. 2019, 56, 2596–2608. [Google Scholar] [CrossRef]
- Bürkner, P.-C. brms: An R Package for Bayesian Multilevel Models Using Stan. J. Stat. Softw. 2017, 80, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Meredith, M.; Ridout, M. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 2018, 14, 322–337. [Google Scholar]
- Ridout, M.S.; Linkie, M. Estimating overlap of daily activity patterns from camera trap data. JABES 2009, 14, 322–337. [Google Scholar] [CrossRef]
- Dodd, C.K.; Barichivich, W.J.; Smith, L.L. Effectiveness of a barrier wall and culverts in reducing wildlife mortality on a heavily traveled highway in Florida. Biol. Conserv. 2004, 118, 619–631. [Google Scholar] [CrossRef]
- Sołowczuk, A.J.S. Effect of Landscape Elements and Structures on the Acoustic Environment on Wildlife Overpasses Located in Rural Areas. Sustainability 2020, 12, 7866. [Google Scholar] [CrossRef]
- Barrueto, M.; Ford, A.T.; Clevenger, A.P. Anthropogenic effects on activity patterns of wildlife at crossing structures. Ecosphere 2014, 5, art27. [Google Scholar] [CrossRef]
- Gagnon, J.W.; Dodd, N.L.; Ogren, K.S.; Schweinsburg, R.E. Factors associated with use of wildlife underpasses and importance of long-term monitoring. J. Wildl. Manag. 2011, 75, 1477–1487. [Google Scholar] [CrossRef]
- Barber, J.R.; Crooks, K.R.; Fristrup, K.M. The costs of chronic noise exposure for terrestrial organisms. Trends Ecol. Evol. 2010, 25, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Hammitt, W.E.; Cole, D.N.; Monz, C.A. Wildland Recreation: Ecology and Management; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Erb, P.L.; McShea, W.J.; Guralnick, R.P. Anthropogenic influences on macro-level mammal occupancy in the Appalachian trail corridor. PLoS ONE 2012, 7, e42574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queiroz, C.; Beilin, R.; Folke, C.; Lindborg, R. Farmland abandonment: Threat or opportunity for biodiversity conservation? A global review. Front. Ecol. Environ. 2014, 12, 288–296. [Google Scholar] [CrossRef]
- Gordon, I.J. Review: Livestock production increasingly influences wildlife across the globe. Animal 2018, 12, s372–s382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schieltz, J.M.; Rubenstein, D.I. Evidence based review: Positive versus negative effects of livestock grazing on wildlife. What do we really know? Environ. Res. Lett. 2016, 11, 113003. [Google Scholar] [CrossRef] [Green Version]
- Clevenger, A.P.; Waltho, N. Dry drainage culvert use and design considerations for small-and medium-sized mammal movement across a major transportation corridor. In Proceedings of the Third International Conference on Wildlife Ecology and Transportation Florida Department of Transportation US Department of Transportation US Forest Service Defenders of Wildlife, Missoula, MT, USA, 13–16 September 1999. [Google Scholar]
- McDonald, W.; Clair, C.C.S. Elements that promote highway crossing structure use by small mammals in Banff National Park. J. Appl. Ecol. 2004, 41, 82–93. [Google Scholar] [CrossRef]
- Roads, roadsides and wildlife conservation: A review: A.F. Bennett, in: Nature conservation 2: The role of corridors, ed D.A. Saunders & R.J. Hobbs, (Surrey Beatty). Biol. Conserv. 1991, 60, 64. [CrossRef]
- Gunson, K.E.; Mountrakis, G.; Quackenbush, L.J. Spatial wildlife-vehicle collision models: A review of current work and its application to transportation mitigation projects. J. Environ. Manag. 2011, 92, 1074–1082. [Google Scholar] [CrossRef]
- Ramp, D.; Wilson, V.K.; Croft, D.B. Assessing the impacts of roads in peri-urban reserves: Road-based fatalities and road usage by wildlife in the Royal National Park, New South Wales, Australia. Biol. Conserv. 2006, 129, 348–359. [Google Scholar] [CrossRef]
- Galinskaitė, L.; Ulevičius, A.; Valskys, V.; Samas, A.; Busher, P.E.; Ignatavičius, G. The Influence of Landscape Structure on Wildlife–Vehicle Collisions: Geostatistical Analysis on Hot Spot and Habitat Proximity Relations. ISPRS Int. J. Geo Inf. 2022, 11, 63. [Google Scholar]
- Gu, H.; Dai, Q.; Wang, Q.; Wang, Y. Factors contributing to amphibian road mortality in a wetland. Curr. Zool. 2011, 57, 768–774. [Google Scholar] [CrossRef]
- Li, T.; Shilling, F.; Thorne, J.; Li, F.; Schott, H.; Boynton, R.; Berry, A.M. Fragmentation of China’s landscape by roads and urban areas. Landsc. Ecol. 2010, 25, 839–853. [Google Scholar] [CrossRef] [Green Version]
- Forman, R.T.; Sperling, D.; Bissonette, J.A.; Clevenger, A.P.; Cutshall, C.D.; Dale, V.H.; Fahrig, L.; France, R.L.; Goldman, C.R.; Heanue, K. Road Ecology: Science and Solutions; Island Press: Washington, DC, USA, 2003. [Google Scholar]
Figure 1.
Study area in Pingwu county, north of Giant Panda National Park, Sichuan, China.

Figure 2.
Coefficient plot displaying the regression coefficients of zero-inflated generalized linear mixed models (ZINB) comparing independent events of wildlife at BC and sites far from roads. Solid circles indicate the estimated coefficients for events in the models, their 95% confidence intervals are presented by the solid line, and the vertical dashed line indicates the value of 0. The color blue means that the count of independent events at sites far from roads was significantly higher than those at BC (the lower limit of confidence interval was greater than 0). The color red indicates that the count of independent events at sites far from roads was significantly less than that at BC (the upper limit of confidence interval was less than 0).
Figure 2.
Coefficient plot displaying the regression coefficients of zero-inflated generalized linear mixed models (ZINB) comparing independent events of wildlife at BC and sites far from roads. Solid circles indicate the estimated coefficients for events in the models, their 95% confidence intervals are presented by the solid line, and the vertical dashed line indicates the value of 0. The color blue means that the count of independent events at sites far from roads was significantly higher than those at BC (the lower limit of confidence interval was greater than 0). The color red indicates that the count of independent events at sites far from roads was significantly less than that at BC (the upper limit of confidence interval was less than 0).

Figure 3.
Overlap between daily visiting patterns of medium animals at BC sites (solid line) and the sites far from roads (dotted line). The grey line shows the traffic volume along the road. Time of day is presented on the x-axis. The thin dotted line represents the standardized sunrise time and sunset time respectively. The blue area in the figure represents night time.
Figure 3.
Overlap between daily visiting patterns of medium animals at BC sites (solid line) and the sites far from roads (dotted line). The grey line shows the traffic volume along the road. Time of day is presented on the x-axis. The thin dotted line represents the standardized sunrise time and sunset time respectively. The blue area in the figure represents night time.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of generalized linear model results for examining the difference in wildlife richness at sites of bridge and culvert and sites for control at sites far from roads.
Table 1.
Summary of generalized linear model results for examining the difference in wildlife richness at sites of bridge and culvert and sites for control at sites far from roads.
| Model | Variable | Coefficient | SE | Lower95% | Upper95% | R2 |
|---|---|---|---|---|---|---|
| Richness~site | Site: Far | 280.65 | 55.08 | 171.74 | 390.29 | 0.50 |
Note: “Site: Far” represent the dummy variable for far from road.
Table 2.
The LOOIC and R2 for zero-inflated Poisson (ZIP) GLMM. WAIC results were consistent with LOOIC. Reduced models include the set of predictors that were considered important in the global models (95% CI did not cross 0). The difference between each model and the best-fit model is shown as Δelpd (expected log predictive density) with standard error (SE).
Table 2.
The LOOIC and R2 for zero-inflated Poisson (ZIP) GLMM. WAIC results were consistent with LOOIC. Reduced models include the set of predictors that were considered important in the global models (95% CI did not cross 0). The difference between each model and the best-fit model is shown as Δelpd (expected log predictive density) with standard error (SE).
| Model | LOOIC | WAIC | Δelpd (SE) | R2 | Variables | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Human | Traffic: Low | DA | Openness | Cover50 | Cover1 km | Zi-Cover1 km | |||||
| Global | 251.70 | 248.60 | −0.30 (0.70) | 0.49 | −0.17 (−0.30, −0.04) | 0.45 (0.15, 0.75) | 0.22 (0.05, 0.39) | −0.56 (−4.29, 3.41) | 0.79 (−19.16, 21.22) | −19.60 (−92.98, 39.78) | 19.12 (0.50, 37.69) |
| Reduced | 251.10 | 248.00 | 0 | 0.49 | −0.17 (−0.31, −0.04) | 0.46 (0.17, 0.76) | 0.21 (0.05, 0.37) | - | - | - | 20.93 (3.89, 38.05) |
Note: Variables statistics include the parameter estimates and 95% confidence intervals. Human: Human independent events of visitation. DA: domestic animals. Traffic: the traffic volume levels were divided into low and high. Cover50: the coverage of scrub and woodland within 50 m. Cover1km: the coverage of scrub and woodland within 1 km. Zi-Cover1 km: the zero-inflation probability for the variable of coverage of scrub and woodland within 1 km.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lu, S.; Yue, Y.; Wang, Y.; Zhang, D.; Yang, B.; Yu, Z.; Lin, H.; Dai, Q. The Factors Influencing Wildlife to Use Existing Bridges and Culverts in Giant Panda National Park. Diversity 2023, 15, 487. https://doi.org/10.3390/d15040487
AMA Style
Lu S, Yue Y, Wang Y, Zhang D, Yang B, Yu Z, Lin H, Dai Q. The Factors Influencing Wildlife to Use Existing Bridges and Culverts in Giant Panda National Park. Diversity. 2023; 15(4):487. https://doi.org/10.3390/d15040487
Chicago/Turabian StyleLu, Song, Ying Yue, Yihong Wang, Dawei Zhang, Biao Yang, Zhen Yu, Honghui Lin, and Qiang Dai. 2023. "The Factors Influencing Wildlife to Use Existing Bridges and Culverts in Giant Panda National Park" Diversity 15, no. 4: 487. https://doi.org/10.3390/d15040487
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.