1. Introduction
During the last century, the world has witnessed the construction of an enormous road network. From 1957 to 2012, the European road network alone increased from 146.7 million to 421.8 million kilometers [
1]. Besides the devastating consequences associated with the destruction and fragmentation of the landscape, road traffic further impacts biodiversity through the emission of pollution (e.g., chemical, light, and noise) [
2,
3,
4] The chemical pollution (e.g., hydrocarbons and heavy metals), washed off the road surface by rain, originates from tires, fuel residues, and the asphalt used to pave roads [
5]. To mitigate the impact of such chemical pollution, roadside stormwater ponds have been constructed [
6,
7]. These stormwater ponds (SWPs) are used to collect, store, and clean up runoff water from the roads before it is released into the environment [
6]. The name “stormwater” pond refers to the design of these water reservoirs to collect all water, even in case of maximum rainfall, i.e., during a storm. Despite their primary role to retain road pollutants, some species, especially amphibians, breed inside SWPs, at least in North America and Western Europe, [
8,
9,
10,
11]. This has led to the suggestion that SWPs might act as ecological traps for such species [
12,
13,
14]. Indeed, numerous studies have demonstrated the negative effects of roadside pollutants (e.g., heavy metals, salts) on amphibians, particularly during larval development [
12,
15,
16,
17]. Some of these pollutants might cause changes in the physiology and morphology of affected animals, such as a reduction in metabolism or an increasing risk of malformations, leading to a reduced larval survival [
12,
18,
19,
20,
21]. It has also been shown that these pollutants (e.g., copper, lead) can negatively affect the behavior of amphibians, such as their capacity to escape due to decreased swim speed [
21,
22]. However, pollutant concentration in SWPs varies greatly between roads and also between countries, as road types, traffic volume, and pond management practices (including dredging frequency) differ [
10,
23].
Although polluted, in a situation where the natural habitats of amphibians are in steep decline, SWPs are increasingly perceived as potential substitute biotopes [
10]. Globally, 41% of amphibian species are threatened with extinction [
24] and the main cause for such population erosion is the loss of natural habitat [
25,
26]. Worldwide wetland surfaces, essential for amphibians, have decreased by more than 70% in the last century alone [
27]. Since SWPs are built alongside roads, they might act as relay sites (in a stepping-stone configuration) for the recolonization of wetlands by providing a corridor for amphibian species [
11,
28].
One species particularly concerned by such a scenario is the European green toad (
Bufotes viridis). In the Bas-Rhin plain region (Alsace, eastern France), which is host to the largest green toad population in France, the majority of individuals are found in SWPs [
29]. This is likely associated with the damming of the Rhine River in the 20th century, which destroyed the natural habitat of green toads in the region (open banks of large rivers). In the following, toads moved to various semi-natural ponds (SNPs; quarries and ponds developed for the species) for survival and, upon their implementation during the 1990s, invaded SWPs. Hence, it is of great importance to understand the ecological role that SWPs play for this protected species (ecological trap or suitable breeding habitat), so that the design and management practices for these structures can be adapted. Conan and colleagues (2022) studied the growth and survival of European green toad tadpoles within SWPs, which was compared with the situation in SNPs. The latter were used as “control sites” because they are located far from major highways and their associated pollution. Results from that study showed a reduced larval survival inside SWPs (~0% in SWPs vs. ~21% in SNPs), supporting the view that SWPs might act as an ecological trap. The exact reason(s) for the reduced survival in SWPs remain(s) unknown. However, the above study showed that sediments inside SWPs had a five-fold higher concentration of pollutants than sediments of the SNPs, while water pollution levels did not differ between pond types. In addition, a leech species (
Helobdella stagnalis) was present in SWPs (and was found attached to adults and tadpoles) but not in SNPs.
Research Objectives
The present study is a follow up to the above-mentioned study [
30] to investigate the potential reasons for the high tadpole mortality observed in SWPs. To this end, we studied the effects of sediment pollution and leech presence on the survival of European green toad tadpoles in a controlled laboratory environment. In the following, our investigation covered the larval development of green toads from hatching to metamorphosis. Firstly, we followed the survival and growth of tadpoles until metamorphosis. In addition, we conducted a number of experiments to study how pollution might affect the behavior of tadpoles (swim tests to assess locomotion capacity) and the metabolism of metamorphosed individuals (respirometry trials to measure standard metabolic rate).
Based on the published literature, we expected that sediment pollution will (1) have an overall negative effect on tadpole survival, lead (2) to an accelerated growth rate, (3) lead to a decrease in swim speed [
12,
31], and (4) lead to an increase in standard metabolic rate. We furthermore expected that (5) the presence of leeches will also reduce tadpole survival.
2. Materials and Methods
2.1. Egg Collection and Housing
On 17 May 2021, we collected fragments from five different clutches of European green toads (
Bufotes viridis) in a semi-natural pond (Bischoffsheim, 48°28′58.4″ N, 7°30′42.7″ E), containing at least 200 eggs each. Larval survival at the concerned pond was ~70% during the year preceding the current study [
30]. Clutch fragments were transported in a plastic bucket of water from the sampling site to the laboratory (IPHC-DEPE, Strasbourg). Eggs were incubated inside an aquarium at 20 °C, until hatching. Water inside the aquarium consisted of water from the collection site (50%) and of clean tap water (50%), continuously oxygenated with an air pump.
2.2. Leech Collection (Helobdella stagnalis)
On 4 May 2021, we collected approximately 200 individuals of Helobdella stagnalis in one of the SWPs (Rosheim, 48°30′12.9″ N, 7°29′57.7″ E), where both leeches and breeding European green toads were present. Leeches were collected by hand, removing them from adult toads and our boots (after boot immersion). They were transported to the laboratory within a plastic bucket, filled with water from the sampling site, where they rested overnight, before being introduced to the experimental setup.
2.3. Sediment Samples (SWPs)
We collected approximatively 5 L of sediment from 3 SWPs in May 2021. The sediment at these sites was found to be the most polluted of all sites during our previous study [
30]. The sediment was thoroughly mixed and freeze-dried in the laboratory to kill the majority of its life forms without modifying its biochemistry. About 2.5 L of sediment were used for the experimental setup, while the rest was stored for pollutant analysis.
2.4. Experimental Design
For our investigation, we set up four large glass tanks (120 × 50 × 35 cm, L × W × H) that each contained three smaller glass tanks (40 × 30 × 30 cm, L × W × H). Different environmental conditions, concerning sediment pollution and leech presence, were realized within the four tanks, while the three smaller glass tanks within each large tank served as replicates (
Figure 1). All tanks were filled with a 2 cm layer of sediment and tap water (up to a height of 28 cm), which was continuously oxygenated by bubbling air through it (air pump “ProSilent a200”, JBL, Neuhofen, Germany) and mechanically filtered (“CristalProfi e1502 greenline”, JBL, Neuhofen, Germany). We set up the following four conditions: (1) in the “
control” condition, tanks were filled with a clean sediment layer (bought from a pet store; “Sansibar white”, JBL, Neuhofen, Germany) and water. (2) Tanks representing the “
leech” condition were filled with clean sediment (as in condition 1), water, and 20 leeches per replicate. (3) The “
SWP sediment” condition contained polluted sediment from the SWPs and water. (4) The “
SWP sediment + leech” condition also contained polluted sediment from the SWPs, water, and, in addition, 20 leeches per replicate. Hence, leech density per tank might seem high but was actually lower than what we observed in enclosures in situ [
30]. The water quality (nitrate, nitrite, ammonium) was analyzed twice a week with colorimetric tests (“Combiset Test Plus NH
4”, JBL, Neuhofen, Germany) to avoid exposure to nitrogen peaks. Each tank was set up carefully, depositing the sediment first and avoiding disturbance when adding water. All tanks were left for a period of 3 weeks so that conditions could stabilize (water quality, settling of suspended sediment). Thereafter (2 June 2021), tadpoles and leeches were added to each replicate tank. A total of 480 tadpoles were used during the experiment (40 tadpoles per replicate tank, 3 replicate tanks per condition). Tadpole distribution was random for each replicate.
2.5. Sediment Pollution Analysis
To ensure that sediment collected from the SWPs had pollution levels similar to those of our previous study, concentrations for the various pollutants were quantified before experimentation. The freeze-dried sediment was first crushed and then sieved to obtain a homogeneous sediment with a particle size of less than 125 µm [
32]. The pollutant analysis was conducted with a portable X-ray Fluorescence Analyzer (“Vanta”; Olympus, Tokyo, Japan) that had been carefully calibrated beforehand (Lenormand et al.
in press). The shooting-time was set to 63 s and three replicates per sediment sample, which were kept inside a 10 cm
3 plastic cup covered with Parafilm
TM, were analyzed (USEPA 2017).
2.6. Survival and Growth Assessment
To monitor the growth and survival of tadpoles, length measurements and counting of survivors were conducted three times per week until all four legs of the tadpoles were well developed (Gosner, stage 42) [
33]. For measuring, tadpoles were moved from the respective setup and placed into a small plastic container (1 cm water layer) with a ruler at its bottom. Pictures were taken with a digital camera (“Tough 5”; Olympus, Tokyo, Japan), the number of surviving tadpoles was counted, and their size determined (from snout to base of tail) using the pictures and Image J software (V1.52a, Development Team, 2020). With all four legs developed (Gosner, stage 42), tadpoles switch from an aquatic to an aerial breathing system. Accordingly, at this stage of development, we placed individuals into a terrarium containing both an aquatic and terrestrial part, to avoid drowning, and size measurements were discontinued (also because body size decreases at that stage). The surviving metamorphosed tadpoles (“toadlets”) were kept and used in respirometry trials.
2.7. Swim Tests
To investigate the potential impact of pollution on the locomotion capacity of tadpoles (i.e., swim speed), individuals with well-developed hind legs (Gosner, stage 40; ~42 days after the start of experimentation) were subjected to swim tests. At the start of a trial, an individual was placed in an open PVC tube that served as a swim channel with a graded scale at the bottom (
Figure 2). Within 30 s after placement, the individual was stimulated to swim by gently touching the tail with a stick. This was repeated three times per trial, leaving 30 s between repetitions. We filmed the trials with a digital camera (“Tough 5”) and analyzed video with Tracker software (V6.0.1, Development Team, 2021) to estimate the swim speed of each individual. Swim tests were conducted with three individuals per replicate tank (i.e., N = 9 individuals per condition). However, since tadpoles exposed to conditions 2 and 4 (leech presence) died within the first 2 weeks of experimentation, these tests were only conducted for conditions 1 and 3.
2.8. Respirometry Trials
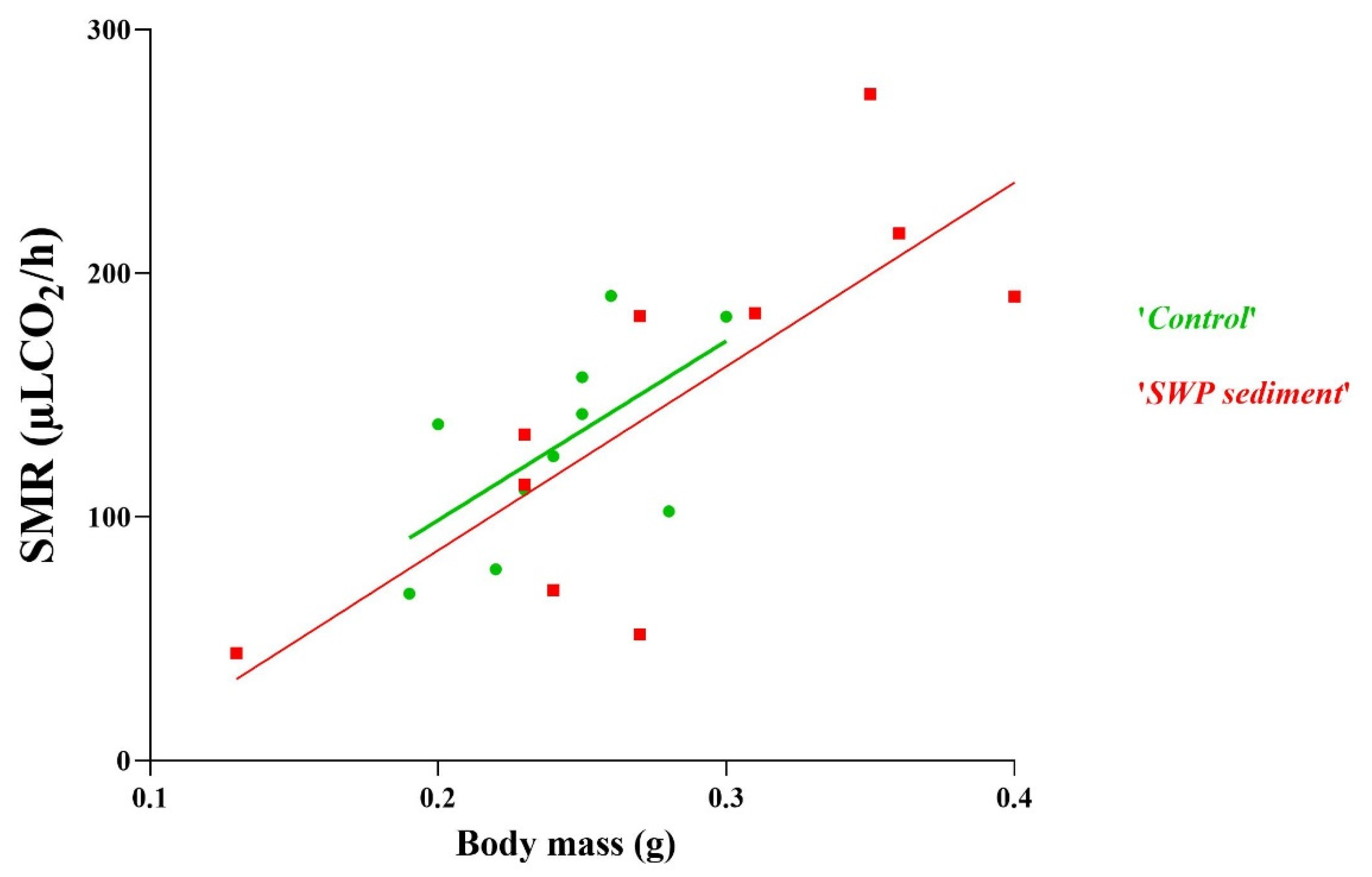
To investigate the potential effect of pollutants on metabolic rate, we recorded the CO2 production rate of freshly metamorphosed toadlets (Gosner, stage 46, four legs and fully resorbed tail), exposed to conditions 1 (‘control’) and 3 (‘SWP sediment’). Given the high mortality of tadpoles in conditions 2 and 4, no individuals reached metamorphosis and, hence, these groups could not be included in respirometry measurements.
We conducted respirometry trials with 10 toadlets per tested condition (i.e., N = 20). Mean body mass of toadlets was 0.26 g (range: 0.13–0.40 g; Aroma-Zone
® precision balance, 0.1–500 g, resolution: 0.01 g). The metabolic rate of individuals was determined by recording the CO
2 production rate within an open circuit respirometry system (FoxBox, Sable Systems, Las Vegas, NV, USA). The respirometry system was calibrated before each trial, using 0.5% CO
2 (Air Liquide, Paris, France) for the span and 0% CO
2 for the zero settings, respectively. For the latter, we used room air that was pulled through a tube with Ascarite and Drierite. A 5 mL transparent plastic syringe with a 12.5 mm diameter served as the metabolic chamber through which air was pulled at a rate of 44 mL/min and fed into the gas analyzer. Water vapor and carbon dioxide were removed from the incurrent air using Drierite and Ascarite, respectively. The time constant of the system was calculated to be 6.5 s. All trials were conducted at a room temperature of 21 °C. Baseline readings of background CO
2 and water vapor pressure were recorded using a baselining unit (CB-2, Sable Systems International, Las Vegas, NV, USA) for at least 1 min before and after a measurement period with toadlets in the respirometry chamber. After the initial disturbance upon introduction to the chamber, a toadlet remained calm for the rest of a trial, during which CO
2 concentration inside the chamber and flow rate through the chamber were recorded. Trials lasted ~15 min for each individual and resting metabolic rate (Standard Metabolic Rate at 21 °C, SMR) was calculated from CO
2 production rate, using a modification of equation 10.4 of Lighton [
34]:
, where
is the CO
2 production rate (µL/h),
is the main flow rate leaving the chamber (ml/min),
is the fractional concentration of CO
2 leaving the chamber, and
is the fractional concentration of CO
2 entering the chamber (equal to zero, as CO
2 in the incurrent air was scrubbed).
was calculated as the lowest stable value over 60 s, taken from a pre-selected stable 8 min section. Data were recorded and analyzed using ExpeData (Sable Systems, Las Vegas, NY, USA).
All procedures were carried out after obtaining the authorizations for capture, transport and the detention of specimen of the various species in captivity (DDPP67-SPAE-FSC-2019-04). The experimental protocol was approved by the French Ethical Committee (CREMEAS) and the French Ministry of Environment: “Ministère de la Transition Écologique” under agreement number (APAFIS #28740-2020120913291952 v4). At the end of experimentation, all surviving toadlets were released at the capture site.
2.9. Statistical Analysis
We used the Kaplan–Meier survival analysis and Log-rank tests to compare the survival rate of tadpoles between the different conditions [
35]. A generalized additive model (GAM) with Gaussian distribution was used to investigate potential size differences over time between the various conditions (size~condition + time; using package “mgcv” in R [
36]). An analysis of variance (ANOVA) based on a linear mixed model (LMM) with the individual as a random factor was used to compare the swim speed of tadpoles exposed to condition 1 (‘
control’) versus condition 3 (‘
SWP sediment’) (speed~condition + 1/individual), using package “nmle” in R [
37]. Finally, to investigate potential differences in metabolic rate between toadlets of conditions 1 and 3, we used a multivariate analysis of variance (MANOVA-Pillai test). In our experimental set-up, we placed three smaller tanks inside each of the four larger glass tanks, which were intended to serve as replicates for each condition. However, since the water of all smaller tanks within each larger tank was on the same water circulation/filtering circuit, they cannot be considered as independent replicates. Hence, we accounted for this during the statistical analysis by removing all “replicates”, so that we only test for differences between the four large glass tanks.
All statistical analyses and graphs were realized with R (v3.5.5, RStudio v1.2.1335, R Core Team, Vienna, Austria, 2020) and GraphPad Prism 8.2.1 (San Diego, CA, USA), respectively. Results are presented as means ± SEM with a statistical significance level of 5%.
4. Discussion
The results of our current study clearly demonstrate the strong effect that the presence of leeches has on tadpole survival. While the possibility remains that the polluted sediments of artificial SWPs contributed to the high mortality of tadpoles that we found previously in situ [
30] and under controlled conditions (this study), the prominent role of
Helobdella stagnalis is now obvious. A leech can detect a tadpole, attach itself to it, and devour it within a few minutes. Leech predation on tadpoles at the abundance level tested (20 leeches for 40 tadpoles) was high, so that no tadpole survived for more than 15 days, regardless of the pollution level of the substrate (conditions 3 and 4). Interestingly, even when providing alternative food for the leeches ad libitum (condition 2b), they continued to devour tadpoles, suggesting a strong attraction for tadpoles. Nevertheless, further studies are needed to investigate the exact relationship between leeches and tadpoles/toads in a natural/semi-natural setting. While in our previous study [
30] we found an even greater leech density inside the tadpole enclosures of the SWPs, it is not clear to what degree this might have reflected leech density in the pond. Leeches likely detected the tadpoles inside the enclosures and were able to target them, while tadpoles were unable to escape. However, in that study, we also regularly detected freely moving adult green toads that had up to 20 leeches attached to them, illustrating the capacity of leeches to successfully target toads under natural conditions [
30].
Not all leech species of the genus
Helobdella possess chitinous jaws and they are better known to prey on invertebrates (e.g., chironomid larvae and small freshwater snails; [
39]). Hence, tadpole predation by
Helobdella stagnalis (
Hs) leeches was somewhat surprising and has not been documented before. By contrast, such predation has been observed in other leeches (e.g.,
Desserodbella picta) that frequently devour frog tadpoles (
Lithobates sylvaticus: [
40];
Rana iberica: [
41]). Little is known about the ability of these two leech species to colonize new sites, so we can only speculate why
Hs has never been observed in the semi-natural ponds of our study area (SNPs). One possible explanation may be the relatively frequent dry up of SNPs during periods of draught. By contrast, during our 3 years of monitoring, a full dry up has never been observed in SWPs. Hence, it might be possible that frequent dry ups eliminate most leech individuals within SNPs.
Our study did not find a negative effect of sediment pollution on tadpole survival or their locomotion capacity (swim speed). Metabolic rates of toadlets (Gosner 46) also did not differ between conditions (“
control” vs. “
SWP sediment”). Since pollutant concentrations in the SWP sediment conditions were in some cases considerably higher than previously reported threshold effect concentrations in sediment for amphibians ([
38];
Table 1), this is surprising. In the context of the suitability of SWPs for amphibian habitat, pollution within SWPs is typically a major concern [
12,
14]. Previous studies found negative effects of Cu and Ni (see
Table 1) on larval survival in other amphibian species (
Lithobates pipiens; [
20],
Limnodynastes tasmaniensis; [
14]). However, since our tadpoles died quickly during the conditions that included the presence of leeches (conditions 2 and 4), our scope for comparisons was somewhat limited. Concerning our experiments that investigated swim speed and metabolic rate, we could only compare results between condition 3 (“
SWP sediment”) and the control condition. The combination of sediment pollution and leech presence (condition 4) led to a much greater predation rate on tadpoles (i.e., no tadpole survivors after 7 days), which might indicate some synergistic effects. Different mechanisms might explain the effects observed with condition 4. For example, (1) pollution might increase the motivation of leeches to feed (or increase their attraction for fatter tadpoles) or (2) pollution might affect the ability of tadpoles to escape from leeches, albeit not via changes in absolute swim speed.
As originally expected, tadpoles exposed to the SWP sediment grew faster and to a larger size than tadpoles exposed to the “clean sediment” (
Figure 3b). This is likely explained by the greater amount of organic matter present within the SWP sediment (plant matter, etc.) when compared with the aquarium sediment, providing better foraging conditions. This might have also offset any potentially negative effects of pollutants on swim speed, while it should have further increased tadpole metabolic rate, which it did not. The lacking difference in metabolic rate between conditions might also be explained, at least in part, by the time lag of up to one week between the removal of toadlets from their experimental condition and metabolic rate measurements. In contrast to our study, previous investigations demonstrated a clear effect of pollutants on metabolic rate (i.e., an increase) and swim behavior (i.e., a reduced escape capacity) in a number of species (
Lithobates catesbeianus—[
42];
Limnodynastes tasmaniensis and
Limnodynastes peronii [
14];
Lithobates pipiens—[
18]). Apart from methodological issues preventing the demonstration of such an effect in our study (though pollutant levels in the above studies were lower than in the current study;
Table 2), it is possible that tadpoles of the European green toad are more resistant to the presence of pollutants than other species. For example, green toads are highly tolerant to salt pollution [
43,
44,
45]. However, Zhelev and colleagues found that green toad tadpoles were more susceptible to pollution effects than marsh frogs (
Pelophylax ridibundus; [
46]). Nevertheless, pollution might have a long-term negative effect on adult reproduction via bioaccumulation [
47]. In this context, it would be very interesting to study pollutant levels in the different tissues of tadpoles (e.g., skin, muscle) in relation to their diet (i.e., stomach content) and to further study pollution dynamics over several years. Moreover, a parental transfer (i.e., transfer of contaminants to offspring) might also occur during future reproductive events of these individuals [
47,
48].
Given the combined results of the current study and those of our previous investigation [
30], which both demonstrate a high mortality of tadpoles in SWPs, we cannot recommend SWPs as a suitable substitute habitat for European green toads. However, there are further aspects to consider. For example, during the last few years, we observed many toadlets emerging from several SWPs, especially from those more recently built. Clearly, after the construction of SWPs in the Bas-Rhin plain, toad populations have greatly increased in their vicinity. During some years, toadlet numbers around these newly built SWPs were the highest recorded within the Bas-Rhin plain [
29]. Hence, we propose the following scenario concerning SWPs: when a SWP is first constructed, a sediment layer is absent and leeches will not be present. The shallowness at the pond edge and the absence of vegetation makes these artificial ponds the ideal breeding habitat for European green toads, whose numbers rapidly increase. However, after several years without sediment removal (dredging) and the absence of complete dry-up events, leeches are able to colonize the ponds, spread rapidly, and feed strongly on tadpoles, decimating the breeding success of toads. Further studies are needed to substantiate support for such a scenario, especially investigating the effects of SWP age and dredging frequency on toad reproduction (Clevenot et al. in prep). Until then, we recommend road managers to regularly dredge SWPs to limit the presence of leeches and to remove polluted sediment.
5. Conclusions
Clearly, recreating natural wetlands, especially in altered landscapes that have experienced a dramatic loss of wetlands, as is the case for the majority of the industrialized and agricultural plains of western Europe, would be the best way to improve the situation of locally endangered amphibians. However, since this is unlikely to happen soon, it would be a strategic mistake to neglect available artificial areas (such as SWPs) in such an effort, especially since such areas can be improved to provide a more adequate habitat for amphibians. The present study confirms that tadpole survival of European Green toads within SWPs might be low if SWPs are not adequately managed. By contrast, if well managed, these structures might not act as ecological traps for sensitive amphibians, but add to conservation measures. Hence, it is important to reconsider the use of SWPs (excluding wildlife from it or adapting it to the needs of wildlife), study their role for different species, and manage them in a way that significantly improves their qualities as a valuable substitute for declining wetland areas.







{kind=link}
{kind=link}
{kind=link}
{kind=link}