
Being Safe, but Not Too Safe: A Nudibranch Feeding on a Bryozoan-Associated Hydrozoan
by
 , , , , and
, , , , and
Davide Maggioni
1,2,* ,
,
Giulia Furfaro
3,*,
Michele Solca
4,
Davide Seveso
1,2,
Paolo Galli
1,2,5 and
and
Simone Montano
1,2 1
Department of Earth and Environmental Sciences (DISAT), University of Milano-Bicocca, 20126 Milano, Italy
2
Marine Research and High Education (MaRHE) Center, University of Milano-Bicocca, Faafu Magoodhoo Island 12030, Maldives
3
Department of Biological and Environmental Sciences and Technologies (DiSTeBA), University of Salento, 73100 Lecce, Italy
4
Museo di Biologia Marina ‘Pietro Parenzan’, 73010 Lecce, Italy
5
Dubai Business School, University of Dubai, Dubai P.O. Box 14143, United Arab Emirates
*
Authors to whom correspondence should be addressed.
Diversity 2023, 15(4), 484; https://doi.org/10.3390/d15040484
Submission received: 27 February 2023
/
Revised: 21 March 2023
/
Accepted: 23 March 2023
/
Published: 24 March 2023
(This article belongs to the Special Issue Systematics and Evolution of Gastropods)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Nudibranchs have a mostly carnivorous diet, and they prey on a wide variety of other animal taxa. Many species, mainly belonging to the Cladobranchia suborder, feed on cnidarians, including member of the class Hydrozoa. Several hydrozoan species display a symbiotic lifestyle, being associated with other benthic invertebrates, including for instance bryozoans, corals, octocorals, and sponges. In our knowledge, no record of nudibranch predation on symbiotic hydrozoans has been reported so far, possibly thanks to the protective action by the host towards its symbiotic hydrozoan. Here, we show the unexpected case of a nudibranch belonging to the recently described species Sakuraeolis marhe (Fernández-Simón and Moles, 2023) feeding on Zanclea sp. 2, a hydrozoan associated with the cheilostome bryozoan Celleporaria sp. This trophic association is confirmed by the presence and storage of the nematocysts into the nudibranch cnidosacs. Moreover, the nudibranch appears to selectively store mostly a single type of nematocyst, that is large size stenotele. The observation here reported represents the first well-documented record of a nudibranch feeding on a symbiotic hydrozoan and the first confirmed case of predation on Zanclea polyps. Moreover, we provide additional genetic information and the first description of the internal anatomy of S. marhe.
1. Introduction
Nudibranchs have a highly diversified, mostly carnivorous diet, preying on a wide range of animal taxa, including ascidians, bryozoans, cnidarians, crustaceans, echinoderms, entoprocts, fishes, other mollusks, polychaetes, and sponges [1,2]. However, most of the species are stenophagous and show a highly specialized diet, being able to feed exclusively on a single or a reduced number of benthic species [3,4,5,6]. Recently, a strong phylogenetic correlation with prey type has been demonstrated for one of the two nudibranch suborders, namely the Cladobranchia, revealing that most of the larger clades in the group feed on specific organisms [7]. In the Cladobranchia suborder, cnidarians are the most common prey items, especially those belonging to the classes Hydrozoa and Anthozoa [8], and the most recent common ancestor of the group has also been hypothesized to prey on them (and specifically on hydrozoans), with only a few subsequent dietary shifts [7]. Among cnidarian-eating cladobranchs, some taxa show a peculiar strategy, that is the sequestration of unfired nematocysts from their prey, made possible by multiple adaptations including behavior, cuticular epithelia, intracellular cuticular structures, and mucus secretions. Sequestered nematocysts, called kleptocnidae, are stored in specific structures in the upper portion of the cerata, the cnidosacs, where they remain functional [9]. There, they may play different roles, including attacking prey items during feeding activities, or they may be stored to prevent damage to the nudibranch itself, even though different studies have shown that the kleptocnidae seem to be most effective for defensive purposes [9]. Some species also appear to selectively store specific types of nematocysts among those available from the prey [10], possibly those that are most effective against predation [11].
Hydrozoan-feeding nudibranchs prey on a variety of species, mostly with a large or erect colony, belonging to the Leptothecata and Anthoathecata orders, and they may be both generalist and specialist in their preferred preys [8,12,13]. Nudibranchs generally feed on the benthic colonial polyp stage, even though some species are known to feed on planktonic polyp colonies belonging to the family Porpitidae Goldfuss, 1818 and the order Siphonophorae [13,14], or even on the medusa stage, such as the curious case of Phylliroe bucephala Lamarck, 1816 parasitizing and preying the medusa of Zanclea costata Gegenbaur, 1857 [15]. Recently, a peculiar trophic interaction between a nudibranch and a hydroid, termed kleptopredation, has also been characterized, where Cratena peregrina (Gmelin, 1791) selectively feeds on polyps of Eudendrium racemosum (Cavolini, 1785) that have recently captured planktonic preys, thus increasing the energy content of its meal [16].
Hydrozoan polyps are generally considered as substrate generalists, but several cases of symbioses with other benthic organisms are known [17], especially in the anthoathecates, with the hydrozoan colonies using the host as a substrate. An example is represented by the capitate family Zancleidae Russel, 1953, whose members associate with multiple organisms, including algae, bivalves, bryozoans, and corals [18,19,20]. Some of these associations have been characterized as mutualistic, with the hydrozoan polyps enhancing the competitive abilities of the host [21,22] and providing protection against predators, including platyhelminthes, nudibranchs, and other gastropods [21,23]. Interestingly, no report of nudibranch predation on these symbiotic hydrozoans has been reported so far. The only record of a nudibranch feeding on zancleid polyps involves the sea slug Pteraeolidia ianthina (Angas, 1864) and an undetermined Zanclea species [8], but, except the names of the two organisms involved, no additional information was reported by the authors, including for instance whether the Zanclea polyps were associated to other organisms or not. Rao and Alagarswami [24] also hypothesized that the nudibranch Anteaeolidiella orientalis (Bergh, 1888) may feed on Zanclea polyps, but they did not observe the interaction and based their assumption solely on the co-occurrence of the two organisms in the same aquarium, leaving this trophic interaction doubtful.
Recently, Cunha et al. [25] performed an extensive photographic and DNA barcoding survey of gastropods in the Central Maldives, and described the species Sakuraeolis marhe Fernández-Simón and Moles, 2023, a species currently known only from the Maldives and the Philippines, reporting that it was observed living and feeding on top of encrusting bryozoans associated with hydrozoans. This species belongs to the polyphyletic family Facelinidae Bergh, 1889, and to the genus Sakuraeolis Baba, 1965, which currently includes nine species, all of them distributed along the Indian and the west Pacific Ocean. Sakuraeolis species are known to feed on a variety of hydrozoan taxa, including the genera Ectopleura Agassiz, 1862, Eudendrium Ehrenberg, 1834, Pennaria Goldfuss, 1820, Rhizophysa Péron and Lesueur, 1807, and Solanderia Duchassaing and Michelin, 1846 [26,27,28,29], and on scyphozoan polyps of the genus Aurelia Lamarck, 1816 [30]. Sakuraeolis marhe could therefore represent the first clear case of nudibranch predation on symbiotic hydrozoans or bryozoans.
With this work, we provide new insights in both the recently described species S. marhe and its trophic interaction with the bryozoan and hydrozoan. Specifically, we provide a detailed description of the association between S. marhe and the bryozoan–hydrozoan symbiotic system, and we contribute additional genetic data of the nudibranch, including 16S rRNA and H3 sequences, assessing its phylogenetic position, we analyze its internal anatomy (buccal apparatus and reproductive system), complementing the original description of the species, and we show its ability to selectively store specific nematocyst types as kleptocnidae in the cnidosacs.
2. Materials and Methods
During surveys of the shallow reefs around Faafu Magoodhoo Island, Republic of the Maldives (3.087 N, 72.956 E), performed via SCUBA diving in March 2018, two nudibranchs were observed on a hydrozoan-associated bryozoan colony. A fragment of the bryozoan colony, together with associated hydrozoans and preying nudibranchs, was collected and brought to the facilities of the Marine Research and High Education (MaRHE) Center in Faafu Magoodhoo for further analyses. The interaction among the three groups of living organisms was observed under a Leica EZ4 D stereo microscope (Leica, Wetzlar, Germany) equipped with a Canon PowerShot G7 X Mark II camera (Canon, Tokyo, Japan). Subsequently, the organisms were anesthetized with menthol crystals and fixed in 10% formalin and 99% ethanol for morphological and molecular analyses, respectively. Fixed organisms were identified morphologically (all organisms) and genetically (hydrozoans and nudibranchs) by investigating the anatomical diagnostic characteristics, and by sequencing portions of informative DNA regions, namely the 16S rRNA and the H3 genes. An anatomical dissection of the nudibranch was carried out under a Nikon SMZ800N stereo microscope (Nikon, Tokyo, Japan) equipped with a Nikon Digital Sight 1000 camera (Nikon). The buccal apparatus was extracted using proteinase K solution (20 mg/mL) [31] for the digestion of the soft tissues and the observation of chitinous structures important for species identification, such as the radula and jaws. The reproductive system was also investigated, and a final schematic and descriptive drawing was reported. The bryozoan and associated hydrozoan species were studied under a Leica EZ4 D stereo microscope to identify them to the lowest level possible. Finally, hydrozoans and nudibranch cerata were analyzed under a Zeiss Axioskop 40 transmitted light compound microscope (Zeiss, Oberkochen, Germany) equipped with a Canon PowerShot G7 X Mark II camera (Canon) at a magnification of 40× and 100×, to identify and measure the nematocyst types. Nematocysts were measured using ImageJ 1.52p software [32], and were identified according to Östman [33].
DNA was extracted from one nudibranch individual using the QIAGEN DNeasy Blood & Tissue Kit, following the manufacturer’s protocols, and from one hydrozoan polyp following the protocol described in Maggioni et al. [20], that is digesting the specimen in proteinase K and MilliQ water for 30 min at 50 °C, and for 10 min at 90 °C. A portion of the 16S rRNA was amplified for the nudibranch and hydrozoan, using the primer pair 16AR and 16BR described in Palumbi et al. [34], and the primer pair SHB and SHA described in Cunningham and Buss [35], respectively. Additionally, a portion of the H3 gene was also amplified for the nudibranch using the primer pair H3F and H3R, described in Colgan et al. [36]. All PCRs were performed following the protocols described in Maggioni et al. [37]: 5 min for the initial denaturation at 94 °C, 30 cycles of 30 s at 94 °C, 1 min at 50 °C (SHB–SHA), 53 °C (16AR–16BR), or 52 °C (H3F–H3R), 1.5 min at 72 °C, and a final elongation of 5 min at 72 °C. All PCR products were checked through electrophoretic runs in 1.5% agarose gels, purified with Illustra ExoStar (GE Healthcare, Chicago, IL, USA), and finally sequenced in both directions with an ABI 3730xl DNA Analyzer (Applied Biosystems, Waltham, MA, USA) at the Macrogen Europe Milan Genome Center (Milan, Italy). The obtained chromatograms were checked and corrected with Geneious 6.1.6, and the protein-coding gene H3 was translated to check for the presence of open reading frames. The sequences obtained in this study were deposited with the relative metadata in GenBank, with the accession numbers OQ110615, OQ145162, and OQ145163. Sequences were checked to avoid possible contamination using the Basic Local Alignment Search Tool (BLAST), and they were compared to those already present in GenBank. Moreover, for the nudibranch, related 16S sequences belonging to the family Facelinidae and the outgroup Duvaucelia odhneri J. Tardy, 1963 (GenBank accession number: OQ145163) were downloaded from GenBank and aligned to our sequence to assess its phylogenetic position. For comparison, a H3 dataset was also assembled and analyzed in the same way, even though this DNA region is known to be weakly informative at the species and genus level [38,39,40]. Sequences were aligned with MAFFT 7.110 [41] with the E-INS-i option; the substitution models were determined using jModelTest 2 [42] under the Akaike Information Criterion, and maximum likelihood phylogenetic analyses were performed with RAxML 8.2.12 [43] with 1000 non-parametric bootstrap replicates.
3. Results
3.1. Morphological Characterization of the Organisms
The bryozoan and associated hydrozoan species were identified as Celleporaria sp. and Zanclea sp. 2 (sensu Maggioni et al. [44]), respectively, following Maggioni et al. [19,44]. Specifically, the hydrozoan colony was characterized by a naked hydrorhiza partially embedded into the bryozoan skeleton, and by monomorphic, cylindrical, and transparent gastrozooids, with short oral and aboral tentacles with inclusions in the capitations. The cnidome was composed of stenoteles of three size classes (small size: ~7 × 5 µm, medium size: ~15 × 12 µm, and large size: ~20 × 16 µm) abundant in the tentacle capitations, and also present in lower quantity in the hydrorhiza, and of macrobasic holotrichous euryteles (~20 × 12 µm) that were very abundant in the hydrorhiza. The encrusting bryozoan colony displayed a typical Celleporaria morphology, also showing a relatively large size, conspicuous pigmentation, and secondary calcification, but it was not possible to identify the specimen at the species level.
The morphological analysis of the nudibranch revealed that the external morphology and its internal anatomy perfectly matched with those reported for the genus Sakuraeolis (Figure 1 and Figure 2), with the general shape of the radula and jaws resembling that described for S. enosimensis (Baba, 1930), the type species of the genus. In particular, it shares a muscular, tongue-shaped penis without an apical hook, the majority of the liver branches in the form of horseshoes, the anus cleioproctic, the rhinophores smooth, the foot-corners tentaculiform, the jaw-plates indented dorsally, and a central radula tooth broadly horseshoe-shaped with several rather strong lateral denticles (Figure 2). Moreover, the external morphology and coloration of our specimens also matched with that of S. marhe, as reported in Cunha et al. [25], whose internal anatomy has not been described yet and is provided here for the first time:
- Facelinidae Bergh, 1889,
- Sakuraeolis Baba, 1965,
- Sakuraeolis marhe Fernández-Simón and Moles, 2023.
Buccal apparatus: The jaw-plates are horny yellow and show a dorsal indentation (Figure 2c,d). The jaw-edge bears a series of 35–40 well-defined and rounded regular denticles (Figure 2d). The radula (Figure 2e–i) has the formula 21 × 0.1.0. The teeth are broadly horseshoe shaped, bearing 7–9 rather strong denticles on either side of a produced median cusp. The teeth show a degree of intraindividual variability; in fact, the number of lateral denticles of the rachidian teeth may vary between 6 and 10, according to the position of the teeth throughout the radula (new or old teeth), and the median cusp can be differently pronounced according to this latter factor (Figure 2g–i).
Reproductive system: the penis is muscular, tongue-shaped, and unarmed. It protrudes from a large sheath visible in fresh and conserved specimens (Figure 2a,b). The prostatic portion is globose and glandular, and is connected to the vas deferens that is folded and that terminates in the ampulla. This is rounded and connected with a sac-shaped bursa copulatrix, and with a small tubular vagina (Figure 2b). A voluminous folded female gland mass is located ventrally between the female part of the reproductive system, and the ampulla and the ovotestis (Figure 2b).
3.2. Molecular Phylogenetics
The obtained sequences showed no contamination and further confirmed the morphological identifications. The hydrozoan 16S sequence (GenBank accession number: OQ145162) perfectly matched the deposited sequences of Zanclea sp. 2 from the Maldives (percentage identity of up to 100%). The nudibranch 16S sequence (GenBank accession number: OQ145163) showed a percentage identity of 95.65% with a sequence ascribed to a Facelina sp. (GenBank accession number: MK478697) collected in Indonesia [45], and it resulted 93.44% identical with one S. enosimensis sequence (GenBank accession number: HM162682) from California. Similarly, the latter specimen (GenBank accession number: HQ010472) showed a percentage identity of 95.27% with the obtained nudibranch H3 sequence (GenBank accession number: OQ110615), whereas the highest match was with Caloria sp. 4 (96.96%; GenBank accession number: JQ996965) from the Philippines. The 16S phylogenetic hypothesis revealed a topology that was broadly concordant with the COX1 tree produced by Cunha et al. [25] (Figure 3). The Sakuraeolis marhe sequence was closely related to the Facelina sp. sequence from Indonesia, followed by two Favorinidae sp. sequences from the Philippines. This group was then closely related to Sakuraeolis arcana Ellis-Diamond, Picton, Tibiriçá & Sigwart, 2021 from Mozambique, S. enosimensis from California, S. japonica (Baba, 1937) from Korea, and Caloria sp. 4 from the Philippines. Finally, this clade was sister to a clade composed of the Cratena Bergh, 1864 and Myja Bergh, 1896 sequences, together with two Facelinidae sp. 2 sequences, again similarly to the data from Cunha et al. [25]. Overall, the nodes were moderately to well-supported, especially regarding the relationships of S. marhe with Facelina sp. and Favorinidae sp., similarly to what was shown in Cunha et al. [25]. The H3 phylogenetic tree (Figure S1) was characterized by a lower resolution, similar to what was shown in previous studies [38,39,40], and the relationships between the S. marhe sequence and other facelinid sequences remained unclear, according to this DNA region. Overall, the results obtained from both morphological and molecular analyses confirmed that the nudibranch belongs to the species S. marhe, the bryozoan to Celleporaria sp., and the hydrozoan to Zanclea sp. 2.
3.3. Behavioral Observations and Kleptocnidae Assessment
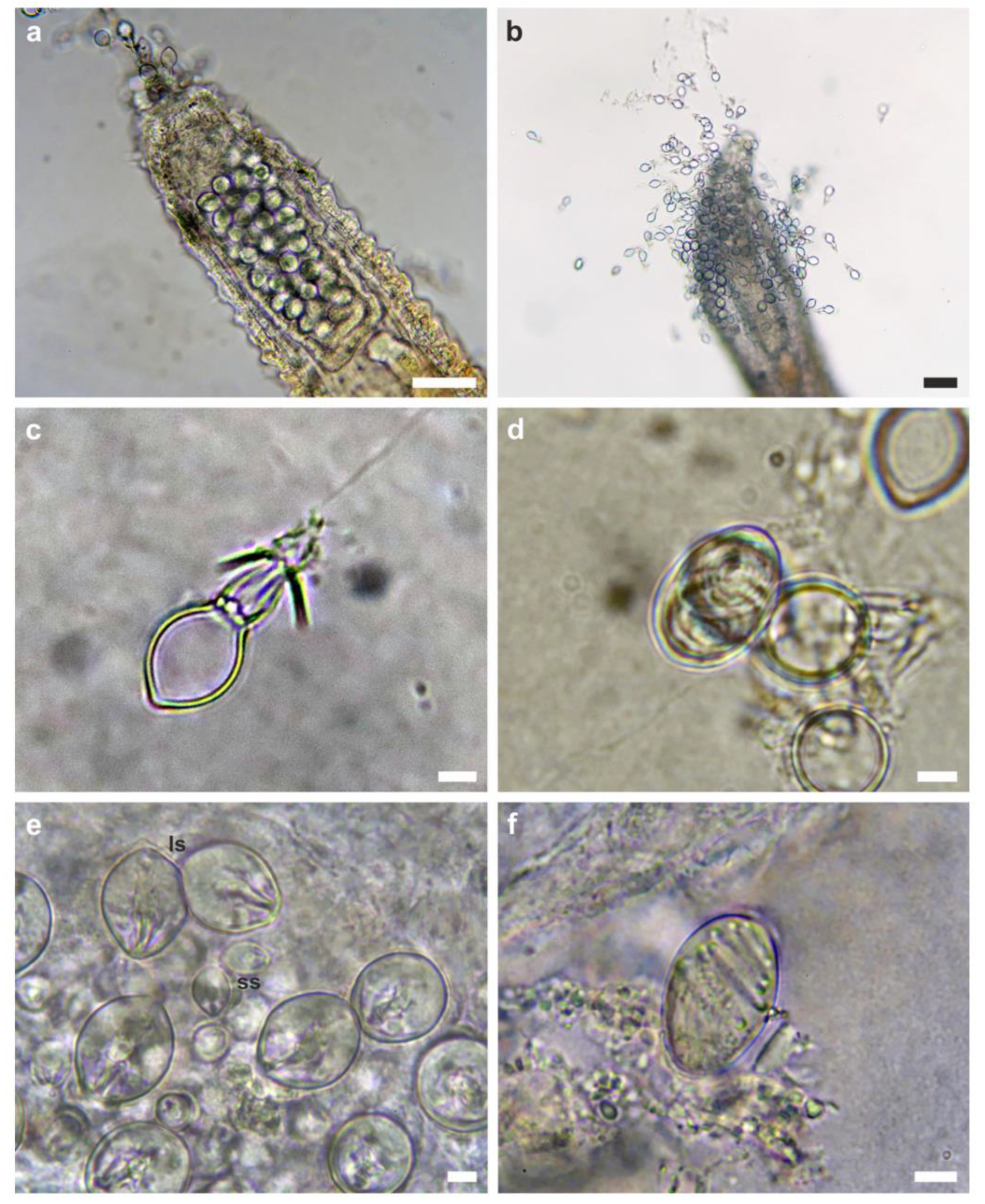
The two S. marhe individuals were observed moving on top of the bryozoan colony and feeding specifically on Zanclea polyps (Figure 1a–c), but not on Celleporaria zooids, which were observed to be intact after the passage of the nudibranchs. Specifically, the nudibranchs were observed crawling on the Celleporaria surface, leaving behind consumed or partially consumed hydrozoan polyps (Figure 1d). The cerata were filled with kleptocnidae (Figure 1e and Figure 4a,b) that were identified as large size stenoteles (Figure 4c) and macrobasic holotrichous euryteles (Figure 4d), the latter being very rare. The analysis of the hydrozoan cnidome revealed the presence of stenoteles (Figure 4e) and macrobasic holotrichous euryteles (Figure 4f), in agreement with Maggioni et al. [19,44]. Only large size stenoteles (~20 × 16 µm) were observed in the cnidosacs, whereas Zanclea sp. 2 polyps also possessed small and medium size stenoteles, and euryteles were rare, despite being common in the Zanclea hydrorhiza, as shown in this work and in Maggioni et al. [44].
4. Discussion
Maldivian coral reefs are home to an extraordinary diversity of organisms, with new species being frequently described from different areas of the archipelago [46,47,48,49]. In addition, previously unknown or understudied inter-specific associations and interactions have been recently described [50,51,52], such as the case of an amphipod associated with the gastropod Coriocella hybiae Wellens, 1991 [53], revealing that the diversity and ecology of these ecosystems strongly need further research, especially given the current coral reef crisis [54].
In this context, Sakuraeolis marhe was recently described from Central Maldives by Cunha et al. [25], based on the external morphology and genetic data. The specimens herein analyzed matched the morphological description of this species, and their identification was further supported by the newly produced genetic data, the overlapping distribution, and the substrate on which they were found. Additionally, the characterization of the internal anatomical diagnostic features (i.e., the buccal apparatus and the reproductive system) complemented the original description of the species. The anatomical description here provided is congruent with the characters considered diagnostic for the genus Sakuraeolis, except for the features of the penial sheath that, however, are also lacking in the newly described congeneric S. arcana [29], suggesting that this structure may not be diagnostic at the genus level. This morphological observation sheds some lights on the gap of knowledge still existing on the synapomorphies characterizing this apparently non-monophyletic genus, for which an integrative systematic assessment is indeed desirable. With this work, we also provided ecological information on the species, regarding the substrate and the feeding preferences. Indeed, S. marhe was observed living on the bryozoan Celleporaria sp. and selectively preying on its associated symbiotic hydrozoan Zanclea sp. 2.
Celleporaria species are known to host different hydrozoan species in the Indo-Pacific, including Zanclea divergens Boero, Bouillon & Gravili, 2000, Zanclea sp. 1, and Zanclea sp. 2 [20,37,55,56], and these associations are supposed to be intimate, with the hydrozoans having lost the perisarc covering the hydrorhiza, and being protected by the bryozoan skeleton through secondary calcification. Advantages for bryozoan-associated hydrozoan colonies may be increased food availability, thanks to water currents produced by the host, and protection from predators, thanks to the bryozoan skeleton [17]. On the other hand, Celleporaria zooids were observed stimulating the release of mucus particles from hydrozoan polyps, which were then ingested [56]. Osman and Haugsness [21] reported on a Celleporaria-associated Zanclea species that enhanced the competitive abilities of the host in the Eastern Pacific, by protecting it from competitors and predators thanks to nematocysts. Even though the latter Zanclea species remains undetermined, it looks like Celleporaria and all associated hydroids live in intimate and possibly mutualistic associations with positive trophic and defensive outcomes for both symbiotic partners. A similar situation was also hypothesized for coral-associated Zanclea species [23], where hydroids might repel predators, including corallivore gastropods and pathogens, thanks to their nematocysts.
The predatory behavior of S. marhe here reported was therefore unexpected, revealing that the beneficial roles of symbiotic hydrozoans seem not to apply when the predators are specialized hydrozoan eaters. Even if the nudibranch does not directly feed on the bryozoan, it may affect the structural integrity of zooids by crawling on the surface, and may reduce its competitive ability by removing the associated hydrozoans. This questions the defensive function of symbiotic hydrozoans, at least in the Zanclea–Celleporaria association here examined. At the same time, the possible protective function of the bryozoan skeleton towards associated hydrozoans is also challenged by the observations here reported, at least for what concerns S. marhe predatory activities.
The analysis of the cnidosacs content confirmed the presence of kleptocnidae previously belonging to Zanclea sp. 2 polyps, namely, large size stenoteles, and very rarely, euryteles, both with sizes concordant with those of Zanclea sp. 2 nematocysts. The modulation of kleptocnidae storage has been demonstrated in a few cases, being, for instance, governed by both the diet and the presence of predators in Flabellina verrucosa (M. Sars, 1829) [57]. The uptake of specific types of nematocysts among those available from the prey has also been shown in other nudibranch species such as Cratena pilata (Gould, 1870), storing only mastigophores from the prey Pennaria disticha Goldfuss, 1820 [58]. The occurrence of kleptocnidae mainly of the stenotele type in S. marhe may suggest that this species is selective in the types of nematocysts that it stores in the cnidosacs or in the hydrozoan structures that it feeds on. Euryteles are concentrated in the hydrorhiza of Zanclea sp. 2 and are not found in polyps, suggesting that S. marhe may selectively feed on the polyps, possibly avoiding hydrorhizal structures. On the other hand, small and medium size stenoteles were not observed in the cnidosacs, and these nematocysts are very common in polyps, indicating that S. marhe selectively stores large size stenoteles in its cnidosacs and leaving open the hypothesis that the nudibranch feeds on the hydrorhiza as well, but does not store euryteles.
In conclusion, the morphological and anatomical descriptions of S. marhe were provided here for the first time, together with useful insights on the diagnostic power of the reproductive structures that are nowadays considered useful to delimit nudibranchs at the genus taxonomic level. Finally, the interaction here described represents the first well-documented record of a nudibranch feeding on a symbiotic hydrozoan and storing the nematocysts in its cnidosacs. This also represents the first record of nudibranch predation on Zanclea polyps, highlighting the need for further research to better characterize and comprehend the roles of these hydrozoans in the many symbiotic systems in which they are involved.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15040484/s1, Figure S1: Maximum likelihood H3 phylogenetic reconstruction of Sakuraeolis marhe.
Author Contributions
Conceptualization, D.M.; methodology, D.M. and G.F.; formal analysis, D.M. and G.F.; writing—original draft preparation, D.M. and G.F.; writing—review and editing, D.M., G.F., M.S., D.S., P.G., and S.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The DNA sequences obtained in this study are openly available in GenBank with the accession numbers OQ110615, OQ145162, and OQ145163.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McDonald, G.; Nybakken, J. A preliminary report on a world-wide review of the food of nudibranchs. J. Molluscan Stud. 1991, 57, 61–663. [Google Scholar] [CrossRef]
- Canessa, M.; Bavestrello, G.; Cattaneo-Vietti, R.; Furfaro, G.; Doneddu, M.; Navone, A.; Trainito, E. Rocky substrate affects benthic heterobranch assemblages and prey/predator relationships. Estuar. Coast. Shelf Sci. 2021, 261, 31. [Google Scholar] [CrossRef]
- Baretta-Bekker, H.J.; Duursma, E.K.; Kuipers, B.R. Encyclopedia of Marine Sciences; Springer: Berlin, Germany, 1998; ISBN 978-3-642-58831-0. [Google Scholar]
- Prkic, J.; Furfaro, G.; Mariottini, P.; Carmona, L.; Cervera, J.L.; Modica, M.V.; Oliverio, M. First record of Calma gobioophaga Calado and Urgorri, 2002 (Gastropoda: Nudibranchia) in the Mediterranean Sea. Mediterr. Mar. Sci. 2014, 15, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Furfaro, G.; Trainito, E.; De Lorenzi, F.; Fantin, M.; Doneddu, M. Tritonia nilsodhneri marcus Ev., 1983 (gastropoda, Heterobranchia, tritoniidae): First records for the Adriatic Sea and new data on ecology and distribution of Mediterranean populations. Acta Adriat. 2017, 58, 261–270. [Google Scholar] [CrossRef]
- Chimienti, G.; Angeletti, L.; Furfaro, G.; Canese, S.; Taviani, M. Habitat, morphology and trophism of Tritonia callogorgiae sp. nov., a large nudibranch inhabiting Callogorgia verticillata forests in the Mediterranean Sea. Deep Sea Res. Oceanogr. Res. Pap. 2020, 165, 103364. [Google Scholar] [CrossRef]
- Goodheart, J.A.; Bazinet, A.L.; Valdés, Á.; Collins, A.G.; Cummings, M.P. Prey preference follows phylogeny: Evolutionary dietary patterns within the marine gastropod group Cladobranchia (Gastropoda: Heterobranchia: Nudibranchia). BMC Evol. Biol. 2017, 17, 221. [Google Scholar] [CrossRef] [Green Version]
- Putz, A.; König, G.M.; Wägele, H. Defensive strategies of Cladobranchia (Gastropoda, Opisthobranchia). Nat. Prod. Rep. 2010, 27, 1386–1402. [Google Scholar] [CrossRef]
- Greenwood, P.G. Acquisition and use of nematocysts by cnidarian predators. Toxicon 2009, 54, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, P.G. 1988. Nudibranch nematocysts. In The Biology of Nematocysts; Hessinger, D.A., Lenhoff, H.M., Eds.; Academic Press: San Diego, CA, USA, 1988; pp. 445–462. ISBN 978-032-314-462-9. [Google Scholar]
- Edmunds, M. Protective mechanisms in the Eolidacea (Mollusca Nudibranchia). Zool. J. Linn. Soc. 1966, 46, 27–71. [Google Scholar] [CrossRef]
- Miller, M.C. Distribution and food of the nudibranchiate Mollusca of the south of the Isle of Man. J. Anim. Ecol. 1961, 30, 95–116. [Google Scholar] [CrossRef]
- McDonald, D.G.; Nybakken, J. A worldwide review of the food of nudibranch mollusks. Part II. The suborder Dendronotacea. Veliger 1999, 42, 62–66. [Google Scholar]
- Arai, M.N. Predation on pelagic coelenterates: A review. J. Mar. Biol. Assoc. UK 2005, 85, 523–536. [Google Scholar] [CrossRef]
- Martin, R.; Brinckmann-Voss, A. Zum brutparasitismus von Phyllirhoe bucephala Per. & Les. (Gastropoda, Nudibranchia) auf der meduse Zanclea costata Gegenb. (Hydrozoa, Anthomedusae). Pubbl. Staz. Zool. Napoli 1963, 33, 206–223. [Google Scholar]
- Willis, T.J.; Berglöf, K.T.; McGill, R.A.; Musco, L.; Piraino, S.; Rumsey, C.M.; Fernández, T.V.; Badalamenti, F. Kleptopredation: A mechanism to facilitate planktivory in a benthic mollusc. Biol. Lett. 2017, 13, 20170447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puce, S.; Cerrano, C.; Di Camillo, C.G.; Bavestrello, G. Hydroidomedusae (Cnidaria: Hydrozoa) symbiotic radiation. J. Mar. Biol. Assoc. UK 2008, 88, 1715–1721. [Google Scholar] [CrossRef]
- Boero, F.; Bouillon, J.; Gravili, C. A survey of Zanclea, Halocoryne and Zanclella (Cnidaria, Hydrozoa, Anthomedusae, Zancleidae) with description of new species. Ital. J. Zool. 2000, 67, 93–124. [Google Scholar] [CrossRef]
- Maggioni, D.; Saponari, L.; Seveso, D.; Galli, P.; Schiavo, A.; Ostrovsky, A.N.; Montano, S. Green fluorescence patterns in closely related symbiotic species of Zanclea (Hydrozoa, Capitata). Diversity 2020, 12, 78. [Google Scholar] [CrossRef] [Green Version]
- Maggioni, D.; Arrigoni, R.; Seveso, D.; Galli, P.; Berumen, M.L.; Denis, V.; Hoeksema, B.W.; Huang, D.; Manca, F.; Pica, D.; et al. Evolution and biogeography of the Zanclea-Scleractinia symbiosis. Coral Reefs 2022, 41, 779–795. [Google Scholar] [CrossRef]
- Osman, R.W.; Haugsness, J.A. Mutualism among sessile invertebrates: A mediator of competition and predation. Science 1981, 211, 846–848. [Google Scholar] [CrossRef] [Green Version]
- Ristedt, H.; Schuhmacher, H. The bryozoan Rhynchozoon larreyi (Audouin, 1826)—A successful competitor in coral reef communities of the Red Sea. Mar. Ecol. 1985, 6, 167–179. [Google Scholar] [CrossRef]
- Montano, S.; Fattorini, S.; Parravicini, V.; Berumen, M.L.; Galli, P.; Maggioni, D.; Arrigoni, R.; Seveso, D.; Strona, G. Corals hosting symbiotic hydrozoans are less susceptible to predation and disease. Proc. Royal Soc. B Biol. Sci. 2017, 284, 20172405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, K.V.; Alagarswami, K. An account of the structure and early development of a new species of a nudibranchiate gastropod, Eolidina (Eolidina) mannarensis. J. Mar. Biol. Assoc. India 1960, 2, 6–16. [Google Scholar]
- Cunha, T.J.; Fernández-Simón, J.; Petrula, M.; Giribet, G.; Moles, J. Photographic Checklist, DNA Barcoding, and New Species of Sea Slugs and Snails from the Faafu Atoll, Maldives (Gastropoda: Heterobranchia and Vetigastropoda). Diversity 2023, 15, 219. [Google Scholar] [CrossRef]
- Arakawa, K.Y. Competitors and fouling organisms in the hanging culture of the Pacific oyster, Crassostrea gigas (Thunberg). Mar. Freshw. Behav. Physiol. 1990, 17, 67–94. [Google Scholar] [CrossRef]
- Hirano, Y. Two new species of Sakuraeolis (Aeolidacea, Facelinidae) from Japan. Venus 1999, 58, 191–199. [Google Scholar] [CrossRef]
- Nagale, P.; Apte, D. Intertidal hydroids (Cnidaria: Hydrozoa: Hydroidolina) from the Gulf of Kutch, Gujarat, India. Mar. Biodivers. Rec. 2014, 7, e116. [Google Scholar] [CrossRef] [Green Version]
- Ellis-Diamond, D.C.; Picton, B.E.; Tibiriçá, Y.; Sigwart, J.D. A new species of Sakuraeolis from Mozambique, described using 3D reconstruction of anatomy and phylogenetic analysis. J. Molluscan Stud. 2021, 87, eyab010. [Google Scholar] [CrossRef]
- Takao, M.; Okawachi, H.; Uye, S.I. Natural predators of polyps of Aurelia aurita sl (Cnidaria: Scyphozoa: Semaeostomeae) and their predation rates. Plankton Benthos Res. 2014, 9, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Furfaro, G.; Trainito, E.; Fantin, M.; D’Elia, M.; Madrenas, E.; Mariottini, P. Mediterranean Matters: Revision of the Family Onchidorididae (Mollusca, Nudibranchia) with the Description of a New Genus and a New Species. Diversity 2023, 15, 38. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Östman, C. A guideline to nematocyst nomenclature and classification, and some notes on the systematic value of nematocysts. Sci. Mar. 2000, 64, 31–46. [Google Scholar] [CrossRef]
- Palumbi, S.R.; Martin, A.; Romano, S.; McMillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR; Department of Zoology and Kewalo Marine Laboratory, University of Hawaii: Honolulu, HI, USA, 1991. [Google Scholar]
- Cunningham, C.W.; Buss, L.W. Molecular evidence for multiple episodes of paedomorphosis in the family Hydractiniidae. Biochem. Syst. Ecol. 1993, 21, 57–69. [Google Scholar] [CrossRef]
- Colgan, D.; McLauchlan, A.; Wilson, G.; Livingston, S.; Edgecombe, G.; Macaranas, J.; Gray, M. Histone H3 and U2 snRNA DNA sequences and arthropod molecular evolution. Aust. J. Zool. 1998, 46, 419–437. [Google Scholar] [CrossRef]
- Maggioni, D.; Schiavo, A.; Ostrovsky, A.N.; Seveso, D.; Galli, P.; Arrigoni, R.; Berumen, M.L.; Benzoni, F.; Montano, S. Cryptic species and host specificity in the bryozoan-associated hydrozoan Zanclea divergens (Hydrozoa, Zancleidae). Mol. Phylogenet. Evol. 2020, 151, 106893. [Google Scholar] [CrossRef] [PubMed]
- Galia-Camps, C.; Carmona, L.; Cabrito, A.; Ballesteros, M.B.V. Double trouble. A cryptic first record of Berghia marinae Carmona, Pola, Gosliner, & Cervera 2014 in the Mediterranean Sea. Mediterr. Mar. Sci. 2020, 21, 191–200. [Google Scholar] [CrossRef]
- Furfaro, G.; Salvi, D.; Trainito, E.; Vitale, F.; Mariottini, P. When morphology does not match phylogeny: The puzzling case of two sibling nudibranchs (Gastropoda). Zool. Scr. 2021, 50, 439–454. [Google Scholar] [CrossRef]
- Garzia, M.; Mariottini, P.; Salvi, D.; Furfaro, G. Variation and Diagnostic Power of the Internal Transcribed Spacer 2 in Mediterranean and Atlantic Eolid Nudibranchs (Mollusca, Gastropoda). Front. Mar. Sci. 2021, 8, 693093. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Maggioni, D.; Arrigoni, R.; Galli, P.; Berumen, M.L.; Seveso, D.; Montano, S. Polyphyly of the genus Zanclea and family Zancleidae (Hydrozoa, Capitata) revealed by the integrative analysis of two bryozoan-associated species. Contr. Zool. 2018, 87, 87–104. [Google Scholar] [CrossRef] [Green Version]
- Schillo, D.; Wipfler, B.; Undap, N.; Papu, A.; Boehringer, N.; Eisenbarth, J.H.; Kaligis, F.; Bara, R.; Schäberle, T.F.; König, G.M.; et al. Description of a new Moridilla species from North Sulawesi, Indonesia (Mollusca: Nudibranchia: Aeolidioidea)—Based on MicroCT, histological and molecular analyses. Zootaxa 2019, 4652, 265–295. [Google Scholar] [CrossRef] [Green Version]
- Maggioni, D.; Puce, S.; Galli, P.; Seveso, D.; Montano, S. Description of Turritopsoides marhei sp. nov.(Hydrozoa, Anthoathecata) from the Maldives and its phylogenetic position. Mar. Biol. Res. 2017, 13, 983–992. [Google Scholar] [CrossRef]
- Voigt, O.; Ruthensteiner, B.; Leiva, L.; Fradusco, B.; Woerheide, G. A new species of the calcareous sponge genus Leuclathrina (Calcarea: Calcinea: Clathrinida) from the Maldives. Zootaxa 2018, 4382, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Malinverno, E.; Leoni, B.; Galli, P. Coccolithophore assemblages and a new species of Alisphaera from the Faafu Atoll, Maldives, Indian Ocean. Mar. Micropaleontol. 2022, 172, 102110. [Google Scholar] [CrossRef]
- Tea, Y.K.; Najeeb, A.; Rowlett, J.; Rocha, L.A. Cirrhilabrus finifenmaa (Teleostei, Labridae), a new species of fairy wrasse from the Maldives, with comments on the taxonomic identity of C. rubrisquamis and C. wakanda. ZooKeys 2022, 1088, 65. [Google Scholar] [CrossRef]
- Montano, S.; Maggioni, D. Camouflage of sea spiders (Arthropoda, Pycnogonida) inhabiting Pavona varians. Coral Reefs 2018, 37, 153. [Google Scholar] [CrossRef] [Green Version]
- Montalbetti, E.; Saponari, L.; Montano, S.; Maggioni, D.; Dehnert, I.; Galli, P.; Seveso, D. New insights into the ecology and corallivory of Culcita sp. (Echinodermata: Asteroidea) in the Republic of Maldives. Hydrobiologia 2019, 827, 353–365. [Google Scholar] [CrossRef]
- Maggioni, D.; Montano, S.; Voigt, O.; Seveso, D.; Galli, P. A mesophotic hotel: The octocoral Bebryce cf. grandicalyx as a host. Ecology 2020, 101, e02950. [Google Scholar] [CrossRef]
- Maggioni, D.; Saponari, L.; Montano, S. Shrimps with a coat: An amphipod hiding in the mantle of Coriocella hibyae (Gastropoda, Velutinidae). Coral Reefs 2018, 37, 647. [Google Scholar] [CrossRef] [Green Version]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Puce, S.; Cerrano, C.; Boyer, M.; Ferretti, C.; Bavestrello, G. Zanclea (Cnidaria: Hydrozoa) species from Bunaken Marine Park (Sulawesi Sea, Indonesia). J. Mar. Biol. Assoc. UK 2002, 82, 943–954. [Google Scholar] [CrossRef]
- Puce, S.; Bavestrello, G.; Di Camillo, C.G.; Boero, F. Symbiotic relationships between hydroids and bryozoans. Symbiosis 2007, 44, 137–143. [Google Scholar]
- Frick, K. Response in nematocyst uptake by the nudibranch Flabellina verrucosa to the presence of various predators in the southern Gulf of Maine. Biol. Bull. 2003, 205, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Kepner, W.A. The manipulation of the nematocysts of Pennaria tiarella by Aeolis pilata. J. Morphol. 1943, 73, 297–311. [Google Scholar] [CrossRef]
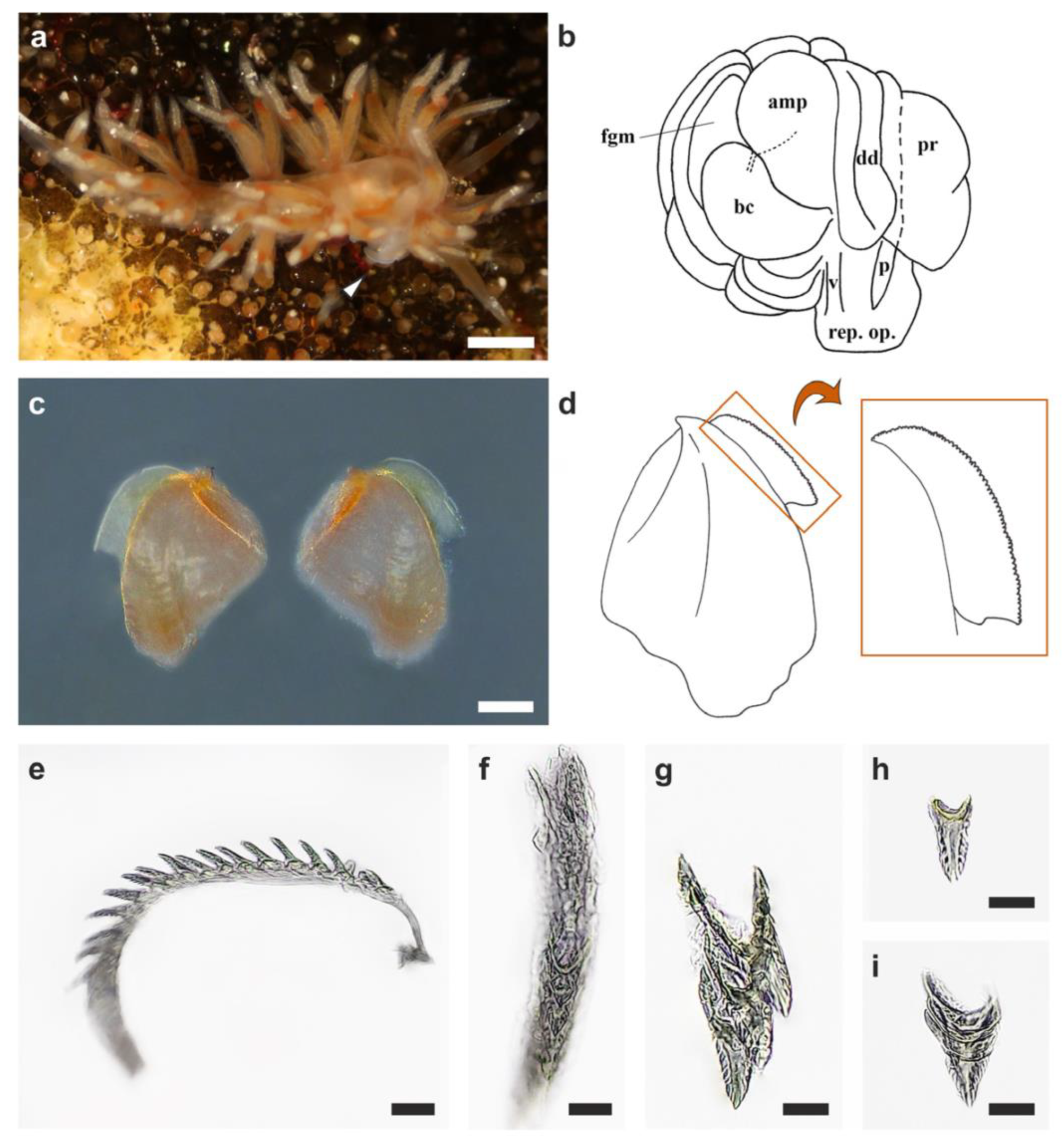
Figure 1.
Sakuraeolis marhe feeding on Zanclea sp. 2 associated with Celleporaria sp. (a,b) An individual of S. marhe crawling on Celleporaria sp. while feeding on Zanclea sp. 2. (c) A detail of the association between Celleporaria sp. and Zanclea sp. 2, showing extended bryozoan zooids (black arrowhead) and hydrozoan polyps (white arrowhead). (d) Signs of predation of S. marhe on Zanclea sp. 2 polyps (two examples of consumed polyps are indicated by black arrowheads). (e) Detail of the cerata with terminal cnidosacs (arrowhead) containing kleptocnidae. Scale bars: (a–c) 0.5 mm, (d,e) 0.2 mm.
Figure 1.
Sakuraeolis marhe feeding on Zanclea sp. 2 associated with Celleporaria sp. (a,b) An individual of S. marhe crawling on Celleporaria sp. while feeding on Zanclea sp. 2. (c) A detail of the association between Celleporaria sp. and Zanclea sp. 2, showing extended bryozoan zooids (black arrowhead) and hydrozoan polyps (white arrowhead). (d) Signs of predation of S. marhe on Zanclea sp. 2 polyps (two examples of consumed polyps are indicated by black arrowheads). (e) Detail of the cerata with terminal cnidosacs (arrowhead) containing kleptocnidae. Scale bars: (a–c) 0.5 mm, (d,e) 0.2 mm.

Figure 2.
Anatomical diagnostic characteristics of Sakuraeolis marhe. (a) In vivo picture of an S. marhe individual with everted male reproductive structures (arrowhead). (b) Schematic drawing of the reproductive system. (c) Picture of the two jaws of the buccal apparatus, and (d) schematic drawing of the jaws and the jaw-edge, drawn at a different magnification scale (orange rectangle). (e,f) Lateral and frontal view of the radula, respectively, and (g–i) intra-individual variability among the teeth. amp: ampulla, bc: bursa copulatrix, dd: vas deferens, fgm: female gland mass, p: penis, pr: prostate, rep. op.: reproductive opening, v: vagina. Scale bar: (a) 0.5 mm, (c) 0.75 mm, (e,f) 25 µm, (g–i) 10 µm.
Figure 2.
Anatomical diagnostic characteristics of Sakuraeolis marhe. (a) In vivo picture of an S. marhe individual with everted male reproductive structures (arrowhead). (b) Schematic drawing of the reproductive system. (c) Picture of the two jaws of the buccal apparatus, and (d) schematic drawing of the jaws and the jaw-edge, drawn at a different magnification scale (orange rectangle). (e,f) Lateral and frontal view of the radula, respectively, and (g–i) intra-individual variability among the teeth. amp: ampulla, bc: bursa copulatrix, dd: vas deferens, fgm: female gland mass, p: penis, pr: prostate, rep. op.: reproductive opening, v: vagina. Scale bar: (a) 0.5 mm, (c) 0.75 mm, (e,f) 25 µm, (g–i) 10 µm.

Figure 3.
Maximum likelihood 16S phylogenetic reconstruction of Sakuraeolis marhe, highlighted in orange, and closely related species. Codes before the species name are the GenBank accession numbers, whereas codes after the species names refer to the voucher numbers or sample IDs. Numbers at nodes are bootstrap values.
Figure 3.
Maximum likelihood 16S phylogenetic reconstruction of Sakuraeolis marhe, highlighted in orange, and closely related species. Codes before the species name are the GenBank accession numbers, whereas codes after the species names refer to the voucher numbers or sample IDs. Numbers at nodes are bootstrap values.

Figure 4.
Kleptocnidae and nematocysts of Sakuraeolis marhe and Zanclea sp. 2. Detail of a cnidosac with (a) undischarged and (b) discharged kleptocnidae. (c) Discharged stenotele and (d) undischarged eurytele from the cnidosac. (e) Small size stenoteles (ss) and large size stenoteles (ls), and (f) eurytele isolated from Zanclea sp. 2 polyps. Scale bars: (a,b) 50 µm; (c–f) 5 µm.
Figure 4.
Kleptocnidae and nematocysts of Sakuraeolis marhe and Zanclea sp. 2. Detail of a cnidosac with (a) undischarged and (b) discharged kleptocnidae. (c) Discharged stenotele and (d) undischarged eurytele from the cnidosac. (e) Small size stenoteles (ss) and large size stenoteles (ls), and (f) eurytele isolated from Zanclea sp. 2 polyps. Scale bars: (a,b) 50 µm; (c–f) 5 µm.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Maggioni, D.; Furfaro, G.; Solca, M.; Seveso, D.; Galli, P.; Montano, S. Being Safe, but Not Too Safe: A Nudibranch Feeding on a Bryozoan-Associated Hydrozoan. Diversity 2023, 15, 484. https://doi.org/10.3390/d15040484
AMA Style
Maggioni D, Furfaro G, Solca M, Seveso D, Galli P, Montano S. Being Safe, but Not Too Safe: A Nudibranch Feeding on a Bryozoan-Associated Hydrozoan. Diversity. 2023; 15(4):484. https://doi.org/10.3390/d15040484
Chicago/Turabian StyleMaggioni, Davide, Giulia Furfaro, Michele Solca, Davide Seveso, Paolo Galli, and Simone Montano. 2023. "Being Safe, but Not Too Safe: A Nudibranch Feeding on a Bryozoan-Associated Hydrozoan" Diversity 15, no. 4: 484. https://doi.org/10.3390/d15040484
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.