

Space Use and Movements of Southeastern Breeding Double-Crested Cormorants (Nannopterum auritum) in the United States

Abstract
:1. Introduction
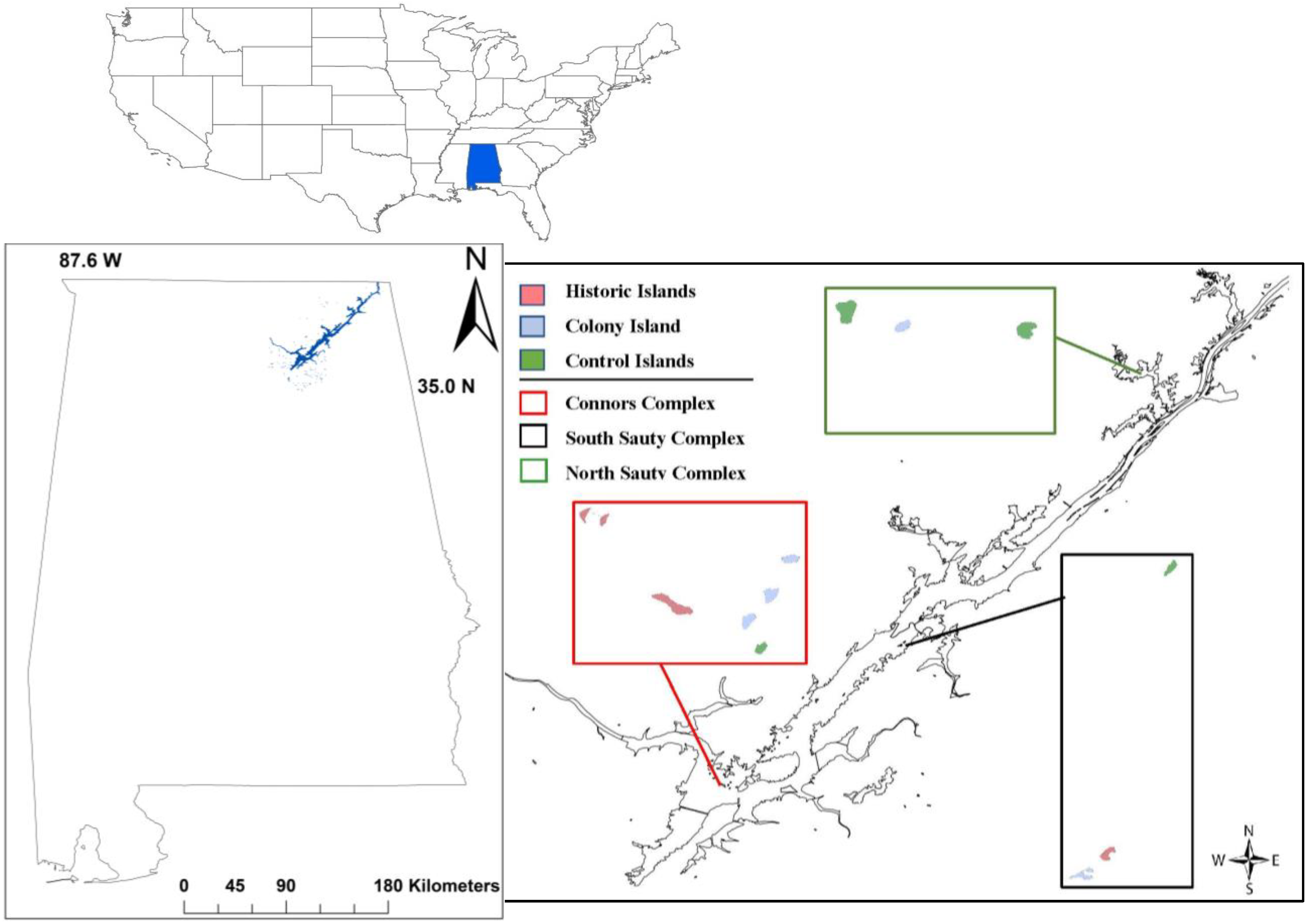
2. Materials and Methods
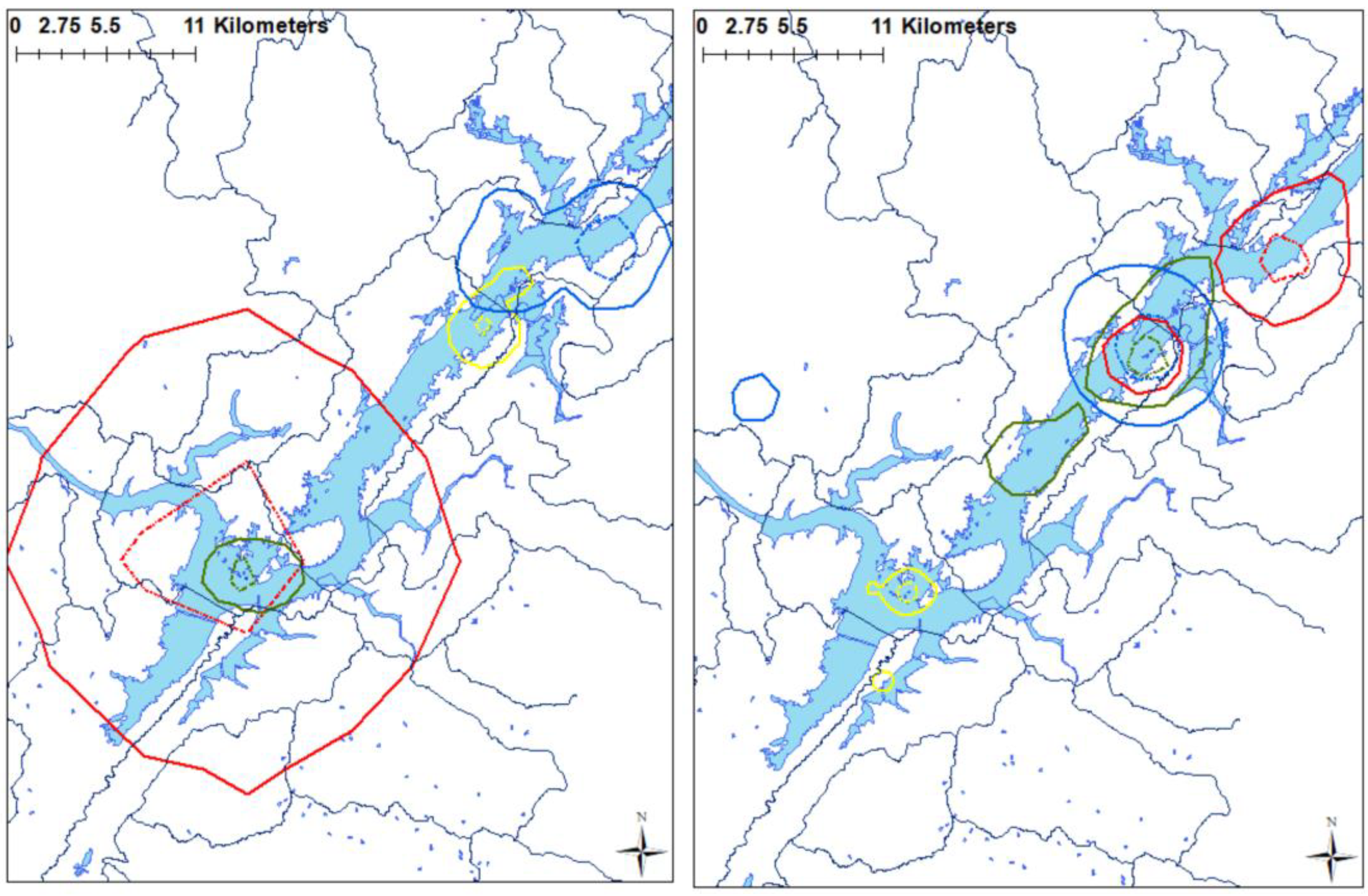
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wires, L.R.; Cuthbert, F.J. Historic Populations of the Double-crested Cormorant (Phalacrocorax auritus): Implications for Conservation and Management in the 21st Century. Waterbirds 2006, 29, 9–37. [Google Scholar] [CrossRef] [Green Version]
- Dorr, B.; King, D.T.; Tobin, M.E.; Harrel, J.B.; Smith, P.L. Double-crested Cormorant movements in relation to aquaculture in eastern Mississippi and western Alabama. Waterbirds 2004, 27, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Dorr, B.S.; Fielder, D.G. Double-Crested Cormorants: Too Much of a Good Thing? Fisheries 2017, 42, 468–477. [Google Scholar] [CrossRef] [Green Version]
- Barras, S.C. Double-Crested Cormorants in Alabama; USDA National Wildlife Research Center-Staff Publications: Washington, DC, USA, 2004; p. 77.
- Lafferty, D.J.; Hanson-Dorr, K.C.; Prisock, A.M.; Dorr, B.S. Biotic and abiotic impacts of Double-crested cormorant breeding colonies on forested islands in the southeastern United States. For. Ecol. Manag. 2016, 369, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Guillaumet, A.; Dorr, B.; Wang, G.; Taylor, J.D.; Chipman, R.B.; Scherr, H.; Bowman, J.; Abraham, K.F.; Doyle, T.J.; Cranker, E. Determinants of local and migratory movements of Great Lakes double-crested cormorants. Behav. Ecol. 2011, 22, 1096–1103. [Google Scholar] [CrossRef] [Green Version]
- Dorr, B.S.; Hatch, J.J.; Weseloh, D.V. Double-crested Cormorant (Phalacrocorax auritus), version 2.0. In Birds of North America; Cornell Lab of Ornithology: Ithaca, NY, USA, 2014. [Google Scholar]
- Coleman, J.T.; Richmond, M.E.; Rudstam, L.G.; Mattison, P.M. Foraging location and site fidelity of the Double-crested Cormorant on Oneida Lake, New York. Waterbirds 2005, 28, 498–510. [Google Scholar] [CrossRef]
- Wires, L.R.; Cuthbert, F.J. Characteristics of Double-crested Cormorant colonies in the U.S. Great Lakes island landscape. J. Great Lakes Res. 2010, 36, 232–241. [Google Scholar] [CrossRef]
- Stapanian, M.A. Interspecific Interactions, Habitat Use, and Management of Double-crested Cormorants (Phalacrocorax auritus) in the Laurentian Great Lakes: An Introduction. J. Great Lakes Res. 2002, 28, 119–124. [Google Scholar] [CrossRef]
- Coleman, J.T.; Richmond, M.E. Daily foraging patterns of adult Double-crested Cormorants during the breeding season. Waterbirds 2007, 30, 189–198. [Google Scholar] [CrossRef]
- Glahn, J.F.; Reinhold, D.S.; Sloan, C.A. Recent population trends of Double-crested Cormorants wintering in the delta region of Mississippi: Responses to roost dispersal and removal under a recent depredation order. Waterbirds 2000, 23, 38–44. [Google Scholar]
- Burr, P.C.; Avery, J.L.; Street, G.M.; Strickland, B.K.; Dorr, B.S. Historic and contemporary use of catfish aquaculture by piscivorous birds in the Mississippi Delta. Condor 2020, 122, duaa036. [Google Scholar] [CrossRef]
- King, D.T.; Blackwell, B.F.; Dorr, B.S.; Belant, J.L. Effects of aquaculture on migration and movement patterns of Double-crested Cormorants. Hum.-Wildl. Interact. 2010, 4, 77–86. [Google Scholar]
- Scherr, H.; Bowman, J.; Abraham, K.F. Migration and Winter Movements of Double-Crested Cormorants Breeding in Georgian Bay, Ontario. Waterbirds 2010, 33, 451–460. [Google Scholar] [CrossRef]
- King, D.T.; Strickland, B.K.; Radomski, A. Winter and summer home ranges and core use areas of Double-crested Cormorants captured near aquaculture facilities in the southeastern United States. Waterbirds 2012, 35, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Dorr, B.S.; Taylor, J.D.; Werner, S.J.; King, D.T.; Farquhar, J.F.; Mazzocchi, I.M.; McCullough, R.D. Summer and Migrational Movements of Satellite-Marked Double-Crested Cormorants from a Breeding Colony Managed by Egg-Oiling in Lake Ontario, USA. Waterbirds 2012, 35, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Dorr, B.S.; Aderman, T.; Butchko, P.H.; Barras, S.C. Management effects on breeding and foraging numbers and movements of double-crested cormorants in the Les Cheneaux Islands, Lake Huron, Michigan. J. Great Lakes Res. 2010, 36, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Ashmole, N.P. The regulation of numbers of tropical oceanic birds. IBIS 1963, 103b, 458–473. [Google Scholar] [CrossRef]
- Encyclopedia of Alabama. Guntersville Dam and Lake. 2015. Available online: http://encyclopediaofalabama.org/ARTICLE/h-2621 (accessed on 27 December 2022).
- Sawyer, C.N. Basic concepts of eutrophication. Water Pollut. Control Fed. 1966, 38, 737–744. [Google Scholar]
- Olin, M.; Rask, M.; Ruuhljärvi, J.; Kurkilahti, M.; Ala-Opas, P.; Ylönen, O. Fish community structure in mesotrophic and eutrophic lakes of southern Finland: The relative abundances of percids and cyprinids along a trophic gradient. J. Fish Biol. 2002, 60, 593–612. [Google Scholar] [CrossRef]
- Soil Survey Staff, Natural Resources Conservation Service, United States Department of Agriculture. Web Soil Survey. Available online: https://websoilsurvey.nrcs.usda.gov (accessed on 23 October 2016).
- Tennessee Wildlife Resource Agency. (n.d.); Tennessee State Government—TN.gov. Guntersville Reservoir Fishing. Available online: https://www.tn.gov/twra/fishing/where-to-fish/cumberland-plateau-r3/guntersville-reservoir.html (accessed on 27 December 2022).
- Moran, L.K.; Dorr, B.S.; Hanson-Dorr, K.C.; Moore, R.J.; Rush, S.A. Stable isotope analyses of movement and pre usage of Double-crested Cormorants in a reservoir system of the southeastern United States. Food Webs 2022, 31, e00220. [Google Scholar] [CrossRef]
- Sheehan, K.L.; Esswein, S.T.; Dorr, B.S.; Yarrow, G.K.; Johnson, R.J. Using species distribution models to define nesting habitat of the eastern metapopulation of Double-crested Cormorants. Ecol. Evol. 2017, 7, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Veum, L.M.; Dorr, B.S.; Hanson-Dorr, K.; Moore, R.J.; Rush, S. Double-crested Cormorant colony effects on soil chemistry, vegetation structure and avian diversity. For. Ecol. Manag. 2019, 453, 117588. [Google Scholar] [CrossRef] [Green Version]
- Migratory Bird Permits; Removal of Depredation Orders for Double-crested Cormorants to Protect Aqua-Culture Facilities and Public Resources; 83 Fed. Reg. 32805 (codified as 50 CFR 21); Fish and Wildlife Service: Washington, DC, USA, 2018.
- King, D.T.; Tobin, M.E. Capture and telemetry techniques for Double-crested Cormorants (Phalacrocorax auritus). Proc. Vertebr. Pest Conf. 2000, 19, 54–57. [Google Scholar] [CrossRef] [Green Version]
- Dunstan, T.C. A harness for radio-tagging raptorial birds. Inland Bird Band. News 1972, 44, 4–8. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 26 November 2017).
- Calenge, C. The package “adehabitat” for the R software: A tool for the analysis of space and habitat use by animals. Ecol. Model. 2006, 197, 516–519. [Google Scholar] [CrossRef]
- Worton, B.J. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 1989, 70, 164–168. [Google Scholar] [CrossRef]
- Seaman, D.E.; Powell, R.A. An evaluation of the accuracy of kernel density estimators for home range analysis. Ecology 1996, 77, 2075–2085. [Google Scholar] [CrossRef] [Green Version]
- Glahn, J.F.; May, A.; Bruce, K.; Reinhold, D. Censusing Double-crested Cormorants (Phalacrocorax auritus) at their winter roosts in the Delta Region of Mississippi. Colonial Waterbirds 1996, 19, 73–81. [Google Scholar] [CrossRef]
- King, D.T. Movements of Double-crested Cormorants among winter roosts in the Delta Region of Mississippi. J. Field Ornithol. 1996, 67, 205–211. [Google Scholar]
- Enstipp, M.R.; Grémillet, D.; Jones, D.R. The effects of depth, temperature and food ingestion on the foraging energetics of a diving endotherm, the Double-crested Cormorant (Phalacrocorax auritus). J. Exp. Biol. 2006, 209, 845–859. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.D.; Roby, D.D.; Collis, K. Foraging patterns of male and female Double-crested Cormorants nesting in the Columbia River estuary. Can. J. Zool. 2004, 82, 541–554. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sex | Colony Affiliation | Breeding Season | Incubation | Chick-Rearing | Post-Fledge | All Movements |
|---|---|---|---|---|---|---|
| May–July | 9–30 May | 31 May–4 July | 5 July–15 August | |||
| Male | South Sauty | 73.8 | 106.8 | 55.8 | - | 73.8 |
| Male | South Sauty | 17.2 | 40.7 | 6.3 | 16.0 | 6.8 |
| Female | South Sauty | - | - | - | 82.1 | 82.1 |
| Female | South Sauty | 80.8 | 172.3 | 36.6 | 62.0 | 64.6 |
| Female | South Sauty | 68.4 | 216.2 | 2.2 | - | 68.4 |
| Male | Connors | 12.3 | 27.3 | 7.6 | 8.1 | 9.5 |
| Female | Connors | 8.4 | 8.4 | - | - | 8.4 |
| Male | Connors | 20.5 | 29.8 | 1.3 | - | 20.5 |
| = | 40.2 ± 32.3 | 98.9 ± 80.6 | 18.3 ± 22.6 | 42.0 ± 31.0 | 41.8 ± 33.2 |
| Breeding Season | Incubation | Chick-Rearing | Post-Fledge | All Movements | |
|---|---|---|---|---|---|
| May–July | 9–30 May | 31 May–4 July | 5 July–15 August | ||
| Female | 52.5 ± 31.6 | 132.3 ± 89.4 | 19.4 ± 17.2 | 72.1 ± 10.1 | 55.9 ± 32.5 |
| Male | 30.95 ± 24.9 | 51.15 ± 32.5 | 17.75 ± 22.1 | 12.05 ± 4.0 | 27.6 ± 31.3 |
| South Sauty | 60.1 ± 25.1 | 134 ± 66.5 | 25.2 ± 22.1 | 53.4 ± 27.7 | 59.1 ± 26.8 |
| Connors | 13.7 ± 5.0 | 21.8 ± 9.6 | 4.5 ± 3.2 | 8.1 ± 0 | 12.8 ± 5.5 |
| Sex | Colony Affiliation | Breeding Season | Incubation | Chick-Rearing | Post-Fledge | All Movements |
|---|---|---|---|---|---|---|
| May–July | 9–30 May | 31 May–4 July | 5 July–15 August | |||
| Male | South Sauty | 11.3 | 16.9 | 10.8 | - | 11.4 |
| Male | South Sauty | 1.6 | 6.3 | 0.5 | 1.5 | 1.0 |
| Female | South Sauty | - | - | - | 13.1 | 13.1 |
| Female | South Sauty | 7.2 | 25.1 | 3.3 | 5.7 | 5.4 |
| Female | South Sauty | 8.2 | 30.1 | 0.3 | - | 8.2 |
| Male | Connors | 1.6 | 3.6 | 1.4 | 1.7 | 1.6 |
| Female | Connors | 1.2 | 1.2 | - | - | 1.2 |
| Male | Connors | 3.7 | 5.6 | 0.2 | - | 3.7 |
| = | 5.0 ± 4.0 | 14.6 ± 11.2 | 2.8 ± 4.1 | 5.5 ± 4.7 | 6.4 ± 5.9 |
| Breeding Season | Incubation | Chick-Rearing | Post-Fledge | All Movements | |
|---|---|---|---|---|---|
| May–July | 9–30 May | 31 May–4 July | 5 July–15 August | ||
| Female | 5.5 ± 3.8 | 18.8 ± 12.6 | 3.2 ± 4.4 | 9.4 ± 3.7 | 7.0 ± 5.0 |
| Male | 4.6 ± 4.6 | 8.1 ± 5.2 | 1.8 ± 1.5 | 1.6 ± 0.1 | 5.4 ± 4.8 |
| South Sauty | 7.1 ± 3.5 | 19.6 ± 9.0 | 3.7 ± 4.3 | 6.8 ± 4.8 | 7.8 ± 4.3 |
| Connors | 2.2 ± 1.1 | 3.5 ± 1.8 | 0.8 ± 0.6 | 1.7 ± 0 | 2.2 ± 1.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moran, L.L.K.; Dorr, B.S.; Hanson-Dorr, K.C.; Moore, R.J.; Rush, S.A. Space Use and Movements of Southeastern Breeding Double-Crested Cormorants (Nannopterum auritum) in the United States. Diversity 2023, 15, 453. https://doi.org/10.3390/d15030453
Moran LLK, Dorr BS, Hanson-Dorr KC, Moore RJ, Rush SA. Space Use and Movements of Southeastern Breeding Double-Crested Cormorants (Nannopterum auritum) in the United States. Diversity. 2023; 15(3):453. https://doi.org/10.3390/d15030453
Chicago/Turabian StyleMoran, Leah L. K., Brian S. Dorr, Katie C. Hanson-Dorr, R. J. Moore, and Scott A. Rush. 2023. "Space Use and Movements of Southeastern Breeding Double-Crested Cormorants (Nannopterum auritum) in the United States" Diversity 15, no. 3: 453. https://doi.org/10.3390/d15030453