
Bizarre Morphology Obscures Real Affiliation: An Integrative Study of Enigmatic Cephalaspid Philine denticulata from Arctic Waters Reveals Its Unique Phylogenetic Position

Abstract
:1. Introduction
2. Material and Methods
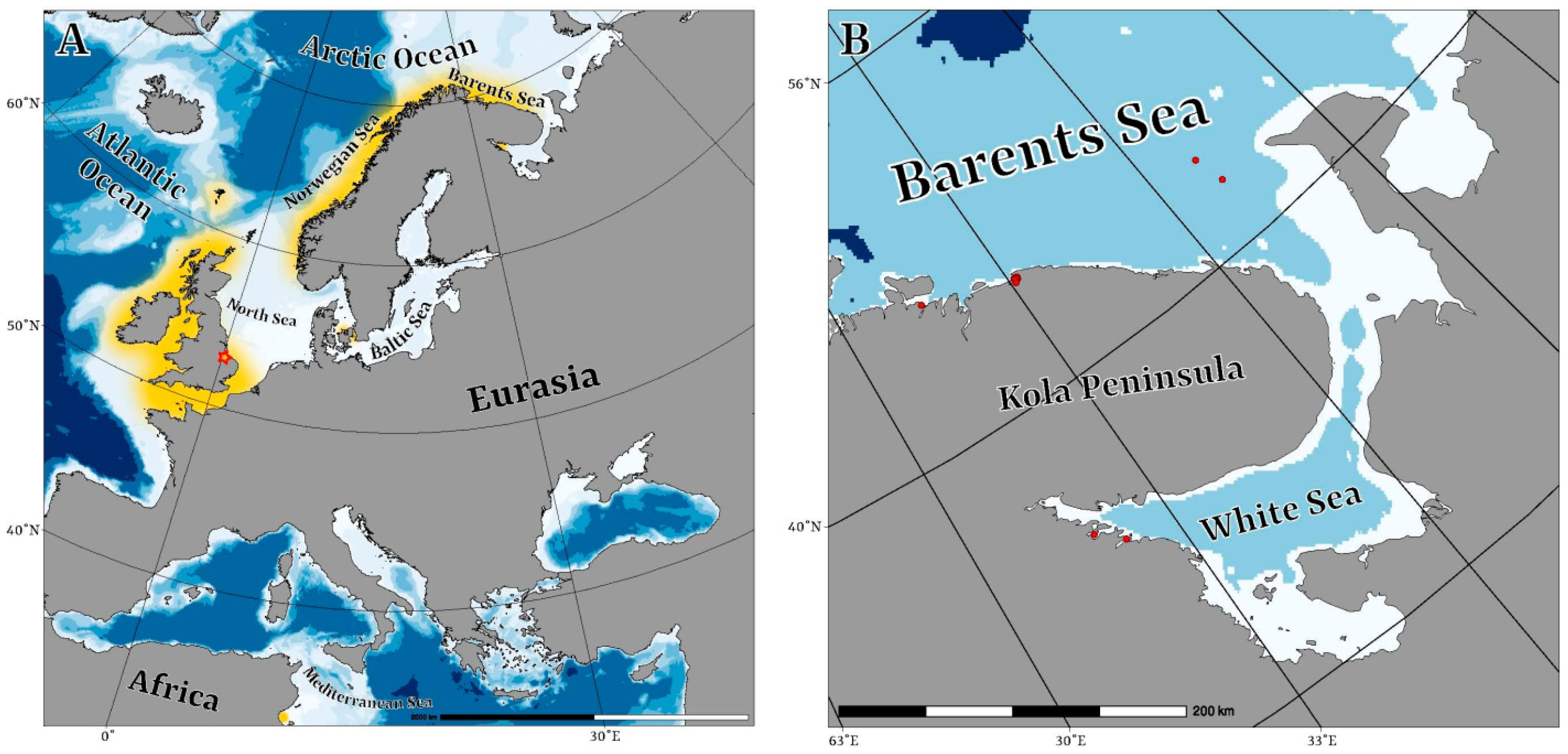
2.1. Collection Data
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Data Analysis
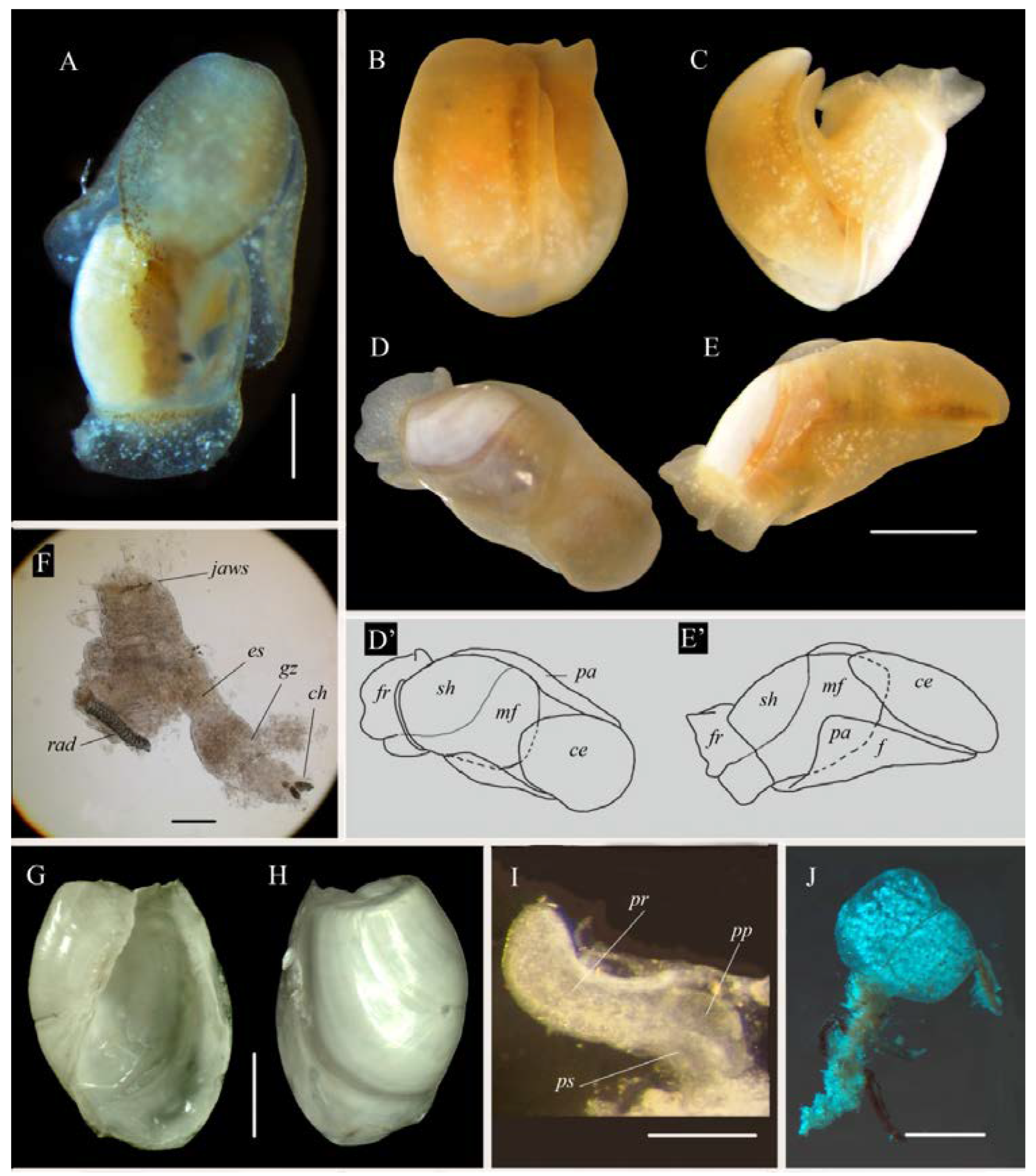
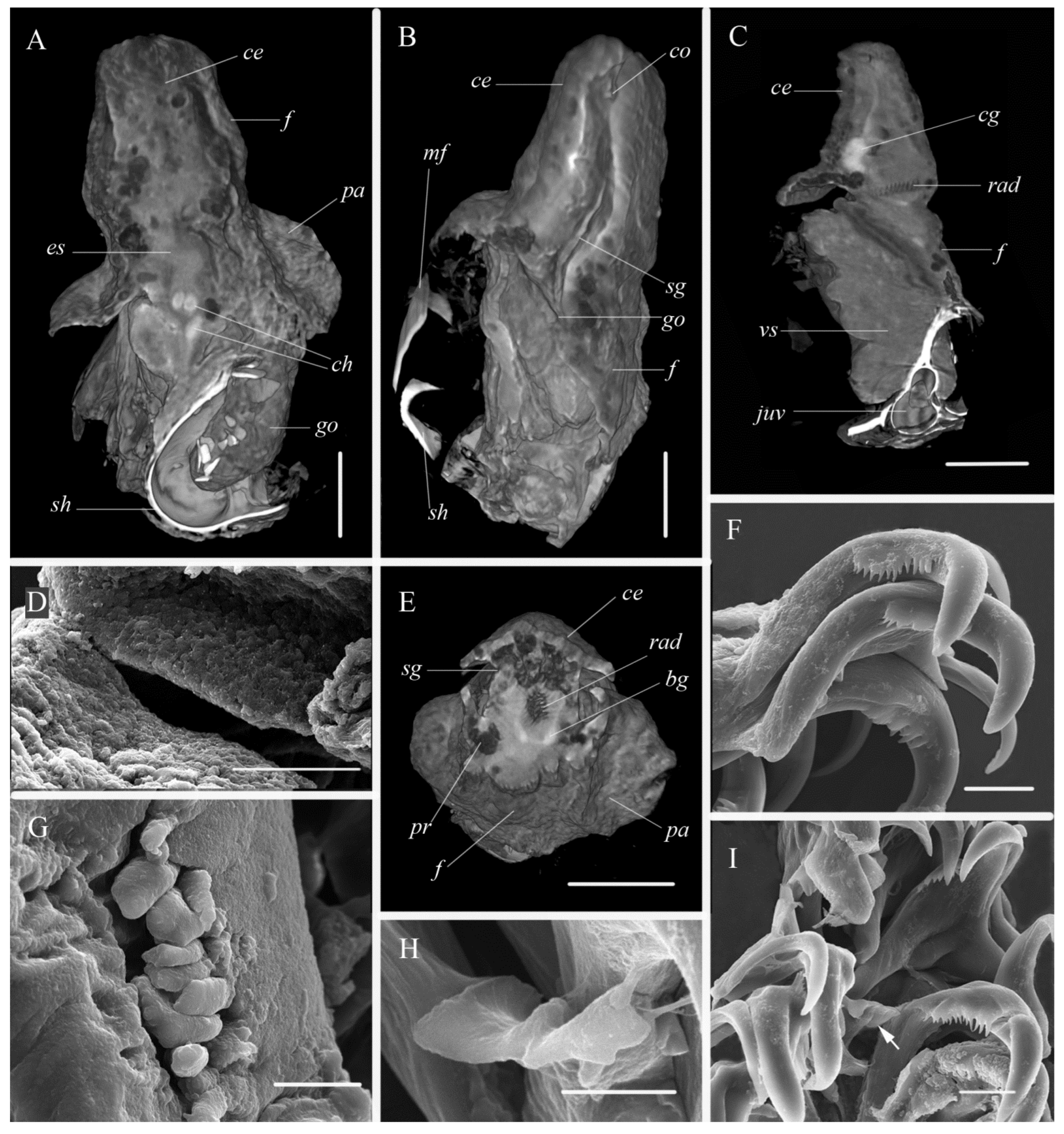
2.4. Morphological Analysis
2.5. Nomenclatural Acts
3. Results
3.1. Systematics
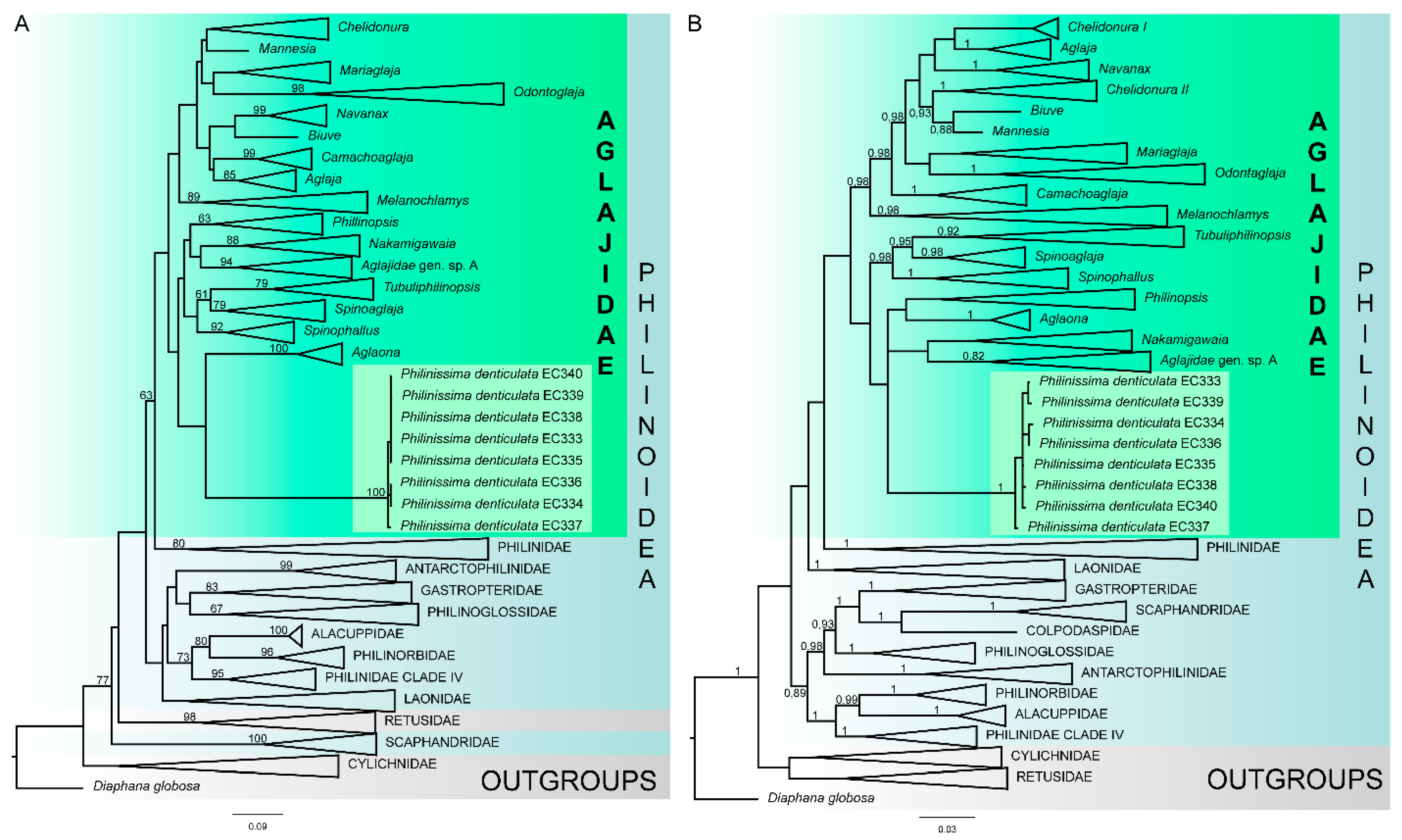
3.2. Molecular Analysis
4. Discussion
4.1. Molecular Data
4.2. Taxonomy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hendry, A.P.; Lohmann, L.G.; Conti, E.; Cracraft, J.; Crandall, K.A.; Faith, D.P.; Donoghue, M.J. Evolutionary biology in biodiversity science, conservation, and policy: A call to action. Evol. Int. J. Organic Evol. 2010, 64, 1517–1528. [Google Scholar] [CrossRef]
- Hoffmann, A.; Griffin, P.; Dillon, S.; Catullo, R.; Rane, R.; Byrne, M.; Sgrò, C. A framework for incorporating evolutionary genomics into biodiversity conservation and management. Clim. Change Responses 2015, 2, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Wiens, J.J. Missing data and the design of phylogenetic analyses. J. Biomed. Inf. 2006, 39, 34–42. [Google Scholar] [CrossRef]
- Graybeal, A.; Cannatella, D. Is It Better to Add Taxa or Characters to a Difficult Phylogenetic Problem? Syst. Biol. 1998, 47, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Zwickl, D.J.; Hillis, D.M. Increased taxon sampling greatly reduces phylogenetic error. Syst. Biol. 2002, 51, 588–598. [Google Scholar] [CrossRef] [Green Version]
- Nabhan, A.R.; Sarkar, I.N. The impact of taxon sampling on phylogenetic inference: A review of two decades of controversy. Brief. Bioinform. 2012, 13, 122–134. [Google Scholar] [CrossRef] [Green Version]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Zhang, N. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef]
- Funch, P.; Kristensen, M.R. Cycliophora is a new phylum with affinities to Entoprocta and Ectoprocta. Nature 1995, 378, 711–714. [Google Scholar] [CrossRef]
- Grell, K.G. Trichoplax adhaerens FE Schulze und die Entstehung der Metazoen. Nat. Rundsch. 1971, 24, 160–161. [Google Scholar]
- Kristensen, R.M. Loricifera, a New Phylum with Aschelminthes Characters from the Meiobenthos. Z. Zoolog. Syst. Evol. 1983, 21, 163–180. [Google Scholar] [CrossRef]
- Price, R.M.; Gosliner, T.M.; Valdés, Á. Systematics and phylogeny of Philine (Gastropoda: Opisthobranchia), with emphasis on the Philine aperta species complex. Veliger 2011, 51, 1–58. [Google Scholar]
- Ohnheiser, L.T.; Malaquias, M.A. Systematic revision of the gastropod family Philinidae (Mollusca: Cephalaspidea) in the north-east Atlantic Ocean with emphasis on the Scandinavian Peninsula. Zool. J. Linn. Soc. 2013, 167, 273–326. [Google Scholar] [CrossRef]
- Gonzales, C.; Gosliner, T. Six new species of Philine (Opisthobranchia: Philinidae) from the tropical Indo-Pacific. In The Coral Triangle: The 2011 Hearst Philippine Biodiversity Expedition; Williams, G.C., Gosliner, T.M., Eds.; California Academy of Sciences: San Francisco, CA, USA, 2014; pp. 351–383. [Google Scholar]
- Oskars, T.R.; Bouchet, P.; Malaquias, M.A. A new phylogeny of the Cephalaspidea (Gastropoda: Heterobranchia) based on expanded taxon sampling and gene markers. Mol. Phylog. Evol. 2015, 89, 130–150. [Google Scholar] [CrossRef]
- Valdés, Á.; Cadien, D.B.; Gosliner, T.M. Philinidae, Laonidae and Philinorbidae (Gastropoda: Cephalaspidea: Philinoidea) from the northeastern Pacific Ocean and the Beaufort Sea (Arctic Ocean). Zootaxa 2016, 4147, 501–537. [Google Scholar] [CrossRef]
- Malaquias, M.A.E.; Ohnheiser, L.T.; Oskars, T.R.; Willassen, E. Diversity and systematics of philinid snails (Gastropoda: Cephalaspidea) in West Africa with remarks on the biogeography of the region. Zool. J. Linn. Soc. 2016, 180, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Chaban, E.M. New genus of opisthobranch molluscs Antarctophiline gen. nov. (Cephalaspidea: Philinoidea) from the Cooperation Sea, Antarctica. Ruthenica 2016, 26, 49–56. [Google Scholar]
- Moles, J.; Avila, C.; Malaquias, M.A. Unmasking Antarctic mollusk lineages: Novel evidence from philinoid snails (Gastropoda: Cephalaspidea). Cladistics 2019, 35, 487–513. [Google Scholar] [CrossRef]
- Chaban, E.M.; Ekimova, I.A.; Schepetov, D.M.; Kohnert, P.C.; Schrodl, M.; Chernyshev, A.V. Euopisthobranch mollusks of the order Cephalaspidea (Gastropoda: Heterobranchia) of the Kuril-Kamchatka Trench and the adjacent Pacific abyssal plain with descriptions of three new species of the genus Spiraphiline (Philinidae). Progr. Oceanol. 2019, 178, 102185. [Google Scholar] [CrossRef]
- Brown, T. Illustrations of Recent Conchology of Great Britain and Ireland: With the Descriptions and Localities of All the Species, Marine, Land and Fresh Water, 2nd ed.; Smith, Elder, and CO.: London, UK, 1844; pp. 1–144. [Google Scholar]
- Sars, G.O. Bidrag til kundskaben om Norges Arktiske Fauna. I. Mollusca Regions Arcticae Norvegiae; Trykt hos A.W. Brøgger: Oslo, Norway, 1878; pp. 1–426. [Google Scholar]
- Odhner, N. Northern and Arctic invertebrates in the collection of the Swedish State Museum (Riksmuseum). III. Opisthobranchia and Pteropoda. Kung. Sven. Vetensk. Akad. Handl. 1907, 41, 1–114. [Google Scholar]
- Thompson, T.E. Biology of Opisthobranch Molluscs; The Ray Society: London, UK, 1976; Volume 1, pp. 1–207. [Google Scholar]
- Horikoshi, M. Reproduction, larval features and life history of Philine denticulata (J. Adams) (Mollusca—Tectibranchia). Ophelia 1967, 4, 43–84. [Google Scholar] [CrossRef]
- Høisæter, T. Distribution of marine, benthic, shell bearing gastropods along the Norwegian coast. Fauna Nor. 2009, 28, 5–106. [Google Scholar] [CrossRef] [Green Version]
- Nekhaev, I.O. Marine shell-bearing Gastropoda of Murman (Barents Sea): An annoteted check-list. Ruthenica 2014, 24, 75–121. [Google Scholar]
- Ivanova, N.V.; deWaard, J.; Hebert, P.D.N. An inexpensive, automation-friendly proto-col for recovering high-quality DNA. Mol. Ecol. Notes 2006, 6, 998–1002. [Google Scholar] [CrossRef]
- Chaban, E.M.; Ekimova, I.A.; Schepetov, D.M.; Chernyshev, A.V. The new genus Aglaona: The first abyssal aglajid (Heterobranchia: Cephalaspidea: Aglajidae) with a description of two new species from the north-western Pacific Oceans. Zool. J. Linn. Soc. 2022, 196, 198–214. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Philippe, H.; Sörhannus, U.; Baroin, A.; Perasso, R.; Gasse, F.; Adoutte, A. Comparison of molecular and paleontological data in diatoms suggests a major gap in the fossil record. J. Evol. Biol. 1994, 7, 247–265. [Google Scholar] [CrossRef]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A New and Scalable Tool for the Selection of DNA and Protein Evolutionary Models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Kobelt, W. Die Familie Bullidae; Verlag von Bauer & Raspe: Nürnberg, Germany, 1896; pp. 1–188. [Google Scholar]
- Nordsieck, F. Die europäischen Meeresschnecken (Opisthobranchia mit Pyramidellidae; Rissoacea) vom Eismeer bis Kapverden, Mittelmeer und Schwarzes Meer; Gustav Fischer Verlag: Stuttgart, Germany, 1972; pp. 1–327. [Google Scholar]
- Franz, D.R.; Clark, K. Occurrence of the cephalaspid Philine sinuate (Stimpson) in southern New England, with a discussion of the species. Veliger 1969, 12, 69–71. [Google Scholar]
- Thompson, T.E. Molluscs: Benthic Opisthobranchs (Mollusca: Gastropoda). Key and notes for the identifications of the species, 2nd ed.; Brill, E.J., Backhuys, W., Eds.; The Bath Press: Avon, UK, 1988; pp. 1–356. [Google Scholar]
- Lemche, H. Northern and Arctic Tectibranch Gestropods. A revision of the Cephalaspid species. Kong. Danske Videnskab. Selskab, Biol. Skrifter 1948, 5, 136. [Google Scholar]
- Chaban, E. Subclass Opisthobranchia: Orders Cephalaspidea, Anaspidea, Acochlidiida, Sacoglossa, Thecosomata, Gymnosomata. In List of Species of Free-Living Invertebrates of Eurasian Arctic Seas and Adjacent Deep Waters; Explorations of the Fauna of the Seas; Sirenko, B.I., Ed.; Zoological Institute of Russian Academy of Sciences: St. Petersburg, Russia, 2001; Volume 51, pp. 108–109. [Google Scholar]
- Pilsbry, H.A. Manual of Conchology; Structural and Systematic; 15; Academy of Natural Sciences of Philadelphia, Conchological Section: Philadelphia, PA, USA, 1895; pp. 1–436. [Google Scholar]
- Marshall, J.T. Addition to “British Conchology” VII. J. Conch. Lond. 1913, 14, 33–35. [Google Scholar]
- Pruvot-Fol, A. Mollusques Opisthobranches. Faune de France; P. Lechevalier: Paris, France, 1954; Volume 58, pp. 1–448. [Google Scholar]
- Eilertsen, M.H.; Malaquias, M.A. 2015. Speciation in the dark: Diversification and biogeography of the deep-sea gastropod genus Scaphander in the Atlantic Ocean. J. Biogr. 2015, 42, 843–855. [Google Scholar] [CrossRef] [Green Version]
- Chaban, E.M.; Kijashko, P.V. On two cephalaspid molluscs (Gastropoda: Opisthobranchia) with two gizzard plates, and description of a new genus and a new family. Zoosystematica Ross. 2016, 25, 204–215. [Google Scholar] [CrossRef]
- Gosliner, T.M. The Philinacea (Mollusca: Gastropoda: Opisthobranchia) of Aldabra Atoll, with descriptions of five new species and a new genus. Bull. Biol. Soc. Wash. 1988, 8, 79–100. [Google Scholar]
- Habe, T. Eight new and little known cephalaspid Opisthobranchia from Japan. Venus 1976, 35, 151–157. [Google Scholar]
- Zamora-Silva, A.; Malaquias, M.A. Molecular phylogeny of the Aglajidae head-shield sea slugs (Heterobranchia: Cephalaspidea): New evolutionary lineages revealed and proposal of a new classification. Zoo. J. Linn. Soc. 2018, 20, 1–51. [Google Scholar] [CrossRef]
- Adams, J. Descriptions of some minute shells. Trans. Linn. Soc. 1800, 5, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Gosliner, T.M. Six new species of aglajids opisthobranch molluscs from the tropical Indo-Pacific. Zootaxa 2011, 2751, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Camacho-García, Y.E.; Ornelas-Gatdula, E.; Gosliner, T.M.; Valdés, Á. Phylogeny of the family Aglajidae (Pilsbry, 1895) (Heterobranchia: Cephalaspidea) inferred from mtDNA and nDNA. Mol. Phyl. Evol. 2014, 71, 113–126. [Google Scholar] [CrossRef]
- Malaquias, M.A.E. New data on the heterobranch gastropods (‘opisthobranchs’) for the Bahamas (tropical western Atlantic Ocean). Mar. Biodiv. Rec. 2014, 7, e27. [Google Scholar] [CrossRef]
- Trianito, E.; Doneddi, M. Contibution to the knowledge of the molluscan fauna in the Marine Protected Area Tavolara-Punta Coda Cavallo: Sacoglossa, Umbraculida, Pleurobranchomorpha, Anaspidea, Cephalaspidea, Thecosomata (Gastropoda; Heterobranchia). Boll. Malac. 2016, 52, 79–89. [Google Scholar]
- Cooke, S.; Hanson, D.; Hirano, Y.; Ornelas-Gatdula, E.; Gosliner, T.M.; Chernyshev, A.V.; Valdés, Á. Cryptic diversity of Melanochlamys sea slugs (Gastropoda, Aglajidae) in the North Pacific. Zool. Scr. 2014, 43, 351–369. [Google Scholar] [CrossRef]
- Valdés, Á. Northeast Pacific benthic shelled sea slugs. Zoosymposia 2019, 13, 242–304. [Google Scholar] [CrossRef] [Green Version]
- Gosliner, T.M. Three new species of aglajid cephalaspidean mollusks from the tropical Indo-Pacific of the Verde Island Passage. Proc. Calif. Acad. Sci. 2014, 62, 191–205. [Google Scholar]
- Chaban, E.M.; Ekimova, I.A.; Chernyshev, A.V. Additional data on the opisthobranch fauna of Peter the Great Bay (Sea of Japan): Melanochlamys chabanae (Heterobranchia: Aglajidae). Bull. Russ. Far-East Malacol. Soc. 2018, 22, 79–91. [Google Scholar]
- Okutani, T. (Ed.) Marine Mollusks in Japan; Tokai University Press: Tokyo, Japan, 2000; pp. 1–1173. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Expedition/Station/Collector | Coordinates | Depth (m) | Gear | Number of Specimens | Density (Specimens/m2) |
|---|---|---|---|---|---|---|
| 11 August 2003 | G/V Pomuald Muklevich, st. 18-3, P. Lubin | 69,5275 N 32,4511 E | 60 | Van Veen grab | 3 | 23 |
| 11 August 2003 | G/V Pomuald Muklevich, st. 18-4, P. Lubin | 69,5275 N 32,4511 E | 60 | Van Veen grab | 1 | |
| 23 August 2003 | G/V Pomuald Muklevich, st. 68-2, P. Lubin | 68,6804 N 41,6753 E | 70.6 | Van Veen grab | 1 | 7 |
| 23 August 2003 | G/V Pomuald Muklevich, st. 68-3, P. Lubin | 68,6804 N 41,6753 E | 70.6 | Van Veen grab | 1 | |
| 23 August 2003 | G/V Pomuald Muklevich, st. 69, P. Lubin | 69,0160 N 41,4670 E | 78.9 | Van Veen grab | 1 | 3 |
| 03 August 2008 | R/V Professor Boiko, st. TS1-1, P. Lubin | 69,1744 N 35,1509 E | 44.7 | Van Veen grab | 16 | 343 |
| 03 August 2008 | R/V Professor Boiko, st. TS1-2, P. Lubin | 69,1744 N 35,1509 E | 44.7 | Van Veen grab | 48 | |
| 03 August 2008 | R/V Professor Boiko, st. TS1-3, P. Lubin | 69,1744 N 35,1509 E | 44.7 | Van Veen grab | 39 | |
| 03 August 2008 | R/V Professor Boiko, st. TS2-1, P. Lubin | 69,1830 N 35,1437 E | 74 | Van Veen grab | 13 | 110 |
| 03 August 2008 | R/V Professor Boiko, st. TS2-2, P. Lubin | 69,1830 N 35,1437 E | 74 | Van Veen grab | 12 | |
| 03 August 2008 | R/V Professor Boiko, st. TS2-3, P. Lubin | 69,1830 N 35,1437 E | 74 | Van Veen grab | 8 | |
| 03 August 2008 | R/V Professor Boiko, st. TS4-1, P. Lubin | 69,2079 N 35,1652 E | 110 | Van Veen grab | 3 | 10 |
| 03 August 2008 | R/V Professor Boiko, st. TS5-1, P. Lubin | 69,2209 N 35,1606 E | 122.3 | Van Veen grab | 2 | 7 |
| 03 August 2008 | R/V Professor Boiko, st. TS7-1, P. Lubin | 69,2089 N 35,2224 E | 88.3 | Van Veen grab | 2 | 13 |
| 03 August 2008 | R/V Professor Boiko, st. TS7-3, P. Lubin | 69,2089 N 35,2224 E | 88.3 | Van Veen grab | 2 | |
| 03 August 2008 | R/V Professor Boiko, st. TS8-3, P. Lubin | 69,2076 N 35.2464 E | 63.3 | Van Veen grab | 5 | 17 |
| 03 August 2008 | R/V Professor Boiko, st. TS9-1, P. Lubin | 69,1996 N 35.2520 E | 58 | Van Veen grab | 2 | 13 |
| 03 August 2008 | R/V Professor Boiko, st. TS9-3, P. Lubin | 69,1996 N 35.2520 E | 58 | Van Veen grab | 2 | |
| 04 August 2008 | R/V Professor Boiko, st. TS11-2, P. Lubin | 69,2132 N 35,2678 E | −2 | Van Veen grab | 1 | 3 |
| 04 August 2008 | R/V Professor Boiko, st. TS12-2, P. Lubin | 69,2215 N 35,2282 E | −2 | Van Veen grab | 8 | 43 |
| 04 August 2008 | R/V Professor Boiko, st. TS12-3, P. Lubin | 69,2215 N 35,2282 E | −2 | Van Veen grab | 5 | |
| 04 August 2008 | R/V Professor Boiko, st. TS13-1, P. Lubin | 69,2031 N 35,1952 E | 100.7 | Van Veen grab | 2 | 20 |
| 04 August 2008 | R/V Professor Boiko, st. TS13-2, P. Lubin | 69,2031 N 35,1952 E | 100.7 | Van Veen grab | 2 | |
| 04 August 2008 | R/V Professor Boiko, st. TS13-3, P. Lubin | 69,2031 N 35,1952 E | 100.7 | Van Veen grab | 2 | |
| June 2015 | A. Chaban, E. Chaban | White Sea, Kandalaksha Bay, Chupa Bay | 2 | scuba diving | 1 | |
| 20 June 2022 | A. Zhadan, E. Chaban | White Sea, Kandalaksha Bay, Ermolinskaya Bay | 8–9 | Sigsbee trawl | 18 |
| Species | Locality | DNA Extract No. | COI | 28S | H3 |
|---|---|---|---|---|---|
| P. denticulata | White Sea | EC333 | OQ445487 | OQ448320 | OQ453177 |
| P. denticulata | White Sea | EC334 | OQ445490 | OQ448321 | - |
| P. denticulata | White Sea | EC335 | - | OQ448322 | OQ453178 |
| P. denticulata | White Sea | EC336 | OQ445491 | OQ448323 | OQ453176 |
| P. denticulata | White Sea | EC337 | OQ445489 | OQ448324 | - |
| P. denticulata | White Sea | EC338 | - | - | OQ453175 |
| P. denticulata | White Sea | EC339 | OQ445488 | - | OQ453174 |
| P. denticulata | White Sea | EC340 | - | - | OQ453179 |
| Species | Shell | Penis | Gizzard Plates | Radula | Current Family | References |
|---|---|---|---|---|---|---|
| Philine infortunata Pilsbry, 1895 | external, globose | no data | no data | no data | Philinidae | [12] |
| “Philine” hemphilli Dall, 1919 | presumably internal, globose | no data | absent | 5:1:0:1:5 | Philinidae | “may be a member of Laona” [15] |
| “Philine” bakeri Dall, 1919 | ?internal, ovate; sculpture chain-like, callus “wing-like” | no data | absent | 2:1:0:1:2 | Philinidae | [15] |
| “Philine” rubrata Gosliner, 1988 | internal, bulloid; sculpture punctate | prostate simple | chitinous ridges | 2:1:0:1:2 | Philinidae | [48] |
| “Philine” baxteri Valdés, Cadien & Gosliner, 2016 | internal, bulloid: sculpture punctuate, callus “wing-like” | prostate simple | absent | 2:1:1:1:2 | Philinidae | [15], this study |
| Philinissima denticulata (J. Adams, 1800) comb. nov. | external, cylindrical; sculpture fine net-like | prostate short, simple; penial papilla unarmed | chitinous ridges | 1:1:1:1:1 | Aglajidae | [12], this study |
| Aglaona rudmani Chaban et al., 2022 | internal, bulloid, oval: sculpture chain-like | prostate short; penial papilla armed | absent | 2:1:0:1:2 | Aglajidae | [28] |
| A. valdesi Chaban et al., 2022 | internal, bulloid, oval; sculpture chain-like | prostate short; penial papilla armed | absent | 2:1:0:1:2 | Aglajidae | [28] |
| Laona zonata A. Adams, 1865 | internal, ovate, with brown band; sculpture net-like | no data | no data | no data | Laonidae | [49] |
| L. confusa (Ohnheiser & Malaquias, 2013) | partly external, globose, smooth | prostate simple | absent | 3:1:?:1:3 | Laonidae | [12,18] |
| Laona nanseni Malaquias, Ohnheiser, Oskars & Willanssen, 2016 | external, oval, with faint rugose sculpture | flat tube | absent | absent | Laonidae | [16] |
| “Philine” retifera (Forbes, 1844) | external, elongate; sculpture with reticulate pattern | no data | no data | no data | insertae sedis | [12,16] as Philine retifera (Forbes, 1844) insertae sedis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaban, E.; Ekimova, I.; Lubin, P.; Nikitenko, E.; Schepetov, D. Bizarre Morphology Obscures Real Affiliation: An Integrative Study of Enigmatic Cephalaspid Philine denticulata from Arctic Waters Reveals Its Unique Phylogenetic Position. Diversity 2023, 15, 395. https://doi.org/10.3390/d15030395
Chaban E, Ekimova I, Lubin P, Nikitenko E, Schepetov D. Bizarre Morphology Obscures Real Affiliation: An Integrative Study of Enigmatic Cephalaspid Philine denticulata from Arctic Waters Reveals Its Unique Phylogenetic Position. Diversity. 2023; 15(3):395. https://doi.org/10.3390/d15030395
Chicago/Turabian StyleChaban, Elena, Irina Ekimova, Pavel Lubin, Ekaterina Nikitenko, and Dimitry Schepetov. 2023. "Bizarre Morphology Obscures Real Affiliation: An Integrative Study of Enigmatic Cephalaspid Philine denticulata from Arctic Waters Reveals Its Unique Phylogenetic Position" Diversity 15, no. 3: 395. https://doi.org/10.3390/d15030395