
Species Composition and Structure of Beetle Associations in Caves of the Częstochowa Upland, Poland

1
Department of Natural History, Upper Silesian Museum, Jana III Sobieskie-go 2 Square, 41-902 Bytom, Poland
2
Institute of Biology, University of Opole, Oleska 22, 45-052 Opole, Poland
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(3), 345; https://doi.org/10.3390/d15030345
Submission received: 23 December 2022
/
Revised: 10 February 2023
/
Accepted: 22 February 2023
/
Published: 1 March 2023
(This article belongs to the Special Issue Women’s Special Issue Series: Diversity)
Abstract
:This paper presents the study’s results on beetles (Insecta: Coleoptera) inhabiting caves in the Częstochowa Upland, southern Poland. During two years of research, 2084 specimens, representing 105 species from 19 beetle families, were collected. The obtained results indicate that many beetle species choose to inhabit caves despite lacking specific adaptations for living in such environments. The cave entrance zone is the most attractive place for surface species to inhabit because its climatic conditions are more stable than outside the cave, some sunlight is present, and the availability of organic matter is high. In the deeper parts of the studied caves, the number of occurring species rapidly decreased. Three species were recognised as troglobionts, four as troglophiles and 32 as subtroglophiles. Speonomus normandi hydrophilus, Choleva lederiana gracilenta and Quedius mesomelinus mesomelinus were identified as the most frequent in the studied caves. Speonomus normandi hydrophilus is endemic to the Arize massif in the Central Pyrenees (France) and was experimentally introduced into the Dzwonnica cave (Poland) in 1982, while Ch. lederiana gracilenta is one of two troglobitic beetles native to Polish caves and Q. imesomelinus mesomelinus is a widespread, very common troglophile.
Keywords:
troglobiont; troglophile; eutroglophile; subtroglophile; trogloxenes; round fungus beetles; rove beetles; cave; Poland1. Introduction
Beetles (Coleoptera) are the most species-rich order of insects, inhabiting various terrestrial and aquatic environments and occurring in almost all climatic zones [1,2]. Beetles can be herbivorous, carnivorous, necrophagous, saprophagous, and mixophagous, and because of this versatility, they have also colonised subterranean environments [1,2]. Some, named troglobionts, are closely associated with caves, while others prefer caves but live in different subterranean habitats (troglophiles) and some temporarily inhabit caves (trogloxenes) [3,4]. Beetles are also found in shallow subterranean habitats, termed the Mesovoid Shallow Substratum (MSS) [5,6,7,8,9,10]. The MSS comprises small crevices and scree slopes in rock debris [8,9,10]. The uniqueness of this environment relies on a connection between subterranean conditions—such as lack of light—with a direct inflow of organic matter from the surface [7,9,10].
Beetles also dwell in caves in Poland, and several species are known from previous research conducted in recent years [11,12]. However, a lack of comprehensive data about beetles inhabiting the subterranean environment in Poland was the main impetus for the authors to undertake this research. In Poland, caves occur in several of its southern regions, such as the Sudety Mountains, Tatra Mountains, Kraków-Częstochowa Upland, Carpathian Flysch Belt, Nidziańska Basin and Świętokrzyskie Mountains. The total number of caves is over 5000; the majority (about 1500) are located in the Kraków-Częstochowa Upland [13].
The purpose of the present research was to observe the species communities of beetles from caves of the Częstochowa Upland, which is a part of the Kraków-Częstochowa Upland. Special attention was paid to the degree of certain species’ relations with the subterranean habitat. Moreover, the particular beetle association structure was also analysed.
2. Materials and Methods
2.1. Study Area
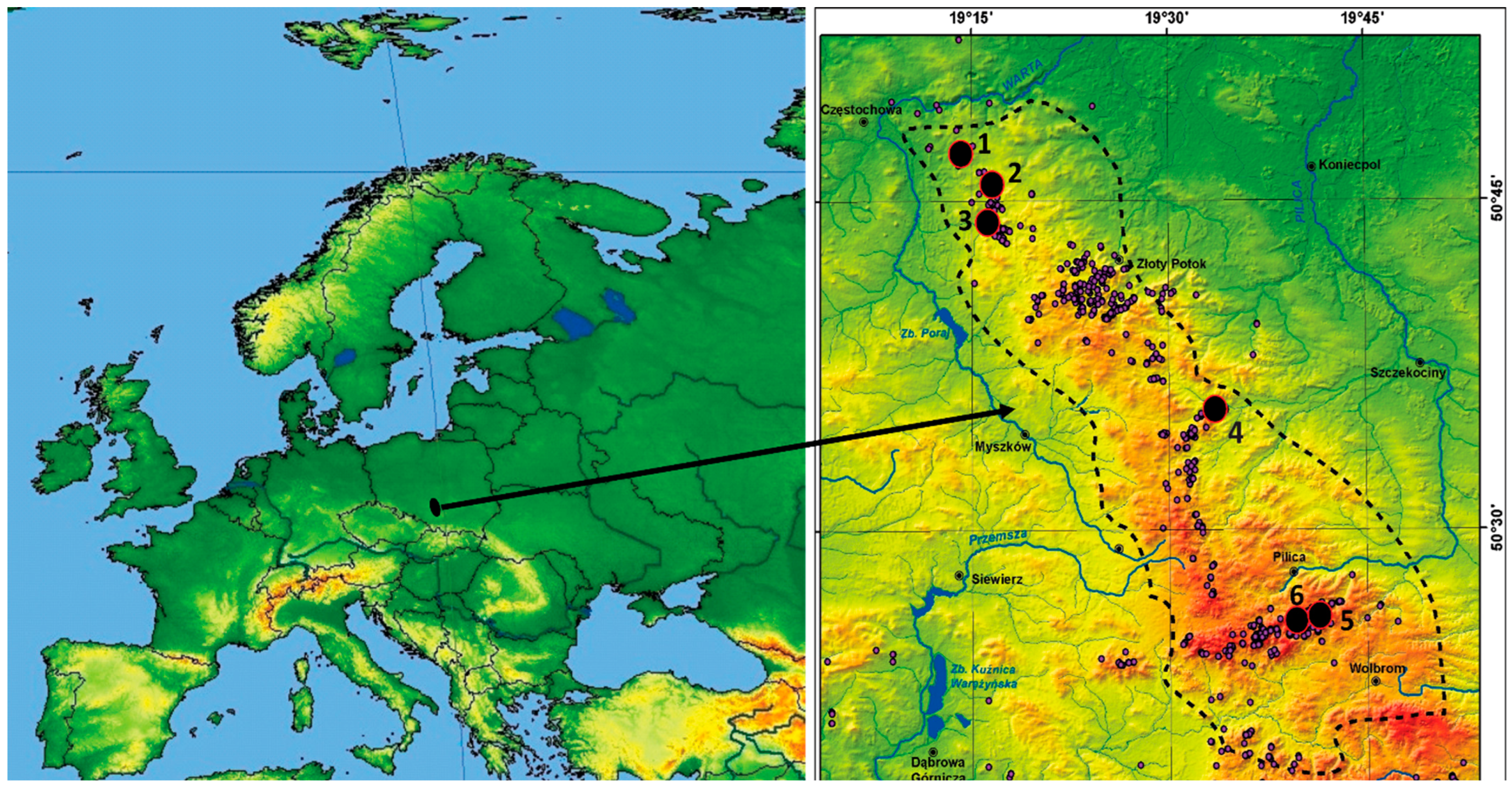
Six caves in the Częstochowa Upland were investigated (Figure 1); these were previously described in detail by Kocot-Zalewska and Woźnica [14]. Graphic diagrams of the studied caves have been taken from the public website of the Polish Geological Institute—National Research Institute [15]. All these caves are horizontal, with similar lengths and altitude a.s.l., but with slight height differences [16,17]. The size and exposure of the entrance, shape, configuration of passages and vegetation around the cave entrance vary for each cave (Figure 2, Figure 3 and Figure 4). The caves are situated in forest zones except for the Towarna cave, which lies in an area with xerothermic grassland. Five of the six studied caves have one entrance; only the Kroczycka cave has two, but both are much smaller than the entrances of other caves.
2.2. Sampling Design
The specimens were sampled from December 2014 to February 2017 using 5 cm-diameter Barber’s traps with 30% propylene glycol (as a preservative liquid). From December 2014 to January 2016, the material was sampled constantly (12 samples from every site per year). The following year, samples were gathered every two months (thus, six times per year). The direct-search method was also used for collecting the insects. However, individuals sampled in such a way were used only for the qualitative analysis and were not included in the quantitative evaluation.
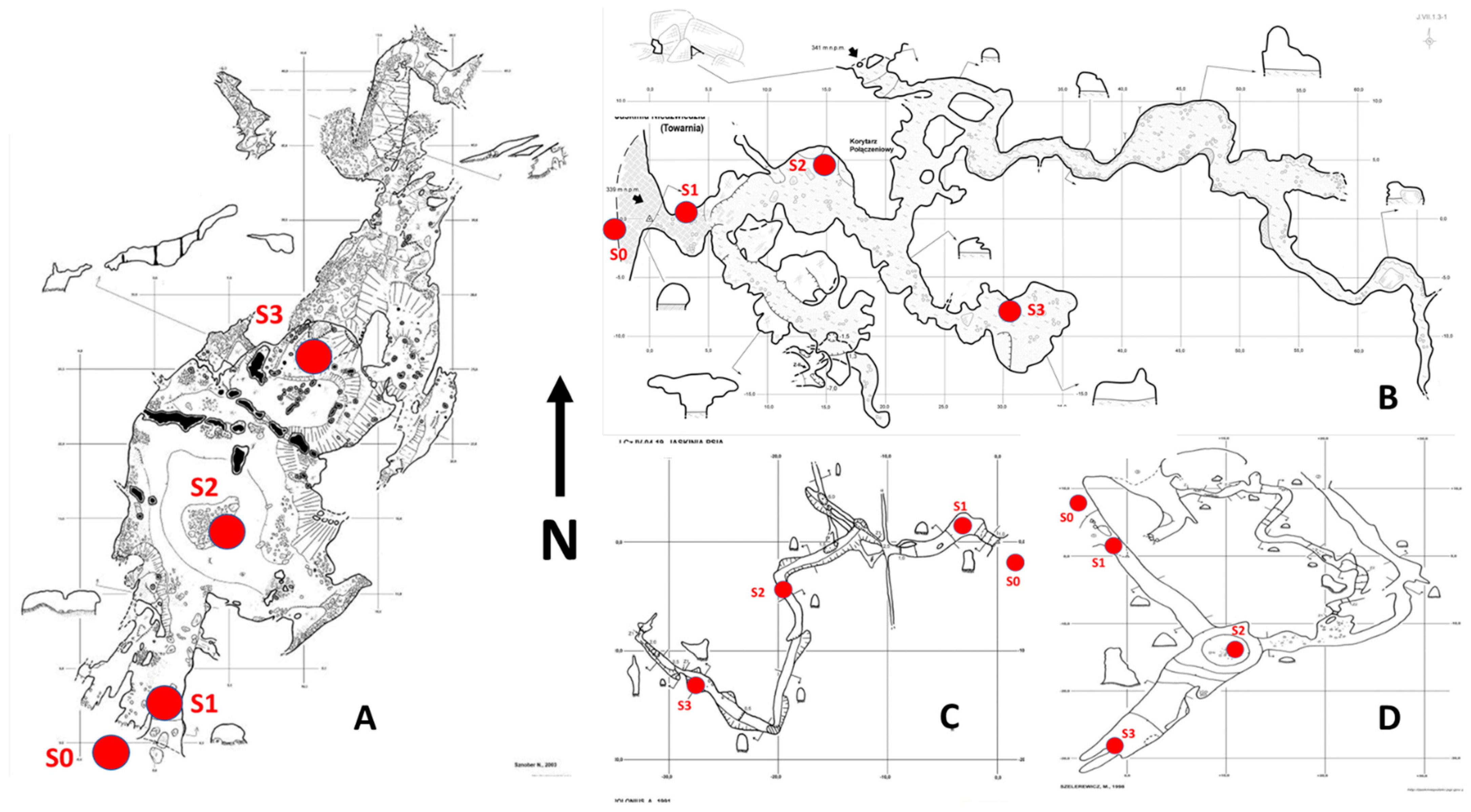
The traps were always situated in the same area in each cave. The first was placed in the entrance zone of the cave (S1), the second in the middle of the passage (S2) and the third at the end of the passage (S3) (Figure 3). Additionally, one trap was always placed in the direct surroundings of each cave entrance (S0) (Figure 3).
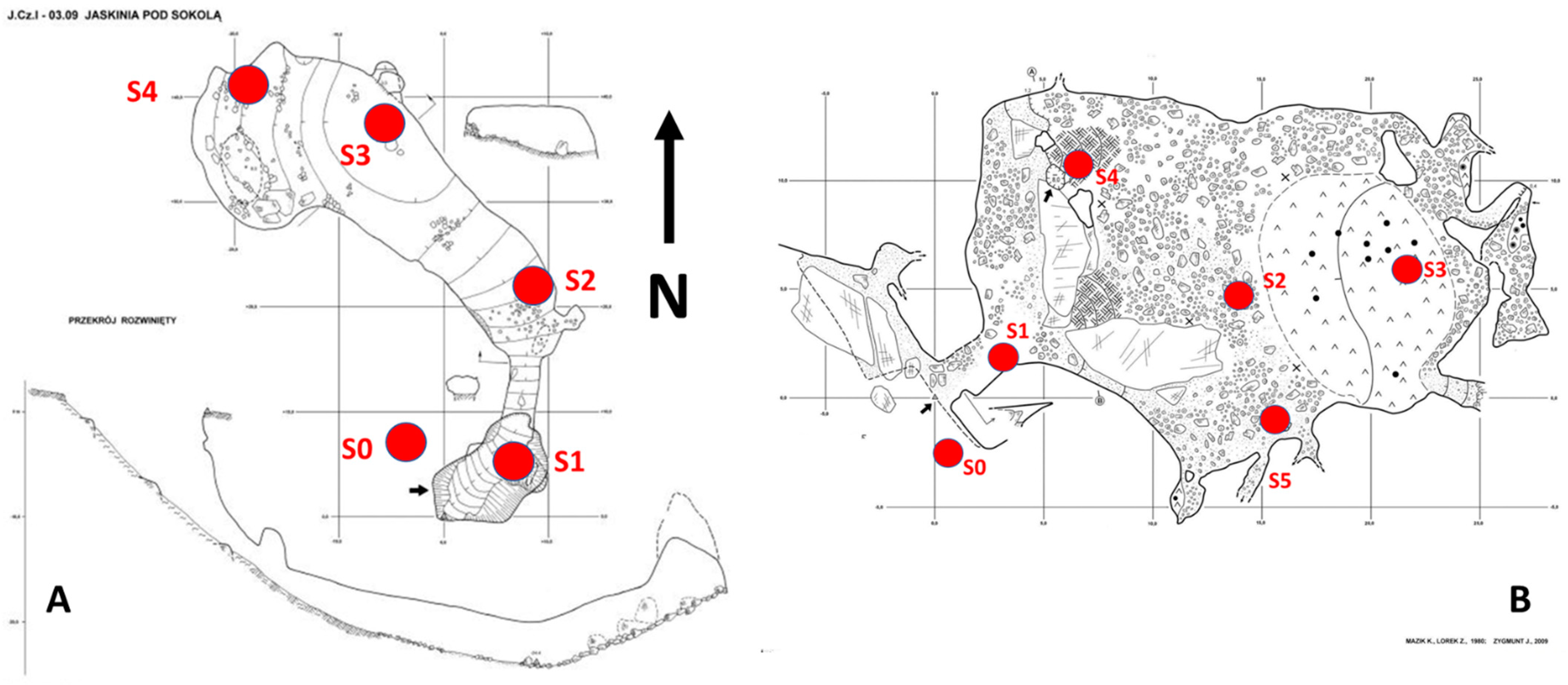
Because of the enormous entrance of the Pod Sokolą cave, the entrance zone (ca 3–4 m inside the cave) was not characterised by specific features. Therefore, an additional trap was used there. In this cave, for the statistical analyses, the trap in S2 was treated as the entrance zone trap, the trap in S3 as a transition zone trap, and that in S4 as a deep zone trap (Figure 4A).
In the Kroczycka cave, four traps were also used, due to the cave morphology resembling a rectangle with two entrances (Figure 4B). Traps S1 and S4 were located near the entrances, and, in the statistical analyses, they were used collectively as the entrance zone traps. Trap S5 was placed approximately in the middle of the cave length, and was thus treated as a trap appropriate for the transition zone. Trap S3 was placed at the end of the cave and represented a deeper part of the cave. Trap S2 marked a microclimate measurement point.
2.3. Microclimate Measurements
The temperature and humidity were measured from December 2014 to February 2017 in the studied caves. The Assmann psychrometer was set up monthly at the same points (S1 to S3 in every cave, and S4 in the Pod Sokolą cave). The psychrometer consisted of two thermometers (wet and dry) and an electric fan. In situ, the temperature was measured on both thermometers, and then, using psychrometric tables, the relative humidity was estimated. It was not possible to use automatic data loggers.
2.4. Data Analysis
In order to identify species associated with the subterranean environment, the dominance was calculated, and habitat and food preferences were analysed. Additionally, species richness for each cave was estimated using Hill numbers (qD, sensu Jost [18]). It was parameterised with q values of 1 and 2 to obtain the Shannon–Wiener diversity (q = 1) and Simpson diversity (q = 2) [19,20] (Appendix A, Formulas (A1) and (A2)). Both formulas are modified versions of the Shannon–Wiener and Simpson indices, respectively [18,19,20]. However, modification of the first index enables the elimination of favouritism of rare or common species, while Simpson diversity puts more weight on the frequencies of species and discounts rare species [19,20]. The analyses were performed using the PAST3 software ver. 4.03 [21].
The dominance index was calculated to detect the dominant species in particular caves (Formula (A3)). All species were classified within four groups, namely, eudominants (above 10.0%), dominants (5.1–10.0%), subdominants (2.1–5.0%) and recedents (1.0–2.0%) [22,23].
The analyses of species habitat and food preferences were based on the literature data [24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54]. According to the habitat, species were assigned to the proper environment groups, e.g., species living in the forest, grasslands, MSS (inhabiting mammal burrows and nests, living under moss, stones, in cave entrances) and others. Regarding diet, the studied beetle species were divided into five groups: herbivores, predators, fungivores, detritivores and omnivores. The group of detritivores includes all the phytosaprophages, zoosaprophages, coprophages and necrophages.
For subterranean species, we used four categories of ecological classification based on Sket’s concept [55], i.e., the group of troglobionts consists of species exclusively found in caves and, in the literature, considered as this form. The group of eutroglophiles is formed by species consistently found in caves throughout the year. However, they are generally epigean species. The group of subtroglophiles is composed of species that temporarily occur in caves; however, they were collected several times in many caves. Those species prefer living in mesovoid shallow substratum. The trogloxenes group includes accidental species for which caves and other subterranean shelters are not a typical habitat.
Moreover, a different group found in shallow subterranean habitats—MSS—was recognised [5,6,7]. In order to assign each of the analysed beetle species to the appropriate ecological category, we used previously published data [56,57,58,59,60,61]. All usable information was involved, i.e., the presence of species throughout the year, food and habitat preference, the area of the cave in which the species was most frequently caught.
3. Results
3.1. Species Dispersal
A total of 2084 specimens of Coleoptera were collected from the traps (Table S1). These individuals represented 105 species in 61 genera and 19 families (Table S2). Three families accounted for over 96% of total individuals, i.e., Leiodidae (74%), Staphylinidae (12.6%) and Cryptophagidae (9.7%).
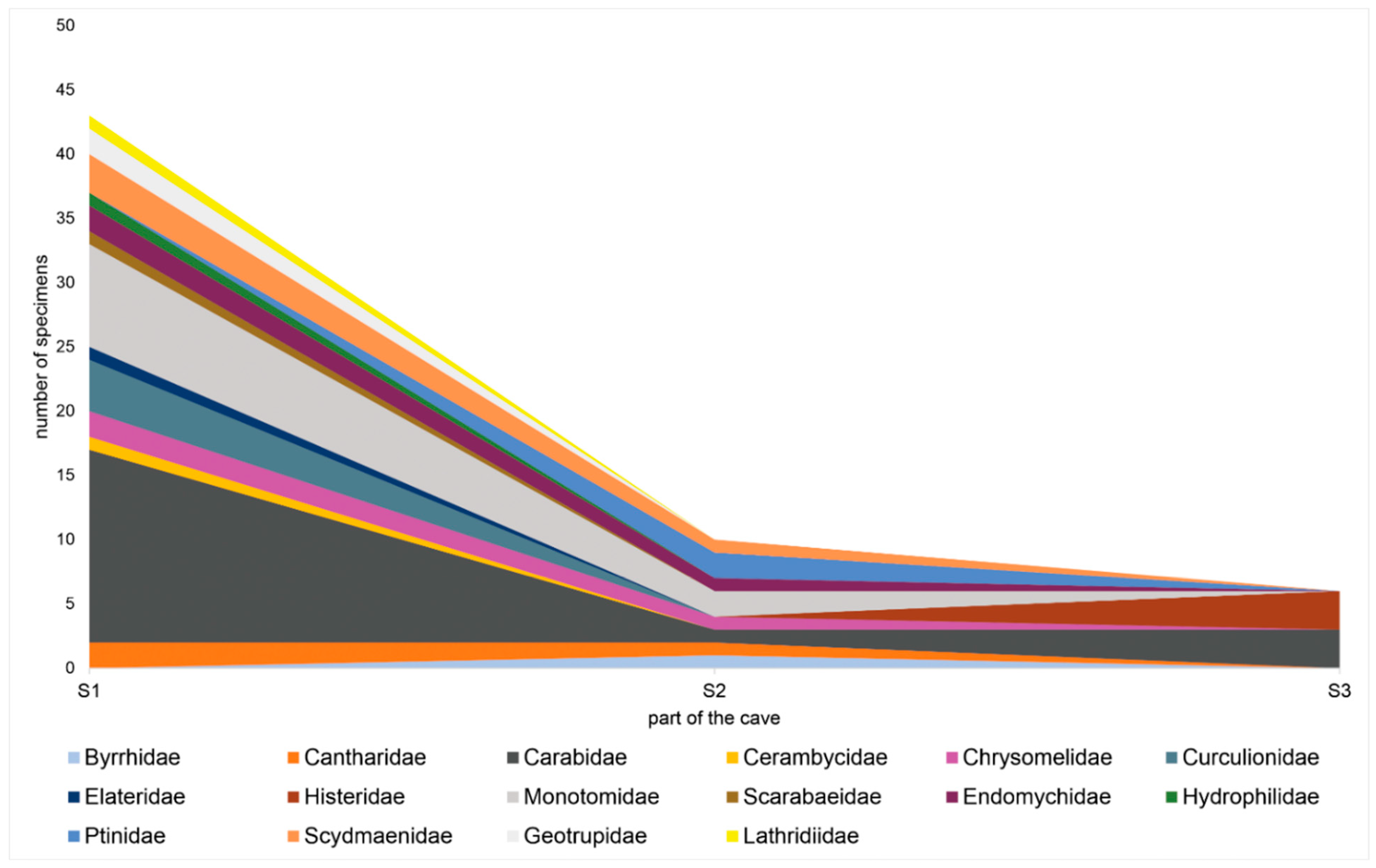
Additionally, we compared the frequency of particular species collected from caves with results obtained from the traps outside the studied caves (especially those species considered eutroglophiles and troglobionts). None of the troglobionts and eutroglophiles, was sampled outside the cave, with one exception. Several specimens of Catops picipes (Fabricius, 1787) were collected from traps located outside the caves. However, sample sites S0 were still situated among rocks and debris near the cave entrances. The presence of the beetles inside the caves varied and depended on the zones within the caves (Figure 5). Many collected specimens for each family were also correlated with the cave zones (Figure 6). When certain zones of the caves were considered, the entrance zone (S1) was the most numerous in species (10 to 35 species). Five to eleven species were collected in the middle of the passages (S2), whereas at the end of the caves (S3), one to eleven species were recorded. The results presenting the number of recorded species in particular parts of each cave are provided in Table 1, and the total number of species in each cave is presented in Table 2.
3.2. Species Richness
The number of species in particular caves differed (Table 2). The highest numbers were observed in the Pod Sokolą cave, while the lowest were observed in the Psia and Kroczycka caves. The highest species richness was calculated for the Zegar and Kroczycka caves, and the lowest in the Towarna cave (Table 2).
3.3. Species Dominance
Speonomus normandi hydrophilus (Jeannel, 1907), representing the family Leiodidae, was the most abundant among collected specimens (a total of 1120 individuals, 53% of all sampled specimens). Despite its abundance, all its specimens were exclusively found in the Towarna cave. Choleva lederiana gracilenta Szymczakowski, 1957, also from Leiodidae, was the second most abundant beetle, comprising 11% of the total collected specimens. As with S. normandi hydrophilus, all its individuals were exclusively sampled in one cave, namely, the Pod Sokolą cave. The third most abundant species found (representing 3.8% of the assemblage) was the staphylinid Quedius mesomelinus mesomelinus Marsham, 1802, which was found in each of the studied caves. Moreover, each cave had its own different set of eudominants and dominants (Table 3).
3.4. Habitat and Food Preferences
The forest beetles appeared to be the most numerous group (45 species, 43%), whereas the smallest group comprised species that occur in grasslands (three species, 3%) (Table S3). Beetles that occur in forests and grasslands were represented by eight species (7.5%). Thirty-two species were classified as those inhabiting MSS environments (30.5%). Most importantly, four species were assigned to the group of strict cave-dwellers (4%). Eight per cent of the collected material represented species inhabiting other environments (e.g., Oxypoda annularis (Mannerheim, 1830) or Anotylus sculpturatus (Gravenhorst, 1806)). Food preferences for 78 species (74% of the collection) were possible to estimate (Table S3). The most numerous groups were the detritivores (36%, 28 species) and predators (33%, 26 species). Fifteen species were classified as fungivores (18%). The minor groups comprised herbivores (three species, 4%), mixophagous and coprophagous (one species in each group, 1.5%).
Unfortunately, for all other species (especially those representing the family Staphy-linidae), these data were impossible to collect due to inadequate knowledge regarding their biology.
3.5. Ecological Classification
Among the collected beetles, three were classified as troglobionts (Speonomus normandi hydrophilus, Choleva lederiana gracilenta and Catops tristis infernus Szymczakowski, 1957), four as eutroglophiles (Quedius mesomelinus mesomelinus, Tachinus subterraneus (Linnaeus, 1758), Omalium validum Kraatz, 1857, Catops picipes), 32 as subtroglophiles or MSS species and 66 as the trogloxenes (Table S2).
3.6. Microclimatic Analyses
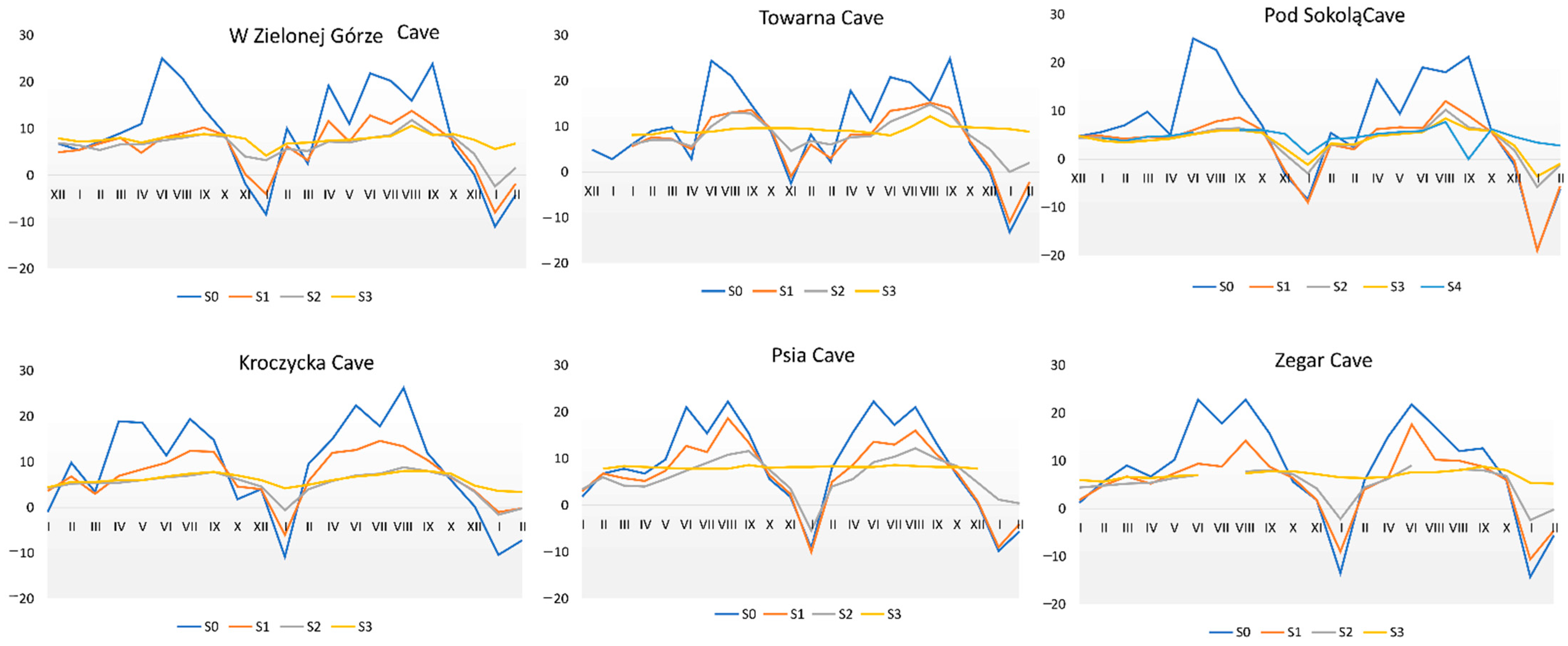
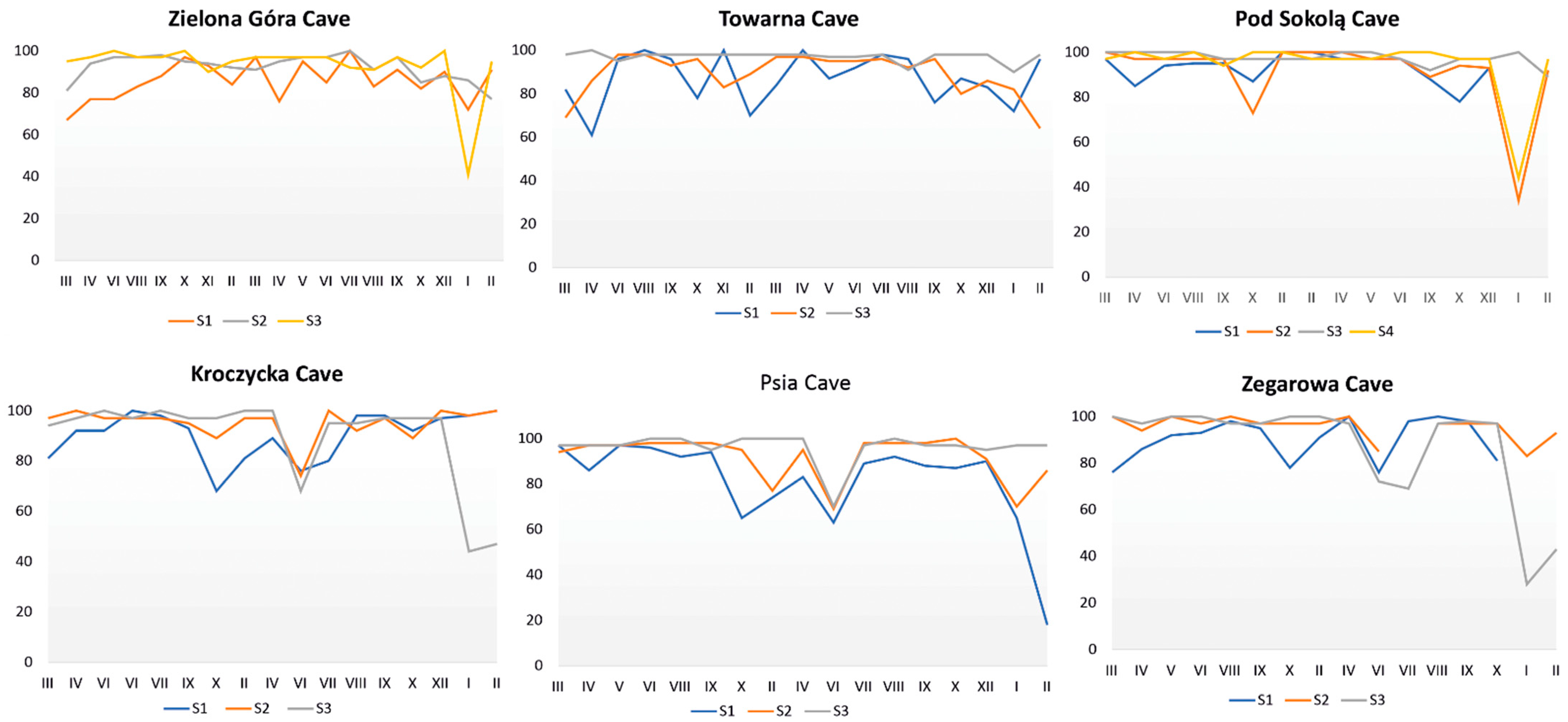
The temperature in all studied caves fluctuated, but the greatest temperature changes were detected in the entrance zone (S1). Additionally, it was observed to decrease towards the deeper parts of the cave. The most stable temperatures were observed at the terminal parts of the caves (Figure 7). Humidity in the studied caves was generally high. However, it fluctuated similarly to temperature, especially in the entrance zone (S1). Notably, humidity fluctuations during the summer and winter periods were recorded even in the deeper parts of the caves (Figure 8).
4. Discussion
4.1. Species Richness
Altogether, 105 Coleoptera species were detected in the studied caves (Table S2). The beetle species number provided in the present study is twice larger than that reported in all previous studies in Polish caves [11,12].
In our study, we confirmed the presence of a beetle species group that commonly inhabits Polish caves [11,12,56,62,63,64,65]. These include: Quedius mesomelinus mesomelinus (Staphylinidae), occurring in all cave regions of Poland [12,56,62,63,64,65]; Choleva cisteloides (Frölich, 1799) and Catops picipes (Leiodidae), in caves in the Ojców region [12,64]; Catops fuscus (Panzer, 1794) (Leiodidae), in caves in the Sudety Mountains [12,62,63,65]; and Choleva bicolor Jeannel, 1923 (Leiodidae), Catops fuliginosus Erichson, 1837 (Leiodidae), Tachinus subterraneus (Staphylinidae), Omalium rivulare (Paykull, 1789) and O. excavatum Stephens, 1834 (Staphylinidae), and Otiorhynchus scaber (Linnaeus, 1758) (Curculionidae) in caves in the Tatra Mountains [12,56].
The highest species frequencies were observed in Leiodidae, Staphylinidae and Cryptophagidae. The occurrence of round fungus beetles (Leiodidae) in the caves was not surprising, because many species of this family are highly associated with subterranean environments [66]. Many rove beetles (Staphylinidae) are also known from subterranean areas [8,66]. However, silken-fungus beetles (Cryptophagidae) are not usually associated with subterranean environments, being closely associated with forest ecosystems and often considered within the ranks of saproxylic and xylobiont Coleoptera [67]. Therefore, the high species frequency of silken-fungus beetles in the studied caves was slightly unexpected. However, most of the Cryptophagidae specimens were collected from the entrance zone of one cave (the W Zielonej Górze cave), and only a few occurred in its deeper zones and in the other caves. Thus, despite collecting about 200 specimens, only two species were classified as subtroglophiles, and five were classified as trogloxenes.
The highest species richness among the studied caves was recorded for the Zegar cave (15.95863401 and 1.051524711 in Shannon diversity and Simpson diversity indices, respectively) (Table 2). The high frequency of those species among the samples may explain such a result. The Towarna cave represented the lowest value of the Shannon diversity (1.447734615), but the highest of the Simpson diversity (7.692307692). It can be explained by the extremal domination of one species (among 1206 specimens in the collection, 1120 belonged to Speonomus normandi hydrophilus). The Psia cave represented rather high values of the abovementioned indices (8.758284041 and 2.949852507, respectively). It was surprising due to the generally poor habitat quality of the cave, which was also confirmed by the low frequency of specimens representing other arthropod groups compared to other studied caves. In addition, we examined the contents of organic carbon in every sampling place of the studied caves only once. The Psia cave is characterised by twice the lower value of organic carbon than other caves. However, this parameter was not elaborated on further due to the small amount of data.
In terms of cave zonation, the highest values of the analysed indices were associated with the entrance zones. This was an expected result because the entrance zones were always characterised by toned temperature fluctuation (in comparison to the surface), a lack of violent weather phenomena, a high accumulation of organic matter, and the presence of direct or indirect sunlight. All these conditions significantly impact cave-dwelling by invertebrate fauna, including beetles. Stable temperatures, high relative humidity and extreme organic matter accumulation were associated with the deepest zones. Despite this, the lowest values of species richness indices resulted from complete darkness, which is not conducive to penetration by surface fauna.
4.2. Species Dominance
The analyses of species dominance indicated the overwhelming contribution of accidental species in the entrance zone (S1), especially in the caves with large entrances. This was evident, for instance, in the Pod Sokolą, Zegar and W Zielonej Górze caves, where the accidental species reached 60% to 80% of the total amount of collected species (Table 3).
Moreover, each studied cave had a different set of eudominant or dominant species (Table 3). In the W Zielonej Górze cave, two species (Cryptophagus distinguendus J. Sturm, 1845 and C. punctipennis Ch.N.F. Brisout de Barneville, 1863) represented the eudominant class. Speonomus normandi hydrophilus, in the Towarna cave, and Choleva lederiana gracilenta and Catops tristis infernus, in the Pod Sokolą cave, exhibited a spectacular dominance. This reached 98% in the deepest part of Towarna (S3) for S. normandi hydrophilus and 87.5% and 20% in the deepest part of Pod Sokolą (S4) for the latter two beetles, respectively.
In the caves of the central and southern part of the investigated area—i.e., the Kroczycka, Psia and Zegar caves—none of the analysed species achieved a high dominance value.
4.3. Habitat and Food Preferences
The examination of food preferences indicated that beetles mostly choose detritus or fungi, and some are predators. Phytophagous species represented only a tiny part of the collected material. Similar results were demonstrated for invertebrates collected in caves in Germany [68]. The analyses of habitat preferences showed that most of the sampled beetle species prefer living in a forest (under rocks, fallen leaves and rotten wood). Beetles preferring open spaces were less abundant (only four per cent of the identified species).
4.4. Ecological Classification
When all data regarding beetle species occurring in Polish caves were taken into consideration, the corroboration in the presently studied caves of three troglobionts (Choleva lederiana gracilenta, Catops tristis infernus and Speonomus normandi hydrophilus) and four eutroglophiles (Quedius mesomelinus mesomelinus, Tachinus subterraneus, Omalium validum and Catops picipes) was an expected result.
Despite Ch. lederiana gracilenta and C. tristis infernus not presenting particular adaptations for living underground, both species were described from caves [69], and thus far, they have never been found in the epigean environment. In turn, S. normandi hydrophilus is a highly specialised subterranean species; however, it can still be found in caves and the MSS environment, such as scree and rock debris [46].
While Q. mesomelinus mesomelinus and T. subterraneus are commonly accepted as (eu)troglophiles species, C. picipes and O. validum are not so obviously classified. C. picipes is a common, widely distributed European species. It is found in both epigean and hypogean environments. However, its preference to occur in subterranean environments and high frequency in samples from various caves in Europe influenced considering this species as an (eu)troglophile [61,70].
Similar results concern O. validum, which occurs primarily in mountain regions and lives in burrows, rock debris and caves [29,71]. The species was found not only in Polish caves but also in many caves in Czechia [72], Germany [61], and Slovakia [8]. Moreover, it is also known for living in the riparian-sylvan microhabitat [73]. It was considered an (eu)troglophile by both Zaenker et al. [61] and Rendoš et al. [8]. In our investigation, we observed it every season through both years of the study. Burakowski [29] indicated that faint pigmentation, small eyes and more extended feet are an adaptation for subterranean living.
It is worth noting that among all collected species, more than 30 were indicated as subtroglophiles or MSS species, which is approximately 30% of all collected species. It is an interesting result because it proves that many species penetrating caves prefer this habitat. The attractiveness of the MSS environment was already pointed out [8]. On the other hand, more than 50% of collected beetle species were trogloxenes. However, a majority of them were sampled only in the entrance zone. This is not entirely unexpected, since the high ratio of trogloxene diversity (3.560852562–5.640653908 Shannon diversity index value) in the entrance zone was also reported in a temperate marble cave in the Sierra Nevada Range, California, USA [74]. Moreover, the occurrence of many trogloxenes was also observed in the entrance zones of caves in Slovakia [59], Romania [73] and Italy [75], where it was mainly attributed to the contact between subterranean and surface environments and a high accumulation of decomposed organic matter.
The presence of very few species that could be classified as troglobionts and eutroglophiles most probably resulted from the environmental conditions peculiar to caves of the Częstochowa Upland. The temperatures inside the studied caves were unstable and varied throughout the year. Temperature fluctuations were higher in the entrance zones (about several dozen degrees Celsius) than in the depths of the caves (several degrees Celsius). Similarly, relative humidity was inconstant, especially in the entrance zone. Moreover, in some caves, the direct or diffracted light reached even a half-length of the cave passages. Because species strictly associated with subterranean environments prefer stable environmental factors, such as temperature and humidity [76,77], they cannot find such conditions in the investigated caves.
Nevertheless, despite conditions that are not very suitable for dwelling in the analysed caves, two strict troglobionts, Choleva lederiana gracilenta and Catops tristis infernus, have populations living in the Pod Sokolą cave. This was a slightly unexpected situation, since some diffracted light reaches even the very end of this cave, and a minor fluctuation in air temperature occurs there. In this case, however, stable humidity was a crucial factor for the presence of both these cave-obligated beetles.
In contrast, the deepest terminal part of the Psia cave boasts a very stable temperature (the yearly fluctuation was less than 1 °C), high and stable humidity, and complete darkness. Despite these preferable conditions, this cave zone had outstandingly poor fauna (Tables S1 and S2). Within a two-year study period, a single beetle specimen was sampled in this zone of the Psia cave.
However, there was also a cave in which the climatic conditions appeared to be very dynamic throughout the year, but it still possessed its troglobiont beetle, namely, Speonomus normandi hydrophilus. Moreover, this troglobiont was the most abundant beetle in this cave, being extremely dominant (Table 3). It is worth mentioning that S. n. hydrophilus is alien to the fauna of Poland. Endemic to the Pyrenees, it is a beetle that was intentionally introduced to the Dzwonnica cave in 1982 [78]. Within 40 years, this species colonised nearby caves (i.e., the Towarna and Cabanowa caves) [79,80].
This case of colonisation success proves that, in Poland, the environmental conditions in caves are sufficient for habitation by troglobionts, but for some reason, are only barely adequate or inadequate for highly specialised dwelling species. As suggested by some authors [56,58,64], this phenomenon can be explained by the geological history of Central Europe, where, during the Pleistocene, glaciations influenced the entire fauna to such an extent that it could not even survive in the caves, which are normally refugial environments.
There is one more reason for this hypogean environment to be vastly attractive for many beetles. Though the epigean surroundings have their own environmental and climatic conditions, they are not as stable as those in the cave entrances. Additionally, as was suggested in many previous studies, organic matter from the external environment can easily flow into the caves via physical or biological means [4,9,81,82,83,84]. Most importantly, the entrance and transition zones of the studied caves exhibit conditions similar to the MSS environment to a greater extent than the deeper cave zones. The MSS, as a specific space [5,6], connects components originating from the underground and the surface environments. Hence, it seems to provide an additional explanation for why so many beetle species live in these caves, albeit being weakly associated with them ecologically.
5. Conclusions
- Many beetle species choose to dwell in caves, despite lacking specific adaptations for living in such environments.
- The entrance zone is the most attractive place for surface species to inhabit because the climatic conditions there are more stable than outside the cave (due to the presence of some light and high availability of organic matter).
- In the deeper parts of the studied caves, the number of occurring species rapidly decreased, though this zone sometimes had a stable environmental condition (namely, humidity).
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15030345/s1, Table S1. Numbers of sampled specimens in each studied cave. Table S2. A list of sampled species with their ecological classification. Table S3. Habitat and food preferences of beetles inhabiting caves in the Częstochowa Upland.
Author Contributions
Conceptualization, B.L. and J.K.-Z.; data curation, B.L.; formal analysis, J.K.-Z.; methodology, J.K.-Z.; resources, J.K.-Z.; software, J.K.-Z.; supervision, B.L.; visualization, J.K.-Z.; writing—original draft, J.K.-Z.; writing—review and editing, B.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The material was collected with permissions of the Regional Directorate for Environmental Protection (RDOŚ) in Katowice and Kraków. We want to thank the following persons for their help in species identification: the late H. Szołtys (Brynek, Poland), K. Komosiński (Department of Zoology, Faculty of Biology and Biotechnology, University of Warmia and Mazury in Olsztyn, Poland), M. Mazur (Institute of Biology, University of Opole, Opole, Poland), the late A. Melke (Independent Researcher), and P. Sienkiewicz (Department of Entomology and Environmental Protection, Poznań University of Life Sciences, Poland). Special thanks go to Jerzy A. Lis (Institute of Biology, University of Opole, Opole, Poland) for his comments on the first version of the manuscript and to three anonymous reviewers for their helpful comments in the preparation of its final improved version.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
The Shannon diversity index was calculated as:
where the following are defined:
1D = exp(H′) = exp(−Σ(pi log pi2)
- 1D—Shannon diversity;
- H′—the Shannon index;
- pi —the proportion (ni/N) of individuals of ith species found (ni) divided by the total number of individuals found (N).
The Simpson diversity was calculated as:
where the following are defined:
2D = 1/Σ pi2
- Σ pi2—the Simpson index;
- pi—number of specimens of species ith in the community.
The dominance was calculated by using the formula:
where the following are defined:
Di = ni/N × 100%
- Di—dominance index;
- ni—number of individuals in ith species;
- N—total number of individuals for all species.
References
- Stack, C. (Ed.) Beetles: Biodiversity, Ecology and Role in the Environment; Nova Science Publishers Inc.: New York, NY, USA, 2015; p. 262. [Google Scholar]
- Marshall, S.A. Beetles: The Natural History and Diversity of Coleoptera; Firefly Books Ltd.: Richmond Hill, ON, Canada, 2018; p. 784. [Google Scholar]
- Racovitza, E.G. Essai sur les problèmes biospéologiques. Arch. Zool. Exp. Gén. 1907, 4, 371–488. [Google Scholar]
- Culver, D.C. Cave Life. Evolution and Ecology; Harvard University Press: Cambridge, MA, USA, 1982; p. 189. [Google Scholar]
- Juberthie, C.; Delay, B.; Bouillon, M. Extension du milieu souterrain en zone non-calcaire: Description d’un nouveau milieu et de son peuplement par les Coléoptéres troglobies. Mém. Biospéol. 1980, 7, 19–52. [Google Scholar]
- Juberthie, C. Le milieu souterrain: Étendue et Composition. Mém. Biospéol. 1983, 10, 17–65. [Google Scholar]
- Pipan, T.; Culver, D.C. Shallow Subterranean Habitats. In Encyclopedia of Caves; White, W.B., Culver, D.C., Eds.; Academic Press: London, UK, 2012; pp. 886–895. [Google Scholar]
- Rendoš, M.; Mock, A.; Jászay, T. Spatial and temporal dynamics of invertebrates dwelling karstic mesovoid shallow substratum of Sivec National Nature Reserve (Slovakia), with emphasis on Coleoptera. Biologia 2012, 67, 1143–1151. [Google Scholar] [CrossRef] [Green Version]
- Nae, I.; Băncilă, R.I. Mesovoid shallow substratum as a biodiversity hotspot for conservation priorities: Analysis of oribatid mite (Acari: Oribatida) fauna. Acarologia 2017, 57, 855–868. [Google Scholar] [CrossRef]
- Ledesma, E.; Jiménez-Valverde, A.; Baquero, E.; Jordana, R.; de Castro, A.; Ortuño, V.M. Arthropod biodiversity patterns point to the Mesovoid Shallow Substratum (MSS) as a climate refugium. Zoology 2020, 141, 125771. [Google Scholar] [CrossRef] [PubMed]
- Skalski, A.W. Origin and composition of underground fauna in Poland. In Proceedings of the 8th International Congress of Speleology, Volumes I & II, Department of Geography and Geology, Western Kentucky University, Bowling Green, KY, USA, 18–24 July 1981. [Google Scholar]
- Kocot-Zalewska, J.; Domagała, P. Terrestrial invertebrate fauna of Polish caves—A summary of 100 years of research. Subterr. Biol. 2020, 33, 45–69. [Google Scholar] [CrossRef] [Green Version]
- Gradziński, M.; Kicińska, D. Caves in Poland. In Polish Caving 2013–2017, Proceedings of the 17th International Congress of Speleology, Sydney, Australia, 23–29 July 2017; Kicińska, D., Ed.; Pracownia Kreatywna Bezliku; Komisja Taternictwa Jaskiniowego Polskiego Związku Alpinizmu: Warszawa, Poland, 2017; pp. 4–6. [Google Scholar]
- Kocot-Zalewska, J.; Woźnica, A.J. Cave-dwelling heleomyzid flies (Diptera: Heleomyzidae) from the Polish caves. Historical overview and new data. Int. J. Speleol. 2021, 50, 203–211. [Google Scholar] [CrossRef]
- Jaskinie Polski. Available online: http://geoportal.pgi.gov.pl/jaskinie_polski (accessed on 27 April 2020).
- Szelerewicz, M.; Górny, A. Jaskinie Wyżyny Krakowsko-Wieluńskiej; Wydawnictwo PTTK Kraj: Warszawa-Kraków, Poland, 1986; p. 200. [Google Scholar]
- Zygmunt, J. Jaskinie Okolic Olsztyna; ZH-U Kontur: Częstochowa, Poland, 2013; p. 316. [Google Scholar]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Kobuta, Y.; Zelený, D.; Chiu, C.H.; Li, C.F.; Kosumuto, B.; Yasuhara, M.; Thorn, S.; Wei, C.L.; Costello, M.J.; et al. Quantifying sample completeness and comparing diversities among assemblages. Ecol. Res. 2020, 35, 292–314. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Kasprzak, K.; Niedbała, W. Wskaźniki biocenotyczne stosowane przy porządkowaniu i analizie danych w badaniach ilościowych. In Metody Stosowane w Zoologii Gleby; Górny, M., Grüm, L., Eds.; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1981; pp. 397–416. [Google Scholar]
- Biesiadka, E.; Kowalik, W. Water miters (Hydracarina) of the Western Bieszczady Mountains. I. Stagnant waters. Acta Hydrobiol. 1980, 22, 279–298. [Google Scholar]
- Szymczakowski, W. Klucze Do Oznaczania Owadów Polski. Część XIX: Chrząszcze—Coleoptera. Zeszyt 13: Catopidae; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1961; p. 69. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Biegaczowate—Carabidae, Część 1. Katalog Fauny Polski, XXIII (2); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1973; p. 430. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Biegaczowate—Carabidae, Część 2. Katalog Fauny Polski, XXIII (3); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1974; p. 430. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Adephaga prócz Carabidae, Myxophaga, Polyphaga: Hydrophiloidea. Katalog Fauny Polski, XXIII (4); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1976; p. 307. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Histeroidea i Staphylinoidea prócz Staphylinidae. Katalog Fauny Polski, XXIII (5); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1978; p. 356. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Kusakowate—Staphylinidae, Część 1. Katalog Fauny Polski, XXIII (6); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1979; p. 309. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Kusakowate—Staphylinidae, Część 2. Katalog Fauny Polski, XXIII (7); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1980; p. 271. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Kusakowate—Staphylinidae, Część 3: Aleocharinae. Katalog Fauny Polski, XXIII (8); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1981; p. 329. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Scarabaeoidea, Dascilloidea, Byrrhoidea i Parnoidea. Katalog Fauny Polski, XXIII (9); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1983; p. 394. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Buprestoidea, Elateroidea i Cantharoidea. Katalog Fauny Polski, XXIII (10); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1985; p. 400. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Cucujoidea, Część 1. Katalog Fauny Polski, XXIII (12); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1986; p. 265. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Cucujoidea, Część 2. Katalog Fauny Polski, XXIII (13); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1986; p. 277. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Cucujoidea, Część 3. Katalog Fauny Polski, XXIII (14); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1987; p. 308. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Cerambycidae i Bruchidae. Katalog Fauny Polski, XXIII (15); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1990; p. 311. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Ryjkowcowate prócz ryjkowców—Curculionioidea prócz Curculionidae. Katalog Fauny Polski, XXIII (18); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1992; p. 323. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Ryjkowce—Curculionidae, Część 1. Katalog Fauny Polski, XXIII (19); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1993; p. 304. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Ryjkowce—Curculionidae, Część 2. Katalog Fauny Polski, XXIII (20); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1995; p. 310. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Ryjkowce—Curculionidae, Część 3. Katalog Fauny Polski, XXIII (21); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1997; p. 307. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Uzupełnienia Tomów 2–21. Katalog Fauny Polski, XXIII (22); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 2000; p. 252. [Google Scholar]
- Koch, K. Die Käfer Mitteleuropas. Ökologie. Band 1; Goecke & Evers: Krefeld, Germany, 1989; p. 440. [Google Scholar]
- Koch, K. Die Käfer Mitteleuropas. Ökologie. Band 2; Goecke & Evers: Krefeld, Germany, 1989; p. 382. [Google Scholar]
- Koch, K. Die Käfer Mitteleuropas. Ökologie. Band 3; Goecke & Evers: Krefeld, Germany, 1992; p. 389. [Google Scholar]
- Crouau-Roy, B.; Crouau-Roy, Y.; Ferre, C. Dynamic and temporal structure of the troglobitic beetle Speonomus hydrophilus (Coleoptera: Bathysciinae). Ecography 1992, 15, 12–18. [Google Scholar] [CrossRef]
- Smoleński, M. Kusakowate (Coleoptera: Staphylinidae) borów bażynowych (Empetro-nigri Pinetum) Mierzei Łebskiej w Słowińskim Parku Narodowym. Wiad. Entomol. 2000, 18, 207–222. [Google Scholar]
- Päivinen, J.; Ahlroth, P.; Kaitala, V. Ant-associated beetles of Fennoscandia and Denmark. Entomol. Fenn. 2002, 13, 20–40. [Google Scholar] [CrossRef] [Green Version]
- Ruta, R.; Jałoszyński, P.; Konwerski, S. Nowe stanowiska gnilików (Coleoptera: Histeridae) w Polsce. Część 1. Onthophilinae —Dendrophilinae. Wiad. Entomol. 2004, 23, 13–20. [Google Scholar]
- Tréfás, H.; Van Lenteren, J. Egg-laying-site preferences of Pterostichus melanarius in mono- and intercrops. Bull. Insectology 2008, 61, 225–231. [Google Scholar]
- Shockley, F.W.; Tomaszewska, K.W.; McHugh, J.V. Review of the Natural History of the Handsome Fungus Beetles (Coleoptera: Cucujoidea: Endomychidae). Insecta Mundi 2009, 72, 1–24. [Google Scholar]
- Huruk, S.; Huruk, A. Zgrupowania biegaczowatych (Coleoptera: Carabidae) borów sosnowych oraz jodłowych w Górach Świętokrzyskich. Wiad. Entomol. 2010, 29, 247–258. [Google Scholar]
- Huruk, S.; Barševskis, A. Charakterystyka Biegaczowatych (Coleoptera, Carabidae) Gór Świętokrzyskich; Wydawnictwo Emporium: Kielce, Poland, 2014; p. 219. [Google Scholar]
- Szujecki, A. Kusakowate (Staphylinidae) Lasów Polski. Aspekt Różnorodności i Monitoringu Zooindykacyjnego; Lasy Państwowe: Warszawa, Poland, 2017; p. 150. [Google Scholar]
- Sket, B. Can we agree on an ecological classification of subterranean animals? J. Nat. Hist. 2008, 42, 1549–1563. [Google Scholar] [CrossRef]
- Kowalski, K. Fauna jaskiń Tatr Polskich. Ochr. Przyr. 1955, 23, 283–333. [Google Scholar]
- Skalski, A.W. Materiały do znajomości bezkręgowców jaskiń Wyżyny Krakowsko-Częstochowskiej. Roczn. Muz. Częstochowskiego 1973, 3, 161–200. [Google Scholar]
- Skalski, A.W. Charakterystyka fauny podziemnej Wyżyny Krakowsko-Częstochowskiej. Roczn. Muz. Okręg. Częstochowie 1981, 5, 51–60. [Google Scholar]
- Kováč, Ľ.; Mock, A.; Ľuptáčik, P.; Košel, V.; Fenďa, P.; Svatoň, J.; Mašán, P. Terrestrial Arthropods of the Domica Cave system and the Ardovská Cave (Slovak Karst)—Principal microhabitats and diversity. In Contributions to Soil Zoology in Central Europe, Proceedings of the 7th Central European Workshop on Soil Zoology, České Budějovice, Czech Republic, 14–16 April 2003; Tajovský, K., Schlaghamerský, J., Pižl, V., Eds.; Institute of Soil Biology, Academy of Sciences of the Czech Republic: České Budějovice, Czech Republic, 2005; pp. 61–70. [Google Scholar]
- Weber, D. Die Höhlenfauna Luxemburgs. Ferrantia 69; Musée National d’Histoire Naturelle: Luxembourg, 2013; p. 408. [Google Scholar]
- Zaenker, S.; Weber, D.; Weigand, A. Liste der cavernicolen Tierarten Deutschlands mit Einschluss der Grundwasserfauna (Version 1.9). Available online: https://www.hoehlentier.de/taxa.pdf (accessed on 27 April 2020).
- Arndt, W. Beitrag zur Kenntnis der Höhlenfauna. Ergebnis einer faunistischen Untersuchung der Höhlen Schlesiens. Zool. Anz. 1921, 52, 310–315. [Google Scholar]
- Arndt, W. Speläobiologische Untersuchungen in Schlesien. Speläologisch. Jahrb. 1923, 4, 95–114. [Google Scholar]
- Demel, K. Fauna jaskiń Ojcowskich. Sprawozd. Posiedz. Tow. Nauk. Warsz. Wydz. Nauk Mat.-Przyr. 1918, 11, 623–659. [Google Scholar]
- Pax, F.; Maschke, K. Höhlenfauna des Glatzer Schneeberges. Die rezente Metazoenfauna. Beitr. Biol. Glatzer Schneeberges 1935, 1, 4–72. [Google Scholar]
- Moldovan, E. Beetles. In Encyclopedia of Caves; White, W.B., Culver, D.C., Eds.; Academic Press: London, UK, 2012; pp. 96–107. [Google Scholar]
- Buse, J.; Alexander, K.N.A.; Ranius, T.; Assmann, T. Saproxylic Beetles—Their Role and Diversity in European Woodland and Tree Habitats. In Proceedings of the 5th Symposium and Workshop on the Conservation of Saproxylic Beetles, Lüneberg, Germany, 14–16 June 2008; Pensoft Publishers: Sofia, Bulgaria, 2009; p. 236. [Google Scholar]
- Plachter, H.; Plachter, J. Ökologische Studien zur terrestrischen Höhlenfauna Südduetschlands. Zoologica 1988, 139, 1–67. [Google Scholar]
- Szymczakowski, W. Catopidae (Coleoptera) des grottes dans les Sokole Góry près de Częstochowa. Acta Zool. Cracov. 1957, 1, 65–115. [Google Scholar]
- Nitzu, E. The Cholevinae of Romania (exclusive of Leptodirini) (Coleoptera, Leiodidae) with special reference to the hypogeal records. Zootaxa 2013, 3620, 351–378. [Google Scholar] [CrossRef] [PubMed]
- Mazur, A. Omalium validum KRAATZ, 1858 (Coleoptera, Staphylinidae) im Bielengebirge (poln. Góry Bialskie) und Glatzer Schneegebirge (poln. Masyw Śnieżnika). Przyr. Sudet. 2007, 10, 97–100. [Google Scholar]
- Mlejnek, R.; Hamet, A.; Růžička, J. Brouci (Coleoptera) v jeskyních a propastech České republiky: Beetles (Coleoptera) in caves and chasms of the Czech Republic. Acta Speleol. 2015, 6, 1–111. [Google Scholar]
- Nitzu, E. Microhabitats—Integrative environmental factors for species communities of Coleoptera in the karst landscape. Biologia 2021, 76, 1775–1783. [Google Scholar] [CrossRef]
- Tobin, B.W.; Hutchins, B.T.; Schwartz, B.F. Spatial and temporal changes in invertebrate assemblage structure from the entrance to deep-cave zone of a temperate marble cave. Int. J. Speleol. 2013, 42, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Do cave features affect underground habitat exploitation by non-troglobite species? Acta Oecol. 2014, 55, 29–35. [Google Scholar] [CrossRef]
- Killing-Heinze, M.; Pflitsch, A.; Furian, W.; Allison, S. The importance of air temperature as a key parameter to identify climatic processes inside Carlsbad Cavern, New Mexico, USA. J. Caves Karst Stud. 2017, 79, 153–167. [Google Scholar] [CrossRef]
- Mayoral, J.; Barranco, P.; Fernandez-Cortes, A. First biological data, associated fauna, and microclimate preferences of the enigmatic cave-dwelling beetle Dalyat mirabilis Mateu, 2002 (Coleoptera, Carabidae). Subterr. Biol. 2022, 44, 103–121. [Google Scholar] [CrossRef]
- Skalski, A.W. Experimental acclimatization of Speonomus hydrophilus (Jeannel 1907) (Coleoptera, Catopidae, Bathysciinae) in Poland. Mem. Biospeol. 1994, 21, 127–131. [Google Scholar]
- Kocot-Zalewska, J. Speonomus hydrophilus (Jeannel 1907) w Jaskini Towarnej. In Materiały 50 Sympozjum Speleologicznego, Proceedings of the 50th Speleological Symposium, Kielce, Poland, 20–23 October 2016; Urban, J., Ed.; Sekcja Speleologiczna Polskiego Towarzystwa Przyrodników im. M. Kopernika: Kraków, Poland, 2016; p. 124. [Google Scholar]
- Kocot-Zalewska, J.; Domagała, P.J.; Lis, B. Living in isolation for almost 40 years: Molecular divergence of the 28S rDNA and COI sequences between French and Polish populations of the cave beetle Speonomus normandi hydrophilus (Jeannel, 1907). Subterr. Biol. 2021, 37, 75–88. [Google Scholar] [CrossRef]
- Howarth, F.G. Ecology of cave arthropods. Ann. Rev. Entomol. 1983, 28, 365–389. [Google Scholar] [CrossRef]
- Poulson, T.L. Food sources. In Encyclopedia of Caves; Culver, D.C., White, W.B., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005; pp. 255–264. [Google Scholar]
- Culver, D.C.; Pipan, T. Shallow Subterranean Habitats: Ecology, Evolution, and Conservation; Oxford University Press: Oxford, UK, 2014; p. 258. [Google Scholar]
- Prous, X.; Ferreira, R.L.; Jacobi, C.M. The entrance as a complex ecotone in a Neotropical cave. Int. J. Speleol. 2015, 44, 177–189. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Location of the Częstochowska Upland in Poland; studied caves marked with numbers: 1—W Zielonej Górze cave; 2—Towarna cave; 3—Pod Sokolą cave; 4—Kroczycka cave; 5—Psia cave; 6—Zegar cave.
Figure 1.
Location of the Częstochowska Upland in Poland; studied caves marked with numbers: 1—W Zielonej Górze cave; 2—Towarna cave; 3—Pod Sokolą cave; 4—Kroczycka cave; 5—Psia cave; 6—Zegar cave.

Figure 2.
Entrances of studied caves. (A)—W Zielonej Górze cave; (B)—Towarna cave; (C)—Pod Sokolą cave; (D)—Kroczycka cave; (E)—Psia cave; (F)—Zegar cave.
Figure 2.
Entrances of studied caves. (A)—W Zielonej Górze cave; (B)—Towarna cave; (C)—Pod Sokolą cave; (D)—Kroczycka cave; (E)—Psia cave; (F)—Zegar cave.

Figure 3.
Diagrams of the studied caves with marked sampling points (for explanations, see the text). (A)—W Zielonej Górze cave; (B)—Towarna cave; (C)—Psia cave; (D)—Zegar cave.
Figure 3.
Diagrams of the studied caves with marked sampling points (for explanations, see the text). (A)—W Zielonej Górze cave; (B)—Towarna cave; (C)—Psia cave; (D)—Zegar cave.

Figure 4.
Diagrams of the studied caves with marked sampling points (for explanations, see the text). (A)—Pod Sokolą cave; (B)—Kroczycka cave.
Figure 4.
Diagrams of the studied caves with marked sampling points (for explanations, see the text). (A)—Pod Sokolą cave; (B)—Kroczycka cave.

Figure 5.
Distribution of beetles inside the caves; richness in particular parts of the caves is shown for all beetle families.
Figure 5.
Distribution of beetles inside the caves; richness in particular parts of the caves is shown for all beetle families.

Figure 6.
Distribution of beetles inside the caves; richness in particular parts of the caves is shown for beetle families, except the three most numerous (Staphylinidae, Leiodidae, Cryptophagidae).
Figure 6.
Distribution of beetles inside the caves; richness in particular parts of the caves is shown for beetle families, except the three most numerous (Staphylinidae, Leiodidae, Cryptophagidae).

Figure 7.
Temperature fluctuations at the measurement points in the studied caves.

Figure 8.
The relative humidity fluctuations at the measurement points in the studied caves.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Number of species recorded in particular parts of studied caves. S0–S5—sampling sites.
| Cave | S0 | S1 | S2 | S3 | S4 | S5 |
|---|---|---|---|---|---|---|
| W Zielonej Górze | 22 | 34 | 11 | 7 | ---- | ---- |
| Towarna | 53 | 24 | 9 | 8 | ---- | ---- |
| Pod Sokolą | 13 | 35 | 11 | 5 | 3 | ---- |
| Kroczycka | 17 | 10 | ---- | 5 | 11 | 7 |
| Psia | 20 | 10 | 8 | 1 | ---- | ---- |
| Zegar | 23 | 33 | 8 | 11 | ---- | ---- |
Table 2.
Number of species collected in the studied caves and the value of the species richness indices.
Table 2.
Number of species collected in the studied caves and the value of the species richness indices.
| Cave | Number of Species | Shannon Diversity Index | Simpson Diversity Index |
|---|---|---|---|
| W Zielonej Górze | 38 | 7.614086359 | 1.375137514 |
| Towarna | 27 | 1.447734615 | 7.692307692 |
| Pod Sokolą | 42 | 5.053090317 | 1.497005988 |
| Psia | 17 | 8.758284041 | 2.949852507 |
| Zegar | 37 | 15.95863401 | 1.051524711 |
| Kroczycka | 21 | 8.846306259 | 1.112718371 |
Table 3.
Dominant species in the studied caves.
| Cave | Eudominant | Dominant |
|---|---|---|
| W Zielonej Górze | Cryptophagus distinguendus J. Sturm, 1845 (25.76%) Cryptophagus punctipennis Ch.N.F. Brisout de Barneville, 1863 (28.46%) Quedius mesomelinus mesomelinus Marsham, 1802 (12.30%) | |
| Towarna | Speonomus normandi hydrophilus (Jeannel, 1907) (92.86%) | |
| Pod Sokolą | Choleva lederiana gracilenta Szymczakowski, 1957 (55.6%) Catops picipes (Fabricius, 1787) (10.97%) | Catops tristis infernus Szymczakowski, 1957 (9%). |
| Kroczycka | Quedius mesomelinus mesomelinus Marsham, 1802 (22.2%) Othius subuliformis Stephens, 1833 (16.65%) Cryptophagus distinguendus J. Sturm, 1845 (12.5%). | Choleva paskoviensis E. Reitter, 1913 (8.3%) Proteinus brachypterus (Fabricius, 1792) (6.9%). |
| Psia | Omalium caesum Gravenhorst, 1806 (23.8%) Choleva cisteloides (Frölich, 1799) (14.28%). | |
| Zegar | Catops picipes (Fabricius, 1787) (13.56%) Omalium validum Kraatz, 1857 (10.17%) | Omalium caesum Gravenhorst, 1806 (8.47%), Choleva glauca Britten, 1918 (5.1%) Lesteva longoelytrata (Goeze, 1777) (5.1%) Quedius mesomelinus mesolmelinus Marsham, 1802 (5.1%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kocot-Zalewska, J.; Lis, B. Species Composition and Structure of Beetle Associations in Caves of the Częstochowa Upland, Poland. Diversity 2023, 15, 345. https://doi.org/10.3390/d15030345
AMA Style
Kocot-Zalewska J, Lis B. Species Composition and Structure of Beetle Associations in Caves of the Częstochowa Upland, Poland. Diversity. 2023; 15(3):345. https://doi.org/10.3390/d15030345
Chicago/Turabian StyleKocot-Zalewska, Joanna, and Barbara Lis. 2023. "Species Composition and Structure of Beetle Associations in Caves of the Częstochowa Upland, Poland" Diversity 15, no. 3: 345. https://doi.org/10.3390/d15030345
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.