

New Records of the Alien Chinese Ricefish (Oryzias sinensis) and Its Dispersal History across Eurasia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods

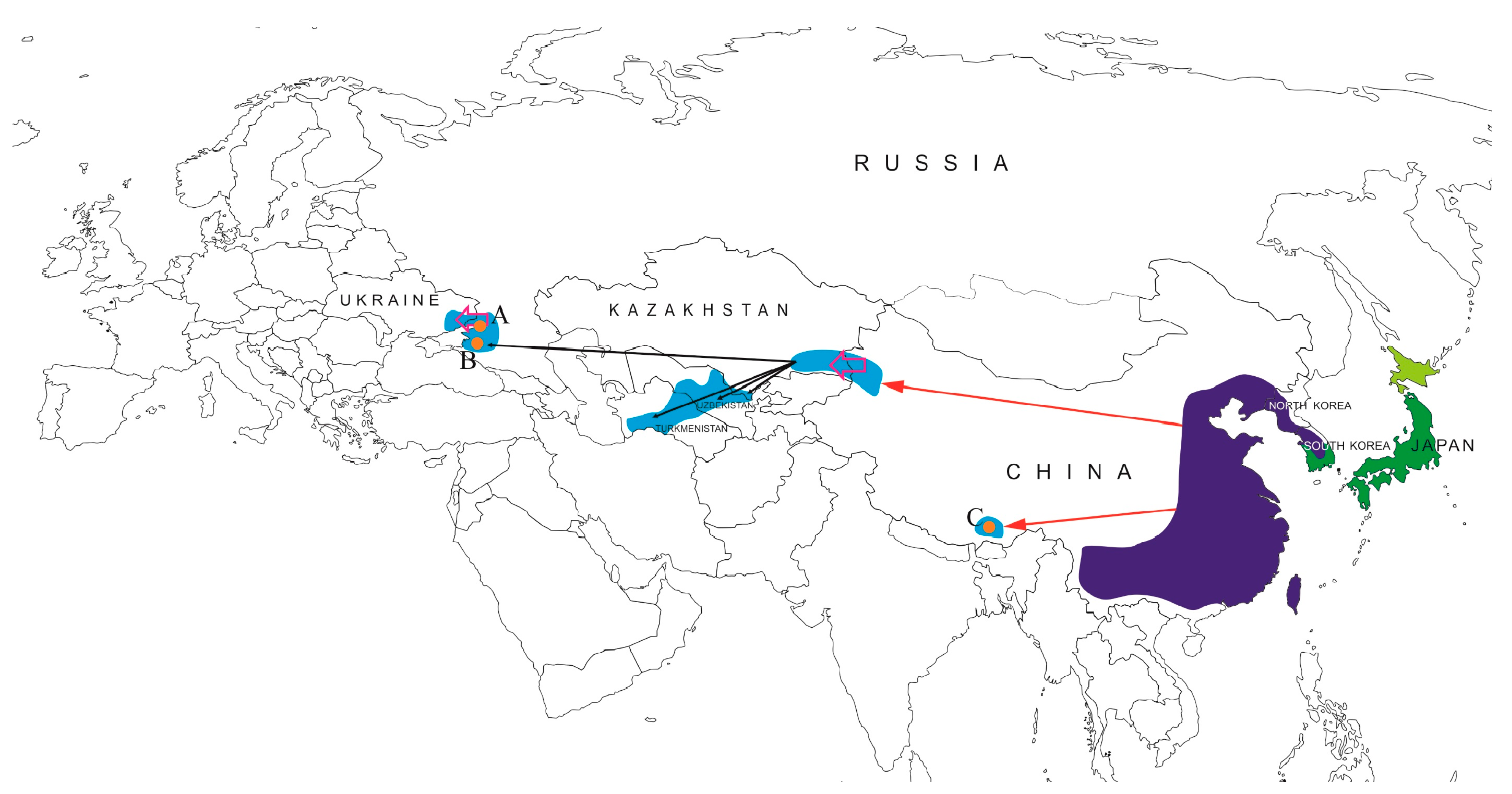{kind=link}
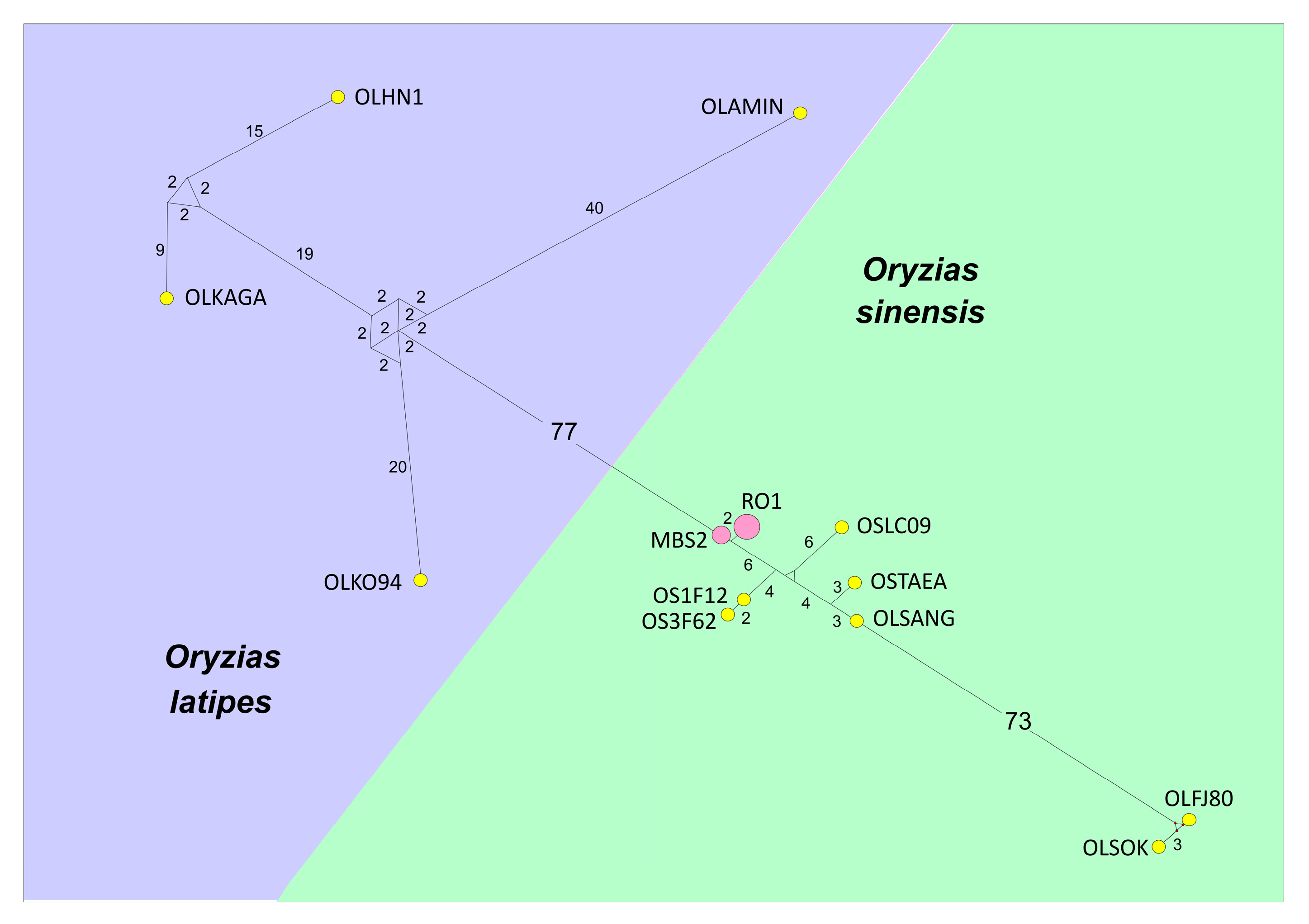{kind=link}
| Designation on Figure 2 | GenBank ID | Species | Locality | Source |
|---|---|---|---|---|
| OS3F62 | HQ536423.1 | O. sinensis | Korea | [50] |
| OS1F128 | HQ536422.1 | O. sinensis | Korea | [50] |
| OSTaean | GU013788.1 | O. sinensis | Korea: the Dongjin River | [40] |
| OLSangh | AP008948.1 | O. latipes (O. sinensis following Yoon et al., 2011) | China: Shanghai | [51] |
| OSLC09 | LC153109.1 | O. sinensis | Japan: Aichi, Higashiyama Zoo | Suzuki-Matsubara M., Murase Y., Moriyama A., unpublished |
| OLFJ80 | FJ197680.1 | O. latipes | not specified | Park J.-Y., An H.-S., Cho Y.-A., Kim K.-K., unpublished |
| OLSOK | AP008947.1 | O. latipes (O. sinensis following Yoon et al., 2011) | inbred strain, derived from eastern Korea | [51] |
| OLKaga | AP008940.1 | O. latipes | Japan: Kaga | [51] |
| OLKO94 | LC335803.1 | O. latipes | Japan: Kanagava | [52] |
| OLAmino | AP008944.1 | O. latipes | Japan: Amino | [51] |
| OLHN1 | AB498066.1 | O. latipes | Japan: HNI | [53] |
| RO1 | MW695407 | O. sinensis | Russia: Rostov province | This paper |
| RO1 | MW695408 | O. sinensis | Russia: Krasnodar Territory | This paper |
| MBS2 | MW695409 | O. sinensis | China, TibetanAutonomous Region | This paper |
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pimm, S.L.; Raven, P. Extinction by numbers. Nature 2000, 403, 843–845. [Google Scholar] [CrossRef]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef]
- Seebens, H.; Bacher, S.; Blackburn, T.M.; Capinha, C.; Dawson, W.; Dullinger, S.; Genovesi, P.; Hulme, P.E.; van Kleunen, M.; Kühn, I.; et al. Projecting the continental accumulation of alien species through to 2050. Glob. Change Biol. 2021, 27, 970–982. [Google Scholar] [CrossRef]
- Pimentel, D. Biological Invasions. Economic and Environmental Costs of Alien Plant, Animal, and Microbe Species, 2nd ed.; CRC Press: Boca Raton, CA, USA; London, UK; New York, NY, USA, 2011. [Google Scholar]
- Ricciardi, A.; Hoopes, M.F.; Marchetti, M.P.; Lockwood, J.L. Progress toward understanding the ecological impacts of nonnative species. Ecol. Monogr. 2013, 83, 263–282. [Google Scholar] [CrossRef] [Green Version]
- Douglas, M.R.; Slynko, Y.V.; Dgebuadze, Y.Y.; Olenin, S.; Aleksandrov, B.; Boltachev, A.; Slynko, E.E.; Khristenko, D.; Minchin, D.; Pavlov, D.F.; et al. Invasion Ecology: An International Perspective Centered in the Holarctic. Fisheries 2015, 40, 464–470. [Google Scholar] [CrossRef]
- Jacobs, L.E.O.; Richardson, D.M.; Lepschi, B.J.; Wilson, J.R.U. Quantifying errors and omissions in alien species lists: The introduction status of Melaleuca species in South Africa as a case study. NeoBiota 2017, 32, 89–105. [Google Scholar] [CrossRef] [Green Version]
- Leuven, R.S.E.W.; Boggero, A.; Bakker, E.S.; Elgin, A.K.; Verreycken, H. Invasive species in inland waters: From early detection to innovative management approaches. Aquat. Invasions 2017, 12, 269–273. [Google Scholar] [CrossRef]
- Zhivoglyadova, L.A.; Revkov, N.K. Invasion of the bivalve Corbicula fluminea (O.F. Müller, 1774) (Bivalvia: Cyrenidae) into the lower Don basin (in Russian, English summary). Aquat. Bioresour. Environ. 2018, 1, 44–50. [Google Scholar]
- Borovikova, E.A.; Alekseeva, J.A.; Bagirov, N.E.; Makhrov, A.A.; Popov, I.Y. Genetic identification of a crayfish (Astacus) species at the northern edge of their distribution area (Solovetsky Islands, White Sea). Biochem. Syst. Ecol. 2016, 65, 205–208. [Google Scholar] [CrossRef]
- Jussila, J.; Maguire, I.; Kokko, H.; Tiitinen, V.; Makkonen, J. Narrow-clawed crayfish in Finland: Aphanomyces astaci resistance and genetic relationship to other selected European and Asian populations. Knowl. Manag. Aquat. Ecosyst. 2020, 421, 30. [Google Scholar] [CrossRef]
- Berezina, N.A.; Terentiev, P.M.; Sharov, A.N.; Maximov, A.A. New records and disappearance from old sites of narrow-clawed crayfish Pontastacus leptodactylus in northwestern Russia. BioInvasions Rec. 2021, 10, 894–903. [Google Scholar] [CrossRef]
- Pryanichnikova, E.G.; Voroshilova, I.S.; Sabitova, R.Z. Introduction of Corbicula fluminea (Müller, 1774) (Mollusca: Bivalvia: Corbiculidae) in the Volga River Basin. Inland Water Biol. 2019, 12, S95–S97. [Google Scholar] [CrossRef]
- Karaouzas, I.; Zogaris, S.; Froufe, E.; Lopes-Lima, M. Rival at the gate: First record of the Asian clam Corbicula fluminea Müller, 1774 (Bivalvia: Corbiculidae) in Greece. Knowl. Manag. Aquat. Ecosyst. 2020, 421, 24. [Google Scholar] [CrossRef]
- Kondakov, A.V.; Bespalaya, Y.V.; Vikhrev, I.V.; Konopleva, E.S.; Gofarov, M.Y.; Tomilova, A.A.; Vinarski, M.V.; Bolotov, I.N. The Asian pond mussels rapidly colonize Russia: Successful invasions of two cryptic species to the Volga and Ob rivers. BioInvasions Rec. 2020, 9, 504–518. [Google Scholar] [CrossRef]
- Bruni, G.; Mirabella, I.; Domeneghetti, D.; Fasola, M.; Bellati, A. Will there be a second extinction? Molecular identification of multiple alien water frogs (Pelophylax ridibundus sensu lato) in Tuscany, Central Italy, reveals genetic pollution within a unique hybridogenetic system. Herpetol. J. 2020, 30, 147–158. [Google Scholar] [CrossRef]
- Reshetnikov, A.N.; Chestnut, T.; Brunner, J.L.; Charles, K.; Nebergall, E.E.; Olson, D.H. Detection of the emerging amphibian pathogens Batrachochytrium dendrobatidis and ranavirus in Russia. Dis. Aquat. Org. 2014, 110, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Chiesa, S.; Scalici, M.; Lucentini, L.; Marzano, F.N. Molecular identification of an alien temnocephalan crayfish parasite in Italian freshwaters. Aquat. Invasions 2015, 10, 209–216. [Google Scholar] [CrossRef]
- Parenti, L.R. A phylogenetic analysis and taxonomic revision of ricefishes, Oryzias and relatives (Beloniformes, Adrianichthyidae). Zool. J. Linn. Soc. 2008, 154, 494–610. [Google Scholar] [CrossRef] [Green Version]
- Hilgers, L.; Schwarzer, J. The untapped potential of ricefish and its wild relatives. eLife 2019, 8, e46994. [Google Scholar] [CrossRef]
- Karpov, V.E. Oryzias latipes (Temminck et Schlegel)—Ricefish (in Russian). In Fishes of Kazakhstan. V. 5.; Gvozdev, E.V., Mitrofanov, V.P., Eds.; Gylym: Alma-Ata, Kazakhstan, 1992; pp. 231–241. [Google Scholar]
- Sokolovskij, V.R.; Strelnikov, A.S.; Tereshchenko, V.G.; Timirhanov, S.R. Reaction of the fish assemblage of lakes with an endemic ichthyofauna on acclimatization new fish species (in Russian). Probl. Fish. 2008, 9, 34–48. [Google Scholar]
- Mamilov, N.; Sharakhmetov, S.; Amirbekova, F.; Bekkozhayeva, D.; Sapargaliyeva, N.; Kegenova, G.; Tanybayeva, A.; Abilkasimov, K. Past, Current and Future of Fish Diversity in the Alakol Lakes (Central Asia: Kazakhstan). Diversity 2022, 14, 11. [Google Scholar] [CrossRef]
- Yuldashov, M.A. Introduction of Alien Fish Species to Waterbodies of Uzbekistan. Int. J. Sci. Res. 2018, 7, 1213–1219. [Google Scholar]
- Shakirova, F.M. Present-Day Condition of Ichthyofauna in Reservoirs of Turkmenistan. Hdb. Env. Chem. 2014, 28, 233–260. [Google Scholar]
- Tsunikova, E.P.; Poznyak, V.G. Ricefish in irrigation water bodies of Krasnodar Territory (in Russian). In Fauna and Ecology of Animals under Conditions of Irrigated Lands; Kukish, A.I., Ed.; Kalmykian State University: Elista, Russia, 1990; pp. 89–91. [Google Scholar]
- Naseka, A.M.; Diripasko, O.A. New Invasive Fish Species in Freshwater Fish Fauna in the Northern Coastal Region of the Sea of Azov (in Russian). Vestnik zoologii 2005, 39, 89–94. [Google Scholar]
- Ishikawa, T.; Tachihara, K. Introduction history of non-native freshwater fish in Okinawa-jima Island: Ornamental aquarium fish pose the greatest risk for future invasions. Ichthyol. Res. 2014, 61, 17–26. [Google Scholar] [CrossRef]
- Fan, L.-Q.; Zhang, X.-J.; Pan, G. Length–weight and length–length relationships for nine fish species from Lhasa River Basin, Tibet, China. J. Appl. Ichthyol. 2015, 31, 807–808. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, Z. Unveiling the status of alien animals in the arid zone of Asia. PeerJ 2016, 4, e1545. [Google Scholar] [CrossRef] [Green Version]
- Roberts, E.R. Systematic observations on tropical Asian medakas or ricefishes of the genus Oryzias, with descriptions of four new species. Ichthyol. Res. 1998, 45, 213–224. [Google Scholar] [CrossRef]
- Kim, I.-S.; Park, J.-Y. Freshwater Fishes of Korea (in Korean); Kyo-Hak Publishing Co. Ltd.: Seoul, Republic of Korea, 2002. [Google Scholar]
- Zhang, C.-g.; Zhao, Y.-h. Species Diversity and Distribution of Inland Fishes in China (in Chinese); Science Press: Beijing, China, 2016. [Google Scholar]
- Asai, T.; Senou, H.; Hosoya, K. Oryzias sakaizumii, a new ricefish from northern Japan (Teleostei: Adrianichthyidae). Ichthyol. Explor. Freshwaters. 2011, 22, 289–299. [Google Scholar]
- Chen, Y.-R.; Uwa, H.; Chu, X.-L. Taxonomy and distribution of the genus Oryzias in Yunnan, China (Cyprinodontiformes, Oryziidae) (in Chinese, summary in English). Acta Zootaxonomica Sinica 1989, 14, 239–246. [Google Scholar]
- Park, M.-K.; Kim, I.-S.; Park, J.-Y. Morphological Characters and Geographical Distributions of Korean Ricefish (Pisces, Adrianichthyidae), Oryzias. Korean J. Ichthyol. 2006, 18, 163–169. [Google Scholar]
- Kim, I.-S.; Kim, S.-Y. Interspecific hybridization between Oryzias sinensis and O. latipes from Korea (in Korean, summary in English). Korean J. Ichthyol. 1993, 5, 113–121. [Google Scholar]
- Sakaizumi, M.; Shimizu, Y.; Hamaguchi, S. Electrophoretic studies of meiotic segregation in inter- and intraspecific hybrids among east Asian species of the genus Oryzias (Pisces: Oryziatidae). J. Exp. Zool. 1992, 264, 85–92. [Google Scholar] [CrossRef]
- Min, M.-S. Genetic variation and speciation of 2 species of the genus Oryzias (Pisces, Adrianichthyidae) in Korea (in Korean). Korean J. Syst. Zool. 1997, 13, 9–20. [Google Scholar]
- Yoon, M.; Kim, K.-Y.; Bang, I.-C.; Nam, Y.K.; Kim, D.S. Complete mitogenome sequence of the Chinese ricefish Oryzias sinensis (Teleostei: Beloniformes) and its phylogenetic analysis. Genes Genom. 2011, 33, 307–312. [Google Scholar] [CrossRef]
- In, D.-S.; Choi, E.-S.; Yoon, J.-D.; Kim, J.-H.; Min, J.-I.; Baek, S.-H.; Jang, M.-H. Cytochrome oxidase subunit I (COI) DNA sequence divergence between two cryptic species of Oryzias in South Korea. J. Ecol. Environ. 2013, 36, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.H.; Ratnasingham, S.; deWaard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. B 2003, 270 (Suppl. S1), S96–S99. [Google Scholar] [CrossRef] [Green Version]
- Hirai, N.; Torii, Y.; Matsuoka, H.; Ishii, M. Genetic diversity and intrusion of alien populations of Oryzias latipes in Osaka Prefecture, central Japan. Jpn. J. Environ. Entomol. Zool. 2017, 28, 47–54. [Google Scholar]
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Kottelat: Cornol, Switzerland; Freyhof: Berlin, Germany, 2007. [Google Scholar]
- Chikhachev, A.S.; Egorov, A.V. Species composition of ichthyofauna in water bodies of Rostov Region (in Russian). Izvestiya Vuzov. North-Caucasian Region. Nat. Sci. 2009, 6, 73–78. [Google Scholar]
- Vasilyeva, E.D.; Luzhnyak, V.A. Fishes of the Azov Sea Basin (in Russian); Publishing house YNTs RAS: Rostov-on-Don, Russia, 2013. [Google Scholar]
- Audira, G.; Siregar, P.; Chen, K.H.-C.; Roldan, M.J.M.; Huang, J.-C.; Lai, H.-T.; Hsiao, C.-D. Interspecies Behavioral Variability of Medaka Fish Assessed by Comparative Phenomics. Int. J. Mol. Sci. 2021, 22, 5686. [Google Scholar] [CrossRef]
- Artamonova, V.S.; Kolmakova, O.V.; Kirillova, E.A.; Makhrov, A.A. Phylogeny of salmonoid fishes (Salmonoidei) based on mtDNA COI gene sequences (barcoding). Contemp. Probl. Ecol. 2018, 11, 271–285. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA barcoding Australia’s fish species. Phil. Trans. R. Soc. B 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Kim, S.; Koo, H.; Kim, J.-H.; Jung, J.-W.; Hwang, S.Y.; Kim, W. DNA chip for species identification of Korean freshwater fish: A case study. BioChip J. 2011, 5, 72–77. [Google Scholar] [CrossRef]
- Hirayama, M.; Mukai, T.; Miya, M.; Murata, Y.; Sekiya, Y.; Yamashita, T.; Nishida, M.; Watabe, S.; Oda, S.; Mitani, H. Intraspecific variation in the mitochondrial genome among local populations of Ricefish Oryzias latipes. Gene 2010, 457, 13–24. [Google Scholar] [CrossRef]
- Katsumura, T.; Oda, S.; Mitani, H.; Oota, H. Ricefish Population Genome Structure and Demographic History Described via Genotyping-by-Sequencing. G3 (Genes Genomes Genetics) 2019, 9, 217–228. [Google Scholar] [PubMed] [Green Version]
- Setiamarga, D.H.E.; Miya, M.; Yamanoue, Y.; Azuma, Y.; Inoue, J.G.; Ishiguro, N.B.; Mabuchi, K.; Nishida, M. Divergence time of the two regional ricefish populations in Japan as a new time scale for comparative genomics of vertebrates. Biol. Lett. 2009, 5, 812–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Wu, C. The Fishes of the Qinghai-Xizang Plateau (in Chinese); Sichuan Publishing House of Science & Technology: Chengdu, China, 1992. [Google Scholar]
- Zhang, C.; Cai, B.; Xu, T. Fishes and Fish Resources in Xizang, China (in Chinese); China Agriculture Press: Beijing, China, 1995. [Google Scholar]
- Makhrov, A.A.; Artamonova, V.S.; Karabanov, D.P. Finding of topmouth gudgeon Pseudorasbora parva (Temminck et Schlegel) (Actinopterygii: Cyprinidae) in the Brahmaputra river basin (Tibetan plateau, China). Russ. J. Biol. Invasions 2013, 4, 174–179. [Google Scholar] [CrossRef]
- Adbildaev, M.A.; Dubitsky, A.M. The occurrence in the Ili River basin of fish identified as Aplocheilus sp., a species new to the USSR fauna (in Russian). Voprosy Ikhtiologii 1974, 4, 328–330. [Google Scholar]
- Zarechnaya, S.N.; Makhanko, E.V.; Rechkalova, N.I.; Vitlin, L.M.; Zhulev, A.I.; Shumeyko, V.N.; Erlikh, V.D. Experience of introduction of Aplocheilus latipes (Temmick and Schlegel) in Krasnodar Territory and Azerbaijan (in Russian). Med. Parasitol. Parasit. Dis. 1980, 49, 15–18. [Google Scholar]
- Sal’nikov, V.B. Antropogenic migration of fish in Turkmenistan. J. Ichthyol. 1998, 38, 591–602. [Google Scholar]
- Karabanov, D.P.; Kodukhova, Y.V.; Pashkov, A.N.; Reshetnikov, A.N.; Makhrov, A.A. “Journey to the West”: Three phylogenetic lineages contributed to the invasion of stone moroko, Pseudorasbora parva (Actinopterygii: Cyprinidae). Russ. J. Biol. Invasions 2021, 12, 67–78. [Google Scholar] [CrossRef]
- Li, S.; Chen, J.; Wang, X.; Copp, G.H. Invasiveness screening of non-native fishes for the middle reach of the Yarlung Zangbo River, Tibetan Plateau, China. River Res. Applic. 2017, 33, 1439–1444. [Google Scholar] [CrossRef]


| Locality | Coordinates | Date | N | SL, mm |
|---|---|---|---|---|
| Rostov Region, Russia (Don River basin) | N 47°5′43.2″; E 39°19′1.5″ | 1.05.2019 | 10 (2) | 17.0–23.3 (19.8) |
| Krasnodar Territory, Russia (Kuban River basin) | N 45°14′7.4″; E 37°59′52″ | 11.05.2019 | 8 (2) | 18.5–25.5 (22.13) |
| Tibetan Autonomous Region, China (Yarlung Tsangpo (Brahmaputra) basin) | N 29°21′24″; E 90°42′55″ | 27.09.2012 | 1 (2) | 26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makhrov, A.A.; Artamonova, V.S.; Sun, Y.-H.; Fang, Y.; Pashkov, A.N.; Reshetnikov, A.N. New Records of the Alien Chinese Ricefish (Oryzias sinensis) and Its Dispersal History across Eurasia. Diversity 2023, 15, 317. https://doi.org/10.3390/d15030317
Makhrov AA, Artamonova VS, Sun Y-H, Fang Y, Pashkov AN, Reshetnikov AN. New Records of the Alien Chinese Ricefish (Oryzias sinensis) and Its Dispersal History across Eurasia. Diversity. 2023; 15(3):317. https://doi.org/10.3390/d15030317
Chicago/Turabian StyleMakhrov, Alexander A., Valentina S. Artamonova, Yue-Hua Sun, Yun Fang, Andrey N. Pashkov, and Andrey N. Reshetnikov. 2023. "New Records of the Alien Chinese Ricefish (Oryzias sinensis) and Its Dispersal History across Eurasia" Diversity 15, no. 3: 317. https://doi.org/10.3390/d15030317