

‘Unfinished’ Morphogenesis Hides Different Speciation Pathways in Charophytes: Evidence from the 190-Year-Old Original Material of Chara denudata (Charales, Charophyceae)

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Study
Studied Specimens
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Phylogenetic Analyses
3. Results
3.1. Description of the Specimens of Chara denudata
3.2. Description of the Specimens of Chara dissoluta
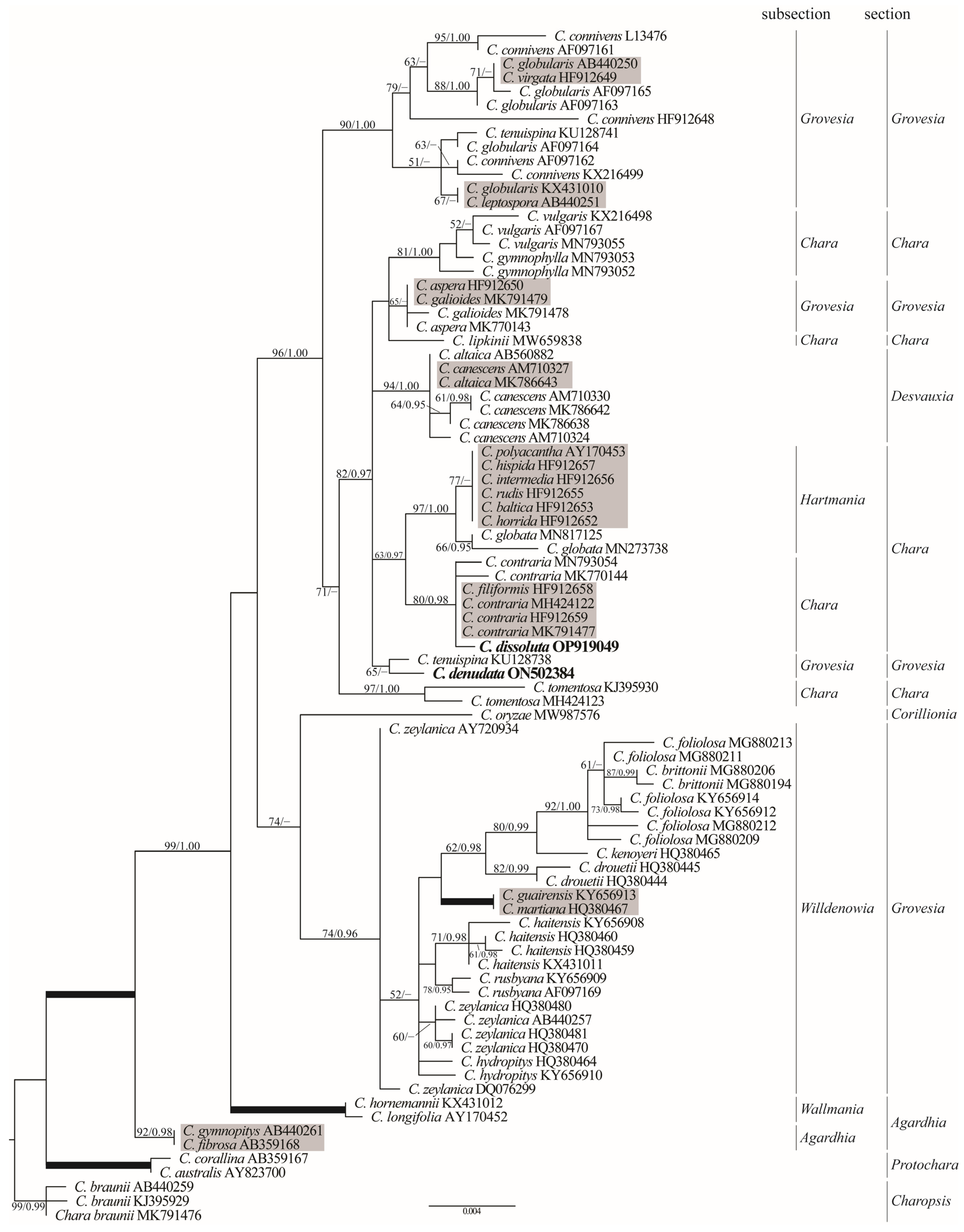
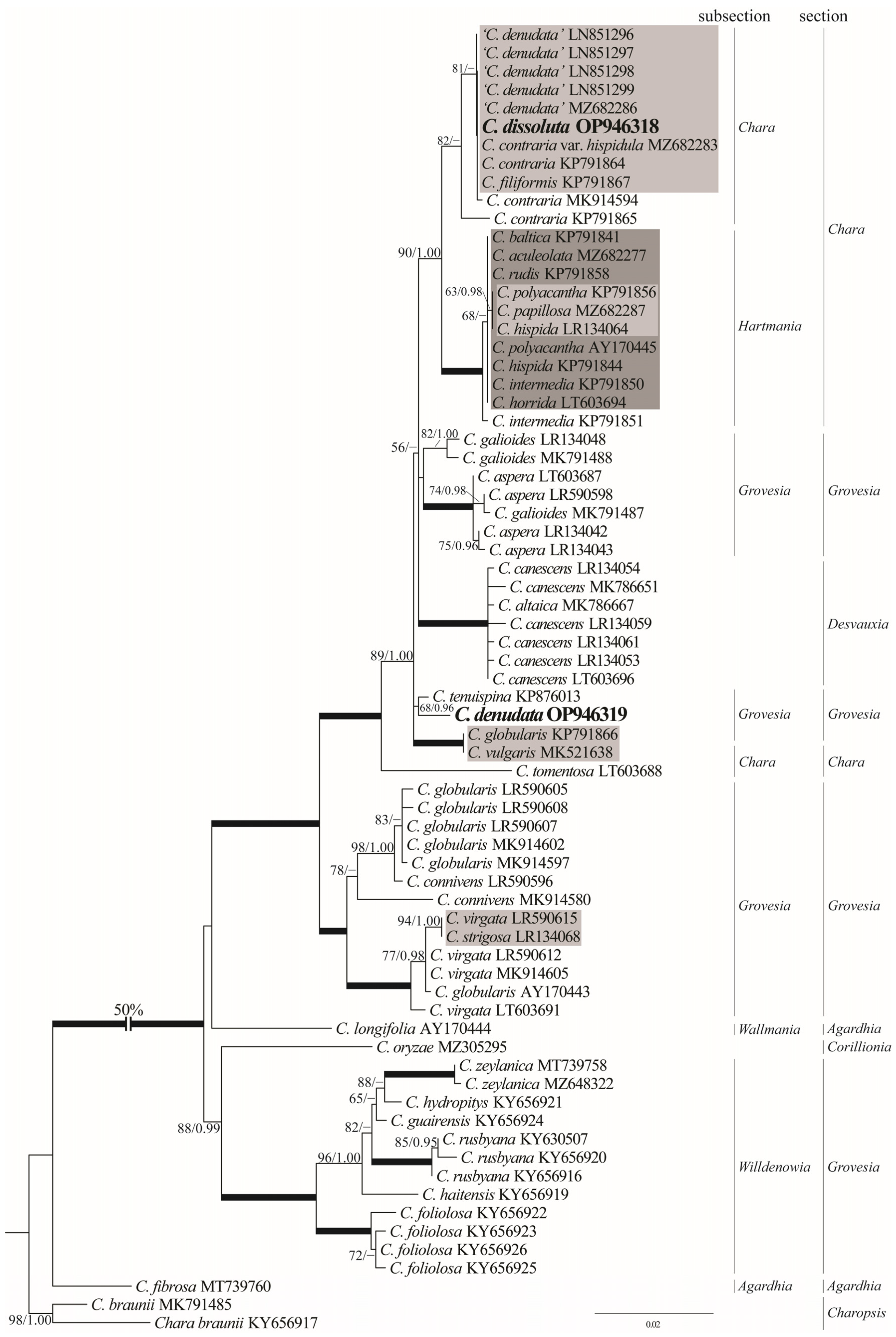
3.3. Phylogenetic Positions
4. Discussion
4.1. Morphological Perspective
4.2. Phylogenetic Perspective
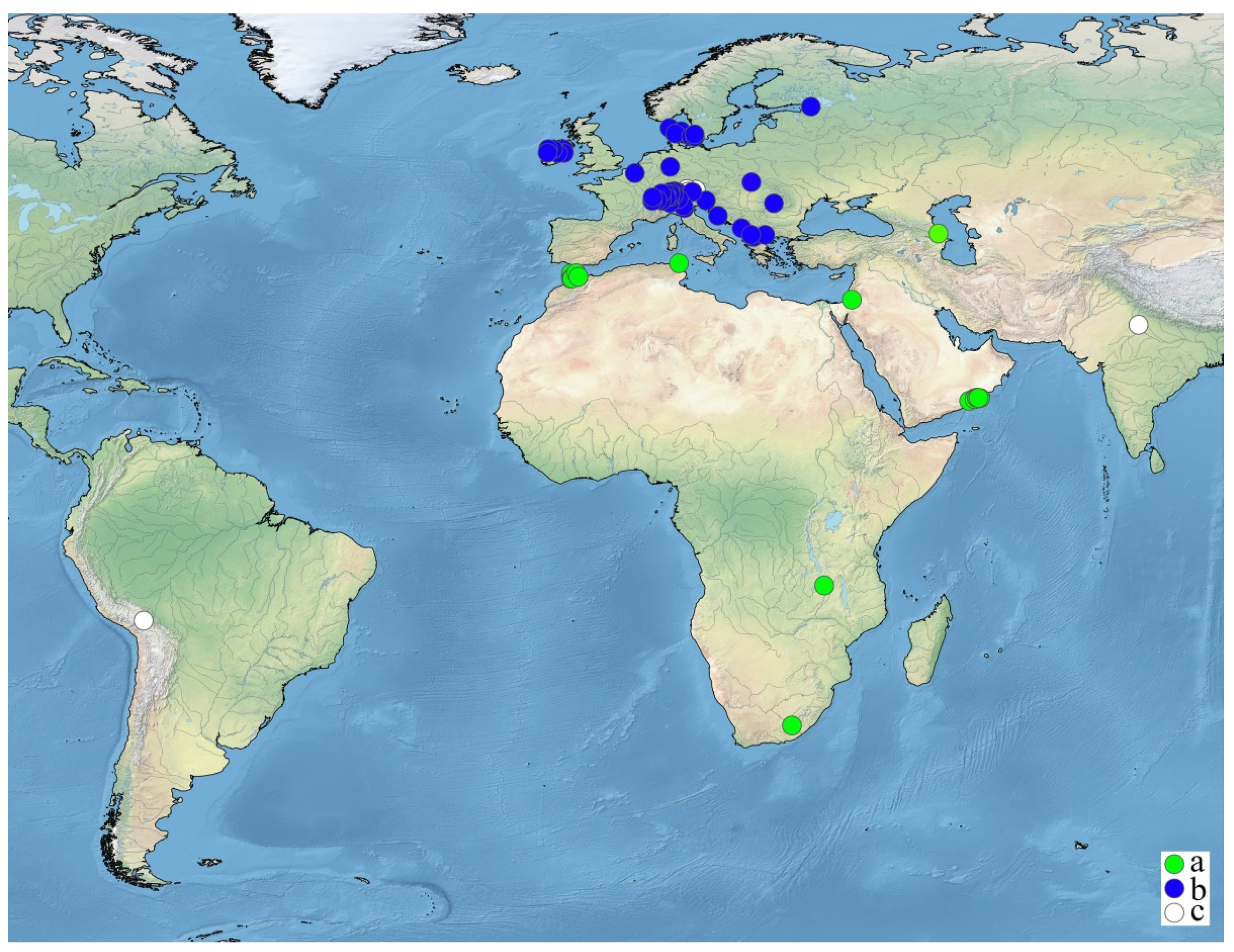
4.3. Ecological Perspective
4.4. Distribution
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wood, R.D.; Imahori, K. Iconograph of the Characeae. In Monograph of the Characeae; Wood, R.D., Imahori, K., Eds.; Verlag von J. Cramer: Weinheim, Germany, 1964; pp. v–xv, 1–5, icons 1–395. [Google Scholar]
- Wood, R.D. A revision of the Characeae. In Monograph of the Characeae; Wood, R.D., Imahori, K., Eds.; Verlag von J. Cramer: Weinheim, Germany, 1965; pp. i–xxiv, 1–904. [Google Scholar]
- Jäger, D. Exemplare des Formenkreises Chara denudata A. Braun 1847 und Chara dissoluta A. Braun ex Leonhard 1864 aus dem Bodensee. Rostock. Meeresbiol. Beiträge 2010, 23, 29–39. [Google Scholar]
- Becker, R.; Blindow, I.; Doege, A.; Franke, T.; Gregor, T.; Hamann, U.; Jäger, D.; Jorda, C.; Kabus, T.; Korsch, H.; et al. Beschreibung der Characeen-Arten Deutschlands. In Armleuchteralgen. Die Characeen Deutschlands; Arbeitsgruppe Characeen Deutschlands Lehrstuhl für Ökologie der Universität Rostock, Ed.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 209–572. [Google Scholar] [CrossRef]
- Nat, E.; Raabe, U.; Romanov, R.; Schubert, H.; Stewart, N. A book about the charophytes of Europe. IRGC News 2017, 28, 21–22. [Google Scholar]
- Sakayama, H. Taxonomy of Nitella (Charales, Charophyceae) based on comparative morphology of oospores and multiple DNA marker phylogeny using cultured material. Phycol. Res. 2008, 56, 202–215. [Google Scholar] [CrossRef]
- Casanova, M.T. An overview of Nitella in Australia (Characeae, Charophyta). Aust. Syst. Bot. 2009, 22, 192–218. [Google Scholar] [CrossRef]
- Sakayama, H.; Kasai, F.; Nozaki, H.; Watanabe, M.M.; Kawachi, M.; Shigyo, M.; Nishihiro, J.; Washitani, I.; Krienitz, L.; Ito, M. Taxonomic reexamination of Chara globularis (Charales, Charophyceae) from Japan based on oospore morphology and rbcL gene sequences, and the description of C. leptospora sp. nov. J. Phycol. 2009, 45, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Pérez, W.; Hall, J.D.; McCourt, R.M.; Karol, K.G. Phylogeny of North American Tolypella (Charophyceae, Charophyta) based on plastid DNA sequences with a description of Tolypella ramosissima sp. nov. J. Phycol. 2014, 50, 776–789. [Google Scholar] [CrossRef] [PubMed]
- Romanov, R.E.; Gontcharov, A.A.; Barinova, S.S. Chara globata Mig. (Streptophyta: Charales): Rare species revised. Fottea Olomouc 2015, 15, 39–50. [Google Scholar] [CrossRef]
- Karol, K.G.; Alix, M.S.; Scribailo, R.W.; Skawinski, P.M.; Sleith, R.S.; Sardina, J.A.; Hall, J.D. New records of the rare North American endemic Chara brittonii (Characeae), with comments on its distribution. Brittonia 2018, 70, 277–288. [Google Scholar] [CrossRef]
- Romanov, R.E.; Barinova, S.S.; Nikulin, Y.V.; Gontcharov, A.A. Chara lipkinii (Charales, Charophyceae): A new dioecious Mediterranean species under risk of extinction in the wild and some implications for the taxonomy of the genus Chara. Fottea 2022, 22, 1–12. [Google Scholar] [CrossRef]
- Romanov, R.E.; Nikulin, A.Y.; Nikulin, V.Y.; Gontcharov, A.A. New species Chara oryzae and a new section Corillionia of Chara (Charales, Charophyceae) from European Mediterranean rice fields. Eur. J. Phycol. 2022, 57, 328–342. [Google Scholar] [CrossRef]
- Schneider, S.C.; Nowak, P.; von Ammon, U.; Ballot, A. Species differentiation in the genus Chara (Charophyceae): Considerable phenotypic plasticity occurs within homogenous genetic groups. Eur. J. Phycol. 2016, 51, 282–293. [Google Scholar] [CrossRef] [Green Version]
- Urbaniak, J.; Sakayama, H. Taxonomical analysis of closely related species of Chara L. section Hartmania (Streptophyta: Charales) based on morphological and molecular data. Fottea 2017, 17, 222–239. [Google Scholar] [CrossRef]
- Urbaniak, J.; Kwiatkowski, P. Taxonomic studies on the Chara section Hartmania in Poland based on morphological and molecular data. PhytoKeys 2019, 135, 71–90. [Google Scholar] [CrossRef] [PubMed]
- Braun, A. Die Characeen Afrika’s; Monatsberichte der Königlich preussischen Akademie der Wissenschaften zu Berlin: Berlin, Germany, 1868; Volume 1867, pp. 873–944. [Google Scholar]
- Groves, J.; Bullock-Webster, G.R. The British Charophyta; II. Charae; Ray Society: London, UK, 1924; pp. v–xi, 1–141, pl. 22–45. [Google Scholar]
- Hollerbach, M.M.; Krassavina, L.K. The Identification Manual of Freshwater Algae of the USSR. Iss. 14. The Charophytes—Charophyta; Nauka: Leningrad, Russia, 1983; pp. 1–190. (In Russian) [Google Scholar]
- Krause, W. Süßwasserflora von Mitteleuropa. Charales (Charophyceae), 18; Spektrum Akademischer Verlag: Heidelberg, Germany, 1997; pp. 1–202. [Google Scholar]
- Bailly, G.; Schaefer, O. Guide Illustré des Characées du Nord-Est de la France; Conservatoire Botanique National de Franche-Comté: Besançon, France, 2010; pp. 1–96. [Google Scholar]
- John, D.M.; Whitton, B.A.; Brook, A.J. (Eds.) Freshwater Algal Flora of the British Isles: An Identification Guide to Freshwater and Terrestrial Algae, 2nd ed.; Cambridge University Press: Cambridge, UK, 2011; pp. 1–878. [Google Scholar]
- Drège, J.F. Zwei pflanzengeographische Documente. Bes. Beigabe Zur Flora 1843, 26, 1–230. [Google Scholar]
- Romanov, R.; Stewart, N.; van de Weyer, K.; Roden, C.; Trbojević, I.; Biberdžić, V.; Trajanovska, S.; Alegro, A.; Kashta, L. Chara dissoluta. In Characeae of Europe; Schubert, H., Gregor, T., Blindow, I., Nat, E., Stewart, N., Romanov, R., van der Weyer, K., Denys, L., Korsch, H., Casanova, M.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2023; in press. [Google Scholar]
- Marcin, D. Hydrogeologická štruktúra baldovce—Sivá Brada. Podzemná Voda 2000, VI, 114–121. [Google Scholar]
- Echt, C.S.; Erdahl, L.A.; McCoy, T.J. Genetic segregation of random amplified polymorphic DNA in diploid cultivated alfalfa. Genome 1992, 35, 84–87. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Dubrovina, A.S.; Tyunin, A.P. The methylation status of plant genomic DNA influences PCR efficiency. J. Plant Physiol. 2015, 175, 59–67. [Google Scholar] [CrossRef]
- Bonfield, J.K.; Smith, K.F.; Staden, R. A new DNA sequence assembly program. Nucleic Acids Res. 1995, 23, 4992–4999. [Google Scholar] [CrossRef]
- Galtier, N.; Gouy, M.; Gautier, C. Seaview and phylo-win: Two graphic tools for sequence alignment and molecular phylogeny. Comput. Appl. Biosci. 1996, 12, 543–548. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*. Phylogenetic Analysis using Parsimony (*and Other Methods). Version 4.0b10; Sinauer Associates: Sunder-land, MA, USA, 2002. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef]
- Casanova, M.T.; Karol, K.G. A revision of Chara sect. Protochara, comb. et stat. nov. (Characeae: Charophyceae). Aust. Syst. Bot. 2014, 27, 23–37. [Google Scholar] [CrossRef]
- Proctor, V.W.; Griffin, D.G.; Hotchkiss, A.T. A synopsis of the genus Chara, series Gymnobasalia (subsection Willdenowia RDW). Am. J. Bot. 1971, 58, 894–901. [Google Scholar] [CrossRef]
- Casanova, M.T. Typification and circumscription of Nitella sonderi (Characeae, Charophyceae). Aust. Syst. Bot. 2007, 20, 464–472. [Google Scholar] [CrossRef]
- Casanova, M.T. Review of the species concepts Chara fibrosa and C. flaccida (Characeae, Charophyceae). Aust. Syst. Bot. 2013, 26, 291–297. [Google Scholar] [CrossRef]
- Milozza, A.; Abdelahad, N. The contribution of historical and morphological studies on herbarium specimens to a better definition of Chara pelosiana Avetta (Charales, Charophyceae). Plants 2021, 10, 2488. [Google Scholar] [CrossRef]
- Migula, W. Die Characeen Deutschlands, Oesterreichs und der Schweiz; Unter Berücksichtigung aller Arten Europas. Dr. L. Rabenhorst—Kryptogamenflora von Deutschland, Oesterreich und der Schweiz; Verlag Eduard Kummer: Leipzig, Germany, 1897; pp. VII, 1–765. [Google Scholar]
- Sluiter, C.P. Beiträge zur Kenntnis von Chara contraria A. Braun und Chara dissoluta A. Braun. Bot. Ztg. 1910, 68, 125–167. [Google Scholar]
- Hussain, M.I.; Victor, R.; Khoja, T.M. Charophytes of the Sultanate of Oman, Southern Arabia. Nova Hedwig. 2003, 77, 429–444. [Google Scholar] [CrossRef]
- Olsen, N.S. Danish Charophyta. Chorological, ecological and biological investigations. Det K. Dan. Vidensk. Selsk. Biol. Skr. 1944, 3, 1–240. [Google Scholar]
- Krause, W.; Krause, H. Exsikkate Europäischer Characeeen; 2. Aufl. 4 Bände, 1984–1986.
- Moore, J.A. Charophytes of Great Britain and Ireland; B.S.B.I. Handbook No. 5; Botanical Society of the British Isles: London, UK, 1986; pp. 1–144. [Google Scholar]
- Allen, G.O. Notes on charophytes from British Columbia. Proc. Linn. Soc. Lond. 1951, 162, 148–152. [Google Scholar] [CrossRef]
- Daily, F.K. The Characeae of Indiana. Butl. Univ. Bot. Stud. 1953, 11, 5–49. [Google Scholar]
- Naz, S.; Diba, N.J. Some morphological observations of charophytes (Characeae) from Bangladesh. J. Life Earth Sci. 2014, 7, 71–77. [Google Scholar] [CrossRef]
- Han, F.S.; Li, Y.Y. (Eds.) Flora Algarum Sinicarum Aquae Dulcis. Tomus 3. Charophyta; Science Press: Beijing, China, 1994; pp. 1–267. (In Chinese) [Google Scholar]
- Daily, F.K. Chara Kieneri, a new species from Nebraska. Butl. Univ. Bot. Stud. 1949, 9, 127–130. [Google Scholar]
- John, D.M.; Moore, J.A.; Green, D.R. Preliminary observations on the structure and ornamentation of the oosporangial wall in Chara (Charales, Chlorophyta). Br. Phycol. J. 1990, 25, 1–24. [Google Scholar] [CrossRef]
- Leitch, A.R.; John, D.M.; Moore, J.A. The oosporangium of the Characeae (Chlorophyta, Charales). Prog. Phycol. Res. 1990, 7, 213–268. [Google Scholar]
- Urbaniak, J.; Blaženčić, J. SEM study of oospore characteristics in endemic and endangered Balkan charophytes. Cryptogam. Algol. 2012, 33, 277–288. [Google Scholar] [CrossRef]
- Soulie–Märsche, I. Etude Comparée de Gyrogonites de Charophytes Actuelles et Fossiles et Phylogénie des Genres Actuels; (Thèse–ès–Sci. Univ. Montpellier 1979, Rev. Edit.); Imprimerie. des Tilleuls: Millau, France, 1989; pp. 1–237. [Google Scholar]
- Ray, S.; Pekkari, S.; Snoeijs, P. Oospore dimensions and wall ornamentation patterns in Swedish charophytes. Nord. J. Bot. 2001, 21, 207–224. [Google Scholar] [CrossRef]
- Mandal, D.K.; Blaženčić, J.; Ray, S. SEM study of compound oospore wall ornamentation of some members of Charales from Yugoslavia, Croatia and Slovenia. Arch. Biol. Sci. 2002, 54, 29–34. [Google Scholar] [CrossRef]
- Mandal, D.K.; Ray, S. Taxonomic significance of micromorphology and dimensions of oospores in the genus Chara L. (Charales, Chlorophyta). Arch. Biol. Sci. 2004, 56, 131–138. [Google Scholar] [CrossRef]
- Casanova, M.T. An overview of Chara L. in Australia (Characeae, Charophyta). Aust. Syst. Bot. 2005, 18, 25–39. [Google Scholar] [CrossRef]
- Urbaniak, J. A SEM and light microscopy study of the oospore wall ornamentation in Polish charophytes (Charales, Charophyceae)—Genus Chara. Nova Hedwig. 2011, 93, 1–28. [Google Scholar] [CrossRef]
- Ahmadi, A.; Riahi, H.; Sheidai, M.; van Raam, J.C. A study of the oospore characteristics in some charophytes (Characeae) of Iran. Nova Hedwig. 2012, 94, 487–504. [Google Scholar] [CrossRef]
- Romanov, R.E.; (Komarov Botanical Institute of the Russian Academy of Sciences, Saint-Petersburg, Russia). Personal communication, 2022. Unpublished Data.
- Mann, H.; Hanel, C.; Langangen, A.; Nowak, P. Chara contraria var. hispidula Braun (Charales) in Newfoundland, Canada a new variety described from North America. Bot. Lett. 2022, 169, 250–258. [Google Scholar] [CrossRef]
- Taylor, R.; (Independent Ecologist, Mtunzini, 3867, KwaZulu-Natal, South Africa). Personal communication, 2020.
- Janks, M.R.; (GroundTruth Wetlands cc, Hilton, Pietermaritzburg Area, South Africa). Personal communication, 2021.
- Janks, M.R. Montane Wetlands of the South African Great Escarpment: Plant Communities and Environmental Drivers. Master’s Thesis, Rhodes University, Makhanda, South Africa, December 2014. [Google Scholar]
- Rayss, T. Materiaux pour la Flore Algologique de la Palestine. II. Les Algues des Eaux Continentales. Palest. J. Bot. 1951, 5, 71–95. [Google Scholar]
- Guerlesquin, M. Nouvelle contribution à l’étude des Charophycées du Maroc nordoccidental (II). CNRS 1978, 249, 109–137. [Google Scholar]
- Stewart, N.F.; Church, J.M. Red Data Books of Britain and Ireland: Stoneworts; Joint Nature Conservation Committee: Peterborough, UK, 1992; pp. 1–144. [Google Scholar]
- Blaženčić, J.; Kashta, L.; Vesić, A.; Biberdžić, V.; Stevanović, B. Charophytes (Charales) of Lake Skadar/Shkodra: Ecology and Distribution. In The Skadar/Shkodra Lake Environment. The Handbook of Environmental Chemistry; Pešić, V., Karaman, G., Kostianoy, A., Eds.; Springer: Cham, Switzerland, 2018; Volume 80, pp. 169–202. [Google Scholar] [CrossRef]
- Corillion, R. Contribution à l’étude des Characées de Tunisie et bilan actuel de la flore charologique tunisienne. Bull. De La Société Phycol. De Fr. 1977, 22, 47–59. [Google Scholar]
- Barinova, S.; Smith, T. Flora of algae and cyanobacteria of continental waters of Israel in the XXI century: Taxonomy, autecology and water quality indicators. Diversity 2022, 14, 328. [Google Scholar] [CrossRef]
- Corillion, R. Sur la présence et la signification de formes à cortication incomplète du Chara contraria Kützing dans l’Est armoricain. Bull. De La Société Mayenne-Sci. 1960, 49–60. [Google Scholar]
- Pundhir, H.S. Vidyavati New karyomorphological observations on charophytes from Uttar Pradesh, India. Nucleus 1994, 37, 39–45. [Google Scholar]
- Allen, G.O. IX. Charophyta. Trans. Linn. Soc. Lond. 3rd Ser. 1940, 1, 155–160. [Google Scholar] [CrossRef]
- Guerlesquin, M. Charophytes. In Lake Titicaca: A Synthesys of Limnological Knowledge; Dejoux, C., Iltis, A., Eds.; Kluwer Academie Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 1992; pp. 232–240. [Google Scholar]
- Grant, M.C.; Proctor, V.W. Chara vulgaris and Chara contraria: Patterns of reproductive isolation for two cosmopolitan species complexes. Evolution 1972, 26, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Proctor, V.W. Chara globularis Thuillier (=C. fragilis Desvaux): Breeding patterns within a cosmopolitan complex. Limnol. Oceanogr. 1971, 16, 422–436. [Google Scholar] [CrossRef]
- Proctor, V.W. Genetics of Charophyta. In The Genetics of Algae; Botanical Monographs. Vol. 12; Lewin, R.A., Ed.; Blackwell Scientific Publications: Oxford, UK, 1976; pp. 210–218. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphological Traits | Values | |
|---|---|---|
| C. denudata | C. dissoluta | |
| Length of plant, cm | 7–8, 13–14 | >20/10–11 |
| Diameter of stem, μm | (486)513–945 | 453–757/243–392, 653–720 |
| Diameter of nodal bulbil, mm | 0.9–1.1 | –/– |
| Length of branchlet, mm | 7–13 | 12–33/6–14 |
| Length of adaxial bract cell, μm | 420–6380 | (233)286–960/(91)455–887 |
| Length of oogonium without coronula, μm | (456)651–810 | (427)495–785/455–721 |
| Width of oogonium, μm | 419–513 | (214) 256–486/(232)405–450 |
| Length of oogonium coronula, μm | –/– | –/130–170 |
| Width of oogonium coronula, μm | –/– | –/208–263 |
| Length of oospore without basal cage, μm | 538–723 | –/486–587 |
| Width of oospore, μm | 350–423 | –/233–328 |
| Length of basal cage, μm | 69–79 | –/– |
| Diameter of antheridium, μm | (214)228–288(317) | 209–428 / 243–500(645) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romanov, R.E.; Barinova, S.S.; Nikulin, V.Y.; Gontcharov, A.A. ‘Unfinished’ Morphogenesis Hides Different Speciation Pathways in Charophytes: Evidence from the 190-Year-Old Original Material of Chara denudata (Charales, Charophyceae). Diversity 2023, 15, 249. https://doi.org/10.3390/d15020249
Romanov RE, Barinova SS, Nikulin VY, Gontcharov AA. ‘Unfinished’ Morphogenesis Hides Different Speciation Pathways in Charophytes: Evidence from the 190-Year-Old Original Material of Chara denudata (Charales, Charophyceae). Diversity. 2023; 15(2):249. https://doi.org/10.3390/d15020249
Chicago/Turabian StyleRomanov, Roman E., Sophia S. Barinova, Vyacheslav Yu. Nikulin, and Andrey A. Gontcharov. 2023. "‘Unfinished’ Morphogenesis Hides Different Speciation Pathways in Charophytes: Evidence from the 190-Year-Old Original Material of Chara denudata (Charales, Charophyceae)" Diversity 15, no. 2: 249. https://doi.org/10.3390/d15020249