
Effect of Grape Pomace Intake on the Rumen Bacterial Community of Sheep
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
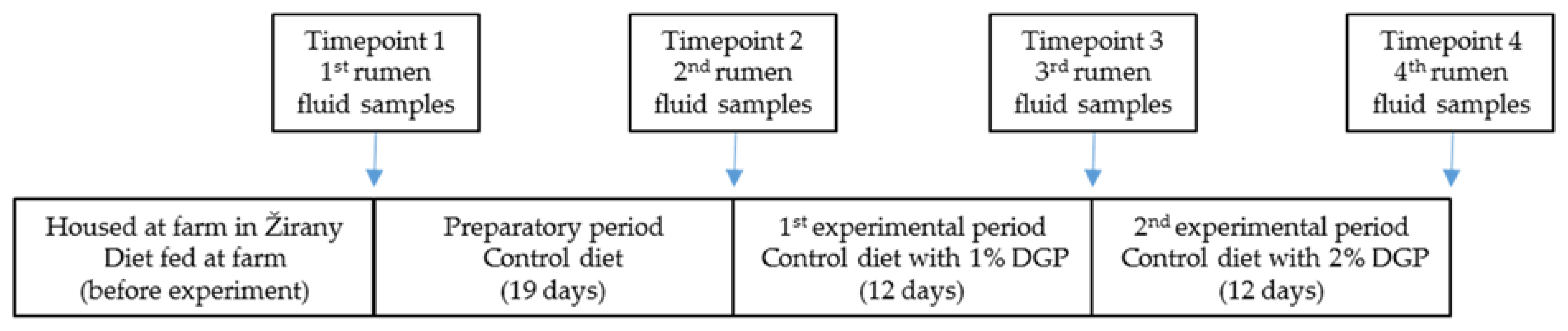
2.1. Animals and Experiment Design
2.2. Diet Composition and Analysis
2.3. Rumen Fluid Sample Collection
2.4. DNA Analysis
2.5. Statistical Analysis
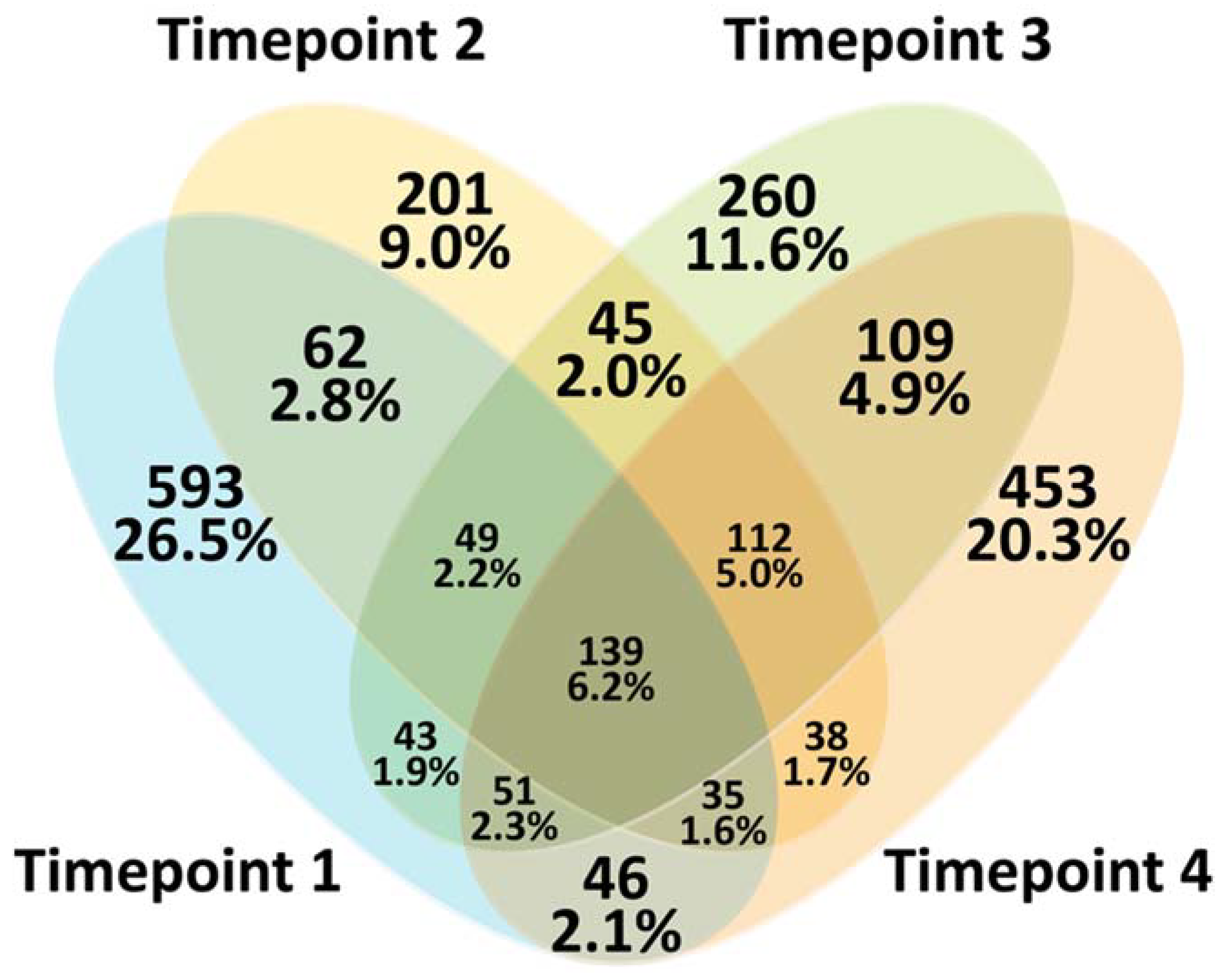
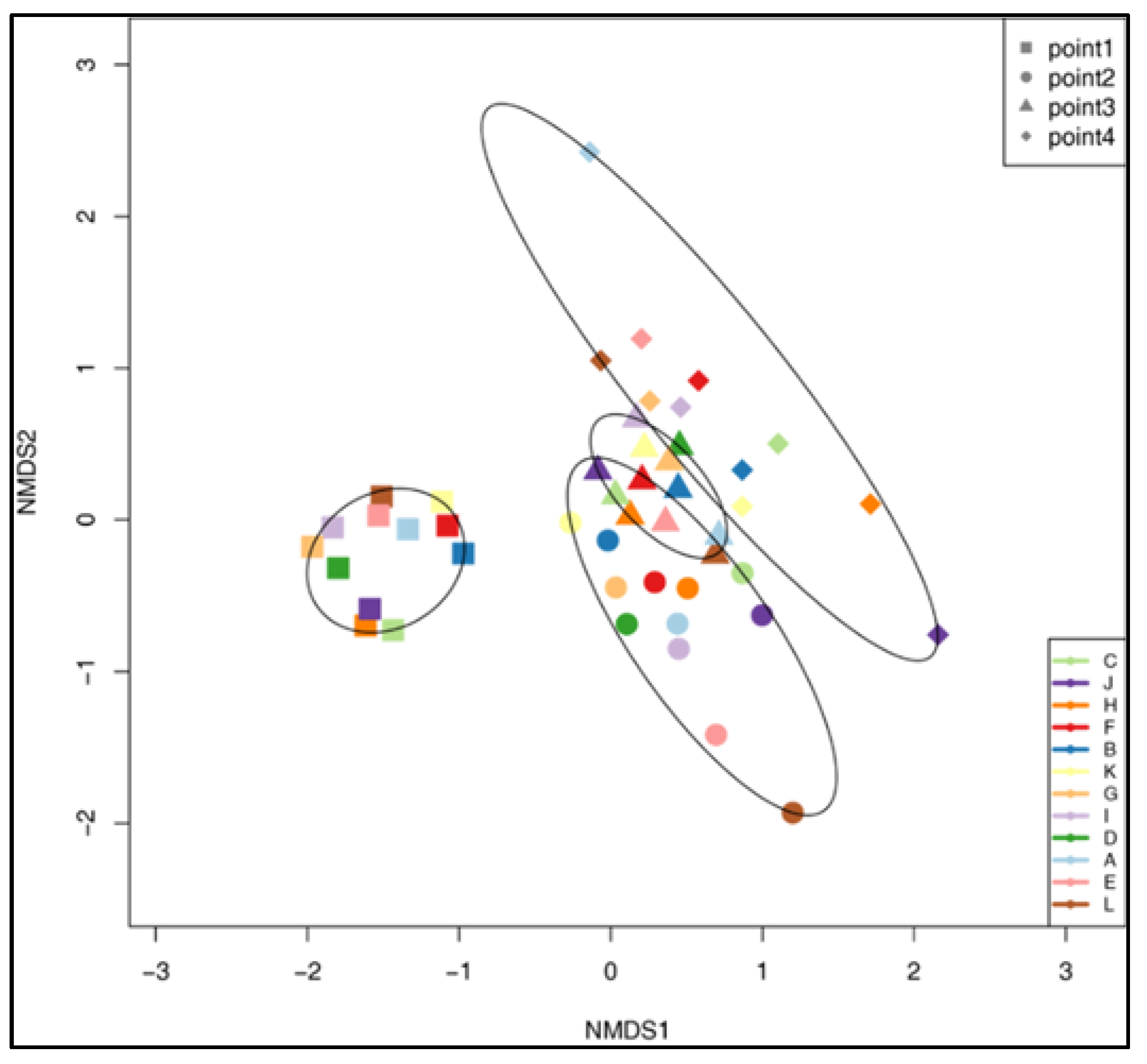
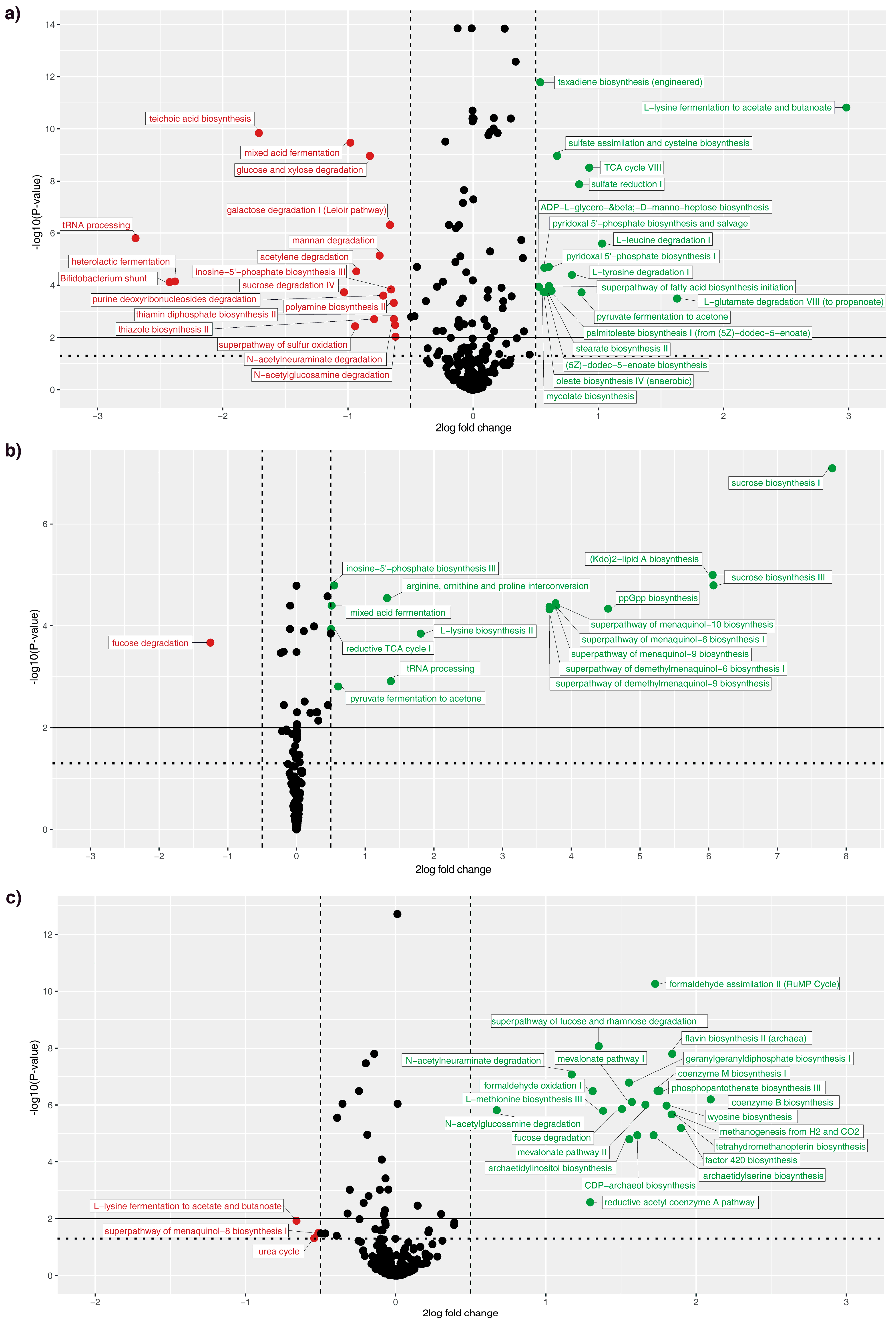
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanušovský, O.; Bíro, D.; Šimko, M.; Gálik, B.; Juráček, M.; Rolinec, M.; Majlát, M.; Herkeľ, R. Continual monitoring of reticulorumenal pH of dairy cows during 45 days. Acta Fytotechn. Zootechn. 2015, 18, 53–55. [Google Scholar] [CrossRef]
- Imrich, I.; Čopík, Š.; Mlyneková, E.; Mlynek, J.; Haščík, P.; Kanka, T. The effect of Saccharomyces cerevisiae additive to cattle ration on milk yield of dairy cows. Acta Fytotechn. Zootechn. 2021, 24, 45–48. [Google Scholar] [CrossRef]
- Hanušovský, O.; Bíro, D.; Šimko, M.; Gálik, B.; Juráček, M.; Rolinec, M.; Balušíková, Ľ. The dynamic of the ruminal content pH change and its relationship to milk composition. Acta Vet. Brno 2018, 87, 119–126. [Google Scholar] [CrossRef]
- Clark, H.J. Lactational responses to postruminal administration of proteins and amino acids. J. Dairy Sci. 1975, 58, 1178–1197. [Google Scholar] [CrossRef] [PubMed]
- OIV. International Organisation of Vine and Wine (OIV). 2018. Available online: https://www.oiv.int/what-we-do/global-report?oiv (accessed on 20 January 2023).
- Hanušovsky, O.; Gálik, B.; Bíro, D.; Šimko, M.; Juráček, M.; Rolinec, M.; Zábranský, L.; Philipp, C.; Puntigam, R.; Slama, J.A.; et al. The nutritional potential of grape by-products from the area of Slovakia and Austria. Emirates J. Food. Agri. 2020, 32, 1–10. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin-rich feeds. Small Rum. Res. 2003, 49, 241–256. [Google Scholar] [CrossRef]
- Mezzomo, R.; Paulino, P.V.R.; Barbosa, M.M.; Martins, T.S.; Pereira, L.G.R.; Silva, J.C.; Paulino, M.F.; Silva, M.C.; Serão, N.V. Protein dietary efficiency and methane emission in cattle fed soybean meal treated with tannins. Anim. Prod. Sci. 2017, 58, 2233–2241. [Google Scholar] [CrossRef]
- Nudda, A.; Correddu, F.; Marzano, A.; Battacone, G.; Nicolussi, P.; Bonelli, P.; Pulina, G. Effects of diets containing grape seed, linseed, or both on milk production traits, liver and kidney activities, and immunity of lactating dairy ewes. J. Dairy Sci. 2015, 98, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Ianni, A.; Martino, G. Dietary Grape Pomace Supplementation in Dairy Cows: Effect on Nutritional Quality of Milk and Its Derived Dairy Products. Foods 2020, 9, 168. [Google Scholar] [CrossRef]
- Bennato, F.; Ianni, A.; Florio, M.; Grotta, L.; Pomilio, F.; Saletti, M.A.; Martino, G. Nutritional properties of milk from dairy ewes fed with a diet containing grape pomace. Foods 2022, 11, 1878. [Google Scholar] [CrossRef]
- Vasta, V.; Daghio, M.; Cappucci, A.; Buccioni, A.; Serra, A.; Viti, C.; Mele, M. Invited review: Plant polyphenols and rumen microbiota responsible for fatty acid biohydrogenation, fiber digestion, and methane emission: Experimental evidence and methodological approaches. J. Dairy Sci. 2019, 102, 3781–3804. [Google Scholar] [CrossRef] [PubMed]
- Salami, S.A.; Valenti, B.; Luciano, G.; Lanza, M.; Umezurike-Amahah, N.M.; Kerry, J.P.; O’Grady, M.N.; Newbold, C.J.; Priolo, A. Dietary cardoon meal modulates rumen biohydrogenation and bacterial community in lambs. Sci. Rep. 2021, 11, 16180. [Google Scholar] [CrossRef] [PubMed]
- Biscarini, F.; Palazzo, F.; Castellani, F.; Masetti, G.; Grotta, L.; Cichelli, A.; Martino, G. Rumen microbiome in dairy calves fed copper and grape-pomace dietary supplementations: Composition and predicted functional profile. PLoS ONE 2018, 13, e0205670. [Google Scholar] [CrossRef] [PubMed]
- Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes (Text with EEA relevance). Off. J. Europ. Union 2010, L276.
- AOAC. Official Methods of Analysis AOAC, 17th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef]
- Větrovský, T.; Baldrian, P.; Morais, D. SEED 2: A user-friendly platform for amplicon high-throughput sequencing data analyses. Bioinformatics 2018, 34, 2292–2294. [Google Scholar] [CrossRef]
- Boylen, E.; Ram Rideout, J.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Env. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Love, M.I.; Huber, W.; Andres, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘Vegan’. Community Ecology Package, Version 2.5-6. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 2 October 2022).
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Morgavi, D.P.; Kelly, W.J.; Janssen, P.H.; Attwood, G.T. Rumen microbial (meta)genomics and its application to ruminant production. Animal 2013, 7, 184–201. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wang, Z.; Dong, C.; Li, F.; Wang, W.; Yuan, Z.; Mo, F.; Weng, X. Rumen Bacteria Communities and Performances of Fattening Lambs with a Lower or Greater Subacute Ruminal Acidosis Risk. Front. Microbiol. 2017, 12, 2506. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xu, T.; Xu, S.; Ma, L.; Han, X.; Wang, X.; Zhang, X.; Hu, L.; Zhao, N.; Chen, Y.; et al. Effect of dietary concentrate to forage ratio on growth performance, rumen fermentation and bacterial diversity of Tibetan sheep under barn feeding on the Qinghai-Tibetan plateau. PeerJ 2019, 5, e7462. [Google Scholar] [CrossRef] [PubMed]
- Diaz Carrasco, J.M.; Cabral, C.; Redondo, L.M.; Pin Viso, N.D.; Colombatto, D.; Farber, M.D.; Fernández Miyakawa, M.E. Impact of chestnut and quebracho tannins on rumen microbiota of bovines. BioMed. Res. Int. 2017, 2017, 9610810. [Google Scholar] [CrossRef]
- Ivanišová, E.; Terentjeva, M.; Kántor, A.; Frančáková, H.; Kačániová, M. Phytochemical and antioxidant profile of different varietes of grape from the Small Carpathians wine region of Slovakia. Erwerbs Obstbau 2019, 61, 53–59. [Google Scholar] [CrossRef]
- Frutos, P.; Hervas, G.; Giraldez, F.J.; Mantecon, A.R. Review. Tannins and ruminant nutrition. Span J Agric Res 2004, 2, 191–202. [Google Scholar] [CrossRef]
- Jones, G.A.; McAllister, T.A.; Muir, A.D.; Cheng, K.-J. Effects of sainfoin (Onobrichys viciifolia Scop.) condensed tannins on growth and proteolysis by four strains of ruminal bacteria. Appl. Env. Microbiol. 1994, 60, 1374–1378. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Wu, W.; Tu, Y.; Zhang, N.; Diao, Q. Resveratrol affects in vitro rumen fermentation, methane production and prokaryotic community composition in a time- and diet-specific manner. Microb. Biotecnol. 2020, 13, 1118–1131. [Google Scholar] [CrossRef]
- Witzig, M.; Zeder, M.; Rodehutscord, M. Effect of the ionophore monensin and tannin extracts supplemented to grass silage on populations of ruminal cellulolytics and methanogens in vitro. Anaerobe 2018, 50, 44–54. [Google Scholar] [CrossRef]
- Tavendale, M.H.; Meagher, L.P.; Pacheco, D.; Walker, N.; Attwood, G.T.; Sivakumaran, S. Methane production from in vitro rumen incubations with Lotus pedunculatus and Medicago sativa, and effects of extractable condensed tannin fractions on methanogenesis. Anim. Feed Sci. Technol. 2005, 123, 403–419. [Google Scholar] [CrossRef]
- Newbold, C.J.; Ramos-Morales, E. Review: Ruminal microbiome and microbial metabolome: Effects of diet and ruminant host. Animal 2020, 14 (Suppl. S1), 78–86. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.M.; Moate, P.J.; Marett, L.; Cocks, B.G.; Hayes, B.J. Investigating the effect of two methane- mitigating diets on the rumen microbiome using massively parallel sequencing. J. Dairy Sci. 2013, 96, 6030–6046. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Vinitchaikul, P.; Deng, M.; Zhang, G.; Sun, L.; Gou, X.; Mao, H.; Yang, S. Host and altitude factors affect rumen bacteria in cattle. Braz J. Microbiol. 2020, 51, 573–1583. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 14567. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.H.; Bian, G.R.; Zhu, W.Y.; Mao, S.Y. High-grain feeding causes strong shifts in ruminal epithelial bacterial community and expression of Toll-like receptor genes in goats. Front. Microbiol. 2015, 6, 167. [Google Scholar] [CrossRef]
- Belanche, A.; Kingston-Smith, A.H.; Griffith, G.W.; Newbold, C.J. A Multi-Kingdom Study Reveals the Plasticity of the Rumen Microbiota in Response to a Shift From Non-grazing to Grazing Diets in Sheep. Front. Microbiol. 2019, 10, 122. [Google Scholar] [CrossRef]
- Langda, S.; Zhang, C.; Zhang, K.; Gui, B.; Ji, D.; Deji, C.; Cuoji, A.; Wang, X.; Wu, Y. Diversity and Composition of Rumen Bacteria, Fungi, and Protozoa in Goats and Sheep Living in the Same High-Altitude Pasture. Animals 2020, 10, 186. [Google Scholar] [CrossRef]
- Vašeková, P.; Juráček, M.; Biro, D.; Šimko, M.; Gálik, B.; Rolinec, M.; Hanušovský, O.; Kolláthová, R.; Ivanišová, E. Bioactive compounds and fatty acid profile of grape pomace. Acta Fytotechn Zootechn 2020, 23, 230–235. [Google Scholar] [CrossRef]
- Juráček, M.; Vašeková, P.; Massányi, P.; Kováčik, A.; Bíro, D.; Šimko, M.; Gálik, B.; Rolinec, M.; Hanušovský, O.; Kolláthová, R.; et al. The effect of dried grape pomace feeding on nutrients digestibility and serum biochemical profile of wethers. Agriculture 2021, 11, 1194. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Fed at Farm in Žirany | Control Diet | Control Diet with 1% DGP | Control Diet with 2% DGP | |
|---|---|---|---|---|
| Feeds (g) | ||||
| Maize silage | 400 | - | - | - |
| Alfalfa silage | 300 | - | - | - |
| Barley grounded | 100 | - | - | - |
| Meadow hay | 500 | 700 | 700 | 700 |
| Wheat grounded | 100 | 118.6 | 118.6 | 118.6 |
| Soybean meal | - | 238.6 | 238.6 | 238.6 |
| DGP | - | - | 10.3 * | 20.6 ** |
| Nutrient concentrations in diet (g) | ||||
| Dry matter | 838.4 | 934.1 | 943.8 | 953.5 |
| Crude protein | 88.1 | 179.0 | 180.0 | 181.0 |
| Ether extract | 16.4 | 13.0 | 13.9 | 14.8 |
| Crude fiber | 265.8 | 288.0 | 289.9 | 291.8 |
| ADF | 310.5 | 351.4 | 355.3 | 359.2 |
| NDF | 474.8 | 529.8 | 534.5 | 539.3 |
| NFE | 491.2 | 520.6 | 526.8 | 532.9 |
| NFC | 282.2 | 278.8 | 282.1 | 285.4 |
| Organic matter | 861.4 | 1000.6 | 1010.5 | 1020.4 |
| Ash | 49.0 | 56.6 | 57.0 | 57.4 |
| Ca | 4.2 | 4.1 | 4.1 | 4.2 |
| P | 2.4 | 3.9 | 4.0 | 4.0 |
| Mg | 1.5 | 2.1 | 2.1 | 2.1 |
| Na | 0.3 | 0.3 | 0.3 | 0.3 |
| K | 11.1 | 15.5 | 15.6 | 15.8 |
| Phylum (%) | TP1 | TP2 | TP3 | TP4 | p-Value TP1 to TP2 | p-Value TP2 to TP3 | p-Value TP3 to TP4 | p-Value TP2 to TP4 |
|---|---|---|---|---|---|---|---|---|
| Actinobacteria | 2.50 | 0.15 | 0.03 | 0.29 | <0.001 | NA | NA | 0.291 |
| Bacteroidetes | 47.99 | 61.35 | 58.35 | 56.07 | 0.946 | 0.953 | 0.838 | 0.561 |
| Candidatus Saccharibacteria | 2.98 | 0.30 | 0.34 | 1.97 | <0.001 | 0.893 | 0.006 | 0.001 |
| Chloroflexi | 0.30 | 1.28 | 0.87 | 1.30 | 0.052 | 0.632 | 0.996 | 0.395 |
| Euryarchaeota | 1.34 | 1.30 | 2.20 | 7.74 | 0.946 | 0.976 | <0.001 | <0.001 |
| Fibrobacteres | 1.64 | 0.46 | 0.64 | 0.25 | <0.001 | 0.976 | 0.006 | 0.261 |
| Firmicutes | 23.85 | 20.15 | 15.59 | 16.92 | 0.005 | 0.428 | 0.644 | 0.056 |
| Proteobacteria | 3.59 | 1.56 | 3.80 | 3.83 | 0.001 | 0.368 | 0.514 | 0.516 |
| Spirochaetes | 1.59 | 0.82 | 0.08 | 0.11 | 0.001 | 0.367 | 0.838 | <0.001 |
| SR1 | 5.25 | 0.17 | 0.51 | 0.18 | <0.001 | 0.976 | 0.609 | 0.326 |
| Synergistetes | 0.90 | 1.44 | 5.19 | 4.57 | <0.001 | <0.001 | 0.838 | n. d. |
| Tenericutes | 1.02 | 0.18 | 0.21 | 0.13 | 0.011 | 0.976 | 0.060 | 0.001 |
| Verrucomicrobia | 6.87 | 10.44 | 11.36 | 5.86 | 0.478 | 0.989 | <0.001 | <0.001 |
| Family (%) | TP1 | TP2 | TP3 | TP4 | p-Value TP1 to TP2 | p-Value TP2 to TP3 | p-Value TP3 to TP4 | p-Value TP2 to TP4 |
|---|---|---|---|---|---|---|---|---|
| Acidaminococcaceae | 3.80 | 5.21 | 2.88 | 2.49 | 0.760 | 0.005 | 0.559 | <0.001 |
| Anaerolineaceae | 0.29 | 1.28 | 0.87 | 1.30 | 0.088 | 0.994 | 0.964 | n. d. |
| Bdellovibrionaceae | 1.56 | 0.33 | 0.97 | 0.22 | 0.009 | 0.994 | 0.016 | 0.332 |
| Bifidobacteriaceae | 2.44 | 0.00 | 0.00 | 0.02 | <0.001 | 0.994 | 0.964 | 0.738 |
| Chitinophagaceae | 1.40 | 0.40 | 0.13 | 0.43 | 0.005 | 0.487 | 0.051 | 0.719 |
| Fibrobacteraceae | 1.64 | 0.46 | 0.64 | 0.25 | <0.001 | 0.994 | 0.003 | 0.222 |
| Flavobacteriaceae | 1.20 | 1.00 | 0.87 | 1.20 | 0.453 | 0.994 | 0.559 | 0.791 |
| Gracilibacteraceae | 1.10 | 1.23 | 1.75 | 1.69 | 0.657 | 0.994 | 0.964 | n. d. |
| Lachnospiraceae | 3.35 | 2.46 | 2.38 | 3.89 | 0.001 | 0.994 | <0.001 | 0.011 |
| Marinilabiliaceae | 1.65 | 2.34 | 1.97 | 1.05 | 0.760 | 0.994 | 0.001 | <0.001 |
| Methanobacteriaceae | 1.07 | 1.06 | 1.53 | 6.85 | 0.981 | 0.994 | <0.001 | <0.001 |
| Pasteurellaceae | 0.43 | 0.13 | 1.00 | 0.04 | 0.099 | 0.348 | 0.010 | n. d. |
| Porphyromonadaceae | 8.39 | 23.11 | 17.75 | 22.23 | 0.006 | 0.866 | 0.509 | 0.004 |
| Prevotellaceae | 32.30 | 28.80 | 33.38 | 24.44 | 0.002 | 0.387 | 0.003 | 0.796 |
| Prolixibacteraceae | 0.45 | 1.00 | 0.55 | 1.10 | 0.387 | 0.866 | 0.009 | 0.981 |
| Rikenellaceae | 1.58 | 3.61 | 2.80 | 3.45 | 0.001 | 0.994 | 0.723 | 0.875 |
| Ruminococcaceae | 11.23 | 6.50 | 5.21 | 5.12 | 0.004 | 0.994 | 0.964 | 0.745 |
| Saccharibacteria_genera_incertae_sedis | 2.98 | 0.30 | 0.34 | 1.97 | <0.001 | 0.994 | 0.062 | 0.002 |
| Sphingobacteriaceae | 0.52 | 0.49 | 0.80 | 1.35 | 0.970 | 0.866 | 0.032 | 0.705 |
| Spirochaetaceae | 1.59 | 0.82 | 0.08 | 0.11 | <0.001 | 0.387 | 0.964 | <0.001 |
| SR1_genera_incertae_sedis | 5.25 | 0.17 | 0.51 | 0.18 | <0.001 | 0.994 | 0.964 | 0.413 |
| Synergistaceae | 0.09 | 1.44 | 5.19 | 4.57 | <0.001 | <0.001 | 0.964 | <0.001 |
| Syntrophorhabdaceae | 0.00 | 0.02 | 0.63 | 2.71 | 0.981 | 0.994 | 0.509 | 0.280 |
| Veillonellaceae | 3.00 | 4.18 | 2.89 | 2.84 | 0.803 | 0.994 | 0.964 | 0.997 |
| Verrucomicrobia _Subdivision5_genera_incertae_sedis | 6.79 | 10.23 | 10.37 | 4.99 | 0.466 | 0.994 | 0.001 | 0.023 |
| Genera (%) | TP1 | TP2 | TP3 | TP4 | p-Value TP1 to TP2 | p-Value TP2 to TP3 | p-Value TP3 to TP4 | p-Value TP2 to TP4 |
|---|---|---|---|---|---|---|---|---|
| Acetobacteroides | 0.45 | 10.57 | 9.23 | 10.80 | <0.001 | 0.994 | 0.650 | 0.928 |
| Alloprevotella | 0.92 | 0.43 | 0.61 | 0.10 | <0.001 | 0.994 | <0.001 | 0.139 |
| Barnesiella | 1.16 | 3.43 | 0.94 | 4.24 | 0.148 | 0.007 | <0.001 | 0.560 |
| Bifidobacterium | 2.44 | 0.00 | 0.00 | 0.02 | <0.001 | 0.994 | 0.954 | 0.825 |
| Butyrivibrio | 0.73 | 0.66 | 1.26 | 2.12 | 0.096 | 0.099 | <0.001 | <0.001 |
| Centipeda | 0.31 | 1.15 | 0.64 | 0.70 | 0.148 | 0.994 | 0.985 | <0.001 |
| Clostridium IV | 2.05 | 0.94 | 0.19 | 0.34 | <0.001 | 0.081 | 0.242 | 0.097 |
| Chelonobacter | 0.34 | 0.00 | 1.00 | 0.02 | <0.001 | <0.001 | 0.001 | 0.400 |
| Falsiporphyromonas | 2.93 | 4.97 | 6.17 | 4.54 | 0.913 | 0.253 | 0.588 | 0.003 |
| Fibrobacter | 1.64 | 0.46 | 0.64 | 0.25 | <0.001 | 0.994 | 0.012 | 0.079 |
| Flavonifractor | 1.19 | 0.63 | 0.91 | 0.45 | 0.400 | 0.994 | 0.954 | n. d. |
| Fretibacterium | 0.04 | 1.36 | 5.15 | 4.56 | <0.001 | <0.001 | 0.954 | <0.001 |
| Gracilibacter | 1.05 | 1.21 | 1.72 | 1.68 | 0.594 | 0.984 | NA | n. d. |
| Mangroviflexus | 0.26 | 1.09 | 1.03 | 0.30 | <0.001 | 0.994 | <0.001 | <0.001 |
| Methanobrevibacter | 1.07 | 1.06 | 1.53 | 6.84 | 0.873 | 0.994 | <0.001 | <0.001 |
| Methanomassiliicoccus | 0.27 | 0.18 | 0.66 | 0.90 | 0.647 | 0.692 | 0.808 | 0.057 |
| Mucinivorans | 1.58 | 3.55 | 2.75 | 3.41 | 0.067 | 0.994 | 0.422 | 0.441 |
| Ornatilinea | 0.29 | 1.12 | 0.87 | 1.29 | 0.319 | 0.994 | 0.954 | n. d. |
| Oscillibacter | 0.75 | 1.50 | 0.52 | 0.46 | 0.924 | 0.994 | 0.954 | n. d. |
| Parafilimonas | 1.40 | 0.40 | 0.13 | 0.39 | 0.001 | 0.467 | 0.062 | n. d. |
| Paraprevotella | 3.33 | 2.02 | 3.38 | 1.47 | <0.001 | 0.031 | <0.001 | <0.001 |
| Petrimonas | 0.03 | 0.68 | 0.96 | 0.77 | <0.001 | 0.185 | 0.097 | n. d. |
| Prevotella | 28.05 | 26.35 | 29.39 | 22.86 | <0.001 | 0.692 | 0.377 | 0.169 |
| Pseudosphingobacterium | 0.46 | 0.42 | 0.74 | 1.04 | 0.396 | 0.902 | 0.327 | n. d. |
| Ruminococcus | 1.85 | 0.43 | 0.46 | 0.27 | <0.001 | 0.976 | 0.741 | 0.361 |
| Saccharibacteria_genera_incertae_sedis | 2.98 | 0.30 | 0.34 | 1.97 | <0.001 | 0.994 | 0.062 | 0.008 |
| Saccharofermentans | 2.80 | 0.87 | 0.91 | 1.16 | <0.001 | 0.994 | 0.479 | 0.376 |
| Schwartzia | 0.85 | 0.80 | 1.09 | 0.36 | 0.027 | 0.976 | 0.002 | <0.001 |
| Selenomonas | 1.31 | 1.84 | 0.13 | 0.20 | 0.373 | 0.009 | 0.741 | <0.001 |
| Sporobacter | 0.40 | 0.57 | 1.09 | 1.11 | 0.873 | 0.994 | 0.954 | 0.155 |
| SR1_genera_incertae_sedis | 5.25 | 0.17 | 0.51 | 0.18 | <0.001 | 0.994 | 0.954 | 0.629 |
| Succiniclasticum | 3.78 | 5.10 | 2.88 | 2.48 | 0.755 | 0.001 | 0.954 | <0.001 |
| Syntrophorhabdus | 0.00 | 0.02 | 0.63 | 2.71 | 0.924 | 0.994 | 0.422 | 0.365 |
| Tannerella | 3.30 | 2.63 | 0.30 | 1.71 | 0.179 | 0.016 | 0.077 | n. d. |
| Treponema | 1.44 | 0.80 | 0.07 | 0.10 | 0.001 | 0.383 | 0.954 | <0.001 |
| Vampirovibrio | 1.56 | 0.33 | 0.97 | 0.22 | 0.005 | 0.994 | 0.059 | 0.194 |
| Verrucomicrobia _Subdivision3_genera_incertae_sedis | 0.00 | 0.06 | 0.98 | 0.85 | 0.265 | <0.001 | 0.954 | n. d. |
| Verrucomicrobia _Subdivision5_genera_incertae_sedis | 6.79 | 10.23 | 10.37 | 4.99 | 0.924 | 0.994 | 0.003 | n. d. |
| Indices (Mean ± SD) | TP1 | TP2 | TP3 | TP4 |
|---|---|---|---|---|
| Richness | 275 ± 39 a | 144 ± 45 b | 192 ± 30 c | 204 ± 75 c |
| Evenness | 0.88 ± 0.023 ab | 0.90 ± 0.013 a | 0.89 ± 0.013 ab | 0.87 ± 0.047 b |
| Shannon | 7.17 ± 0.32 a | 6.39 ± 0.46 b | 6.73 ± 0.21 ab | 6.61 ± 0.74 b |
| ANOSIM | TP1 | TP2 | TP3 | TP4 |
|---|---|---|---|---|
| TP1 | p = 0.001 | p = 0.001 | p = 0.001 | |
| TP2 | R = 0.819 | p = 0.001 | p = 0.001 | |
| TP3 | R = 0.958 | R = 0.306 | p = 0.001 | |
| TP4 | R = 0.934 | R = 0.588 | R = 0.320 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rolinec, M.; Medo, J.; Gábor, M.; Miluchová, M.; Šimko, M.; Gálik, B.; Hanušovský, O.; Schubertová, Z.; Bíro, D.; Zábranský, L.; et al. Effect of Grape Pomace Intake on the Rumen Bacterial Community of Sheep. Diversity 2023, 15, 234. https://doi.org/10.3390/d15020234
Rolinec M, Medo J, Gábor M, Miluchová M, Šimko M, Gálik B, Hanušovský O, Schubertová Z, Bíro D, Zábranský L, et al. Effect of Grape Pomace Intake on the Rumen Bacterial Community of Sheep. Diversity. 2023; 15(2):234. https://doi.org/10.3390/d15020234
Chicago/Turabian StyleRolinec, Michal, Juraj Medo, Michal Gábor, Martina Miluchová, Milan Šimko, Branislav Gálik, Ondrej Hanušovský, Zuzana Schubertová, Daniel Bíro, Luboš Zábranský, and et al. 2023. "Effect of Grape Pomace Intake on the Rumen Bacterial Community of Sheep" Diversity 15, no. 2: 234. https://doi.org/10.3390/d15020234