
Basal Anseriformes from the Early Paleogene of North America and Europe †


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. The Fossil Record of Anseriformes
3. Methods
4. Systematic Paleontology
4.1. Class Aves Linnaeus, 1758
Family Anachronornithidae nov. fam. Houde, Dickson, and Camarena
4.2. Anseriformes Familia Incertae Sedis
4.2.1. Anseriformes Familia Incertae Sedis, Willwood Formation
4.2.2. Anseriformes Familia Incertae Sedis, Green River Formation
4.2.3. Anseriformes Familia Incertae Sedis, London Clay Formation
London Clay Group A
London Clay Group B Referred Specimens
London Clay Group C Referred Specimens
London Clay Group D Referred Specimen
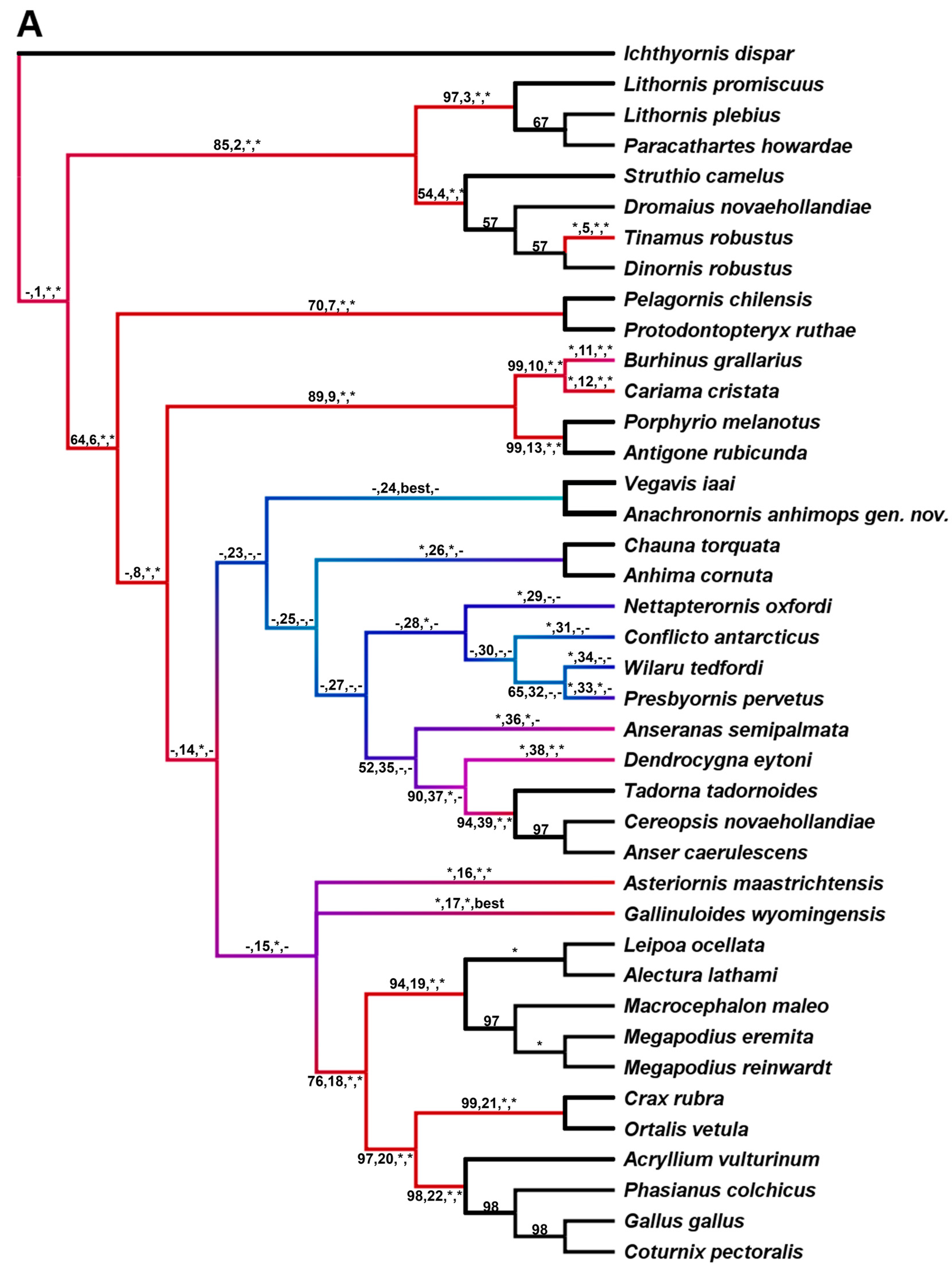
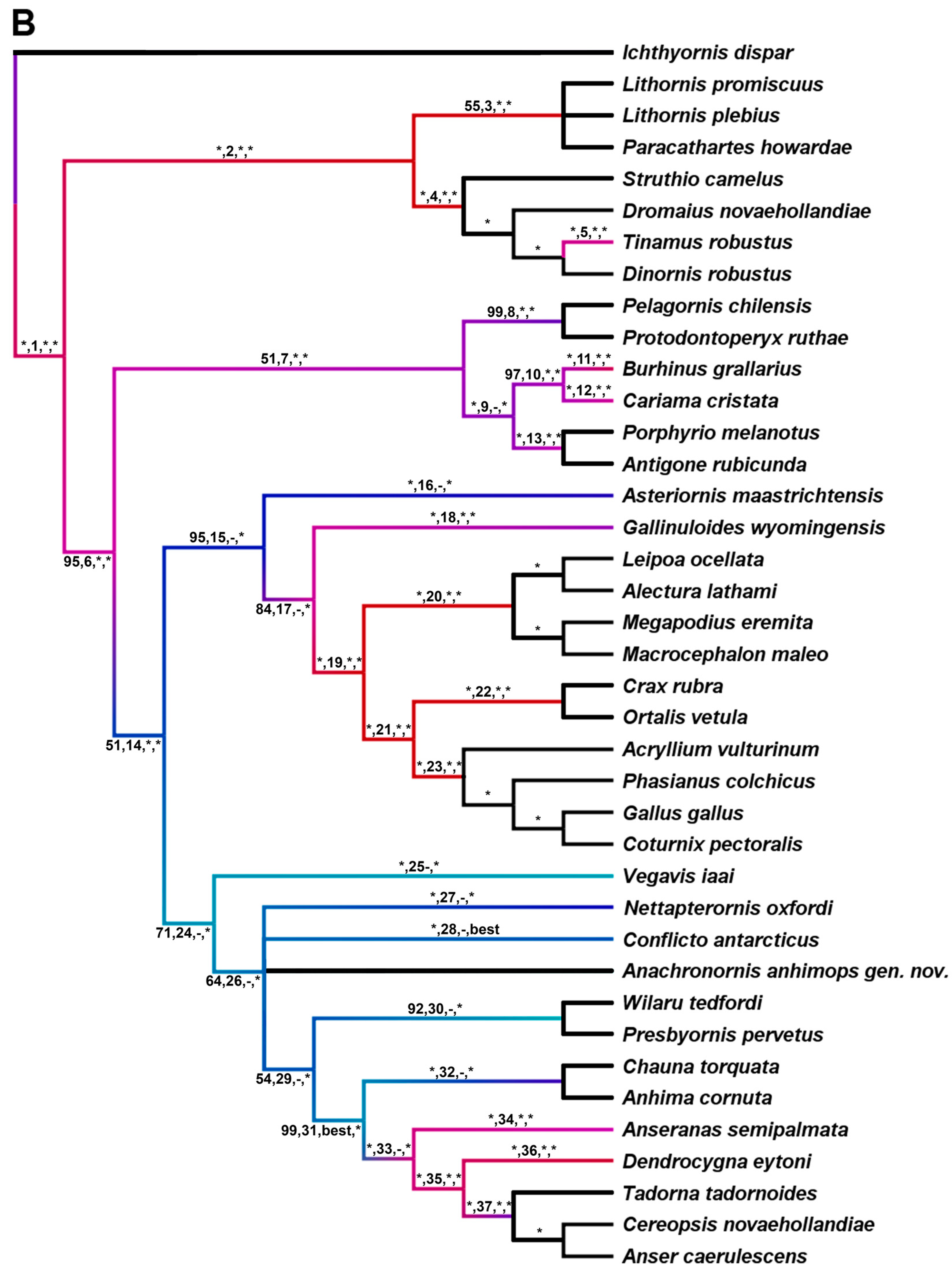
5. Results of Phylogenetic Analyses
5.1. Dataset 1
5.2. Dataset 2
5.3. Dataset 3
5.4. Dataset 4
5.5. Dataset 5
5.6. Dataset 6
5.7. Dataset 7
5.7.1. Phylogenetically Constrained Parsimony Tree
5.7.2. Fully Unconstrained Bayesian Tree
5.7.3. Phylogenetically Unconstrained Total-Evidence Bayesian Tree
5.7.4. Phylogenetically Constrained Total-Evidence Bayesian Tree
6. Discussion
6.1. Phylogenetic Analysis
6.2. Character Distributions of the Datasets
6.3. Neornithine Symplesiomorphies
6.4. Comparison to Presbyornis, Nettapterornis, and Conflicto
6.5. Character Polarity of the Spatulate Bill in Anseriformes
6.6. Ecology of Anachronornis nov. gen.
6.7. Postorbital and Zygomatic Processes
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Livezey, B.C. A Phylogenetic Analysis of Basal Anseriformes, the Fossil Presbyornis, and the Interordinal Relationships of Waterfowl. Zool. J. Linn. Soc. 1997, 121, 361–428. [Google Scholar] [CrossRef]
- Olson, S.L.; Feduccia, A. Presbyornis and the Origin of the Anseriformes (Aves: Charadriomorphae). Smithson. Contrib. Zool. 1980, 323, 1–24. [Google Scholar] [CrossRef]
- Livezey, B.C.; Zusi, R.L. Higher-Order Phylogeny of Modern Birds (Theropoda, Aves: Neornithes) Based on Comparative Anatomy. II. Analysis and Discussion. Zool. J. Linn. Soc. 2007, 149, 1–95. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, E.D.; Mirarab, S.; Aberer, A.J.; Li, B.; Houde, P.; Li, C.; Ho, S.Y.W.; Faircloth, B.C.; Nabholz, B.; Howard, J.T.; et al. Whole-Genome Analyses Resolve Early Branches in the Tree of Life of Modern Birds. Science 2014, 346, 1320–1331. [Google Scholar] [CrossRef] [PubMed]
- Claramunt, S.; Cracraft, J. A New Time Tree Reveals Earth History’s Imprint on the Evolution of Modern Birds. Sci. Adv. 2015, 1, e1501005. [Google Scholar] [CrossRef]
- Prum, R.O.; Berv, J.S.; Dornburg, A.; Field, D.J.; Townsend, J.P.; Lemmon, E.M.; Lemmon, A.R. A Comprehensive Phylogeny of Birds (Aves) Using Targeted next-Generation DNA Sequencing. Nature 2015, 526, 569–573. [Google Scholar] [CrossRef]
- Kimball, R.T.; Oliveros, C.H.; Wang, N.; White, N.D.; Barker, F.K.; Field, D.J.; Ksepka, D.T.; Chesser, R.T.; Moyle, R.G.; Braun, M.J.; et al. A Phylogenomic Supertree of Birds. Diversity 2019, 11, 109. [Google Scholar] [CrossRef]
- Olson, S.L. The Fossil Record of Birds. In Avian Biology; Farner, D.S., King, J., Parkes, K.C., Eds.; Academic Press: New York, NY, USA, 1985; pp. 79–238. [Google Scholar]
- Feduccia, A. The Origin and Evolution of Birds; Yale University Press: New Haven, CT, USA, 1996. [Google Scholar]
- Ericson, P.G.P. Systematic Relationships of the Palaeogene Family Presbyornithidae (Aves: Anseriformes). Zool. J. Linn. Soc. 1997, 121, 429–483. [Google Scholar] [CrossRef]
- Waddell, P.J.; Cao, Y.; Hasegawa, M.; Mindell, D.P. Assessing the Cretaceous Superordinal Divergence Times within Birds and Placental Mammals by Using Whole Mitochondrial Protein Sequences and an Extended Statistical Framework. Syst. Biol. 1999, 48, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Dyke, G.J. The Fossil Waterfowl (Aves: Anseriformes) from the Eocene of England. Am. Mus. Novit. 2001, 3354, 1–15. [Google Scholar] [CrossRef]
- Kurochkin, E.N.; Dyke, G.J.; Karhu, A.A. A New Presbyornithid Bird (Aves, Anseriformes) from the Late Cretaceous of Southern Mongolia. Am. Mus. Novit. 2002, 3386, 1–11. [Google Scholar] [CrossRef]
- Clarke, J.A.; Tambussi, C.P.; Noriega, J.I.; Erickson, G.M.; Ketcham, R.A. Definitive Fossil Evidence for the Extant Avian Radiation in the Cretaceous. Nature 2005, 433, 305–308. [Google Scholar] [CrossRef]
- Tambussi, C.P.; Degrange, F.J.; De Mendoza, R.S.; Sferco, E.; Santillana, S. A Stem Anseriform from the Early Palaeocene of Antarctica Provides New Key Evidence in the Early Evolution of Waterfowl. Zool. J. Linn. Soc. 2019, 186, 673–700. [Google Scholar] [CrossRef]
- Parham, J.F.; Donoghue, P.C.J.; Bell, C.J.; Calway, T.D.; Head, J.J.; Holroyd, P.A.; Inoue, J.G.; Irmis, R.B.; Joyce, W.G.; Ksepka, D.T.; et al. Best Practices for Justifying Fossil Calibrations. Syst. Biol. 2012, 61, 346–359. [Google Scholar] [CrossRef] [PubMed]
- Hope, S. The Mesozoic Radiation of Neornithes. In Mesozoic Birds: Above the Heads of Dinosaurs; Chiappe, L.M., Witmer, L.M., Eds.; University of California Press: Berkeley, CA, USA, 2002; pp. 339–388. [Google Scholar]
- Ericson, P.G.P.; Parsons, T.J.; Johansson, U.S. Morphological and Molecular Support for the Nonmonophyly of the Galloanserae. In Proceedings of the Proceedings of the International Symposium in Honor of John, H. Ostrom; Gauthier, J., Gall, J.E., Eds.; Yale University Press: New Haven, CT, USA, 2001; pp. 159–168. [Google Scholar]
- Elzanowski, A.; Stidham, T.A. Morphology of the Quadrate in the Eocene Anseriform Presbyornis and Extant Galloanserine Birds. J. Morphol. 2010, 271, 305–323. [Google Scholar] [CrossRef]
- Field, D.J.; Benito, J.; Chen, A.; Jagt, J.W.M.; Ksepka, D.T. Late Cretaceous Neornithine from Europe Illuminates the Origins of Crown Birds. Nature 2020, 579, 397–401. [Google Scholar] [CrossRef]
- Mlíkovský, J. Cenozoic Birds of the World, Part 1: Europe; Ninox Press: Prauge, Czech Republic, 2002. [Google Scholar]
- Ericson, P.G.P. Systematic Revision, Skeletal Anatomy, and Paleoecology of the New World Early Tertiary Presbyornithidae (Aves: Anseriformes). PaleoBios 2000, 20, 1–23. [Google Scholar]
- Dyke, G.J.; Van Tuinen, M. The Evolutionary Radiation of Modern Birds (Neornithes): Reconciling Molecules, Morphology and the Fossil Record. Zool. J. Linn. Soc. 2004, 141, 153–177. [Google Scholar] [CrossRef]
- Mayr, G. The Paleogene Fossil Record of Birds in Europe. Biol. Rev. 2005, 80, 515. [Google Scholar] [CrossRef] [PubMed]
- Mayr, G. Phylogenetic Affinities and Morphology of the Late Eocene Anseriform Bird Romainvillia Stehlini Lebedinsky, 1927. Neues Jahrb. Für Geol. Und Paläontologie-Abh. 2008, 248, 365–380. [Google Scholar] [CrossRef]
- Mayr, G. Paleogene fossil birds. In Fascinating Life Sciences, 2nd ed.; Springer International Publishing: Cham, Switzerland, 2022; ISBN 978-3-030-87644-9. [Google Scholar]
- Kurochkin, E.N.; Dyke, G.J. A Large Collection of Presbyornis (Aves, Anseriformes, Presbyornithidae) from the Late Paleocene and Early Eocene of Mongolia. Geol. J. 2010, 45, 375–387. [Google Scholar] [CrossRef]
- Mayr, G.; De Pietri, V.L. A Goose-Sized Anseriform Bird from the Late Oligocene of France: The Youngest Record and Largest Species of Romainvilliinae. Paläontol. Z. 2013, 87, 423–430. [Google Scholar] [CrossRef]
- Stidham, T.A.; Ni, X.-J. Large Anseriform (Aves: Anatidae: Romainvilliinae?) Fossils from the Late Eocene of Xinjiang, China. Vertebr. PalAsiatica 2014, 52, 98–111. [Google Scholar]
- De Pietri, V.L.; Scofield, R.P.; Zelenkov, N.; Boles, W.E.; Worthy, T.H. The Unexpected Survival of an Ancient Lineage of Anseriform Birds into the Neogene of Australia: The Youngest Record of Presbyornithidae. R. Soc. Open Sci. 2016, 3, 150635. [Google Scholar] [CrossRef]
- Worthy, T.H.; Lee, M.S.Y. Affinities of Miocene Waterfowl (Anatidae: ManuherikiaI, Dunstanetta, and Miotadorna) from the St Bathans Fauna, New Zealand. Palaeontology 2008, 51, 677–708. [Google Scholar] [CrossRef]
- Zelenkov, N.V. Cenozoic Evolution of Eurasian Anatids (Aves: Anatidae s. l.). Biol. Bull. Rev. 2020, 10, 417–426. [Google Scholar] [CrossRef]
- Zelenkov, N.V. A Revision of the Palaeocene–Eocene Mongolian Presbyornithidae (Aves: Anseriformes). Paleontol. J. 2021, 55, 323–330. [Google Scholar] [CrossRef]
- Alvarenga, H.M.F. A Fossil Screamer (Anseriformes: Anhimidae) from the Middle Tertiary of Southeastern Brazil. Smithson. Contrib. Zool. 1999, 89, 223–230. [Google Scholar]
- Elzanowski, A.; Boles, W.E. Australia’s Oldest Anseriform Fossil: A Quadrate from the Early Eocene Tingamarra Fauna. Palaeontology 2012, 55, 903–911. [Google Scholar] [CrossRef]
- Zelenkov, N.V. A Swan-Sized Anseriform Bird from the Late Paleocene of Mongolia. J. Vertebr. Paleontol. 2018, 38, e1531879. [Google Scholar] [CrossRef]
- Mayr, G. A New Avian Species with Tubercle-Bearing Cervical Vertebrae from the Middle Eocene of Messel (Germany). Rec. Aust. Mus. 2010, 62, 21–28. [Google Scholar] [CrossRef]
- Zelenkov, N.V.; Martynovich, N.V. A rich bird fauna from the Miocene locality Tagay (Olkhon Island, Baikal Lake). Arch. Menzbier. Ornithol. Soc. 2013, 2, 73–93. [Google Scholar]
- Noriega, J.I.; Tambussi, C.P. A Late Cretaceous Presbyornithidae (Aves: Anseriformes) from Vega Island, Antarctic Peninsula: Paleo-Biogeographic Implications. Ameghiniana 1995, 32, 57–61. [Google Scholar]
- Clarke, J.A.; Norell, M.A. New Avialan Remains and a Review of the Known Avifauna from the Late Cretaceous Nemegt Formation of Mongolia. Am. Mus. Novit. 2004, 3447, 1–12. [Google Scholar] [CrossRef]
- Novas, F.; Agnolin, F.; Rozadilla, S.; Aranciaga-Rolando, A.; Brissón-Eli, F.; Motta, M.; Cerroni, M.; Ezcurra, M.; Martinelli, A.; D’Angelo, J.; et al. Paleontological Discoveries in the Chorrillo Formation (Upper Campanian-Lower Maastrichtian, Upper Cretaceous), Santa Cruz Province, Patagonia, Argentina. MACN 2019, 21, 217–293. [Google Scholar] [CrossRef] [Green Version]
- Agnolín, F.L.; Egli, F.B.; Chatterjee, S.; Alexis, J.; Marsa, G.; Novas, F.E. Vegaviidae, a New Clade of Southern Diving Birds That Survived the K/T Boundary. Sci. Nat. 2017, 104, 87. [Google Scholar] [CrossRef]
- Worthy, T.H.; Degrange, F.J.; Handley, W.D.; Lee, M.S.Y. The Evolution of Giant Flightless Birds and Novel Phylogenetic Relationships for Extinct Fowl (Aves, Galloanseres). R. Soc. Open Sci. 2017, 4, 170975. [Google Scholar] [CrossRef]
- Mayr, G.; De Pietri, V.L.; Scofield, R.P.; Worthy, T.H. On the Taxonomic Composition and Phylogenetic Affinities of the Recently Proposed Clade Vegaviidae Agnolín et al., 2017–Neornithine Birds from the Upper Cretaceous of the Southern Hemisphere. Cretac. Res. 2018, 86, 178–185. [Google Scholar] [CrossRef]
- McLachlan, S.M.S.; Kaiser, G.W.; Longrich, N.R. Maaqwi cascadensis: A Large, Marine Diving Bird (Avialae: Ornithurae) from the Upper Cretaceous of British Columbia, Canada. PLoS ONE 2017, 12, e0189473. [Google Scholar] [CrossRef]
- Clarke, J.A.; Chatterjee, S.; Li, Z.; Riede, T.; Agnolin, F.; Goller, F.; Isasi, M.P.; Martinioni, D.R.; Mussel, F.J.; Novas, F.E. Fossil Evidence of the Avian Vocal Organ from the Mesozoic. Nature 2016, 538, 502–505. [Google Scholar] [CrossRef]
- Mayr, G. A Partial Skeleton of a New Fossil Loon (Aves, Gaviiformes) from the Early Oligocene of Germany with Preserved Stomach Content. J. Ornithol. 2004, 145, 281–286. [Google Scholar] [CrossRef]
- Cracraft, J. Phylogenetic Relationships and Monophyly of Loons, Grebes, and Hesperonithiformes Birds, with Comments on the Early History of Birds. Syst. Biol. 1982, 31, 35–56. [Google Scholar] [CrossRef]
- Bourdon, E. Osteological Evidence for Sister Group Relationship between Pseudo-Toothed Birds (Aves: Odontopterygiformes) and Waterfowls (Anseriformes). Naturwissenschaften 2005, 92, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Wetmore, A. Fossil Birds from the Green River Deposits of Eastern Utah. Ann. Carnegie Mus. 1926, 16, 391–402. [Google Scholar] [CrossRef]
- Feduccia, A.; McGrew, P.O. A Flamingo-like Wader from the Eocene of Wyoming. Contrib. Geol. 1974, 13, 49–61. [Google Scholar]
- Olson, S.L.; Parris, D.C. The Cretaceous Birds of New Jersey. Smithson. Contrib. Zool. 1987, 63, 1–22. [Google Scholar] [CrossRef]
- Olson, S.L. A Giant Presbyornis (Aves: Anseriformes) and Other Birds from the Paleocene Aquia Formation of Maryland and Virginia. Proc. Biol. Soc. Wash 1994, 107, 429–435. [Google Scholar]
- Boles, W.E.; Finch, M.A.; Hofheins, R.H.; Walters, M.; Rich, T.H. A Fossil Stone-Curlew (Aves: Burhinidae) from the Late Oligocene/Early Miocene of South Australia. In Proceedings of the 8th International Meeting of the Society of Avian Paleontology and Evolution, Wien, Austria, 11–16 June 2012; pp. 43–62. [Google Scholar]
- Olson, S.L. The Anseriform Relationships of Anatalavis Olson and Parris (Anseranatidae), with a New Species from the Lower Eocene London Clay. Smithson. Contrib. Zool. 1999, 89, 231–243. [Google Scholar]
- Shufeldt, R.W. Fossil Birds in the Marsh Collection of Yale University. Trans. Conn. Acad. Arts Sci. 1915, 19, 1–110. [Google Scholar]
- Mourer-Chauviré, C. [Review of] Cenozoic Birds of the World, Part 1: Europe by Jiri Mlíkovský, 2002. Auk 2004, 121, 623–627. [Google Scholar] [CrossRef]
- Worthy, T.H.; Scanlon, J.D. An Oligo-Miocene Magpie Goose (Aves: Anseranatidae) from Riversleigh, Northwestern Queensland, Australia. J. Vertebr. Paleontol. 2009, 29, 205–211. [Google Scholar] [CrossRef]
- Zelenkov, N. The Oldest Diving Anseriform Bird from the Late Eocene of Kazakhstan and the Evolution of Aquatic Adaptations in the Intertarsal Joint of Waterfowl. Acta Palaeontol. Pol. 2020, 65, 733–742. [Google Scholar] [CrossRef]
- Mourer-Chauviré, C.; Berthet, D.; Hungueney, M. The Late Oligocene Birds of the Créchy Quarry (Allier, France), with a Description of Two New Genera (Aves: Pelecaniformes: Phalacrocoracidae, and Anseriformes: Anseranatidae). Senckenberg. Lethaea 2004, 84, 303–315. [Google Scholar] [CrossRef]
- Mayr, G.; Smith, R. Ducks, Rails, and Limicoline Waders (Aves: Anseriformes, Gruiformes, Charadriiformes) from the Lowermost Oligocene of Belgium. Geobios 2001, 34, 547–561. [Google Scholar] [CrossRef]
- Milne-Edwards, A. Sur La Distribution Géologique des Oiseaux Fossiles et Description de Quelques Espèces Nouvelles. C. R. Séances Sci. Acad. Sci. 1863, 56, 1219–1222. [Google Scholar]
- Milne-Edwards, A. Recherches Anatomiques et Paleontologiques Pour Servir a l’Histoire Des Oiseaux Fossiles de La France; Victor Masson: Paris, France, 1867. [Google Scholar]
- Mayr, G.; De Pietri, V.L.; Love, L.; Mannering, A.; Scofield, R.P. Oldest, Smallest and Phylogenetically Most Basal Pelagornithid, from the Early Paleocene of New Zealand, Sheds Light on the Evolutionary History of the Largest Flying Birds. Pap. Palaeontol. 2021, 7, 217–233. [Google Scholar] [CrossRef]
- Benito, J.; Kuo, P.-C.; Widrig, K.E.; Jagt, J.W.M.; Field, D.J. Cretaceous Ornithurine Supports a Neognathous Crown Bird Ancestor. Nature 2022, 612, 100–105. [Google Scholar] [CrossRef]
- Andors, A.V. Reappraisal of the Eocene Groundbird Diatryma (Aves: Anserimorphae). Nat. His. Mus. Los Angeles Sci. Ser. 1992, 36, 109–125. [Google Scholar]
- Angst, D.; Lécuyer, C.; Amiot, R.; Buffetaut, E.; Fourel, F.; Martineau, F.; Legendre, S.; Abourachid, A.; Herrel, A. Isotopic and Anatomical Evidence of an Herbivorous Diet in the Early Tertiary Giant Bird Gastornis. Implications for the Structure of Paleocene Terrestrial Ecosystems. Naturwissenschaften 2014, 101, 313–322. [Google Scholar] [CrossRef]
- Andors, A.V. Giant Groundbirds of North America (Aves, Diatrymidae). Ph.D. Thesis, Columbia University, New York, NY, USA, 1988. [Google Scholar]
- Angst, D. Successeurs des Dinosaures? Paléobiologie et Paléoécologie d’un oiseau géant terrestre du Paléogène. Ph.D. Thesis, Université Claude Bernard-Lyon 1, Lion, France, 2014. [Google Scholar]
- Buffetaut, E. The Giant Bird Gastornis in Asia: A Revision of Zhongyuanus Xichuanensis Hou, 1980, from the Early Eocene of China. Paleontol. J. 2013, 47, 1302–1307. [Google Scholar] [CrossRef]
- Murray, P.F.; Vickers-Rich, P. Magnificent Mihirungs: The Colossal Flightless Birds of the Australian Dreamtime; Indiana University Press: Bloomington, IN, USA, 2004. [Google Scholar]
- Mayr, G.; Smith, T. Bony-Toothed Birds (Aves: Pelagornithidae) from the Middle Eocene of Belgium. Palaeontology 2010, 53, 365–376. [Google Scholar] [CrossRef]
- Mayr, G.; Rubilar-Rogers, D. Osteology of a New Giant Bony-Toothed Bird from the Miocene of Chile, with a Revision of the Taxonomy of Neogene Pelagornithidae. J. Vertebr. Paleontol. 2010, 30, 1313–1330. [Google Scholar] [CrossRef]
- Mayr, G. Cenozoic Mystery Birds–on the Phylogenetic Affinities of Bony-toothed Birds (Pelagornithidae). Zool. Scr. 2011, 40, 448–467. [Google Scholar] [CrossRef]
- Baumel, J.J.; Witmer, L.M. Osteologica. In Handbook of Avian Anatomy: Nomina Anatomica Avium; King, A.S., Breazile, J.E., Evans, H.E., Vanden Berge, J.C., Eds.; Publications of the Nuttall Ornithological Club: Cambridge, MA, USA, 1993; pp. 45–132. [Google Scholar]
- Zusi, R.L.; Livezey, B.C. Homology and Phylogenetic Implications of Some Enigmatic Cranial Features in Galliform and Anseriform Birds. Ann. Carnegie Mus. 2000, 69, 157–193. [Google Scholar] [CrossRef]
- Livezey, B.C.; Zusi, R.L. Phylogeny of Neornithes. Bull. Carnegie Mus. Nat. Hist. 2006, 37, 1–544. [Google Scholar] [CrossRef]
- Howard, H. The Avifauna of Emryville Shellmound; University of California Press: Berkeley, CA, USA, 1929; Volume 32. [Google Scholar]
- Bock, W.J. Secondary Articulation of the Avian Mandible. Auk 1960, 77, 19–55. [Google Scholar] [CrossRef]
- Ballmann, P. Les Oiseaux Miocènes de La Grive-Saint-Alban (Isère). Geobios 1969, 2, 157–204. [Google Scholar] [CrossRef]
- Dyke, G.J.; Gulas, B.E. The Fossil Galliform Bird Paraortygoides from the Lower Eocene of the United Kingdom. Am. Mus. Novit. 2002, 3360, 1–14. [Google Scholar] [CrossRef]
- Bourdon, E.; De Ricqlès, A.; Cubo, J. A New Transantarctic Relationship: Morphological Evidence for a Rheidae-Dromaiidae-Casuariidae Clade (Aves, Palaeognathae, Ratitae). Zool. J. Linn. Soc. 2009, 156, 641–663. [Google Scholar] [CrossRef]
- Worthy, T.; Scofield, R. Twenty-First Century Advances in Knowledge of the Biology of Moa (Aves: Dinornithiformes): A New Morphological Analysis and Moa Diagnoses Revised. N. Z. J. Zool. 2012, 39, 87–153. [Google Scholar] [CrossRef]
- Zinoviev, A.V. Notes on Pelvic and Hindlimb Myology and Syndesmology of Emeus Crassus and Dinornis Robustus (Aves: Dinornithiformes). In Proceedings of the 8th International Meeting of the Society of Avian Paleontology and Evolution, Wien, Austria, 11–16 June 2012; pp. 253–278. [Google Scholar]
- Ellerby, D.J.; Marsh, R.L. The Energetic Costs of Trunk and Distal-Limb Loading during Walking and Running in Guinea Fowl Numida Meleagris. J Exp Biol 2006, 209, 2064–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas, L.P.; Main, R.P.; Hutchinson, J.R. Ontogenetic Scaling Patterns and Functional Anatomy of the Pelvic Limb Musculature in Emus (Dromaius Novaehollandiae). PeerJ 2014, 2, e716. [Google Scholar] [CrossRef]
- Clifton, G.T.; Carr, J.A.; Biewener, A.A. Comparative Hindlimb Myology of Foot-Propelled Swimming Birds. J. Anat. 2018, 232, 105–123. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Clarke, J.A.; Eliason, C.M.; Stidham, T.A.; Deng, T.; Zhou, Z. Vocal Specialization through Tracheal Elongation in an Extinct Miocene Pheasant from China. Sci. Rep. 2018, 8, 8099. [Google Scholar] [CrossRef]
- Swofford, D.L. Phylogenetic Analysis Using Parsimony (* and Other Methods). Available online: https://paup.phylosolutions.com (accessed on 27 January 2022).
- Livezey, B.C. A Phylogenetic Analysis of Recent Anseriform Genera Using Morphological Characters. Auk 1986, 103, 737–754. [Google Scholar] [CrossRef]
- Livezey, B.C. Erratum–A Phylogenetic Analysis of Basal Anseriformes, the Fossil Presbyornis, and the Interordinal Relationships of Waterfowl. Zool. J. Linn. Soc. 1998, 124, 397–398. [Google Scholar] [CrossRef]
- Acosta Hospitaleche, C.; Worthy, T.H. New Data on the Vegavis Iaai Holotype from the Maastrichtian of Antarctica. Cretac. Res. 2021, 124, 104818. [Google Scholar] [CrossRef]
- Smith, M.E.; Chamberlain, K.R.; Singer, B.S.; Carroll, A.R. Eocene Clocks Agree: Coeval 40Ar/39Ar, U-Pb, and Astronomical Ages from the Green River Formation. Geology 2010, 38, 527–530. [Google Scholar] [CrossRef]
- Allentoft, M.E.; Heller, R.; Oskam, C.L.; Lorenzen, E.D.; Hale, M.L.; Gilbert, M.T.P.; Jacomb, C.; Holdaway, R.N.; Bunce, M. Extinct New Zealand Megafauna Were Not in Decline before Human Colonization. Proc. Natl. Acad. Sci. USA 2014, 111, 4922–4927. [Google Scholar] [CrossRef]
- Solórzano, A.; Rincón, A.D. The Earliest Record (Early Miocene) of a Bony-Toothed Bird from South America and a Reexamination of Venezuelan Pelagornithids. J. Vertebr. Paleontol. 2015, 35, e995188. [Google Scholar] [CrossRef]
- Collinson, M.E.; Adams, N.F.; Manchester, S.R.; Stull, G.W.; Herrera, F.; Smith, S.Y.; Andrew, M.J.; Kenrick, P.; Sykes, D. X-Ray Micro-Computed Tomography (Micro-CT) of Pyrite-Permineralized Fruits and Seeds from the London Clay Formation (Ypresian) Conserved in Silicone Oil: A Critical Evaluation. Botany 2016, 94, 697–711. [Google Scholar] [CrossRef] [Green Version]
- Benito, J.; Chen, A.; Wilson, L.E.; Bhullar, B.-A.S.; Burnham, D.; Field, D.J. 40 New Specimens of Ichthyornis Provide Unprecedented Insight into the Postcranial Morphology of Crownward Stem Group Birds. PeerJ 2022, 10, e13919. [Google Scholar] [CrossRef]
- Welch, J.L.; Foreman, B.Z.; Malone, D.; Craddock, J. Provenance of Early Paleogene Strata in the Bighorn Basin (Wyoming, USA): Implications for Laramide Tectonism and Basin-Scale Stratigraphic Patterns. In Tectonic Evolution of the Sevier-Laramide Hinterland, Thrust Belt, and Foreland, and Postorogenic Slab Rollback (180-20 Ma); GSA Special Papers: Boulder, CO, USA, 2022; Volume 555, ISBN 978-0-8137-9555-3. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarisation in Bayesian Phylogenetics Using Tracer 1.7.2. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Kimball, R.T.; Pandey, A.; Hosner, P.A.; Braun, M.J.; Hackett, S.J.; Han, K.-L.; Harshman, J.; Huddleston, C.J.; Kingston, S.; et al. Why Do Phylogenomic Data Sets Yield Conflicting Trees? Data Type Influences the Avian Tree of Life More than Taxon Sampling. Syst. Biol. 2017, 66, 857–879. [Google Scholar] [CrossRef]
- Burleigh, J.G.; Kimball, R.T.; Braun, E.L. Building the Avian Tree of Life Using a Large-Scale, Sparse Supermatrix. Mol. Phylogenet. Evol. 2015, 84, 53–63. [Google Scholar] [CrossRef]
- Goldman, N.; Anderson, J.P.; Rodrigo, A.G. Likelihood-Based Tests of Topologies in Phylogenetics. Syst. Biol. 2000, 49, 652–670. [Google Scholar] [CrossRef]
- Dzerzhinsky, F.Y. Evidence for Common Ancestry of the Galliformes and Anseriformes. Cour. Forsch. Senckenb. 1995, 181, 325–336. [Google Scholar]
- O’Connor, P.M. Pulmonary Pneumaticity in the Postcranial Skeleton of Extant Aves: A Case Study Examining Anseriformes. J. Morphol. 2004, 261, 141–161. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, P.M. Postcranial Pneumaticity: An Evaluation of Soft-Tissue Influences on the Postcranial Skeleton and the Reconstruction of Pulmonary Anatomy in Archosaurs. J. Morphol. 2006, 267, 1199–1226. [Google Scholar] [CrossRef] [PubMed]
- De Mendoza, R.S.; Gómez, R.O.; Tambussi, C.P. The Lacrimal/Ectethmoid Region of Waterfowl (Aves, Anseriformes): Phylogenetic Signal and Major Evolutionary Patterns. J. Morphol. 2020, 281, 1486–1500. [Google Scholar] [CrossRef]
- Mayr, G. On the Occurrence of Lateral Openings and Fossae (Pleurocoels) in the Thoracic Vertebrae of Neornithine Birds and Their Functional Significance. VZ 2021, 71, 453–463. [Google Scholar] [CrossRef]
- Secord, R.; Gingerich, P.D.; Smith, M.E.; Clyde, W.C.; Wilf, P.; Singer, B.S. Geochronology and Mammalian Biostratigraphy of Middle and Upper Paleocene Continental Strata, Bighorn Basin, Wyoming. Am. J. Sci. 2006, 306, 211–245. [Google Scholar] [CrossRef]
- Secord, R. The Tiffanian Land-Mammal Age (Middle and Late Paleocene) in the Northern Bighorn Basin, Wyoming. Uni. Mich. Pap. Paleo. 2008, 35, 1–192. [Google Scholar]
- Bowen, G.J.; Maibauer, B.J.; Kraus, M.J.; Röhl, U.; Westerhold, T.; Steimke, A.; Gingerich, P.D.; Wing, S.L.; Clyde, W.C. Two Massive, Rapid Releases of Carbon during the Onset of the Palaeocene–Eocene Thermal Maximum. Nat. Geosci. 2014, 8, 44–47. [Google Scholar] [CrossRef]
- Field, D.J.; Lynner, C.; Brown, C.; Darroch, S.A.F. Skeletal Correlates for Body Mass estimation in Modern and Fossil Flying Birds. PLoS ONE 2013, 8, e82000. [Google Scholar] [CrossRef] [PubMed]
- Houde, P.; Olson, S.L. A Radiation of Coly-like Birds from the Eocene of North America (Aves: Sandcoleiformes, New Order). Nat. Hist. Mus. Los Angeles Cty. Sci. Ser. 1992, 36, 137–160. [Google Scholar]
- Van Tyne, J.; Berger, A.J. Fundamentals of Ornithology; Dover Publications: New York, NY, USA, 1971. [Google Scholar]
- Campbell, B.; Lack, E. A Dictionary of Birds; British Ornithologist’s Union, Buteo Books: Vermillion, SD, USA, 1985; ISBN 0-931130-12-3. [Google Scholar]
- Buchheim, H.P.; Biaggi, R.E.; Cushman, R.A. Stratigraphy and Interbasinal Correlations Between Fossil and the Green River Basin, Wyoming. In Stratigraphy and Paleolimnology of the Green River Formation, Western USA; Smith, M.E., Carroll, A.R., Eds.; Syntheses in Limnogeology; Springer: Dordrecht, The Netherlands, 2015; Volume 1, pp. 127–151. ISBN 978-94-017-9905-8. [Google Scholar]
- Grande, L. Birds. In The Lost World of Fossil Lake; University of Chicago Press: Chicago, IL, USA, 2013; pp. 215–257. [Google Scholar]
- Puttick, M.N.; O’Reilly, J.E.; Pisani, D.; Donoghue, P.C.J. Probabilistic Methods Outperform Parsimony in the Phylogenetic Analysis of Data Simulated without a Probabilistic Model. Palaeontology 2019, 62, 1–17. [Google Scholar] [CrossRef]
- King, B. Bayesian Tip-Dated Phylogenetics in Paleontology: Topological Effects and Stratigraphic Fit. Syst. Biol. 2021, 70, 283–294. [Google Scholar] [CrossRef]
- Feng, S.; Bai, M.; Rivas-González, I.; Li, C.; Liu, S.; Tong, Y.; Yang, H.; Chen, G.; Xie, D.; Sears, K.E.; et al. Incomplete Lineage Sorting and Phenotypic Evolution in Marsupials. Cell 2022, 185, 1646–1660.e18. [Google Scholar] [CrossRef]
- Marsh, O.C. Odontornithes: Monograph on the Extinct Toothed Birds of North America; United States Geological Exploration of the Fortieth Parallel; Government Printing Office: Washington, DC, USA, 1880. [Google Scholar]
- Sanz, J.L.; Chiappe, L.M.; Buscalioni, A.D. The Osteology of Concornis Lacustris (Aves: Enantiomithes) from the Lower Cretaceous of Spain and a Reexamination of Its Phylogenetic Relationships. Am. Mus. Novit. 1995, 3133, 1–23. [Google Scholar]
- Chiappe, L.M. Late Cretaceous Birds of Southern South America: Anatomy and Systematics of Enantiornithes and Patagopteryx deferrariisi. Mün. Geo. Abh. 1996, 30, 203–244. [Google Scholar]
- Zelenkov, N.V.; Stidham, T.A. Possible Filter-Feeding in the Exinct Presbyornis and the Evolution of Anseriformes (Aves). Zool. Z. 2018, 97, 943–956. [Google Scholar] [CrossRef]
- Rose, K.D. The Clarkforkian Land Mammal Age. Uni. Mich. Papers in Paleo. 1981, 26, 1–197. [Google Scholar]
- Houde, P. Paleognathous Birds from the Early Tertiary of the Northern Hemisphere. In Publications of the Nuttall Ornithological Club; Paynter, R.A., Jr., Ed.; Nuttall Ornithological Club: Cambridge, MA, USA, 1988. [Google Scholar]
- Bowen, G.J.; Bloch, J.I. Petrography and geochemistry of floodplain limestones from the Clarks Fork Basin, Wyoming, USA: Carbonate deposition and fossil accumulation on a Paleocene-Eocene floodplain. J. Sediment. Res. 2002, 72, 46–58. [Google Scholar] [CrossRef]
- du Toit, C.J.; Chinsamy, A.; Cunningham, S.J. Cretaceous Origins of the Vibrotactile Bill-Tip Organ in Birds. Proc. R. Soc. B 2020, 287, 20202322. [Google Scholar] [CrossRef] [PubMed]
- Mayr, G.; Gingerich, P.D.; Smith, T. Calcardea Junnei Gingerich, 1987 from the Late Paleocene of North America Is Not a Heron, but Resembles the Early Eocene Indian Taxon Vastanavis Mayr et al., 2007. J. Paleontol. 2019, 93, 359–367. [Google Scholar] [CrossRef]
- Mayr, G.; Gingerich, P.D.; Smith, T. Skeleton of a New Owl from the Early Eocene of North America (Aves, Strigiformes) with an Accipitrid-like Foot Morphology. J. Vertebr. Paleontol. 2020, 40, e1769116. [Google Scholar] [CrossRef]
- Degrange, F.J. Hind Limb Morphometry of Terror Birds (Aves, Cariamiformes, Phorusrhacidae): Functional Implications for Substrate Preferences and Locomotor Lifestyle. Earth Env. Sci. Trans. R. Soc. Edinb. 2017, 106, 257–276. [Google Scholar] [CrossRef]
- IUCN Red List 2022-2 White Backed Duck. Available online: https://www.iucnredlist.org/species/22679785/92830036#habitat-ecology (accessed on 23 January 2023).
- Zweers, G.A. Structure, movement, and myography of the feeding apparatus of the mallard (Anas platyrhynchos L.) A study in functional anatomy. Neth. J. Zool. 1973, 24, 323–467. [Google Scholar] [CrossRef]
- Clyde, W.C.; Gingerich, P.D. Mammalian Community Response to the Latest Paleocene Thermal Maximum: An Isotaphonomic Study in the Northern Bighorn Basin, Wyoming. Geology 1998, 26, 1011. [Google Scholar] [CrossRef]
- Woodburne, M.O.; Gunnell, G.F.; Stucky, R.K. Climate Directly Influences Eocene Mammal Faunal Dynamics in North America. Proc. Natl. Acad. Sci. USA 2009, 106, 13399–13403. [Google Scholar] [CrossRef] [PubMed]
- Hooker, J.J.; Collinson, M.E. Mammalian Faunal Turnover across the Paleocene-Eocene Boundary in NW Europe: The Roles of Displacement, Community Evolution and Environment. Austrian J. Earth Sci. 2012, 105, 17–28. [Google Scholar]
- Widlansky, S.J.; Secord, R.; Snell, K.E.; Chew, A.E.; Clyde, W.C. Carbon Isotope Stratigraphy and Mammal Turnover during Post-PETM Hyperthermals. Clim. Past 2022, 18, 681–712. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Houde, P.; Dickson, M.; Camarena, D. Basal Anseriformes from the Early Paleogene of North America and Europe. Diversity 2023, 15, 233. https://doi.org/10.3390/d15020233
Houde P, Dickson M, Camarena D. Basal Anseriformes from the Early Paleogene of North America and Europe. Diversity. 2023; 15(2):233. https://doi.org/10.3390/d15020233
Chicago/Turabian StyleHoude, Peter, Meig Dickson, and Dakota Camarena. 2023. "Basal Anseriformes from the Early Paleogene of North America and Europe" Diversity 15, no. 2: 233. https://doi.org/10.3390/d15020233