
The Facet of Human Impact: Solenopsis invicta Buren, 1972 Spreading around the Atlantic Forest
 , , , ,
, , , ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
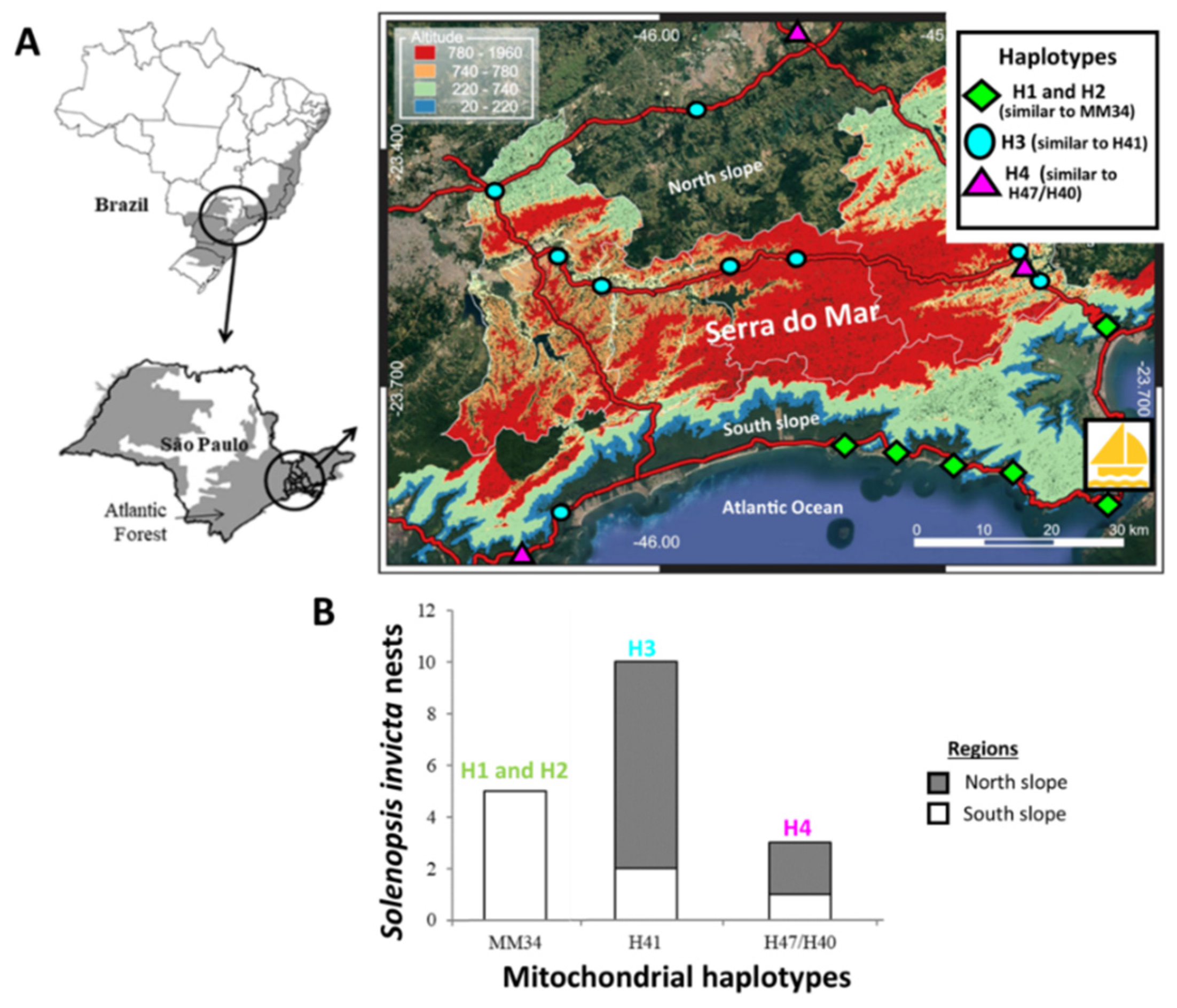
2.1. Collection and Experimental Design
2.2. Molecular Identification
2.3. Molecular Diversity and Bayesian Inference
2.4. Correlation between Genetic and Geographic Distances
3. Results
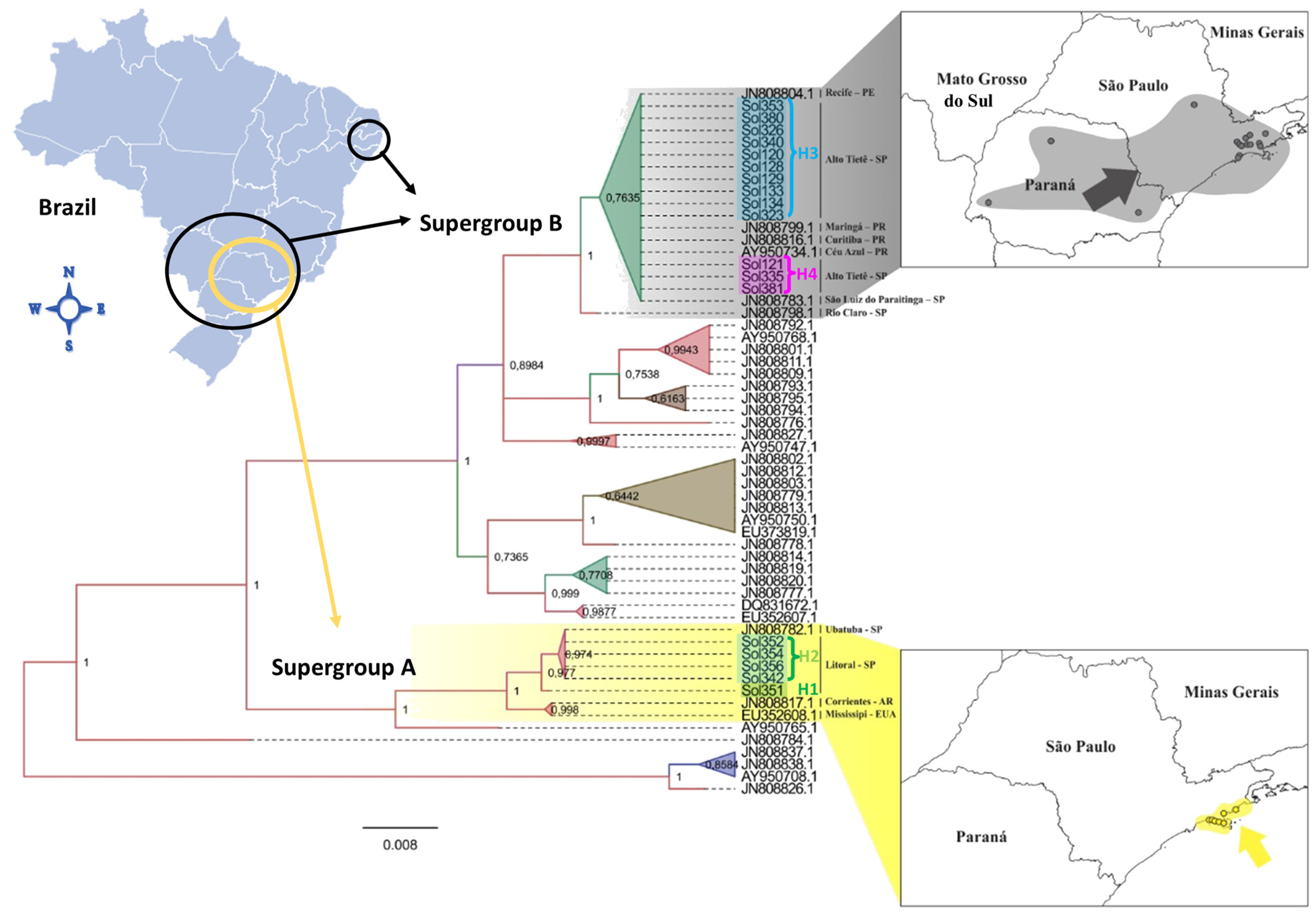
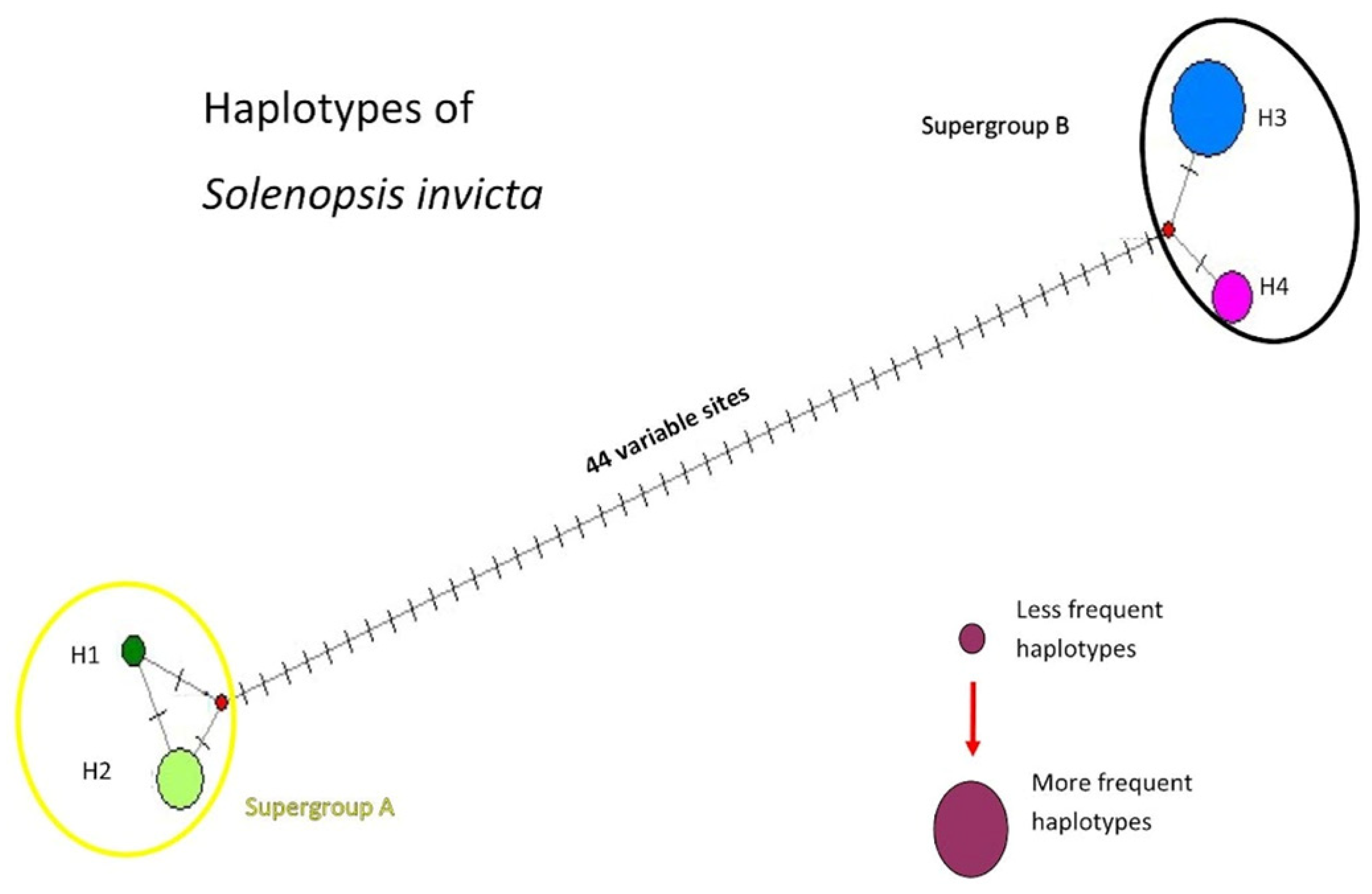
3.1. Molecular Identification and Diversity
3.2. Bayesian Inference and Correlation between Genetic and Geographic Distances
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bolton, B. An Online Ctalog of the Ants of the World. Available online: https://antcat.org (accessed on 6 June 2022).
- Thompson, C.R. The thief ants, Solenopsis molesta group of Florida (Hymenoptera: Formicidae). Fla Entomol. 1989, 72, 268–283. [Google Scholar] [CrossRef]
- Andersen, A.N.; Brault, A. Exploring a new biodiversity frontier: Subterranean ants in northern Australia. Biodiv. Conserv. 2010, 19, 2741–2750. [Google Scholar] [CrossRef]
- Ohyama, L.; King, J.R.; Jenkins, D.G. Diversity and distribution of Solenopsis (Hymenoptera: Formicidae) thief ants belowground. Myrmecol. News 2018, 27, 47–57. [Google Scholar]
- Hernández, V.; Castaño-Meneses, G. Checklist, biological notes and distribution of ants (Hymenoptera: Formicidae) from Barranca de Metztitlán Biosphere Reserve, Hidalgo, Mexico. Sociobiology 2010, 56, 397–434. [Google Scholar]
- Pitts, J.P.; Camacho, G.P.; Gotzek, D.; Mchugh, J.V.; Ross, K.G. Revision of the fire ants of the Solenopsis saevissima species-group (Hymenoptera: Formicidae). Proc. Entomol. Soc. Wash. 2018, 120, 308–411. [Google Scholar] [CrossRef]
- Gusmão, F.A.; Harakava, R.; Campos, A.E.C. Fire-ants of the Solenopsis saevissima species-group (Hymenoptera: Formicidae) nesting in parks in the city of São Paulo: Identification based on mtDNA sequences and morphological characters. Sociobiology 2010, 56, 353–362. [Google Scholar]
- Tschinkel, W.R. The organization of foraging in the fire ant, Solenopsis invicta. J. Insect Sci. 2011, 2011, 11–26. [Google Scholar] [CrossRef]
- Dejean, A.; Céréghino, R.; Leponce, M.; Rossi, V.; Roux, O.; Compin, A.; Delabie, J.H.C.; Corbara, B. The fire ant Solenopsis saevissima and habitat disturbance alter ant communities. Biol. Conserv. 2015, 187, 145–153. [Google Scholar] [CrossRef]
- Chialvo, P.; Gotzek, D.; Shoemaker, D.; Ross, K.G. Genetic analysis reveal cryptic diversity in the native North American fire ants (Hymenoptera: Formicidae: Solenopsis). Syst. Entomol. 2018, 43, 109–122. [Google Scholar] [CrossRef]
- Nagatani, V.H.; Ramalho, M.O.; Alves, J.M.C.; Souza, R.F.; Kayano, D.Y.; Silva, N.S.; Silva, O.G.M.; Harakava, R.; Bueno, O.C.; Hilsdorf, A.W.S.; et al. Impact of native vegetation cover near crops on the occurrence and molecular diversity of fire ants. Agric. Forest Entomol. 2022, 25, 1–11. [Google Scholar] [CrossRef]
- Wang, W.; Feng, X.; Chen, X. Biological invasion and coexistence in intraguild predation. J. Appl. Mathem. 2013, 2013, 1–12. [Google Scholar] [CrossRef]
- Wang, L.; Xu, Y.J.; Zeng, L.; Lu, Y.Y. A review of the impact of the red imported fire ant Solenopsis invicta Buren on biodiversity in South China. J. Integr. Agric. 2018, 17, 1–10. [Google Scholar] [CrossRef]
- Ascunce, M.S.; Yang, C.-C.; Oakey, J.; Calcaterra, L.; Wu, W.-J.; Shih, C.-J.; Goudet, J.; Ross, K.G.; Shoemaker, D. Global invasion history of the fire ant Solenopsis invicta. Science 2011, 331, 1066–1068. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hou, Y.-M. Invasion history of Solenopsis invicta (Hymenoptera: Formicidae) in Fujian, China based on mitochondrial DNA and its implications in development of a control strategy. Insect Sci. 2014, 21, 493–498. [Google Scholar] [CrossRef]
- Vinson, S.B.; Sorensen, A.A. Imported Fire Ants: Life History and Impact; Texas Department of Agriculture: Austin, TX, USA, 1986; 28 p.
- Seebens, H.; Essl, F.; Dawson, W.; Fuentes, N.; Moser, D.; Pergl, J.; Pysek, P.; Kleunen, M.V.; Weber, E.; Winter, M.; et al. Global trade will accelerate plant invasions in emerging economies under climate change. Glob. Change Biol. 2015, 21, 4128–4140. [Google Scholar] [CrossRef]
- Shoemaker, D.D.; Deheer, C.J.; Krieger, M.J.; Ross, K.G. Population genetics of the invasive fire ant Solenopsis invicta (Hymenoptera: Formicidae) in the United States. Ann. Entomol. Soc. Am. 2006, 99, 1213–1233. [Google Scholar] [CrossRef]
- Morrison, L.W.; Porter, S.D.; Daniels, E.; Korzukhin, M.D. Potential global range expansion of the invasive fire ant, Solenopsis invicta. Biol. Invasions 2004, 6, 183–191. [Google Scholar] [CrossRef]
- Benkman, C.W.; Siepielski, A.M.; Parchman, T.L. The local introduction of strongly interacting species and the loss of geographic variation in species and species interactions. Mol. Ecol. 2008, 17, 395–404. [Google Scholar] [CrossRef]
- Dyer, E.E.; Cassey, P.; Redding, D.W.; Collen, B.; Franks, V.; Gaston, K.J.; Jones, K.E.; Kark, S.; Orme, C.D.L.; Blackburn, T.M. The global distribution and drivers of alien bird species richness. PLoS Biol. 2017, 15, 1–25. [Google Scholar] [CrossRef]
- Massoni, A.; Coppi, A.; Balzani, P.; Frizzi, F.; Fani, R.; Zaccaroni, M.; Santini, G. Assessing molecular diversity in native and introduced populations of red wood ant Formica paralugubris. Animals 2022, 12, 3165. [Google Scholar] [CrossRef]
- Henshaw, M.T.; Kunzman, N.; Vanderwoude, C.; Sanetra, M.; Crozier, R.H. Population genetics and history of the introduced fire ant Solenopsis invicta Buren (Hymenoptera: Formicidae), in Australia. Aust. J. Entomol. 2005, 44, 37–44. [Google Scholar] [CrossRef]
- Shoemaker, D.D.; Ahrens, M.E.; Ross, K.G. Molecular phylogeny of fire ants of the Solenopsis saevissima species-group based on mtDNA sequences. Mol. Phylogenet. Evol. 2006, 38, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.C.; Shoemaker, D.D.; Wu, W.J.; Shih, C.J. Population genetic structure of the red imported fire ant, Solenopsis invicta, in Taiwan. Insectes Soc. 2008, 55, 54–65. [Google Scholar] [CrossRef]
- Caldera, E.J.; Ross, K.G.; Deheer, C.J.; Shoemaker, D.D. Putative native source of the invasive fire ant Solenopsis invicta in the USA. Biol. Invasions 2008, 10, 1457–1479. [Google Scholar] [CrossRef]
- Koehler, A.; Galvão, F.; Longhi, S.J. Floresta ombrófila densa altomontana: Aspectos florísticos e estruturais de diferentes trechos na serra do mar, PR. Ciênc. Florest. 2002, 12, 27–39. [Google Scholar] [CrossRef]
- CPTEC-INPE. Centro de Previsão de Tempo e Estudos Climáticos. 2022. Available online: http://clima.cptec.inpe.br/ (accessed on 6 June 2022).
- Joly, A.C.; Metzger, J.P.; Tabarelli, M. Experiences from the Brazilian Atlantic Forest: Ecological findings and conservation initiatives. New Phytol. 2014, 204, 459–473. [Google Scholar] [CrossRef]
- IBGE. Instituto Brasileiro de Geografia e Estatística, Censos Demográficos. 2020. Available online: https://www.ibge.gov.br/ (accessed on 20 April 2020).
- Bueno, O.C. Criação de formigas em laboratório. In Formigas em Ambientes Urbanos no Brasil; Bueno, O.C., Campos, A.E.C., Morini, M.S.C., Eds.; Editora Canal6: Bauru, Brazil, 2018; pp. 125–142. [Google Scholar]
- Pitts, J.P.; Mchughi, J.V.; Ross, K.G. Cladistic analysis of the fire ants of the Solenopsis saevissima species-group (Hymenoptera: Formicidae). Zool. Scr. 2005, 34, 493–505. [Google Scholar] [CrossRef]
- Souza-Campana, D.R.; Wazema, C.T.; Magalhães, F.S.; Silva, N.S.; Nagatani, V.H.; Suguituru, S.S.; Goto, M.A.; Morini, M.S.C. Coleção de referência do Laboratório de Mirmecologia do Alto Tietê, São Paulo, Brasil: Status atual e perspectivas. Bol. Mus. Para. Emílio Goeldi. Sér. Ciênc. Nat. 2020, 15, 317–336. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; De Waard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; De Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Biol. Sci. 2003, 270, 596–599. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of life system (www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef]
- Ross, K.G.; Shoemaker, D.D. Nuclear and mitochondrial genetic structure in two social forms of the fire ants Solenopsis invicta: Insights into transitions to an alternate social organization. Heredity 1997, 78, 590–602. [Google Scholar] [CrossRef]
- Ahrens, M.E.; Ross, K.G.; Shoemaker, D.D. Phylogeographic structure of the fire ant Solenopsis invicta in its native south American range: Roles of natural barriers and habitat connectivity. Evolution 2005, 59, 1733–1743. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.; Souza, R.F.; Bueno, O.C. Presence and distribution of the endosymbiont Wolbachia among Solenopsis spp. (Hymenoptera: Formicidae) from Brazil and its evolutionary history. J. Invertebr. Pathol. 2012, 109, 287–296. [Google Scholar] [CrossRef]
- Valles, S.M.; Porter, S.D. Identification of polygyne and monogyne fire ant colonies (Solenopsis invicta) by multiplex PCR of Gp-9 alleles. Insectes Soc. 2003, 50, 199–200. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1990, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 3, 870–1874. [Google Scholar] [CrossRef]
- Rozas, J.; Sánchez-Delbarrio, J.C.; Messeguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest Version 2. Program Distributed by the Author. Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Martins, C.; Souza, R.F.; Bueno, O.C. Molecular characterization of fire ants, Solenopsis spp., from Brazil based on analysis of mtDNA gene cytochrome oxidase I. J. Insect Sci. 2014, 14, 50. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Bivand, R.; Keitt, T.; Rowlingso, N.B. Rgdal: Bindings for the Geospatial Data Abstraction Library, R Package Version 0.8-11; 2013. Available online: https://cran.r-project.org/web/packages/rgdal/index.html (accessed on 6 June 2022).
- R Core Team. R: A Language and Environment for Statistical Computing, Vienna. 2021. Available online: http://www.R-project.org. (accessed on 15 January 2018).
- Kimura, M.A. Simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 10, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package, R Package Version 2.3-0. 2022. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 6 June 2022).
- Ross, K.G.; Krieger, M.J.; Keller, L.; Shoemaker, D.D. Genetic variation and structure in native populations of the fire ant Solenopsis invicta: Evolutionary and demographic implications. Biol. J. Linn. Soc. 2007, 92, 541–560. [Google Scholar] [CrossRef]
- Fowler, H.G.; Bernardi, J.V.E.; Di Romagnano, L.F.T. Community structure and Solenopsis invicta in São Paulo. In Applied Myrmecology; Vander Meer, R.K., Jaffé, K., Cedeno, A., Eds.; Taylor & Francis Group: London, UK, 1990; pp. 199–207. [Google Scholar]
- Philpott, S.M.; Perfecto, I.; Armbrecht, I.; Parr, C.L. Ant diversity and function in disturbed and changing habitats. In Ecology ants; Lach, L., Parr, C.L., Abbott, K., Eds.; Oxford University Press: New York, NY, USA, 2010; pp. 137–156. [Google Scholar]
- Wetterer, J.K.; Davis, L.R., Jr. Solenopsis invicta (Hymenoptera: Formicidae) in the lesser Antilles. Fla Entomol. 2010, 93, 128–129. [Google Scholar] [CrossRef]
- Ribeiro, M.C.; Metzger, J.P.; Martensen, A.C.; Ponzoni, F.J.; Hirota, M.M. The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biol. Conserv. 2009, 142, 1141–1153. [Google Scholar] [CrossRef]
- Bonamoni, J.; Totato, F.R.; Souza, R.; Gomes, R.; Penha, J.M.; Bueno, A.S.; Peres, C.A. Protecting forests at the expensive of native grasslands: Land-use police encourages open-habitat loss in Brazilian cerrado biome. Perspect. Ecol. Conserv. (PECON) 2019, 17, 26–31. [Google Scholar] [CrossRef]
- Ross, K.G.; Shoemaker, D.D. Estimation of the number of founders of an invasive pest insect population: The fire ant Solenopsis invicta in the USA. Proc. R. Soc. B Biol. Sci. 2008, 275, 2231–2240. [Google Scholar] [CrossRef]
- Souza, R.F.; Cocchi, F.K.; Martins, C.; Morini, M.S.C.; Bueno, O.C. Characterization of allele diversity in a microsatellite locus: A registry for Solenopsis invicta. Adv. Entomol. 2016, 4, 32–36. [Google Scholar] [CrossRef]
- Corin, S.E.; Lester, P.J.; Abbot, K.L.; Ritchie, P.A. Inferring historical introduction pathways with mitochondrial DNA: The case of introduced Argentine ants (Linepithema humile) into New Zealand. Divers. Distrib. 2007, 13, 510–518. [Google Scholar] [CrossRef]
- Vogel, V.; Pedersen, J.S.; Giraud, T.; Krieger, M.J.B.; Keller, L. The worldwide expansion of the Argentine ant. Divers. Distrib. 2010, 16, 170–186. [Google Scholar] [CrossRef]
- Sanllorente, O.; Ruano, F.; Tinaut, A. Large-scale population genetics of the mountain ant Proformica longiseta (Hymenoptera: Formicidae). Popul. Ecol. 2015, 57, 637–648. [Google Scholar] [CrossRef]
- Janda, M.; Matos-Maraví, P.; Borovanska, M.; Zima, J.; Youngerman, E.; Pierce, N.E. Phylogeny and population genetic structure of the ant genus Acropyga (Hymenoptera: Formicidae) in Papua New Guinea. Invertebr. Systemat. 2016, 30, 28–40. [Google Scholar] [CrossRef]
- Ramalho, M.O.; Martins, C.; Silva, L.M.R.; Martins, V.G.; Bueno, O.C. Molecular profile of the brazilian weaver ant Camponotus textor Forel (Hymenoptera, Formicidae). Neotrop. Entomol. 2016, 45, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.-C.; Hua, K.-H.; WU, W.-J. Intraspecific and interspecific aggressive interactions between two species of fire ants, Solenopsis geminata and S. invicta (Hymenoptera: Formicidae), in Taiwan. J. Asia Pac. Entomol. 2015, 18, 93–98. [Google Scholar] [CrossRef]
- Wickings, K.G.; Ruberson, J. Impact of the red imported fire ant (Hymenoptera: Formicidae) on epigeic arthropods of cotton agroecosystems. Ann. Entomol. Soc. Am. 2011, 104, 171–179. [Google Scholar] [CrossRef]
- Tsutsui, N.D.; Suarez, A.V. The colony structure and population biology of invasive ants. Conserv. Biol. 2003, 17, 48–58. [Google Scholar] [CrossRef]
- Ramalho, M.O.; Menino, L.; Souza, R.F.; Kayano, D.Y.; Alves, J.M.C.; Harakava, R.; Nagatani, V.H.; Silva, O.G.M.; Bueno, O.C.; Morini, M.S.C. Fire ants: What do rural and urban areas show us about occurrence, diversity, and ancestral state reconstruction? Genet. Mol. Biol. 2022, 45, e20210120. [Google Scholar] [CrossRef]
- Souza, R.F. Aspectos Bioecológicos e Genéticos de Solenopsis spp. no Mosaico de Paisagens do Alto Tietê Cabeceiras (SP). Ph.D. Thesis, Universidade de Mogi das Cruzes, Mogi das Cruzes, Brazil, 2019; 141p. [Google Scholar]
- Ignazzi, C.A. The Brazilian urban system: The trajectories of Brazilian cities between general dynamics and specific peculiarities. Eur. J. Geogr. 2015, 754, 27349. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Haplotype (Present Study) | Associated Name at NCBI | NCBI Accession Entry | Type Locality at NCBI |
|---|---|---|---|
| H1 H2 | MM34 | EU352608.1 JN808817.1 | Mississipi, EUA Corrientes, Argentina |
| H3 | H41 | AY950736 | Céu Azul, PR, Brazil |
| H4 | H47/H40 * | AY950742.1 AY950735.1 | Campo Grande, MS, Brazil Céu Azul, PR, Brazil |
| Scope of Variation | Sum of Squares | Variance Components | Percentage Variation | p-Value |
|---|---|---|---|---|
| Between supergroups A and B | 328.503 | 22.72043 | 98.61733 | 0.000 |
| Within supergroups A and B | 10.381 | 0.31855 | 1.38267 | |
| Total | 339.333 | 23.03898 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramalho, M.O.; Nagatani, V.H.; Alves, J.M.C.; Silva, O.G.M.; Fox, E.G.P.; Souza, R.F.; Kayano, D.Y.; Harakava, R.; Hilsdorf, A.W.S.; Morini, M.S.C. The Facet of Human Impact: Solenopsis invicta Buren, 1972 Spreading around the Atlantic Forest. Diversity 2023, 15, 194. https://doi.org/10.3390/d15020194
Ramalho MO, Nagatani VH, Alves JMC, Silva OGM, Fox EGP, Souza RF, Kayano DY, Harakava R, Hilsdorf AWS, Morini MSC. The Facet of Human Impact: Solenopsis invicta Buren, 1972 Spreading around the Atlantic Forest. Diversity. 2023; 15(2):194. https://doi.org/10.3390/d15020194
Chicago/Turabian StyleRamalho, Manuela O., Victor H. Nagatani, Juliana M. C. Alves, Otavio G. M. Silva, Eduardo G. P. Fox, Rodrigo F. Souza, Debora Y. Kayano, Ricardo Harakava, Alexandre W. S. Hilsdorf, and Maria S. C. Morini. 2023. "The Facet of Human Impact: Solenopsis invicta Buren, 1972 Spreading around the Atlantic Forest" Diversity 15, no. 2: 194. https://doi.org/10.3390/d15020194