
The Effect of Long-Term Agricultural Use on the Bacterial Microbiota of Chernozems of the Forest-Steppe Zone

Abstract
:1. Introduction
2. Materials and Methods
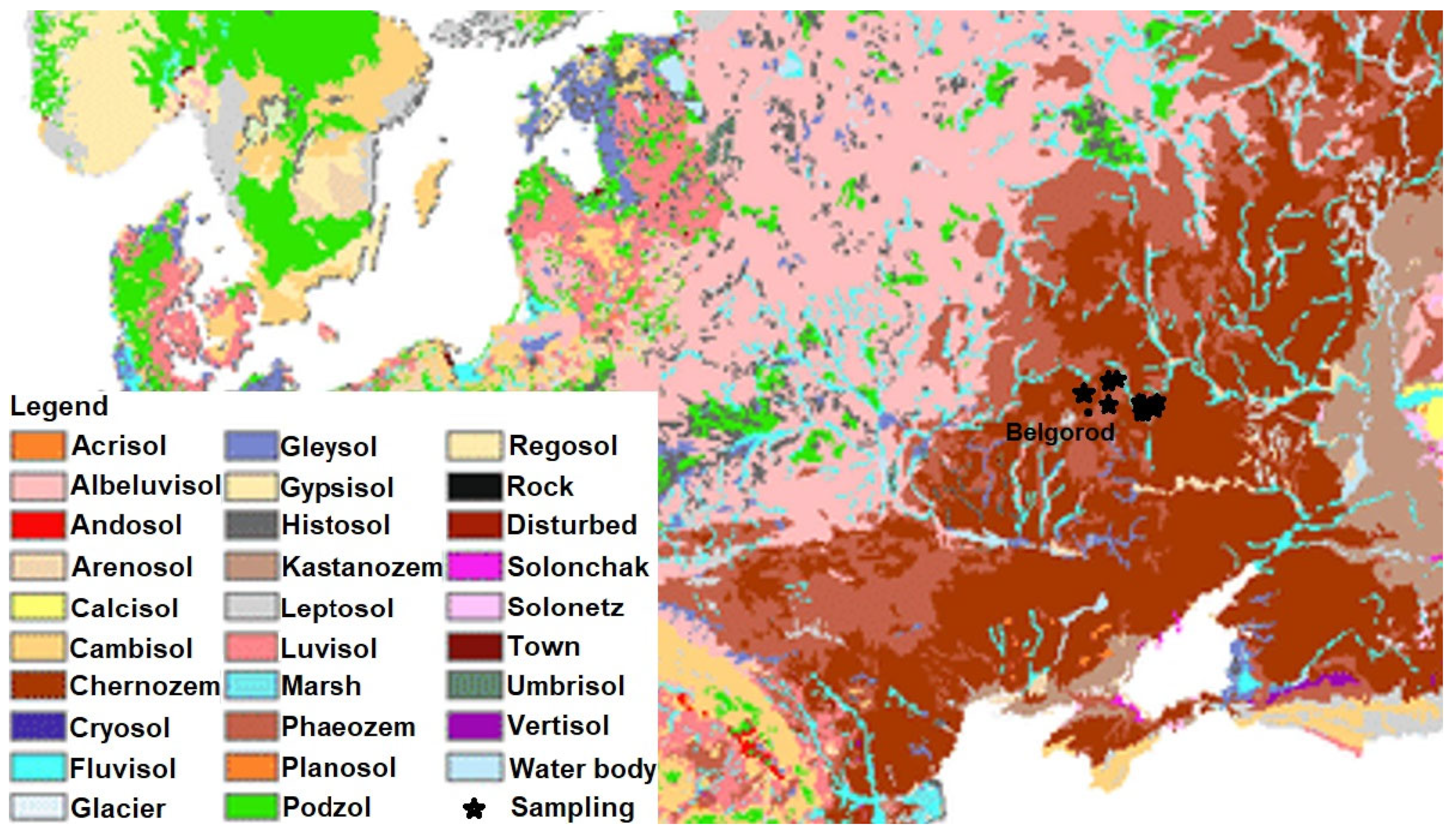
2.1. Sampling Sites and Sample Collection
2.2. Microbiological Profiling
2.3. Statistical Analysis of the Results
3. Results
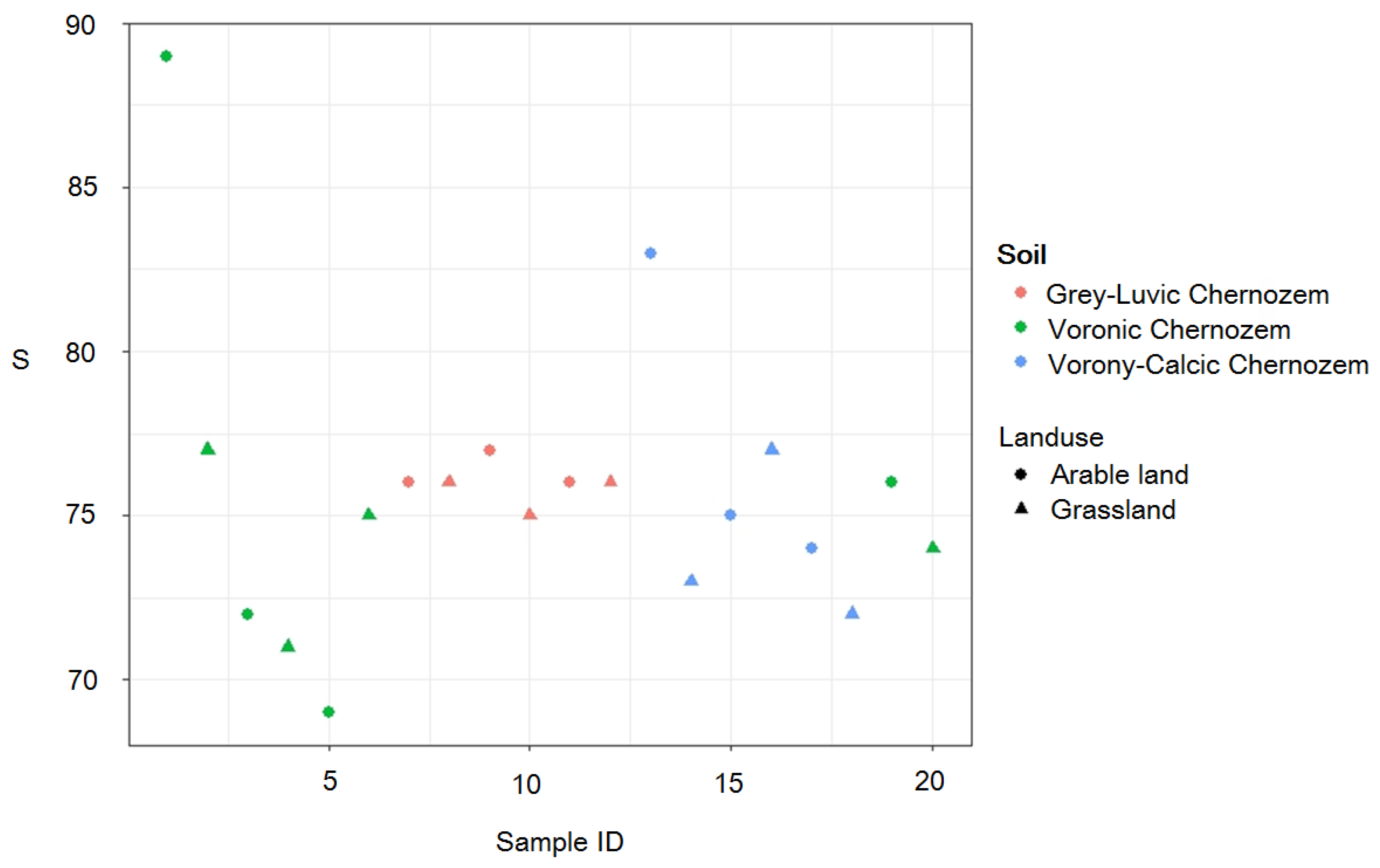
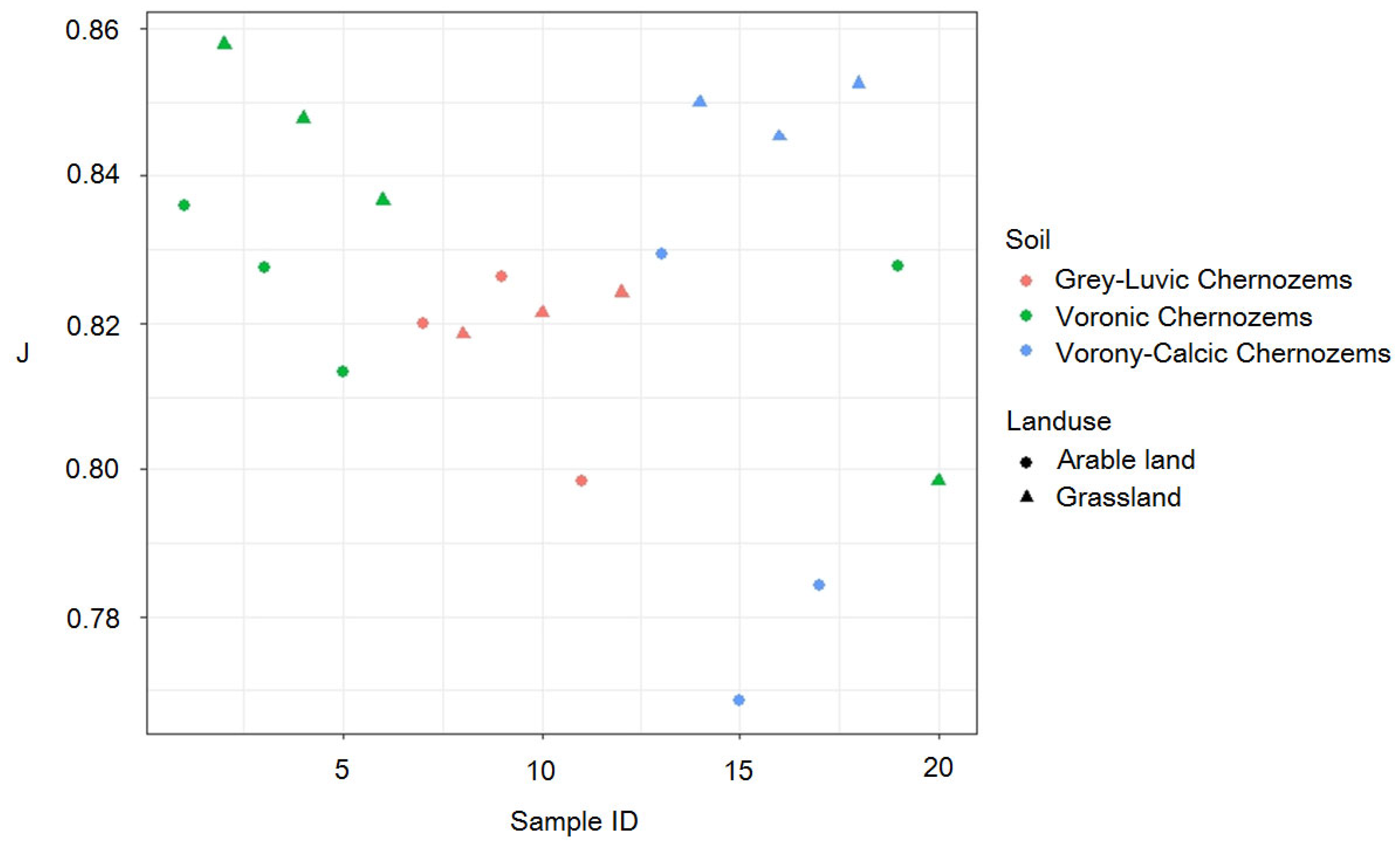
3.1. Comparison of Biodiversity Indicators
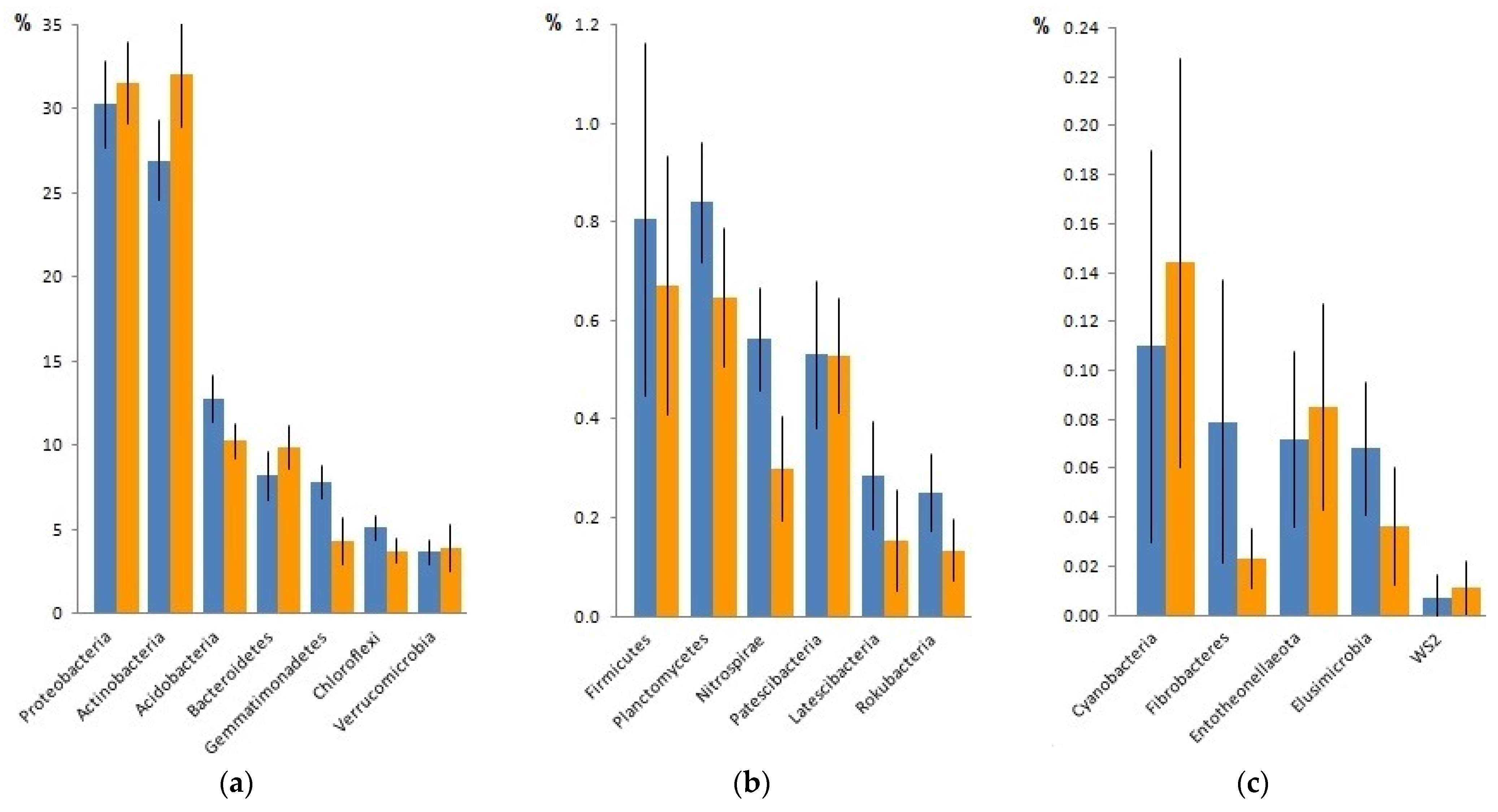
3.2. Comparison of Taxonomic Structure of the Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, X.; Burras, C.L.; Kravchenko, Y.S.; Duran, A.; Huffman, T.; Morras, H.; Studdert, G.; Zhang, X.; Cruse, R.M.; Yuan, X. Overview of Mollisols in the world: Distribution, land use and management. Can. J. Soil Sci. 2012, 92, 383–402. [Google Scholar] [CrossRef]
- Kögel-Knabner, I.; Amelung, W. Dynamics, Chemistry, and Preservation of Organic Matter in Soils. In Treatise on Geochemistry, 2nd ed.; Turekian, K., Holland, H., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2014; Volume 12, pp. 157–215. [Google Scholar]
- Vysloužilová, B.; Ertlen, D.; Schwartz, D.; Šefrna, L. Chernozem. From concept to classification: A review. Acta Univ. Carol. Geogr. 2016, 51, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Van, D.H. Cation binding of humic acids. Geoderma 1971, 5, 53–67. [Google Scholar]
- Szalay, A. Cation exchange properties of humic acids and their importance in the geochemical enrichment of UO2++ and other cations. Geochim. Cosmochim. Acta. 1964, 28, 1605–1614. [Google Scholar] [CrossRef]
- Hawkes, C.V.; DeAngelis, K.M. Chapter 1—Root Interactions with Soil Microbial Communities and Processes. In The Rhizosphere; Cardon, Z.G., Whitbeck, J.L., Eds.; Academic Press: Cambridge, MA, USA, 2007; pp. 1–29. [Google Scholar]
- Wagg, C.; Hautier, Y.; Pellkofer, S.; Banerjee, S.; Schmid, B.; van der Heijden, M.G.A. Diversity and asynchrony in soil microbial communities stabilizes ecosystem functioning. eLife 2021, 10, e62813. [Google Scholar] [CrossRef] [PubMed]
- Xue, P.P.; Carrillo, Y.; Pino, V.; Minasny, B.; McBratney, A.B. Soil Properties Drive Microbial Community Structure in a Large Scale Transect in South Eastern Australia. Sci. Rep. 2018, 8, 11725. [Google Scholar] [CrossRef] [Green Version]
- Lisetskii, F.N.; Zemlyakova, A.V.; Kirichenko, A.D. Variability of Microbiota under Diverse Conditions of Soil Moistening. Biol. Bull. Russ. Acad. Sci. 2018, 45, 337–344. [Google Scholar] [CrossRef]
- Domnariu, H.; Postolache, C.; Avramescu, S.; Lăcătușu, A.R.; Partal, E. Long term effects of tillage and fertilization upon microbiota of a Romanian Chernozem under maize monoculture. Geoderma Reg. 2022, 28, e00463. [Google Scholar] [CrossRef]
- Sun, R.; Li, W.; Dong, W.; Tian, Y.; Hu, C.; Liu, B. Tillage Changes Vertical Distribution of Soil Bacterial and Fungal Communities. Front. Microbiol. 2018, 9, 699. [Google Scholar] [CrossRef] [PubMed]
- Janušauskaite, D.; Kadžienė, G.; Auškalnienė, O. The Effect of Tillage System on Soil Microbiota in Relation to Soil Structure. Pol. J. Environ. Stud. 2013, 22, 1387–1391. [Google Scholar]
- Mathew, R.P.; Feng, Y.; Githinji, L.; Ankumah, R.; Balkcom, K.S. Impact of No-Tillage and Conventional Tillage Systems on Soil Microbial Communities. Appl. Environ. Soil Sci. 2012, 2012, 548620. [Google Scholar] [CrossRef] [Green Version]
- Frøslev, T.G.; Nielsen, I.B.; Santos, S.S.; Barnes, C.J.; Bruun, H.H.; Ejrnæs, R. The biodiversity effect of reduced tillage on soil microbiota. Ambio 2022, 51, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Kraut-Cohen, J.; Zolti, A.; Shaltiel-Harpaz, L.; Argaman, E.; Rabinovich, R.; Green, S.J.; Minz, D. Effects of tillage practices on soil microbiome and agricultural parameters. Sci. Total Environ. 2020, 705, 135791. [Google Scholar] [CrossRef]
- Svistova, I.D.; Stakhurlova, L.D. Microbial succession in the typical chernozems of “Streletskaya steppe” reserve under the influence of long agrogenic impact. Ross. Selskokhoziaistvennaia Nauka 2019, 1, 40–42. [Google Scholar] [CrossRef]
- Semenov, M.; Chernov, T.; Tkhakakhova, A.; Zhelezova, A.; Ivanova, E.; Kolganova, T.; Kutovaya, O. Distribution of prokaryotic communities throughout the Chernozem profiles under different land uses for over a century. Appl. Soil Ecol. 2018, 127, 8–18. [Google Scholar] [CrossRef]
- Gorbacheva, M.A.; Melnikova, N.V.; Chechetkin, V.R.; Kravatsky, Y.V.; Tchurikov, N.A. DNA sequencing and metagenomics of cultivated and uncultivated chernozems in Russia. Geoderma Reg. 2018, 14, e00180. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Powell, J.R.; Hamonts, K.; Reith, F.; Mele, P.; Brown, M.V.; Dennis, P.G.; Ferrari, B.C.; Fitzgerald, A.; Young, A.; et al. Circular linkages between soil biodiversity, fertility and plant productivity are limited to topsoil at the continental scale. New Phytol. 2017, 215, 1186–1196. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Eldridge, D.; Ochoa, V.; Gozalo, B.; Singh, B.; Maestre, F. Soil microbial communities drive the resistance of ecosystem multifunctionality to global change in drylands across the globe. Ecol. Lett. 2017, 20, 1295–1305. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Zhang, C.; Xu, X.; Wang, C.; Liu, G.; Liang, C.; Zuo, X.; Wang, C.; Lv, Y.; Wang, R. Different facets of bacterial and fungal communities drive soil multifunctionality in grasslands spanning a 3500 km transect. Funct. Ecol. 2022, 36, 3120–3133. [Google Scholar] [CrossRef]
- Bucur, D.; Jităreanu, G.; Ailincăi, C.; Tsadilas, C.; Ailinci, D.; Mercus, A. Influence of soil erosion on water, soil, humus and nutrient losses in different crop systems in the Moldavian Plateau, Romania. J. Food Agric. Environ. 2007, 5, 261–264. [Google Scholar]
- Blanco-Canqui, H.; Lal, R. Biological Measures of Erosion Control. In Principles of Soil Conservation and Management; Blanco-Canqui, H., Lal, R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 137–165. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordonet, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucl. Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing, R Core Team: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 25 April 2021).
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; Balakrishnan, N., Colton, T., Eds.; Wiley: New York, NY, USA, 2017; pp. 1–15. [Google Scholar]
- Roger, B.J.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar]
- Whittaker, R.H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 1960, 30, 279–338. [Google Scholar] [CrossRef]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley: NY, USA, 1969; p. 294. [Google Scholar]
- Harrison, S. How natural habitat patchiness affects the distribution of diversity in Californian Serpentine Chaparral. Ecology 1997, 78, 1898–1906. [Google Scholar] [CrossRef]
- Tuomisto, H. A diversity of beta diversities: Straightening up a concept gone awry. Part 1. Defining beta diversity as a function of alpha and gamma diversity. Ecography 2010, 33, 2–22. [Google Scholar] [CrossRef]
- Oksanen, J. Vegan: Ecological diversity. R Proj. 2013, 368, 1–11. [Google Scholar]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Wilcoxon, F. Individual comparisons by ranking methods. Biom. Bull. 1945, 1, 80–83. [Google Scholar] [CrossRef]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.D.; McVeigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef]
- Zhang, H.; Sekiguchi, Y.; Hanada, S.; Hugenholtz, P.; Kim, H.; Kamagata, Y.; Nakamura, K. Gemmatimonas aurantiaca gen. nov., sp. nov., a gram-negative, aerobic, polyphosphate-accumulating micro-organism, the first cultured representative of the new bacterial phylum Gemmatimonadetes phyl. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 1155–1163. [Google Scholar] [CrossRef]
- Zeng, Y.; Selyanin, V.; Lukes, M.; Dean, J.; Kaftan, D.; Feng, F.; Koblizek, M. Characterization of the microaerophilic, bacteriochlorophyll a-containing bacterium Gemmatimonas phototrophica sp. nov., and emended descriptions of the genus Gemmatimonas and Gemmatimonas aurantiaca. Int. J. Syst. Evol. Microbiol. 2015, 65, 2410–2419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Nupur; Wu, N.; Madsen, A.M.; Chen, X.; Gardiner, A.T.; Koblížek, M. Gemmatimonas groenlandica sp. nov. is an aerobic anoxygenic phototroph in the phylum Gemmatimonadetes. Front. Microbiol. 2021, 11, 606612. [Google Scholar] [CrossRef]
- Wang, X.; He, T.; Gen, S.; Zhang, X.-Q.; Wang, X.; Jiang, D.; Li, C.; Li, C.; Wang, J.; Zhang, W.; et al. Soil properties and agricultural practices shape microbial communities in flooded and rainfed croplands. Appl. Soil Ecol. 2020, 147, 103449. [Google Scholar] [CrossRef]
- Kulichevskaya, I.S.; Suzina, N.E.; Liesack, W.; Dedysh, S.N. Bryobacter aggregatus gen. nov., sp. nov., a peat-inhabiting, aerobic chemo-organotroph from subdivision 3 of the Acidobacteria. Int. J. Syst. Evol. Microbiol. 2010, 60, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Daims, H. The Family Nitrospiraceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 733–749. [Google Scholar]
- Vijayan, A.; Jayadradhan, R.V.J.; Pillai, D.; Geetha, P.P.; Valsamma, J.; Sarojini, B.S.I. Nitrospira as versatile nitrifiers: Taxonomy, ecophysiology, genome characteristics, growth, and metabolic diversity. J. Basic Microbiol. 2021, 61, 88–109. [Google Scholar] [CrossRef]
- Prosser, J.I.; Head, I.M.; Stein, L.Y. The Family Nitrosomonadaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 901–918. [Google Scholar]
- Yoon, J.H.; Kang, S.J.; Lee, S.Y.; Lee, J.S.; Park, S. Ohtaekwangia koreensis gen. nov., sp. nov. and Ohtaekwangia kribbensis sp. nov., isolated from marine sand, deep-branching members of the phylum Bacteroidetes. Int. J. Syst. Evol. Microbiol. 2011, 61, 1066–1072. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-J.; Alkawally, M.; Brady, A.L.; Rijpstra, W.I.C.; Damste, S.J.S.; Dunfield, P.F. Chryseolinea serpens gen. nov., sp. nov., a member of the phylum Bacteroidetes isolated from soil. Int. J. Syst. Evol. Microbiol. 2013, 63, 654–660. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.A.; Kim, Y.; Sang, M.-K.; Song, J.; Kwon, S.W.; Weon, H.Y. Chryseolinea soli sp. nov., isolated from soil. J. Microbiol. 2019, 57, 122–126. [Google Scholar] [CrossRef]
- Tóth, E.M.; Borsodi, A.K. The Family Nocardioidaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 651–694. [Google Scholar]
- Matsumoto, A.; Kasai, H.; Matsuo, Y.; Shizuri, Y.; Ichikawa, N.; Fujita, N.; Ōmura, S.; Takahashi, Y. Ilumatobacter nonamiense sp. nov. and Ilumatobacter coccineum sp. nov., isolated from seashore sand. Int. J. Syst. Evol. Microbiol. 2013, 63, 3404–3408. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, A.; Kasai, H.; Matsuo, Y.; Omura, S.; Shizuri, Y.; Takahashi, Y. Ilumatobacter fluminis gen. nov., sp. nov., a novel actinobacterium isolated from the sediment of an estuary. J. Gen. Appl. Microbiol. 2009, 55, 201–205. [Google Scholar]
- Zhang, J.; Song, F.; Xin, Y.H.; Zhang, J.; Fang, C. Microvirga guangxiensis sp. nov., a novel alphaproteobacterium from soil, and emended description of the genus Microvirga. Int. J. Syst. Evol. Microbiol. 2009, 59, 1997–2001. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Ping, W.; Zhang, S.; Chen, Y.; Zhang, Y.; Zhang, J. Description and genome analysis of Microvirga antarctica sp. nov., a novel pink-pigmented psychrotolerant bacterium isolated from Antarctic soil. Antonie Van Leeuwenhoek 2021, 114, 2219–2228. [Google Scholar] [CrossRef]
- Li, J.; Gao, R.; Chen, Y.; Xue, D.; Han, J.; Wang, J.; Dai, Q.; Lin, M.; Ke, X.; Zhang, W. Isolation and Identification of Microvirga thermotolerans HR1, a Novel Thermo-Tolerant Bacterium, and Comparative Genomics among Microvirga Species. Microorganisms 2020, 8, 101. [Google Scholar] [CrossRef] [Green Version]
- Mohr, K.I.; Garcia, R.O.; Gerth, K.; Irschik, H.; Muller, R. Sandaracinus amylolyticus gen. nov., sp. nov., a starch-degrading soil myxobacterium, and description of Sandaracinaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2012, 62, 1191–1198. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Voronic | Vor.-Calcic | Grey-Luvic | Total | |
|---|---|---|---|---|
| Arable | 4 | 3 | 3 | 10 |
| Nonarable | 4 | 3 | 3 | 10 |
| Total | 8 | 6 | 6 | 20 |
| Factors | Diversity | |||
|---|---|---|---|---|
| Alfa | Beta | Gamma | ||
| Land use | Arable land | 76.7 | 0.186 | 91 |
| Grassland | 74.6 | 0.206 | 90 | |
| Soil subtype | Grey-Luvic chernozems | 76 | 0.184 | 90 |
| Vorony-Calcic Chernozems | 75.7 | 0.189 | 90 | |
| Voronic Chernozems | 75.4 | 0.207 | 91 | |
| Factors | Df | Sum Of Sqs | R2 | F | Pr(>F) |
|---|---|---|---|---|---|
| Land use | 1 | 0.14431 | 0.16008 | 4.0736 | 0.002 |
| Soil | 2 | 0.20822 | 0.23097 | 2.9388 | 0.001 |
| Land use:soil | 2 | 0.05302 | 0.05881 | 0.7482 | 0.720 |
| Residual | 14 | 0.49597 | 0.55015 | ||
| Total | 19 | 0.90152 | 1.00000 |
| Phyla (Class) | Familia | %A. | %N.a. | p |
|---|---|---|---|---|
| (Alphaproteobacteria) | Reyranellaceae | 0.44 | 0.71 | 0.016 |
| Beijerinckiaceae | 0.76 | 1.13 | 0.041 | |
| Paracaedibacteraceae | 0.02 | 0.00 | 0.045 | |
| (Gammaproteobacteria) | Nitrosomonadaceae | 1.33 | 0.91 | 0.019 |
| Solimonadaceae | 0.04 | 0.01 | 0.050 | |
| (Deltaproteobacteria) | Sandaracinaceae | 0.16 | 0.49 | 0.013 |
| Actinobacteria | Ilumatobacteraceae | 0.86 | 1.72 | 0.041 |
| Micrococcaceae | 1.21 | 0.50 | 0.007 | |
| Nocardioidaceae | 1.36 | 2.03 | 0.028 | |
| Microbacteriaceae | 0.42 | 0.93 | 0.028 | |
| Glycomycetaceae | 0.04 | 0.00 | 0.005 | |
| Mycobacteriaceae | 0.24 | 0.60 | 0.010 | |
| Solirubrobacteraceae | 1.18 | 1.97 | 0.049 | |
| Acidobacteria | Solibacteraceae | 1.79 | 1.19 | 0.019 |
| Pyrinomonadaceae | 1.05 | 0.53 | 0.004 | |
| Bacteroidetes | Microscillaceae | 0.88 | 2.12 | 0.019 |
| Gemmatimonadetes | Gemmatimonadaceae | 7.59 | 3.87 | 0.007 |
| Chloroflexi | Roseiflexaceae | 0.47 | 0.19 | 0.001 |
| Anaerolineaceae | 0.28 | 0.09 | 0.001 | |
| Nitrospirae | Nitrospiraceae | 0.56 | 0.31 | 0.005 |
| V.—V.c. | V.—G.l. | V.c.—G.l. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Phyla (Class) | Familia | %V. | %V.c. | p | %V. | %G.l. | p | %V.c. | %G.l. | p |
| (Alpha- Proteobacteria) | Sphingomonadaceae | 4.11 | 7.78 | 0.012 | 4.11 | 7.89 | 0.012 | 7.78 | 7.89 | 0.937 |
| Azospirillaceae | 0.12 | 0.52 | 0.035 | 0.12 | 0.20 | 0.065 | 0.52 | 0.20 | 0.065 | |
| Acetobacteraceae | 0.08 | 0.22 | 0.001 | 0.08 | 0.29 | 0.001 | 0.22 | 0.29 | 0.180 | |
| (Gamma- Proteobacteria) | Xanthomonadaceae | 0.75 | 1.93 | 0.024 | 0.75 | 1.45 | 0.030 | 1.93 | 1.45 | 0.485 |
| Rhodanobacteraceae | 0.21 | 0.52 | 0.004 | 0.21 | 0.46 | 0.004 | 0.52 | 0.46 | 0.589 | |
| Pseudomonadaceae | 0.30 | 0.43 | 0.343 | 0.30 | 0.57 | 0.024 | 0.43 | 0.57 | 0.589 | |
| Actinobacteria | Geodermatophilaceae | 0.41 | 1.04 | 0.024 | 0.41 | 0.93 | 0.030 | 1.04 | 0.93 | 0.818 |
| Propionibacteriaceae | 0.23 | 0.14 | 0.282 | 0.23 | 0.05 | 0.008 | 0.14 | 0.05 | 0.039 | |
| Kineosporiaceae | 0.09 | 0.28 | 0.240 | 0.09 | 0.39 | 0.004 | 0.28 | 0.39 | 0.240 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boyarshin, K.S.; Adamova, V.V.; Wentao, Z.; Obuhova, O.Y.; Kolkova, M.V.; Nesterenko, V.A.; Bespalova, O.S.; Kluyeva, V.V.; Degtyareva, K.A.; Kurkina, Y.N.; et al. The Effect of Long-Term Agricultural Use on the Bacterial Microbiota of Chernozems of the Forest-Steppe Zone. Diversity 2023, 15, 191. https://doi.org/10.3390/d15020191
Boyarshin KS, Adamova VV, Wentao Z, Obuhova OY, Kolkova MV, Nesterenko VA, Bespalova OS, Kluyeva VV, Degtyareva KA, Kurkina YN, et al. The Effect of Long-Term Agricultural Use on the Bacterial Microbiota of Chernozems of the Forest-Steppe Zone. Diversity. 2023; 15(2):191. https://doi.org/10.3390/d15020191
Chicago/Turabian StyleBoyarshin, Konstantin S., Valeria V. Adamova, Zheng Wentao, Olga Y. Obuhova, Marina V. Kolkova, Vitaliy A. Nesterenko, Olga S. Bespalova, Violetta V. Kluyeva, Kristina A. Degtyareva, Yulia N. Kurkina, and et al. 2023. "The Effect of Long-Term Agricultural Use on the Bacterial Microbiota of Chernozems of the Forest-Steppe Zone" Diversity 15, no. 2: 191. https://doi.org/10.3390/d15020191