

Compilation, Revision, and Annotation of DNA Barcodes of Marine Invertebrate Non-Indigenous Species (NIS) Occurring in European Coastal Regions
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Compilation of the List of Non-Indigenous Invertebrate Marine Species Occurring in Europe
2.2. Compilation of the Genetic Data and Curation
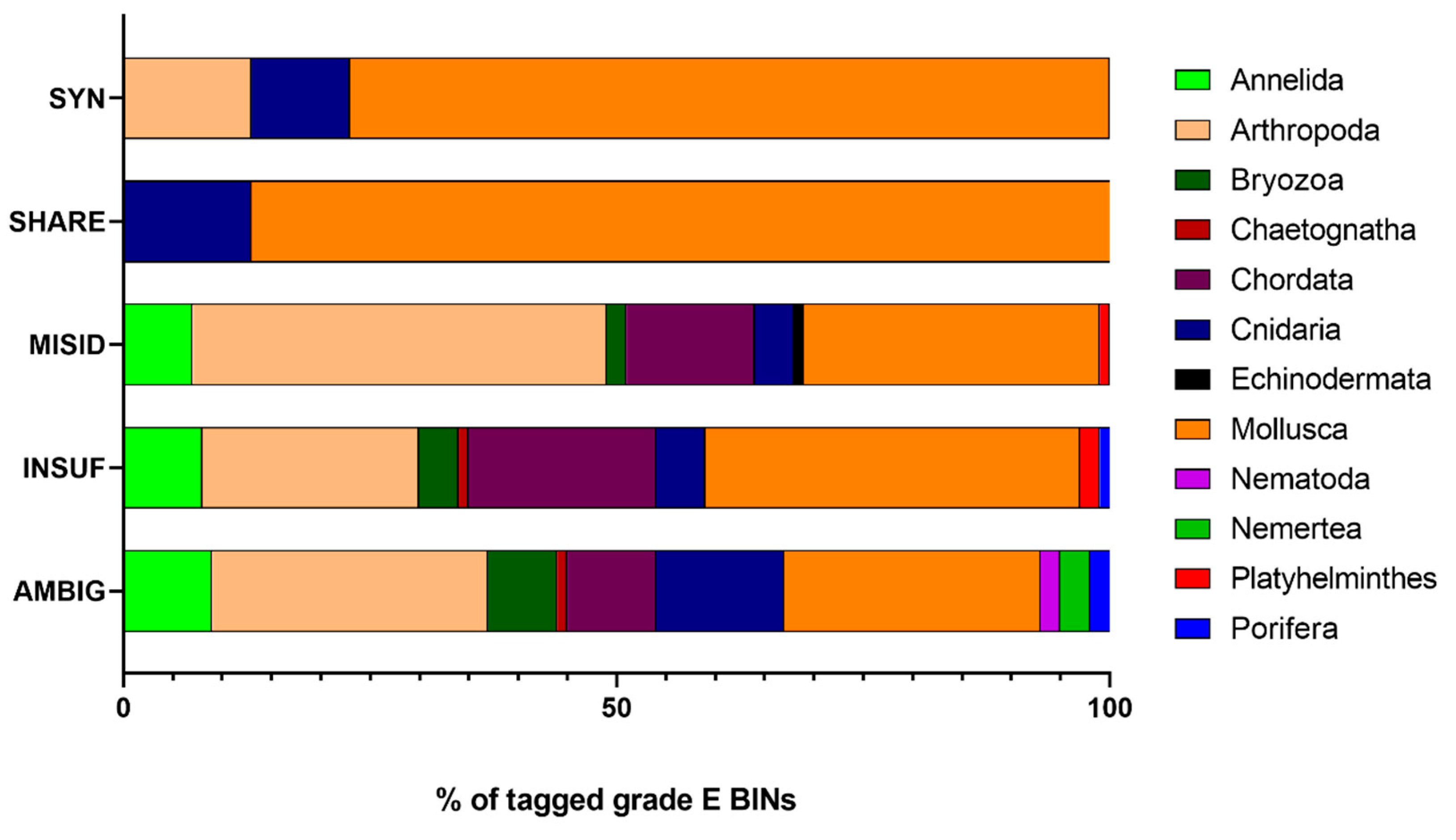
- BINs were tagged SYN if the identifications (IDs) were synonyms of the accepted species name (confirmed using the WoRMS database) and were considered RELIABLE.
- BINs were tagged AMBIG and UNCERTAIN if:
- The BIN comprises more than 3 different IDs (i.e., species, genus, incomplete classification);
- The BIN comprises 3 IDs, but one of the two IDs with the lowest number of records still contains ≥ 10 records.
- BINs were tagged MISID, and the discordance was solved if:
- When there are only two IDs, and the number of records of the putative “incorrect ID” is ≤ 5% of all records in the BIN (rounded up to the unit), the species with the highest number of records is considered the correct ID;
- The ID with the highest number of records is close to other BINs of the same species (when analyzing the NJ tree for the genus), then it is considered the correct ID;
- The ID with the lowest number of records is from a different genus than the one that is considered the “correct ID”, then it is considered the incorrect ID;
- When analyzing the NJ tree for the genus, the ID with the lowest number of records is in a different cluster, containing only records of that same ID, then it is considered the incorrect ID.
2.3. Testing the Impact of the Curated Reference Library on the Accuracy and Amount of NIS Detected through DNA Metabarcoding of Natural Communities
3. Results
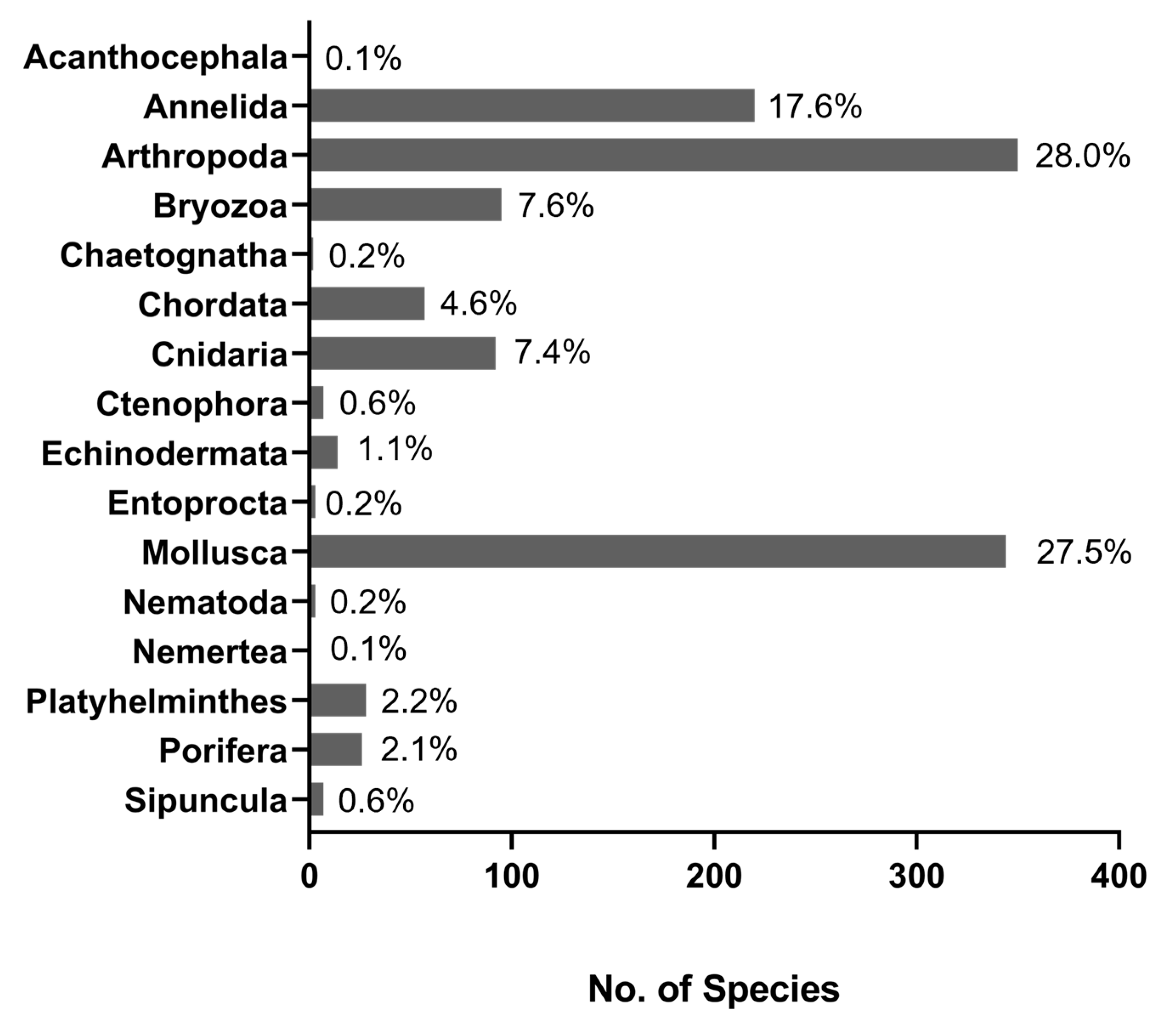
3.1. List of Non-Indigenous Invertebrate Marine Species Occurring in Europe
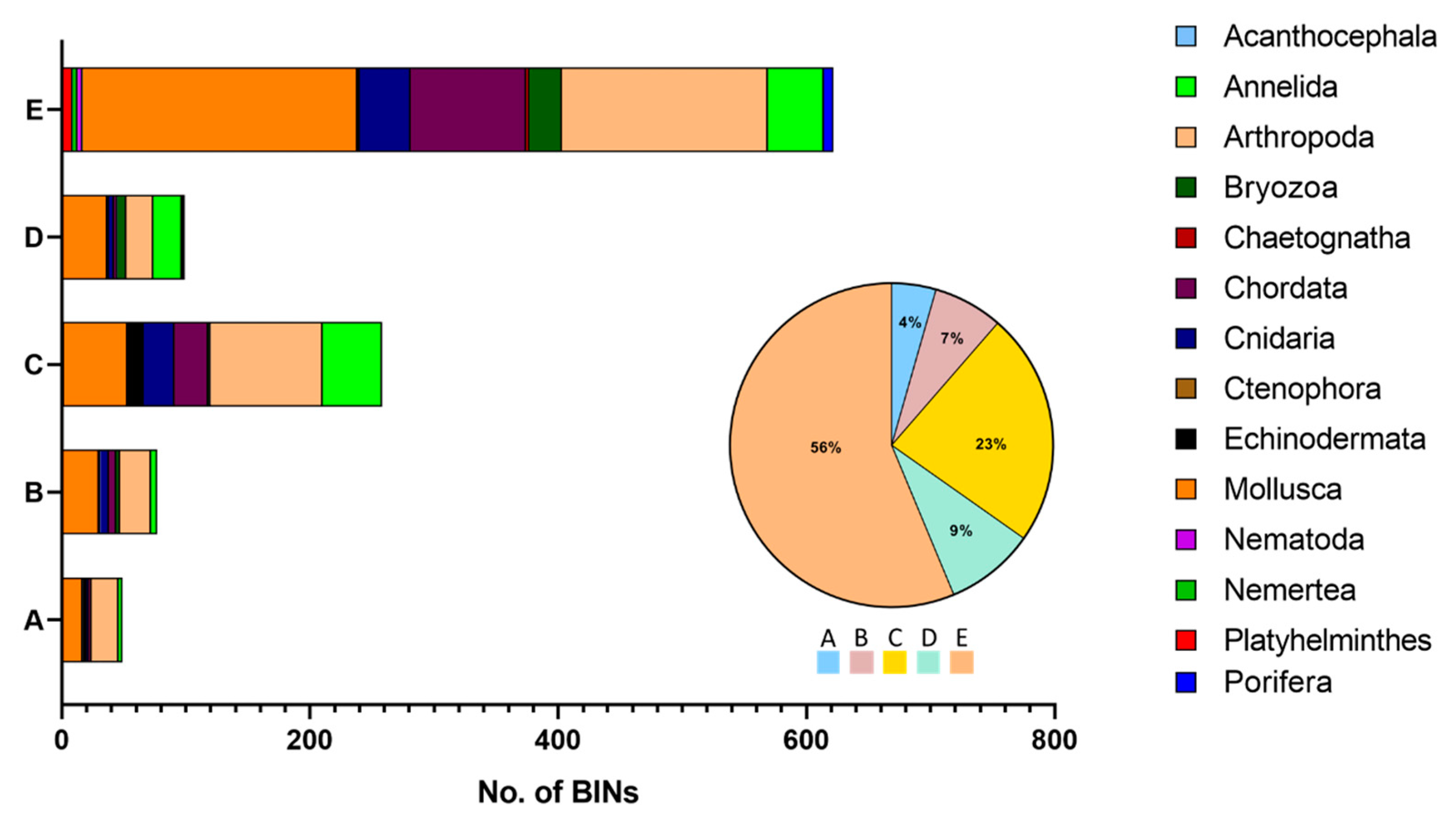
3.2. Curation of the Taxonomic Assignments of Barcode Records
3.3. Impact of the Curated Reference Library on the Accuracy and Amount of NIS Detected through DNA Metabarcoding of Natural Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Jackson, J.B.C.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A.; et al. Historical Overfishing and the Recent Collapse of Coastal Ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Turner, R.E.; Rabalais, N.N. Coastal eutrophication near the Mississipi river Delta. Nature 1994, 368, 619–621. [Google Scholar] [CrossRef]
- Rilov, G.; Crooks, J.A. Biological Invasions in Marine Ecosystems: Ecological, Management and Geographic Perspectives; Springer: Berlin/Heidelberg, Germany, 2009; p. 30. [Google Scholar] [CrossRef]
- Bax, N.; Williamson, A.; Aguero, M.; Gonzalez, E.; Geeves, W. Marine invasive alien species: A threat to global biodiversity. Mar. Policy 2003, 27, 313–323. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of invasive alien marine species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Diagne, C.; Leroy, B.; Vaissière, A.-C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.-M.; Bradshaw, C.J.A.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef] [PubMed]
- EU Regulation 1143/2014; European Union Regulation (EU) No 1143/2014 of the European Parliament and the Council of 22 October 2014 on the Prevention and Management of the Introduction and Spread of Invasive Alien Species. European Commission: Brussel, Belgium, 2014; Volume 317, pp. 35–55.
- Directive 2008/56/EC; European Comission Directive of the European Parliament and the Council Establishing a Framework for Community Action in the Field of Marine Environmental Policy (Marine Strategy Framework Directive). European Parliament and of the Council: Brussel, Belgium, 2008.
- Lehtiniemi, M.; Ojaveer, H.; David, M.; Galil, B.; Gollasch, S.; McKenzie, C.; Minchin, D.; Occhipinti-Ambrogi, A.; Olenin, S.; Pederson, J. Dose of truth—Monitoring marine non-indigenous species to serve legislative requirements. Mar. Policy 2015, 54, 26–35. [Google Scholar] [CrossRef]
- Afonso, I.; Berecibar, E.; Castro, N.; Costa, J.; Frias, P.; Henriques, F.; Moreira, P.; Oliveira, P.; Silva, G.; Chainho, P. Assessment of the colonization and dispersal success of non-indigenous species introduced in recreational marinas along the estuarine gradient. Ecol. Indic. 2020, 113, 106147. [Google Scholar] [CrossRef]
- Chainho, P.; Fernandes, A.; Amorim, A.; Ávila, S.P.; Canning-Clode, J.; Castro, J.J.; Costa, A.C.; Costa, J.L.; Cruz, T.; Gollasch, S.; et al. Non-indigenous species in Portuguese coastal areas, coastal lagoons, estuaries and islands. Estuar. Coast. Shelf Sci. 2015, 167, 199–211. [Google Scholar] [CrossRef]
- Mancinelli, G.; Chainho, P.; Cilenti, L.; Falco, S.; Kapiris, K.; Katselis, G.; Ribeiro, F. The Atlantic blue crab Callinectes sapidus in southern European coastal waters: Distribution, impact and prospective invasion management strategies. Mar. Pollut. Bull. 2017, 119, 5–11. [Google Scholar] [CrossRef]
- Hopkins, G.W.; Freckleton, R.P. Declines in the numbers of amateur and professional taxonomists: Implications for conservation. Anim. Conserv. 2002, 5, 245–249. [Google Scholar] [CrossRef]
- Kim, K.C.; Byrne, L.B. Biodiversity loss and the taxonomic bottleneck: Emerging biodiversity science. Ecol. Res. 2006, 21, 794–810. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; Dewaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cristescu, M.E. From barcoding single individuals to metabarcoding biological communities: Towards an integrative approach to the study of global biodiversity. Trends Ecol. Evol. 2014, 29, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Hajibabaei, M. The golden age of DNA metasystematics. Trends Genet. 2012, 28, 535–537. [Google Scholar] [CrossRef] [PubMed]
- Holman, L.E.; de Bruyn, M.; Creer, S.; Carvalho, G.; Robidart, J.; Rius, M. Detection of introduced and resident marine species using environmental DNA metabarcoding of sediment and water. Sci. Rep. 2019, 9, 11559. [Google Scholar] [CrossRef] [Green Version]
- Pochon, X.; Zaiko, A.; Hopkins, G.A.; Banks, J.C.; Wood, S.A. Early detection of eukaryotic communities from marine biofilm using high-throughput sequencing: An assessment of different sampling devices. Biofouling 2015, 31, 241–251. [Google Scholar] [CrossRef]
- Rey, A.; Basurko, O.C.; Rodriguez-Ezpeleta, N. Considerations for metabarcoding-based port biological baseline surveys aimed at marine nonindigenous species monitoring and risk assessments. Ecol. Evol. 2020, 10, 2452–2465. [Google Scholar] [CrossRef] [Green Version]
- Taberlet, P.; Coissac, E.; Hajibabaei, M.; Rieseberg, L.H. Environmental DNA. Mol. Ecol. 2012, 21, 1789–1793. [Google Scholar] [CrossRef]
- Duarte, S.; Vieira, P.E.; Lavrador, A.S.; Costa, F.O. Status and prospects of marine NIS detection and monitoring through (e)DNA metabarcoding. Sci. Total Environ. 2021, 751, 141729. [Google Scholar] [CrossRef]
- Knowlton, N. Sibling species in the sea. Annu. Rev. Ecol. Syst. 1993, 24, 189–216. [Google Scholar] [CrossRef]
- Radulovici, A.E.; Vieira, P.E.; Duarte, S.; Teixeira, M.A.L.; Borges, L.M.S.; Deagle, B.E.; Majaneva, S.; Redmond, N.; Schultz, J.A.; Costa, F.O. Revision and annotation of DNA barcode records for marine invertebrates: Report of the 8th iBOL conference hackathon. Metabarcoding Metagenomics 2021, 5, e67862. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, P.D.N.; Ratnasingham, S.; De Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Biol. Sci. 2003, 270, S96–S99. [Google Scholar] [CrossRef] [Green Version]
- Paz, G.; Rinkevich, B. Gap analysis of DNA barcoding in ERMS reference libraries for ascidians and cnidarians. Environ. Sci. Eur. 2021, 33, 4. [Google Scholar] [CrossRef]
- Leite, B.R.; Vieira, P.E.; Teixeira, M.A.L.; Lobo-Arteaga, J.; Hollatz, C.; Borges, L.M.S.; Duarte, S.; Troncoso, J.S.; Costa, F.O. Gap-analysis and annotated reference library for supporting macroinvertebrate metabarcoding in Atlantic Iberia. Reg. Stud. Mar. Sci. 2020, 36, 101307. [Google Scholar] [CrossRef]
- Vieira, P.E.; Lavrador, A.S.; Parente, M.I.; Parretti, P.; Costa, A.C.; Costa, F.O.; Duarte, S. Gaps in DNA sequence libraries for Macaronesian marine macroinvertebrates imply decades till completion and robust monitoring. Divers. Distrib. 2021, 27, 2003–2015. [Google Scholar] [CrossRef]
- Weigand, H.; Beermann, A.J.; Čiampor, F.; Costa, F.O.; Csabai, Z.; Duarte, S.; Geigerg, M.F.; Grabowski, M.; Rimet, F.; Rulik, B.; et al. DNA barcode reference libraries for the monitoring of aquatic biota in Europe: Gap-analysis and recommendations for future work. Sci. Total Environ. 2019, 678, 499–524. [Google Scholar] [CrossRef]
- Duarte, S.; Vieira, P.E.; Costa, F.O. Assessment of species gaps in DNA barcode libraries of nonindigenous species (NIS) occurring in European coastal regions. Metabarcoding Metagenomics 2020, 4, 35–46. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-Based Registry for All Animal Species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimet, F.; Aylagas, E.; Borja, A.; Bouchez, A.; Canino, A.; Chauvin, C.; Chonova, T.; Jr, F.C.; Costa, F.O.; Ferrari, B.J.D.; et al. Metadata standards and practical guidelines for specimen and DNA curation when building barcode reference libraries for aquatic life. Metabarcoding Metagenomics 2021, 5, e58056. [Google Scholar] [CrossRef]
- Leese, F.; Altermatt, F.; Bouchez, A.; Ekrem, T.; Hering, D.; Meissner, K.; Mergen, P.; Pawlowski, J.; Piggott, J.; Rimet, F.; et al. DNAqua-Net: Developing new genetic tools for bioassessment and monitoring of aquatic ecosystems in Europe. Res. Ideas Outcomes 2016, 2, e11321. [Google Scholar] [CrossRef] [Green Version]
- Zaiko, A.; Pochon, X.; Garcia-Vazquez, E.; Olenin, S.; Wood, S.A. Advantages and Limitations of Environmental DNA/RNA Tools for Marine Biosecurity: Management and Surveillance of Non-Indigenous Species. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Machida, R.J.; Leray, M.; Ho, S.-L.; Knowlton, N. Metazoan mitochondrial gene sequence reference datasets for taxonomic assignment of environmental samples. Sci. Data 2017, 4, 170027. [Google Scholar] [CrossRef] [Green Version]
- Neto, L.; Pinto, N.; Proença, A.; Amorim, A.; Conde-Sousa, E. 4SpecID: Reference DNA Libraries Auditing and Annotation System for Forensic Applications. Genes 2021, 12, 61. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef]
- Bucklin, A.; Peijnenburg, K.T.C.A.; Kosobokova, K.N.; O’Brien, T.D.; Blanco-Bercial, L.; Cornils, A.; Falkenhaug, T.; Hopcroft, R.R.; Hosia, A.; Laakmann, S.; et al. Toward a global reference database of COI barcodes for marine zooplankton. Mar. Biol. 2021, 168, 78. [Google Scholar] [CrossRef]
- Nugent, C.M.; Elliott, T.A.; Ratnasingham, S.; Adamowicz, S.J. Coil: An R package for cytochrome c oxidase I (COI) DNA barcode data cleaning, translation, and error evaluation. Genome 2020, 63, 291–305. [Google Scholar] [CrossRef]
- Oliveira, L.M.; Knebelsberger, T.; Landi, M.; Soares, P.; Raupach, M.J.; Costa, F.O. Assembling and auditing a comprehensive DNA barcode reference library for European marine fishes. J. Fish Biol. 2016, 89, 2741–2754. [Google Scholar] [CrossRef] [Green Version]
- Fontes, J.T.; Vieira, P.E.; Ekrem, T.; Soares, P.; Costa, F.O. BAGS: An automated Barcode, Audit & Grade System for DNA barcode reference libraries. Mol. Ecol. Resour. 2021, 21, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Landi, M.; Dimech, M.; Arculeo, M.; Biondo, G.; Martins, R.; Carneiro, M.; Carvalho, G.R.; Brutto, S.L.; Costa, F. DNA Barcoding for Species Assignment: The Case of Mediterranean Marine Fishes. PLoS ONE 2014, 9, e106135. [Google Scholar] [CrossRef] [PubMed]
- Grant, D.M.; Brodnicke, O.B.; Evankow, A.M.; Ferreira, A.O.; Fontes, J.T.; Hansen, A.K.; Jensen, M.R.; Kalaycı, T.E.; Leeper, A.; Patil, S.K.; et al. The Future of DNA Barcoding: Reflections from Early Career Researchers. Diversity 2021, 13, 313. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Bogucarskis, K.; Gatto, F.; Vandekerkhove, J.; Deriu, I.; Cardoso, A.C. Building the European Alien Species Information Network (EASIN): A Novel Approach for the Exploration of Distributed Alien Species Data. BioInvasions Rec. 2012, 1, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Olenin, S.; Narščius, A.; Minchin, D.; David, M.; Galil, B.; Gollasch, S.; Marchini, A.; Occhipinti-Ambrogi, A.; Ojaveer, H.; Zaiko, A. Making non-indigenous species information systems practical for management and useful for research: An aquatic perspective. Biol. Conserv. 2014, 173, 98–107. [Google Scholar] [CrossRef]
- Rius, M.; Ahyong, S.; Bieler, R.; Boudouresque, C.; Costello, M.J.; Downey, R.; Galil, B.S.; Gollasch, S.; Hutchings, P.; Kamburska, L.; et al. World Register of introduced Marine Species (WRiMS). Available online: https://www.marinespecies.org/introduced (accessed on 15 September 2021).
- Ahyong, S.; Boyko, C.B.; Bailly, N.; Bernot, J.; Bieler, R.; Brandão, S.N.; Daly, M.; De Grave, S.; Gofas, S.; Hernandez, F.; et al. World Register of Marine Species (WoRMS). Available online: https://www.marinespecies.org (accessed on 17 September 2021).
- Duarte, S.; Vieira, P.E.; Leite, B.R.; Teixeira, M.A.L.; Neto, J.M.; Costa, F.O. Comparing DNA metabarcoding with morphology in the assessment of macrozoobenthos in Portuguese transitional waters in the scope of the water framework directive monitoring. Biorxiv 2022, 1–38. [Google Scholar] [CrossRef]
- Leray, M.; Yang, J.Y.; Meyer, C.P.; Mills, S.C.; Agudelo, N.; Ranwez, V.; Boehm, J.T.; Machida, R.J. A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Front. Zool. 2013, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Lobo, J.; Costa, P.M.; Al Teixeira, M.; Ferreira, M.S.; Costa, M.H.; O Costa, F. Enhanced primers for amplification of DNA barcodes from a broad range of marine metazoans. BMC Ecol. 2013, 13, 34. [Google Scholar] [CrossRef] [Green Version]
- Ratnasingham, S. MBRAVE: The Multiplex Barcode Research and Visualization Environment. Biodivers. Inf. Sci. Stand. 2019, 3, e37986. [Google Scholar] [CrossRef]
- Zouros, E. The exceptional mitochondrial DNA system of the mussel family Mytilidae. Genes Genet. Syst. 2000, 75, 313–318. [Google Scholar] [CrossRef] [Green Version]
- Leese, F.; Bouchez, A.; Abarenkov, K.; Altermatt, F.; Borja, A.; Bruce, K.; Ekrem, T.; Čiampor, F.; Čiamporová-Zaťovičová, Z.; Costa, F.O.; et al. Why We Need Sustainable Networks Bridging Countries, Disciplines, Cultures and Generations for Aquatic Biomonitoring 2.0: A Perspective Derived from the DNAqua-Net COST Action. Adv. Ecol. Res. 2018, 58, 63–99. [Google Scholar] [CrossRef]
- Duarte, S.; Leite, B.R.; Feio, M.J.; Costa, F.O.; Filipe, A.F. Integration of DNA-Based Approaches in Aquatic Ecological Assessment Using Benthic Macroinvertebrates. Water 2021, 13, 331. [Google Scholar] [CrossRef]
- Aylagas, E.; Borja, Á.; Rodríguez-Ezpeleta, N. Environmental Status Assessment Using DNA Metabarcoding: Towards a Genetics Based Marine Biotic Index (gAMBI). PLoS ONE 2014, 9, e90529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, C.O. Taxonomy in times of the taxonomic impediment—Examples from the community of experts on amphipod crustaceans. J. Crustac. Biol. 2015, 35, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Christodoulou, M.; van der Hoorn, B.; Van den Bulcke, L.; Derycke, S.; De Backer, A.; Kröncke, I.; Arbizu, P.M. A reliable DNA barcode reference library for the identification of benthic invertebrates: Essential for biomonitoring of the North Sea. ARPHA Conf. Abstr. 2021, 4, 10–11. [Google Scholar] [CrossRef]
- Gissi, C.; Hastings, K.E.M.; Gasparini, F.; Stach, T.; Pennati, R.; Manni, L. An unprecedented taxonomic revision of a model organism: The paradigmatic case of Ciona robusta and Ciona intestinalis. Zool. Scr. 2017, 46, 521–522. [Google Scholar] [CrossRef]
- Couton, M.; Comtet, T.; Le Cam, S.; Corre, E.; Viard, F. Metabarcoding on planktonic larval stages: An efficient approach for detecting and investigating life cycle dynamics of benthic aliens. Manag. Biol. Invasions 2019, 10, 657–689. [Google Scholar] [CrossRef] [Green Version]
- Viard, F.; Roby, C.; Turon, X.; Bouchemousse, S.; Bishop, J. Cryptic Diversity and Database Errors Challenge Non-Indigenous Species Surveys: An Illustration with Botrylloides spp. in the English Channel and Mediterranean Sea. Front. Mar. Sci. 2019, 6, 615. [Google Scholar] [CrossRef] [Green Version]
- Lacoursière-Roussel, A.; Howland, K.; Normandeau, E.; Grey, E.K.; Archambault, P.; Deiner, K.; Lodge, D.M.; Hernandez, C.; LeDuc, N.; Bernatchez, L. eDNA metabarcoding as a new surveillance approach for coastal Arctic biodiversity. Ecol. Evol. 2018, 8, 7763–7777. [Google Scholar] [CrossRef]
- Costa, F.O.; Antunes, P.M. The contribution of the barcode of life initiative to the discovery and monitoring of biodiversity. In Natural Resources, Sustainability and Humanity; Mendonca, A., Cunha, A., Chakrabarti, R., Eds.; Springer: Dordrecht, The Netherlands, 2012; ISBN 9789400713215. [Google Scholar]
- Rimet, F.; Chaumeil, P.; Keck, F.; Kermarrec, L.; Vasselon, V.; Kahlert, M.; Franc, A.; Bouchez, A. R-Syst: Diatom: An open-access and curated barcode database for diatoms and freshwater monitoring. Database 2016, 2016, baw016. [Google Scholar] [CrossRef] [Green Version]
- Mackie, J.A.; Darling, J.A.; Geller, J.B. Ecology of cryptic invasions: Latitudinal segregation among Watersipora (Bryozoa) species. Sci. Rep. 2012, 2, srep00871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehlauer-Ale, K.H.; Mackie, J.A.; Lim-Fong, G.E.; Ale, E.; Pie, M.R.; Waeschenbach, A. Cryptic species in the cosmopolitan Bugula neritina complex (Bryozoa, Cheilostomata). Zool. Scr. 2014, 43, 193–205. [Google Scholar] [CrossRef]
- Lobo-Arteaga, J.; Ferreira, M.S.; Antunes, I.C.; Teixeira, M.A.L.; Borges, L.M.S.; Sousa, R.; Gomes, P.A.; Costa, M.H.; Cunha, M.R.; Costa, F. Contrasting morphological and DNA barcode-suggested species boundaries among shallow-water amphipod fauna from the southern European Atlantic coast. Genome 2017, 60, 147–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, P.E.; Desiderato, A.; Azevedo, S.L.; Esquete, P.; Costa, F.O.; Queiroga, H. Molecular evidence for extensive discontinuity between peracarid (Crustacea) fauna of Macaronesian islands and nearby continental coasts: Over fifty candidate endemic species. Mar. Biol. 2022, 169, 64. [Google Scholar] [CrossRef]
- Teixeira, M.A.L.; E Vieira, P.; Ravara, A.; O Costa, F.; Nygren, A. From 13 to 22 in a second stroke: Revisiting the European Eumida sanguinea (Phyllodocidae: Annelida) species complex. Zool. J. Linn. Soc. 2022, 196, 169–197. [Google Scholar] [CrossRef]
- Stoeckle, M.Y.; Das Mishu, M.; Charlop-Powers, Z. Improved Environmental DNA Reference Library Detects Overlooked Marine Fishes in New Jersey, United States. Front. Mar. Sci. 2020, 7, 226. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Species | Authority | Non-Curated | Audited | Audited + Curated |
|---|---|---|---|---|---|
| Arthropoda | Amphibalanus amphitrite | (Darwin, 1854) | 0 | 499 | 499 |
| Arthropoda | Amphibalanus improvisus | (Darwin, 1854) | 0 | 0 | 698 |
| Arthropoda | Ampithoe valida | Smith, 1873 | 4 | 4 | 4 |
| Arthropoda | Austrominius modestus | (Darwin, 1854) | 0 | 469 | 469 |
| Bryozoa | Bugula neritina | (Linnaeus, 1758) | 1 | 0 | 1 |
| Arthropoda | Carcinus maenas | (Linnaeus, 1758) | 93 | 93 | 93 |
| Cnidaria | Ectopleura crocea | (Agassiz, 1862) | 657 | 0 | 657 |
| Cnidaria | Haliclystus tenuis* | Kishinouye, 1910 | 2 | 0 | 0 |
| Arthropoda | Monocorophium acherusicum | (Costa, 1853) | 3677 | 3676 | 3676 |
| Mollusca | Mya arenaria | Linnaeus, 1758 | 1 | 0 | 1 |
| Arthropoda | Mytilicola intestinalis | Steuer, 1902 | 0 | 1 | 1 |
| Mollusca | Ostrea stentina | Payraudeau, 1826 | 6 | 0 | 6 |
| Arthropoda | Paracalanus indicus | Wolfenden, 1905 | 0 | 0 | 54 |
| Arthropoda | Perforatus perforatus | (Bruguière, 1789) | 0 | 207 | 209 |
| Annelida | Polydora cornuta | Bosc, 1802 | 5 | 5 | 5 |
| Arthropoda | Pseudodiaptomus marinus | Sato, 1913 | 0 | 54 | 54 |
| Mollusca | Ruditapes philippinarum | (A. Adams and Reeve, 1850) | 0 | 0 | 51 |
| Bryozoa | Scruparia ambigua * | (d’Orbigny, 1841) | 1 | 0 | 0 |
| Annelida | Syllidia armata * | Quatrefages, 1866 | 4 | 0 | 0 |
| Annelida | Terebella lapidaria * | Linnaeus, 1767 | 76 | 0 | 0 |
| Total NIS | 12 | 9 | 16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lavrador, A.S.; Fontes, J.T.; Vieira, P.E.; Costa, F.O.; Duarte, S. Compilation, Revision, and Annotation of DNA Barcodes of Marine Invertebrate Non-Indigenous Species (NIS) Occurring in European Coastal Regions. Diversity 2023, 15, 174. https://doi.org/10.3390/d15020174
Lavrador AS, Fontes JT, Vieira PE, Costa FO, Duarte S. Compilation, Revision, and Annotation of DNA Barcodes of Marine Invertebrate Non-Indigenous Species (NIS) Occurring in European Coastal Regions. Diversity. 2023; 15(2):174. https://doi.org/10.3390/d15020174
Chicago/Turabian StyleLavrador, Ana S., João T. Fontes, Pedro E. Vieira, Filipe O. Costa, and Sofia Duarte. 2023. "Compilation, Revision, and Annotation of DNA Barcodes of Marine Invertebrate Non-Indigenous Species (NIS) Occurring in European Coastal Regions" Diversity 15, no. 2: 174. https://doi.org/10.3390/d15020174