

Resource Partitioning by Corallivorous Snails on Bonaire (Southern Caribbean)


Abstract
:1. Introduction
2. Materials and Methods
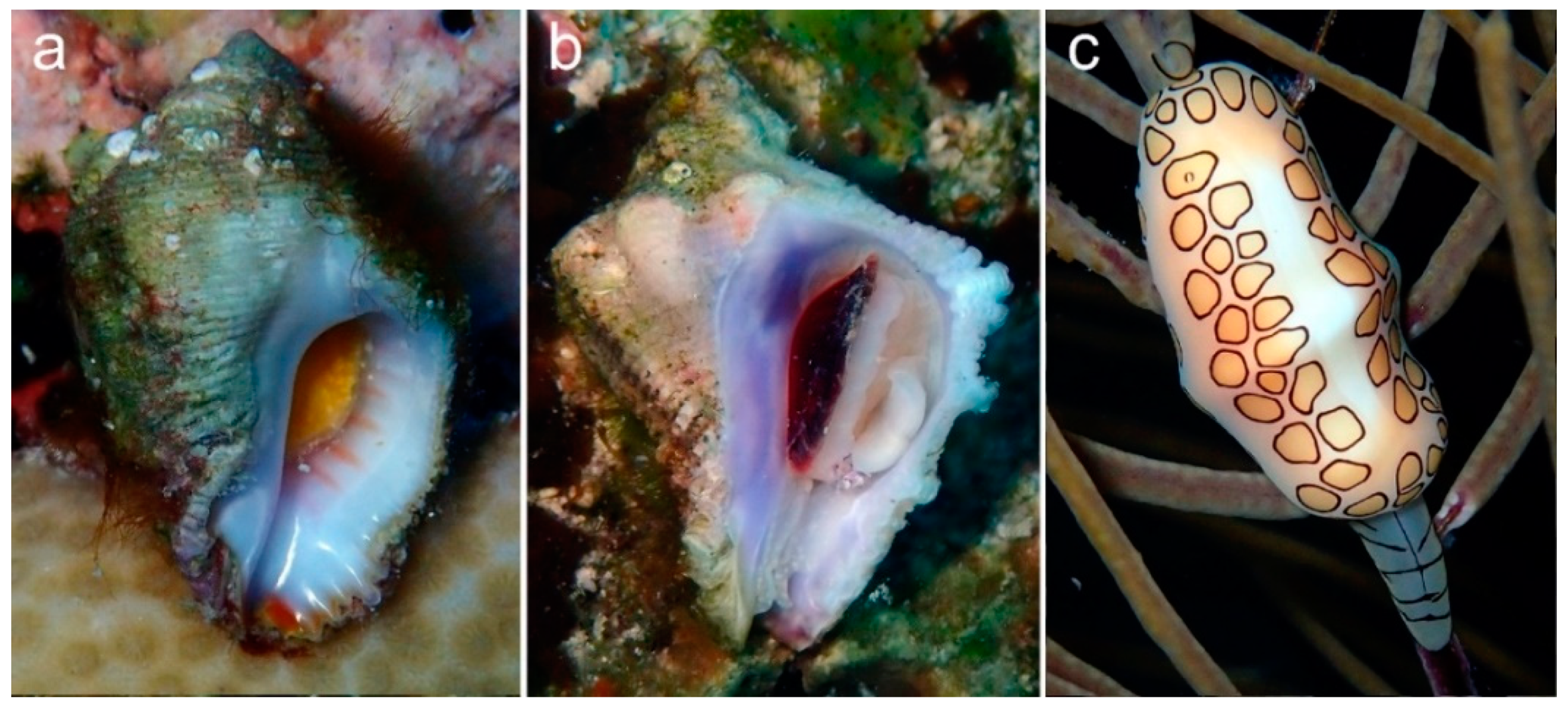
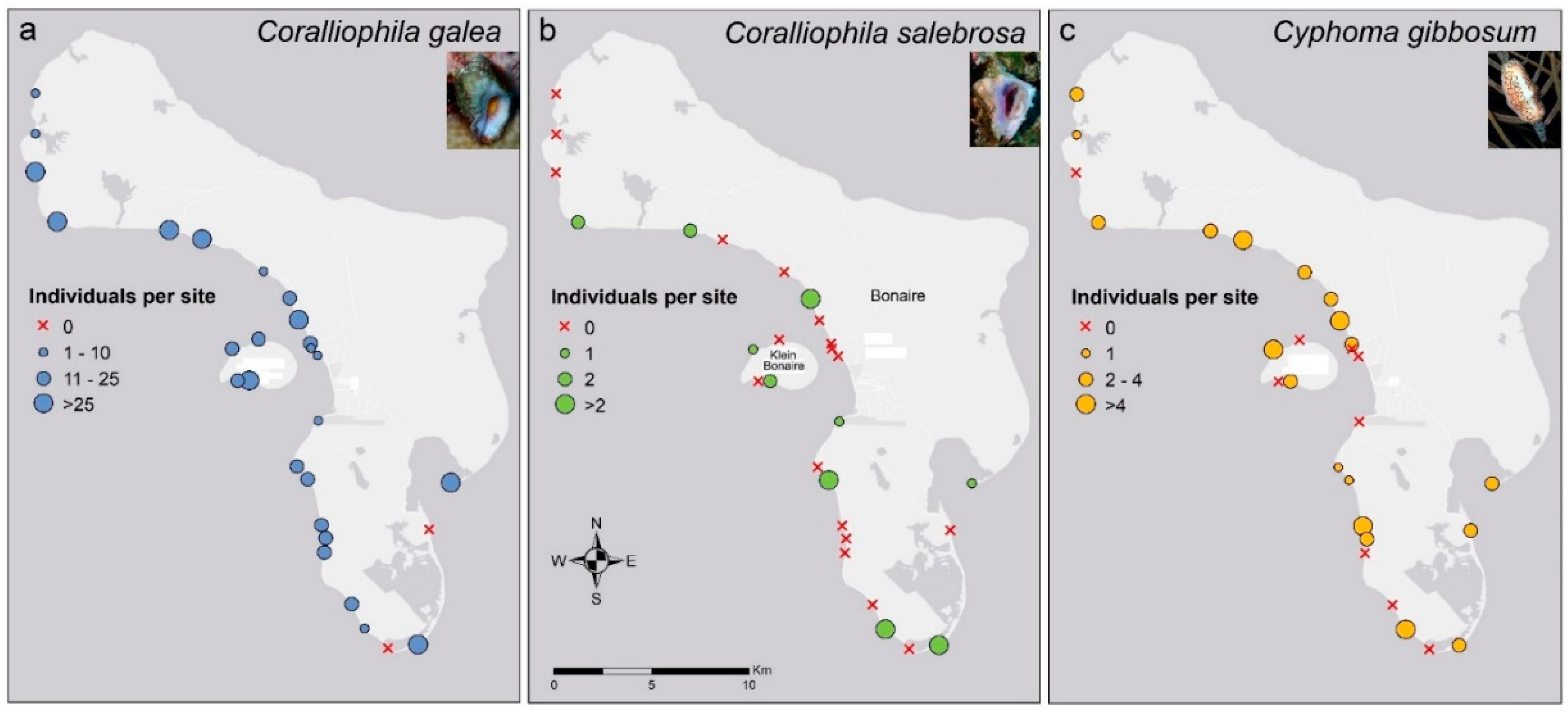
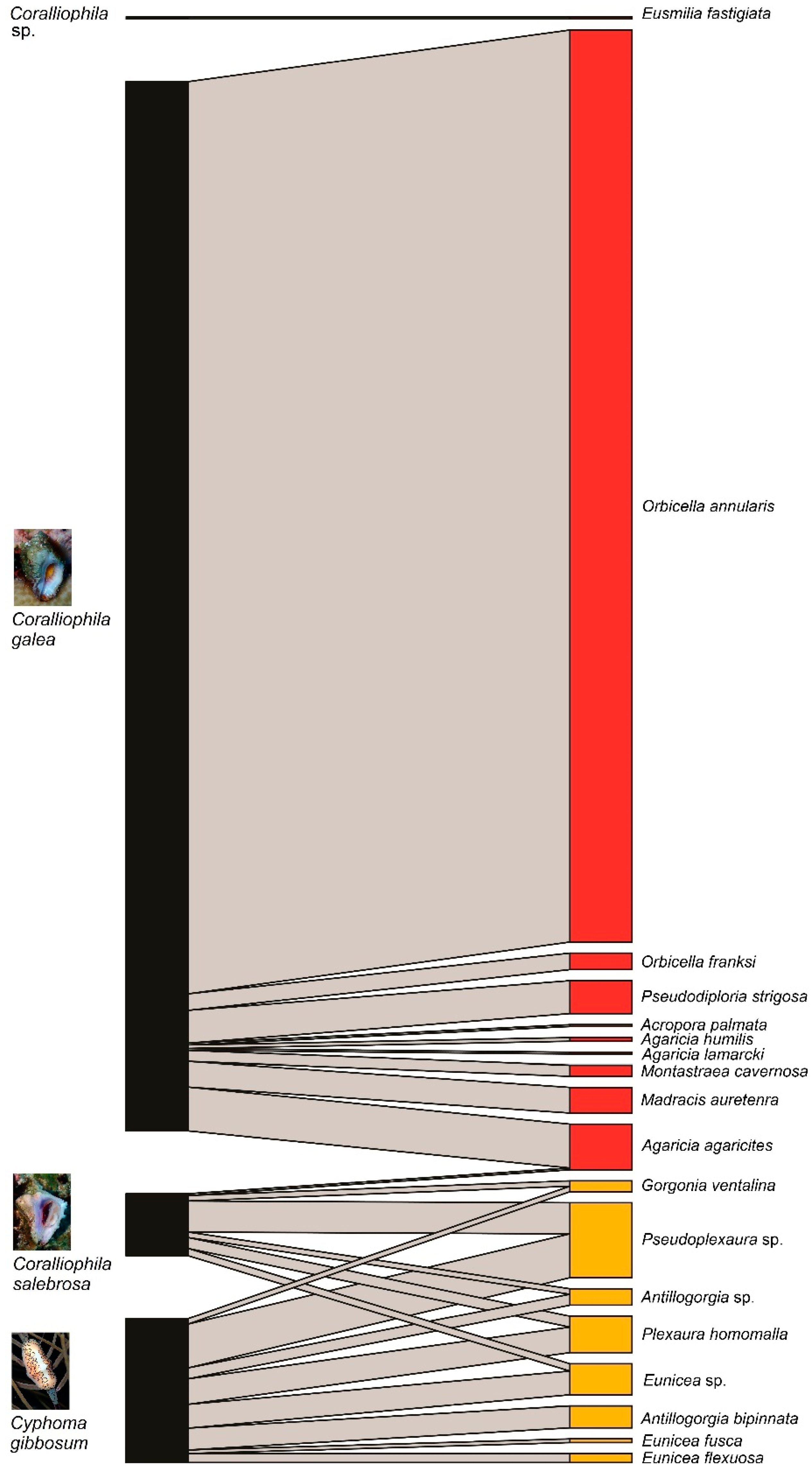
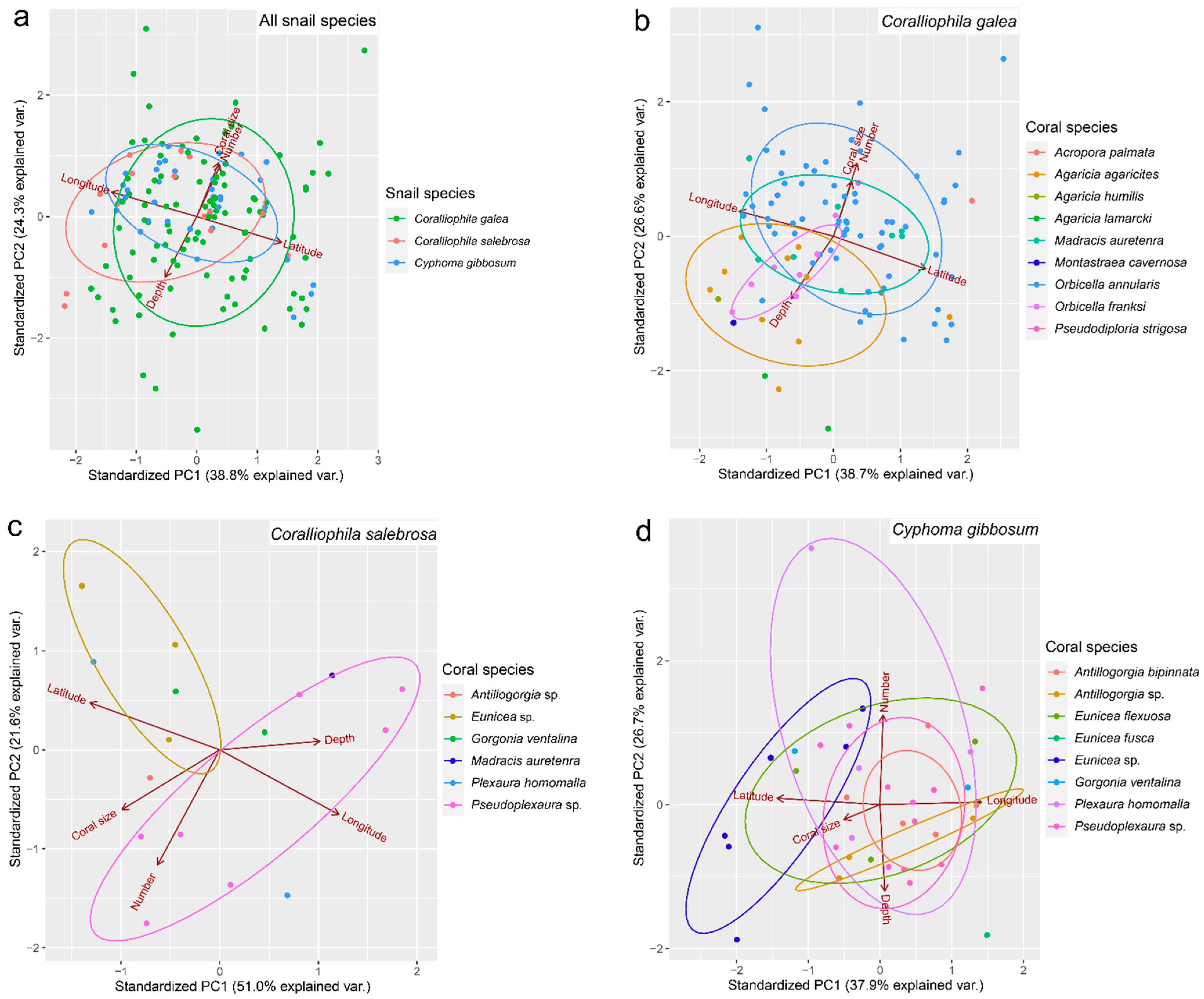
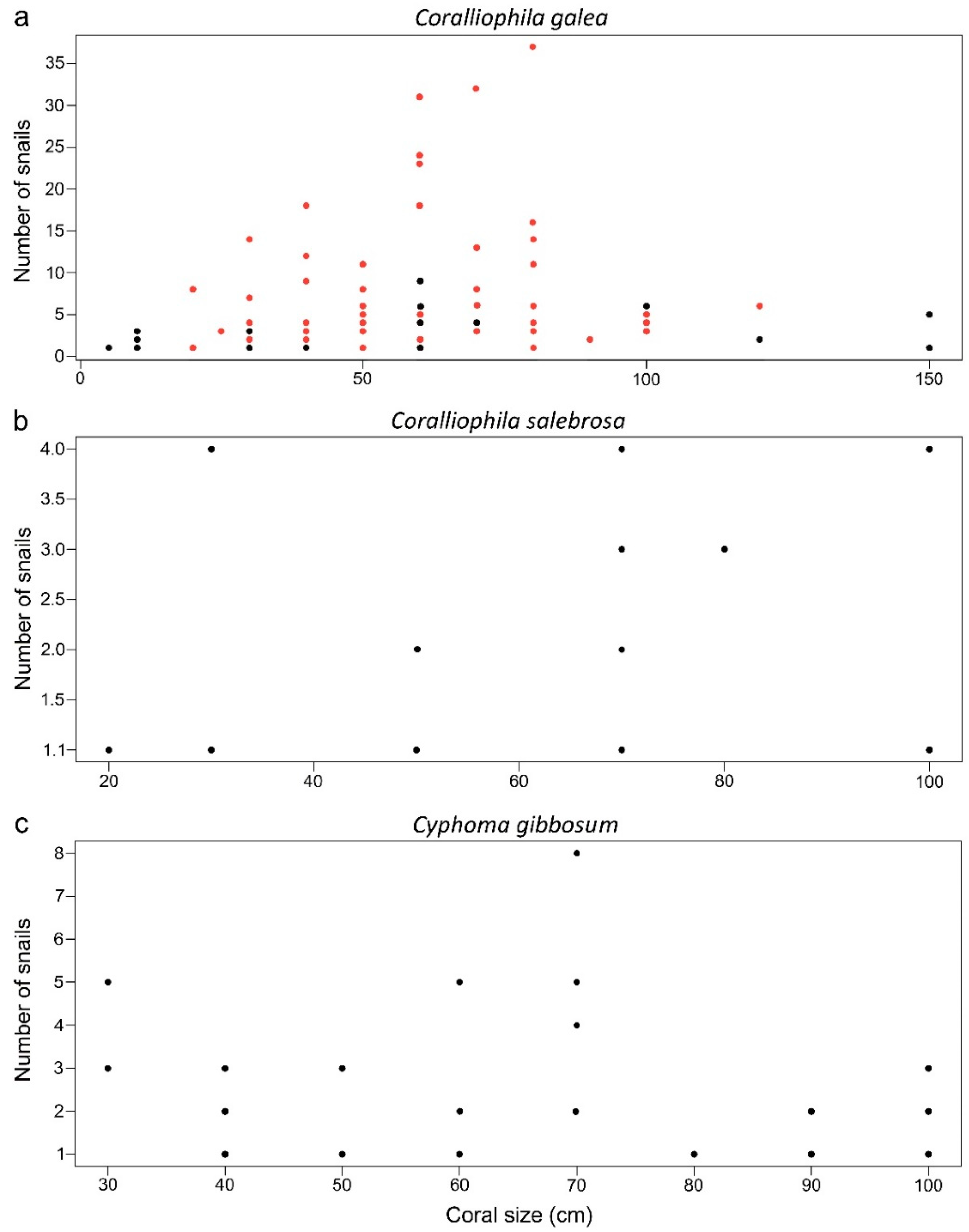
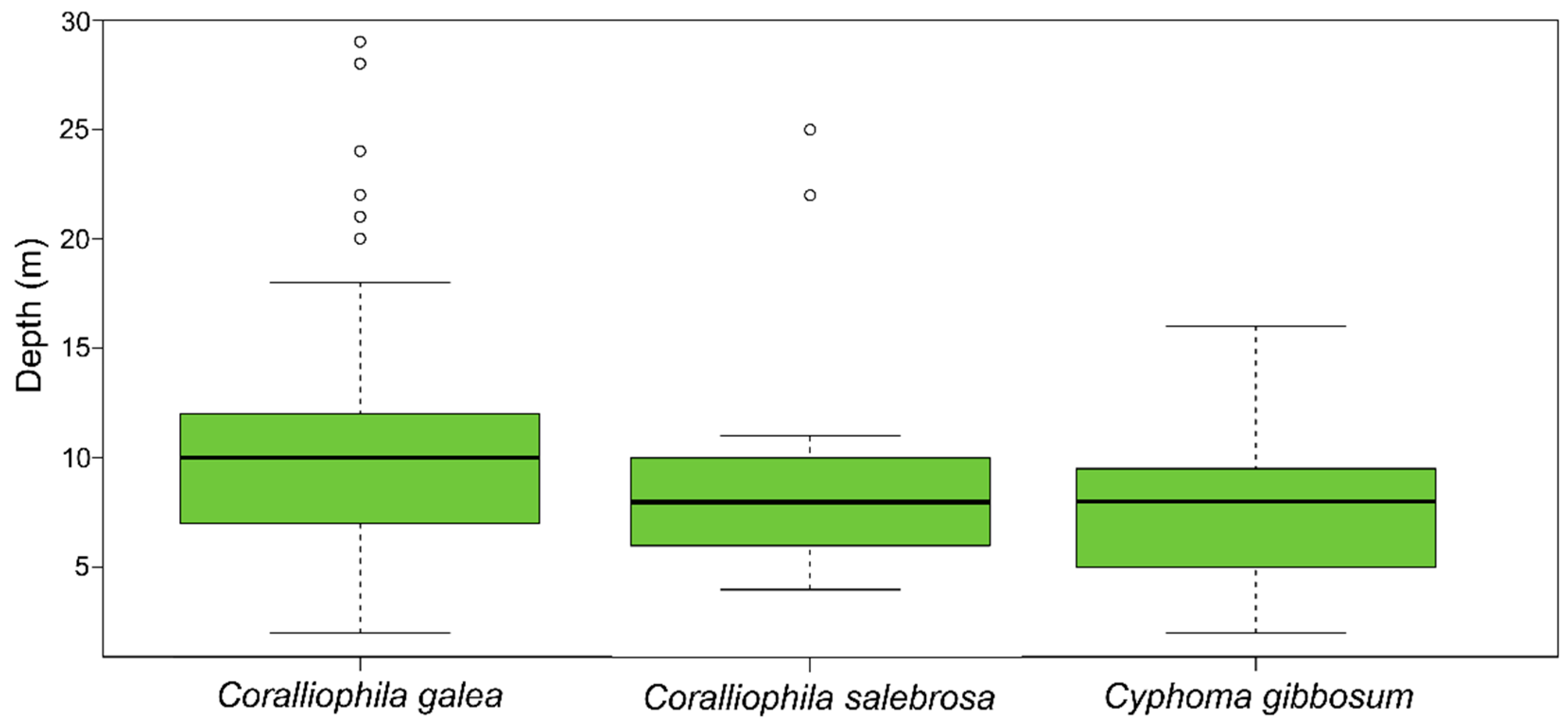
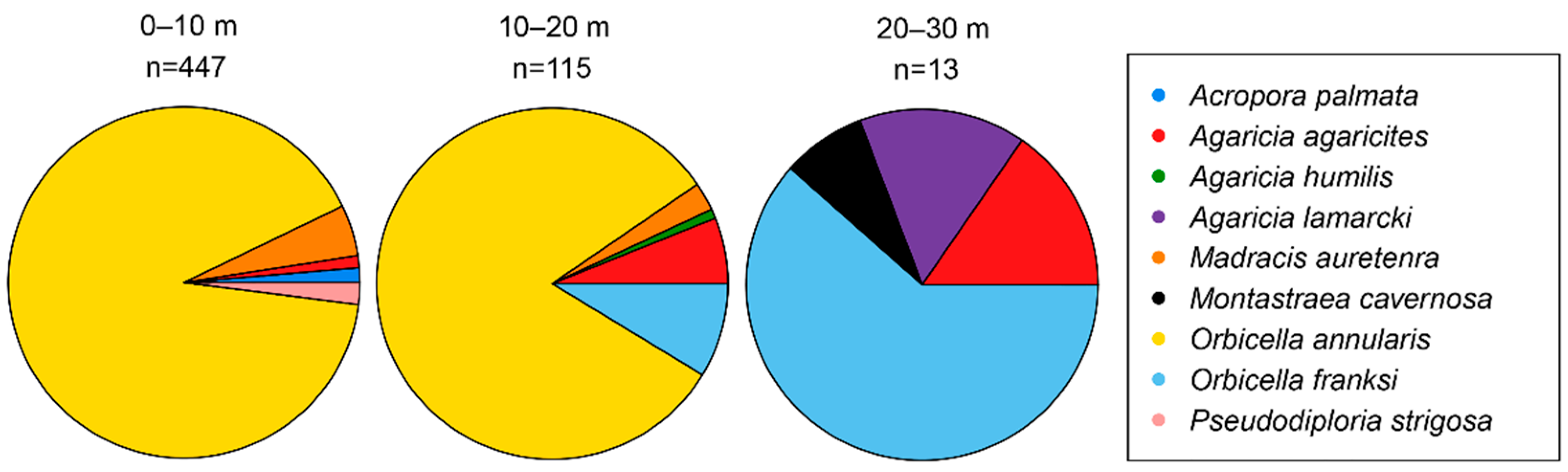
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reaka-Kudla, M.L. Biodiversity of Caribbean coral reefs. In Caribbean Marine Biodiversity: The Known and the Unknown; Miloslavich, P., Klein, E., Eds.; DEStech Publishers: Lancaster, PA, USA, 2005; pp. 259–276. [Google Scholar]
- Miloslavich, P.; Díaz, J.M.; Klein, E.; Alvarado, J.J.; Díaz, C.; Gobin, J.; Escobar-Briones, E.; Cruz-Motta, J.J.; Weil, E.; Cortés, J.; et al. Marine biodiversity in the Caribbean: Regional estimates and distribution patterns. PLoS ONE 2010, 5, e11916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeVantier, L.; Turak, E.; Szava-Kovats, R. Species richness and abundance of reef-building corals in the Indo-West Pacific: The local–regional relation revisited. Front. Mar. Sci. 2020, 7, 487. [Google Scholar] [CrossRef]
- Stella, J.S.; Pratchett, M.S.; Hutchings, P.A.; Jones, G.P. Coral-associated invertebrates: Diversity, ecological importance and vulnerability to disturbance. Oceanogr. Mar. Biol. Annu. Rev. 2011, 49, 43–104. [Google Scholar]
- Hoeksema, B.W.; van der Meij, S.E.T.; Fransen, C.H.J.M. The mushroom coral as a habitat. J. Mar. Biol. Assoc. U. K. 2012, 92, 647–663. [Google Scholar] [CrossRef] [Green Version]
- Nützel, A. Gastropods as parasites and carnivorous grazers: A major guild in marine ecosystems. In The Evolution and Fossil Record of Parasitism; De Baets, K., Huntley, J.W., Eds.; Springer: Cham, Switzerland, 2021; pp. 209–229. [Google Scholar] [CrossRef]
- Lucas, M.Q.; Rodríguez, L.R.; Sanabria, D.J.; Weil, E. Natural prey preferences and spatial variability of predation pressure by Cyphoma gibbosum (Mollusca: Gastropoda) on octocoral communities off La Parguera, Puerto Rico. Int. Sch. Res. Notices 2014, 2014, 742387. [Google Scholar] [CrossRef] [Green Version]
- Pinto, T.K.; Benevides, L.J.; Sampaio, C.L.S. Cyphoma macumba Petuch 1979 (Gastropoda: Ovulidae): A versatile predator of the Brazilian Octocorallia. Mar. Biodivers. 2017, 47, 165–166. [Google Scholar] [CrossRef]
- Shaver, E.C.; Renzi, J.J.; Bucher, M.G.; Silliman, B.R. Relationships between a common Caribbean corallivorous snail and protected area status, coral cover, and predator abundance. Sci. Rep. 2020, 10, 16463. [Google Scholar] [CrossRef]
- Gittenberger, A.; Hoeksema, B.W. Habitat preferences of coral-associated wentletrap snails (Gastropoda: Epitoniidae). Contrib. Zool. 2013, 82, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, B.W.; Scott, C.; True, J.D. Dietary shift in corallivorous Drupella snails following a major bleaching event at Koh Tao, Gulf of Thailand. Coral Reefs 2013, 32, 423–428. [Google Scholar] [CrossRef]
- Potkamp, G.; Vermeij, M.J.A.; Hoeksema, B.W. Genetic and morphological variation in corallivorous snails (Coralliophila spp.) living on different host corals at Curaçao, southern Caribbean. Contrib. Zool. 2017, 86, 111–144. [Google Scholar] [CrossRef]
- Kaullysing, D.; Taleb-Hossenkhan, N.; Kulkarni, B.; Bhagooli, R. Variations in the density of two ectoparasitic gastropods (Coralliophila spp.) on scleractinian corals on a coast-reef scale. Symbiosis 2019, 78, 65–71. [Google Scholar] [CrossRef]
- Moerland, M.S.; Scott, C.M.; Hoeksema, B.W. Prey selection of corallivorous muricids at Koh Tao (Gulf of Thailand) four years after a major coral bleaching event. Contrib. Zool. 2016, 85, 291–309. [Google Scholar] [CrossRef] [Green Version]
- Saponari, L.; Dehnert, I.; Galli, P.; Montano, S. Assessing population collapse of Drupella spp. (Mollusca: Gastropoda) 2 years after a coral bleaching event in the Republic of Maldives. Hydrobiologia 2021, 848, 2653–2666. [Google Scholar] [CrossRef]
- Sánchez, J.A. Remarkable specialization in Eastern Pacific sea fan ectoparasites (Neosimnia). Coral Reefs 2013, 32, 891. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, J.A.; Fuentes-Pardo, A.P.; Almhain, Í.N.; Ardila-Espitia, N.E.; Cantera-Kintz, J.; Forero-Shelton, M. The masquerade game: Marine mimicry adaptation between egg-cowries and octocorals. PeerJ 2016, 4, e2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gittenberger, A.; Gittenberger, E. A hitherto unnoticed adaptive radiation: Epitoniid species (Gastropoda: Epitoniidae) associated with corals (Scleractinia). Contrib. Zool. 2005, 74, 125–203. [Google Scholar] [CrossRef] [Green Version]
- Oliverio, M. Coralliophilinae (Neogastropoda: Muricidae) from the southwest Pacific. Mem. Mus. Natl. Hist. Nat. 2008, 196, 481–585. [Google Scholar]
- Taviani, M.; Angeletti, L.; Dimech, M.; Mifsud, C.; Freiwald, A.; Harasewych, M.G.; Oliverio, M. Coralliophilinae (Gastropoda: Muricidae) associated with deep-water coral banks in the Mediterranean. Nautilus 2009, 123, 106–112. [Google Scholar]
- Claremont, M.; Houart, R.; Williams, S.T.; Reid, D.G. A molecular phylogenetic framework for the Ergalataxinae (Neogastropoda: Muricidae). J. Mollusc. Stud. 2013, 79, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Goud, J.; Hoeksema, B.W. Pedicularia vanderlandi spec. nov., a symbiotic snail (Caenogastropoda: Ovulidae) on the hydrocoral Distichopora vervoorti Cairns and Hoeksema, 1998 (Hydrozoa: Stylasteridae), from Bali, Indonesia. Zool. Verh. 2001, 334, 77–97. [Google Scholar]
- Lorenz, F.; Fehse, D. The Living Ovulidae. A Manual of the Families of Allied Cowries: Ovulidae, Pediculariidae and Eocypraeidae; Conchbooks: Hackenheim, Germany, 2009. [Google Scholar]
- Braga-Henriques, A.; Carreiro-Silva, M.; Porteiro, F.M.; de Matos, V.; Sampaio, Í.; Ocaña, O.; Avila, S.P. The association between a deep-sea gastropod Pedicularia sicula (Caenogastropoda: Pediculariidae) and its coral host Errina dabneyi (Hydrozoa: Stylasteridae) in the Azores. ICES J. Mar. Sci. 2011, 68, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Schiaparelli, S.; Barucca, M.; Olmo, E.; Boyer, M.; Canapa, A. Phylogenetic relationships within Ovulidae (Gastropoda: Cypraeoidea) based on molecular data from the 16S rRNA gene. Mar. Biol. 2005, 147, 411–420. [Google Scholar] [CrossRef]
- Reijnen, B.T.; Hoeksema, B.W.; Gittenberger, E. Host specificity and phylogenetic relationships among Atlantic Ovulidae (Mollusca: Gastropoda). Contrib. Zool. 2010, 79, 69–78. [Google Scholar] [CrossRef]
- Fritts-Penniman, A.L.; Gosliner, T.M.; Mahardika, G.N.; Barber, P.H. Cryptic ecological and geographic diversification in coral-associated nudibranchs. Mol. Phylogenet. Evol. 2020, 144, 106698. [Google Scholar] [CrossRef]
- Mehrotra, R.; Arnold, S.; Wang, A.; Chavanich, S.; Hoeksema, B.W.; Caballer, M. A new species of coral-feeding nudibranch (Mollusca: Gastropoda) from the Gulf of Thailand. Mar. Biodivers. 2020, 50, 36. [Google Scholar] [CrossRef]
- Wang, A.; Conti-Jerpe, I.E.; Richards, J.L.; Baker, D.M. Phestilla subodiosus sp. nov. (Nudibranchia, Trinchesiidae), a corallivorous pest species in the aquarium trade. ZooKeys 2020, 909, 35278. [Google Scholar] [CrossRef]
- Yiu, S.K.F.; Chung, S.S.W.; Qiu, J.W. New observations on the corallivorous nudibranch Phestilla melanobrachia: Morphology, dietary spectrum and early development. J. Mollusc. Stud. 2021, 87, eyab034. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Gittenberger, A. Records of some marine parasitic molluscs from Nha Trang, Vietnam. Basteria 2008, 72, 129–133. [Google Scholar]
- Gittenberger, A.; Gittenberger, E. Cryptic, adaptive radiation of endoparasitic snails: Sibling species of Leptoconchus (Gastropoda: Coralliophilidae) in corals. Org. Divers. Evol. 2011, 11, 21–41. [Google Scholar] [CrossRef] [Green Version]
- Bieler, R.; Petit, R.E. Catalogue of Recent and fossil “worm-snail” taxa of the families Vermetidae, Siliquariidae, and Turritellidae (Mollusca: Caenogastropoda). Zootaxa 2011, 2948, 1–103. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, B.W.; Harper, C.E.; Langdon-Down, S.J.; van der Schoot, R.J.; Smith-Moorhouse, A.; Spaargaren, R.; Timmerman, R.F. Host range of the coral-associated worm snail Petaloconchus sp. (Gastropoda: Vermetidae), a newly discovered cryptogenic pest species in the southern Caribbean. Diversity 2022, 14, 196. [Google Scholar] [CrossRef]
- Knowlton, N.; Lang, J.C.; Keller, B.D. Case study of natural population collapse: Post-hurricane predation on Jamaican staghorn corals. Smithson. Contrib. Mar. Sci. 1990, 31, 1–25. [Google Scholar] [CrossRef]
- Bruckner, A.W.; Bruckner, R.J. Mechanical lesions and corallivory. In Diseases of Coral; Woodley, C.M., Downs, C.A., Bruckner, A.W., Porter, J.W., Galloway, S.B., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 242–265. [Google Scholar] [CrossRef]
- Scott, C.M.; Mehrotra, R.; Hein, M.Y.; Moerland, M.S.; Hoeksema, B.W. Population dynamics of corallivores (Drupella and Acanthaster) on coral reefs of Koh Tao, a diving destination in the Gulf of Thailand. Raffles Bull. Zool. 2017, 65, 68–79. [Google Scholar]
- Boucher, L.M. Coral predation by muricid gastropods of the genus Drupella at Enewetak, Marshall Islands. Bull. Mar. Sci. 1986, 38, 9–11. [Google Scholar]
- Cumming, R.; McCorry, D. Corallivorous gastropods in Hong Kong. Coral Reefs 1998, 17, 178. [Google Scholar] [CrossRef]
- Cumming, R.L. Predation on reef-building corals: Multiscale variation in the density of three corallivorous gastropods, Drupella spp. Coral Reefs 1999, 18, 147–157. [Google Scholar] [CrossRef]
- Schärer, M.T.; Nemeth, M.I. Mass mortality of gorgonians due to a Cyphoma gibbosum (Linnaeus) population outbreak at Mona Island, Puerto Rico. Coral Reefs 2010, 29, 533. [Google Scholar] [CrossRef] [Green Version]
- Bessey, C.; Babcock, R.C.; Thomson, D.P.; Haywood, M.D.E. Outbreak densities of the coral predator Drupella in relation to in situ Acropora growth rates on Ningaloo Reef, Western Australia. Coral Reefs 2018, 37, 985–993. [Google Scholar] [CrossRef]
- Bruckner, A.W.; Coward, G.; Bimson, K.; Rattanawongwan, T. Predation by feeding aggregations of Drupella spp. inhibits the recovery of reefs damaged by a mass bleaching event. Coral Reefs 2017, 36, 1181–1187. [Google Scholar] [CrossRef]
- Hamman, E.A. Aggregation patterns of two corallivorous snails and consequences for coral dynamics. Coral Reefs 2018, 37, 851–860. [Google Scholar] [CrossRef]
- Dalton, S.J.; Godwin, S. Progressive coral tissue mortality following predation by a corallivorous nudibranch (Phestilla sp.). Coral Reefs 2006, 25, 529. [Google Scholar] [CrossRef]
- Gignoux-Wolfsohn, S.; Marks, C.; Vollmer, S. White Band Disease transmission in the threatened coral, Acropora cervicornis. Sci. Rep. 2012, 2, 804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymundo, L.J.; Work, T.M.; Miller, R.L.; Lozada-Misa, P.L. Effects of Coralliophila violacea on tissue loss in the scleractinian corals Porites spp. depend on host response. Dis. Aquat. Organ. 2016, 119, 75–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolet, K.J.; Chong-Seng, K.M.; Pratchett, M.S.; Willis, B.L.; Hoogenboom, M.O. Predation scars may influence host susceptibility to pathogens: Evaluating the role of corallivores as vectors of coral disease. Sci. Rep. 2018, 8, 5258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montano, S.; Aeby, G.; Galli, P.; Hoeksema, B.W. Feeding behavior of Coralliophila sp. on corals affected by Caribbean ciliate infection (CCI): A new possible vector? Diversity 2022, 14, 363. [Google Scholar] [CrossRef]
- Thur, S.M. User fees as sustainable financing mechanisms for marine protected areas: An application to the Bonaire National Marine Park. Mar. Policy 2010, 34, 63–69. [Google Scholar] [CrossRef]
- Trembanis, A.C.; Forrest, A.L.; Keller, B.M.; Patterson, M.R. Mesophotic coral ecosystems: A geoacoustically derived proxy for habitat and relative diversity for the leeward shelf of Bonaire, Dutch Caribbean. Front. Mar. Sci. 2017, 4, 51. [Google Scholar] [CrossRef] [Green Version]
- Bak, R.P.M.; Nieuwland, G.; Meesters, E.H. Coral reef crisis in deep and shallow reefs: 30 years of constancy and change in reefs of Curaçao and Bonaire. Coral Reefs 2005, 24, 475–479. [Google Scholar] [CrossRef]
- Stokes, M.D.; Leichter, J.J.; Genovese, S.J. Long-term declines in coral cover at Bonaire, Netherlands Antilles. Atoll Res. Bull. 2010, 582, 1–21. [Google Scholar] [CrossRef]
- de Bakker, D.M.; Meesters, E.H.; Bak, R.P.M.; Nieuwland, G.; van Duyl, F.C. Long-term shifts in coral communities on shallow to deep reef slopes of Curaçao and Bonaire: Are there any winners? Front. Mar. Sci. 2016, 3, 247. [Google Scholar] [CrossRef] [Green Version]
- de Bakker, D.M.; van Duyl, F.C.; Bak, R.P.M.; Nugues, M.M.; Nieuwland, G.; Meesters, E.H. 40 years of benthic community change on the Caribbean reefs of Curaçao and Bonaire: The rise of slimy cyanobacterial mats. Coral Reefs 2017, 36, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, B.W. (Ed.) Marine Biodiversity of Bonaire: A Baseline Survey, 2nd ed.; Naturalis Biodiversity Center: Leiden, The Netherlands; ANEMOON Foundation: Bennebroek, The Netherlands, 2022. [Google Scholar]
- García-Hernández, J.E.; de Gier, W.; van Moorsel, G.W.N.M.; Hoeksema, B.W. The scleractinian Agaricia undata as a new host for the Caribbean coral gall crab Opecarcinus hypostegus at Bonaire, southern Caribbean. Symbiosis 2020, 81, 303–311. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; García-Hernández, J.E. Host-related morphological variation of dwellings inhabited by the crab Domecia acanthophora in the corals Acropora palmata and Millepora complanata (Southern Caribbean). Diversity 2020, 12, 143. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, B.W.; García-Hernández, J.E.; van Moorsel, G.W.N.M.; Olthof, G.; ten Hove, H.A. Extension of the recorded host range of Caribbean Christmas tree worms (Spirobranchus spp.) with two scleractinians, a zoantharian, and an ascidian. Diversity 2020, 12, 115. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, B.W.; Timmerman, R.F.; Spaargaren, R.; Smith-Moorhouse, A.; van der Schoot, R.J.; Langdon-Down, S.J.; Harper, C.E. Morphological modifications and injuries of corals caused by symbiotic feather duster worms (Sabellidae) in the Caribbean. Diversity 2022, 14, 332. [Google Scholar] [CrossRef]
- Korzhavina, O.A.; Hoeksema, B.W.; Ivanenko, V.N. A review of Caribbean Copepoda associated with reef-dwelling cnidarians, echinoderms and sponges. Contrib. Zool. 2019, 88, 297–349. [Google Scholar] [CrossRef] [Green Version]
- Montano, S.; Reimer, J.D.; Ivanenko, V.N.; García-Hernández, J.E.; van Moorsel, G.W.N.M.; Galli, P.; Hoeksema, B.W. Widespread occurrence of a rarely known association between the hydrocorals Stylaster roseus and Millepora alcicornis at Bonaire, southern Caribbean. Diversity 2020, 12, 218. [Google Scholar] [CrossRef]
- Montenegro, J.; Hoeksema, B.W.; Santos, M.E.A.; Kise, H.; Reimer, J.D. Zoantharia (Cnidaria: Hexacorallia) of the Dutch Caribbean with historical distribution records from the Atlantic and one new species of Parazoanthus. Diversity 2020, 12, 190. [Google Scholar] [CrossRef]
- Reimer, J.D.; Wee, H.B.; García-Hernández, J.E.; Hoeksema, B.W. Same but different? Zoantharian assemblages (Anthozoa: Hexacorallia: Zoantharia) in Bonaire and Curaçao, southern Caribbean. Coral Reefs 2022, 41, 383–396. [Google Scholar] [CrossRef]
- Rotjan, R.D.; Lewis, S.M. Impact of coral predators on tropical reefs. Mar. Ecol. Prog. Ser. 2008, 367, 73–91. [Google Scholar] [CrossRef]
- Miller, M. Corallivorous snail removal: Evaluation of impact on Acropora palmata. Coral Reefs 2001, 19, 293–295. [Google Scholar] [CrossRef]
- Precht, W.; Bruckner, A.; Aronson, R.; Bruckner, R. Endangered acroporid corals of the Caribbean. Coral Reefs 2002, 21, 41–42. [Google Scholar] [CrossRef]
- Garrigues, B.; Lamy, D.; Zuccon, D. The Coralliophilinae from the Antilles and French Guiana with the description of six new species. Xenophora Taxon. 2022, 37, 4–53. [Google Scholar]
- Johnston, L.; Miller, M.W. Variation in life-history traits of the corallivorous gastropod Coralliophila abbreviata on three coral hosts. Mar. Biol. 2007, 150, 1215–1225. [Google Scholar] [CrossRef]
- Johnston, L.; Miller, M.W.; Baums, I.B. Assessment of host-associated genetic differentiation among phenotypically divergent populations of a coral-eating gastropod across the Caribbean. PLoS ONE 2012, 7, e47630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmonds, S.E.; Chou, V.; Cheng, S.H.; Rachmawati, R.; Calumpong, H.P.; Ngurah Mahardika, G.; Barber, P.H. Evidence of host-associated divergence from coral-eating snails (genus Coralliophila) in the Coral Triangle. Coral Reefs 2018, 37, 355–371. [Google Scholar] [CrossRef]
- Miller, A.C. Observations on the associations and feeding of six species of prosobranch gastropods on anthozoans. Atoll Res. Bull. 1972, 152, 4–5. [Google Scholar]
- Martin, D.; Gil, J.; Abgarian, C.; Evans, E.; Turner, E.M.; Nygren, A. Coralliophila from Grand Cayman: Specialized coral predator or parasite? Coral Reefs 2014, 33, 1017. [Google Scholar] [CrossRef] [Green Version]
- Ott, B.; Lewis, J.B. The importance of the gastropod Coralliophila abbreviata (Lamarck) and the polychaete Hermodice carunculata (Pallas) as coral reef predators. Can. J. Zool. 1972, 50, 1651–1656. [Google Scholar] [CrossRef]
- Reijnen, B.T.; van der Meij, S.E.T. Coat of many colours-DNA reveals polymorphism of mantle patterns and colouration in Caribbean Cyphoma Röding, 1798 (Gastropoda, Ovulidae). PeerJ 2017, 5, e3018. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, F. The “Black Morph Cyphoma” from the Netherlands Antilles (Gastropoda: Ovulidae). Acta Conchyl. 2020, 19, 69–75. [Google Scholar]
- Nowlis, J.P. Mate-and oviposition-influenced host preferences in the coral-feeding snail Cyphoma gibbosum. Ecology 1993, 74, 1959–1969. [Google Scholar] [CrossRef]
- Chiappone, M.; Dienes, H.; Swanson, D.W.; Miller, S.L. Density and Gorgonian host-occupation patterns by Flamingo Tongue snails (Cyphoma gibbosum) in the Florida keys. Caribb. J. Sci. 2003, 39, 116–127. [Google Scholar]
- Lasker, H.R.; Coffroth, M.A.; Fitzgerald, L.M. Foraging patterns of Cyphoma gibbosum on octocorals: The roles of host choice and feeding preference. Biol. Bull. 1988, 174, 254–266. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Koh, E.G.L. Depauperation of the mushroom coral fauna (Fungiidae) of Singapore (1860s–2006) in changing reef conditions. Raffles Bull. Zool. Suppl. 2009, 22, 91–101. [Google Scholar]
- Ruesink, J.L.; Harvell, C.D. Specialist predation on the Caribbean gorgonian Plexaurella spp. by Cyphoma signatum (Gastropoda). Mar. Ecol. Prog. Ser. 1990, 65, 265–272. [Google Scholar] [CrossRef]
- Humann, P.; Deloach, N. Reef Coral Identification: Florida, the Caribbean and the Bahamas, 3rd ed.; New World Publications Inc.: Jacksonville, FL, USA, 2013. [Google Scholar]
- Hoeksema, B.W.; van der Loos, L.M.; van Moorsel, G.W.N.M. Coral diversity matches marine park zonation but not economic value of coral reef sites at St. Eustatius, eastern Caribbean. J. Environ. Manag. 2022, 320, 115829. [Google Scholar] [CrossRef]
- Bayer, F.M. The shallow-water Octocorallia of the West Indian region. Stud. Fauna Curaçao Caribb. Isl. 1961, 12, 506065. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.r-project.org/ (accessed on 11 September 2022).
- Dormann, C.F.; Gruber, B.; Fründ, J. Introducing the bipartite Package: Analysing Ecological Networks. R News 2008, 8, 8–11. [Google Scholar]
- Dormann, C.F.; Fruend, J.; Gruber, B.; Beckett, S.; Devoto, M.; Felix, G.M.F.; Iriondo, J.M.; Opsahl, T.; Pinheiro, R.B.P.; Strauss, R.; et al. Visualising Bipartite Networks and Calculating Some (Ecological) Indices. 2022. Available online: https://github.com/biometry/bipartite (accessed on 11 September 2022).
- Pavlopoulos, G.A.; Kontou, P.I.; Pavlopoulou, A.; Bouyioukos, C.; Markou, E.; Bagos, P.G. Bipartite graphs in systems biology and medicine: A survey of methods and applications. GigaScience 2018, 7, giy014. [Google Scholar] [CrossRef]
- Coker, D.J.; DiBattista, J.D.; Stat, M.; Arrigoni, R.; Reimer, J.; Terraneo, T.I.; Villabos, R.; Nowicki, J.P.; Bunce, M.; Berumen, M.L. DNA metabarcoding confirms primary targets and breadth of diet for coral reef butterflyfishes. Coral Reefs 2022, in press. [Google Scholar] [CrossRef]
- Blüthgen, N.; Menzel, F.; Blüthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 6, 9. [Google Scholar] [CrossRef]
- Brawley, S.H.; Adey, W.H. Coralliophila abbreviata: A significant corallivore! Bull. Mar. Sci. 1982, 32, 595–599. [Google Scholar]
- Frade, P.R.; Bongaerts, P.; Baldwin, C.C.; Trembanis, A.C.; Bak, R.P.M.; Vermeij, M.J.A. Bonaire and Curaçao. In Mesophotic Coral Ecosystems. Coral Reefs of the World; Loya, Y., Puglise, K., Bridge, T., Eds.; Springer: Cham, Switzerland, 2019; Volume 12, pp. 149–162. [Google Scholar] [CrossRef]
- Bongaerts, P.; Ridgway, T.; Sampayo, E.M.; Hoegh-Guldberg, O. Assessing the ‘deep reef refugia’ hypothesis: Focus on Caribbean reefs. Coral Reefs 2010, 29, 309–327. [Google Scholar] [CrossRef]
- Potkamp, G.; Vermeij, M.J.A.; Hoeksema, B.W. Host-dependent variation in density of corallivorous snails (Coralliophila spp.) at Curaçao, southern Caribbean. Mar. Biodivers. 2017, 47, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.A. Distribution, movement and impact of the corallivorous gastropod Coralliophila abbreviata (Lamarck) on a Panamánian patch reef. J. Exp. Mar. Biol. Ecol. 1990, 142, 25–42. [Google Scholar] [CrossRef]
- Baums, I.B.; Miller, M.W.; Szmant, A.M. Ecology of a corallivorous gastropod, Coralliophila abbreviata, on two seleractinian hosts. II. Feeding, respiration and growth. Mar. Biol. 2003, 142, 1093–1101. [Google Scholar] [CrossRef]
- Van der Schoot, R.J.; Hoeksema, B.W. Abundance of coral-associated fauna in relation to depth and eutrophication along the leeward side of Curaçao, southern Caribbean. Mar. Environ. Res. 2022, 181, 105738. [Google Scholar] [CrossRef]
- Bruckner, A.; Bruckner, R. Condition of restored Acropora palmata fragments off Mona Island, Puerto Rico, 2 years after the Fortuna Reefer ship grounding. Coral Reefs 2001, 20, 235–243. [Google Scholar] [CrossRef]
- Williams, D.E.; Miller, M.W. Attributing mortality among drivers of population decline in Acropora palmata in the Florida Keys (USA). Coral Reefs 2012, 31, 369–382. [Google Scholar] [CrossRef]
- Bright, A.J.; Rogers, C.S.; Brandt, M.E.; Muller, E.; Smith, T.B. Disease prevalence and snail predation associated with swell-generated damage on the threatened coral, Acropora palmata (Lamarck). Front. Mar. Sci. 2016, 3, 77. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, V.; Sales, L.; Neves, E.; Johnsson, R. Siderastrea stellata Verrill, 1868 as a new host for the corallivore snail Coralliophila caribaea Abbott, 1958 and some biological aspects of their association. Spixiana 2021, 44, 17–18. [Google Scholar]
- Del Mónaco, C.; Villamiza, E.; Narciso, S. Selectividad de presas de Coralliophila abbreviata y C. caribaea en arrecifes coralinos del Parque Nacional Morrocoy, Venezuela: Una aproximación experimental. Lat. Am. J. Aquat. Res. 2010, 38, 57–70. [Google Scholar] [CrossRef]
- Miller, A.C. Cnidarian prey of the snails Coralliophila abbreviata and C. caribaea (Gastropoda: Muricidae) in Discovery Bay, Jamaica. Bull. Mar. Sci. 1981, 31, 932–934. [Google Scholar]
- Oren, U.; Brickner, I.; Loya, Y. Prudent sessile feeding by the corallivore snail, Coralliophila violacea on coral energy sinks. Proc. R. Soc. B 1998, 265, 2043–2050. [Google Scholar] [CrossRef] [Green Version]
- Clements, C.S.; Hay, M.E. Overlooked coral predators suppress foundation species as reefs degrade. Ecol. Appl. 2018, 28, 1673–1682. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; García-Hernández, J.E. Diversity and distribution of stony corals. In Marine Biodiversity of Bonaire: A Baseline Study, 2nd ed.; Hoeksema, B.W., Ed.; Naturalis Biodiversity Center: Leiden, The Netherlands, 2022; pp. 17–19. [Google Scholar]
- De Jong, K.M.; Coomans, H.E. Marine gastropods from Curaçao, Aruba and Bonaire. Stud. Fauna Curaçao Caribb. Isl. 1988, 69, 1–261. Available online: https://repository.naturalis.nl/pub/506113 (accessed on 11 September 2022).
- Whalen, K.E.; Starczak, V.R.; Nelson, D.R.; Goldstone, J.V.; Hahn, M.E. Cytochrome P450 diversity and induction by gorgonian allelochemicals in the marine gastropod Cyphoma gibbosum. BMC Ecol. 2010, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Williams, E.H.; Bunkley-Williams, L. Marine major ecological disturbances of the Caribbean. Infect. Dis. Rev. 2000, 2, 110–127. [Google Scholar]
- Hector, A.; Hooper, R. Ecology: Darwin and the first ecological experiment. Science 2002, 295, 639–640. [Google Scholar] [CrossRef]
- Shaver, E.C.; Burkepile, D.E.; Silliman, B.R. Local management actions can increase coral resilience to thermally-induced bleaching. Nat. Ecol. Evol. 2018, 2, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- De Bakker, D.M.; van Duyl, F.C.; Perry, C.T.; Meesters, E.H. Extreme spatial heterogeneity in carbonate accretion potential on a Caribbean fringing reef linked to local human disturbance gradients. Glob. Change Biol. 2019, 25, 4092–4104. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Snail Species | ||||
|---|---|---|---|---|
| Host Coral Taxa | Coralliophila galea | Coralliophila salebrosa | Coralliophila sp. | Cyphoma gibbossum |
| Scleractinia | ||||
| Acroporidae | ||||
| Acropora palmata | 6 (1) | |||
| Agariciidae | ||||
| Agaricia agaricites | 14 (10) | |||
| Agaricia humilis | 1 (1) | |||
| Agaricia lamarcki | 2 (2) | |||
| Meandrinidae | ||||
| Eusmilia fastigiata | 1 (1) | |||
| Merulinidae | ||||
| Orbicella annularis | 500 (70) | |||
| Orbicella franksi | 18 (7) | |||
| Montastraeidae | ||||
| Montastraea cavernosa | 1 (1) | |||
| Mussidae | ||||
| Pseudodiploria strigosa | 9 (1) | |||
| Pocilloporidae | ||||
| Madracis auretenra | 24 (8) | 1 (1) | ||
| Octocorallia | ||||
| Gorgoniidae | ||||
| Antillogorgia bipinnata | 12 (7) | |||
| Antillogorgia sp. | 3 (1) | 6 (4) | ||
| Gorgonia ventalina | 3 (2) | 3 (2) | ||
| Plexauridae | ||||
| Eunicea flexuosa | 5 (3) | |||
| Eunicea fusca | 2 (1) | |||
| Eunicea sp. | 4 (3) | 13 (6) | ||
| Plexaura homomalla | 6 (2) | 14 (4) | ||
| Pseudoplexaura sp. | 17 (7) | 24 (13) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verboom, L.; Hoeksema, B.W. Resource Partitioning by Corallivorous Snails on Bonaire (Southern Caribbean). Diversity 2023, 15, 34. https://doi.org/10.3390/d15010034
Verboom L, Hoeksema BW. Resource Partitioning by Corallivorous Snails on Bonaire (Southern Caribbean). Diversity. 2023; 15(1):34. https://doi.org/10.3390/d15010034
Chicago/Turabian StyleVerboom, Lukas, and Bert W. Hoeksema. 2023. "Resource Partitioning by Corallivorous Snails on Bonaire (Southern Caribbean)" Diversity 15, no. 1: 34. https://doi.org/10.3390/d15010034