
Epibionts of an Introduced King Crab in the Barents Sea: A Second Five-Year Study


Abstract
:1. Introduction
2. Material and Methods
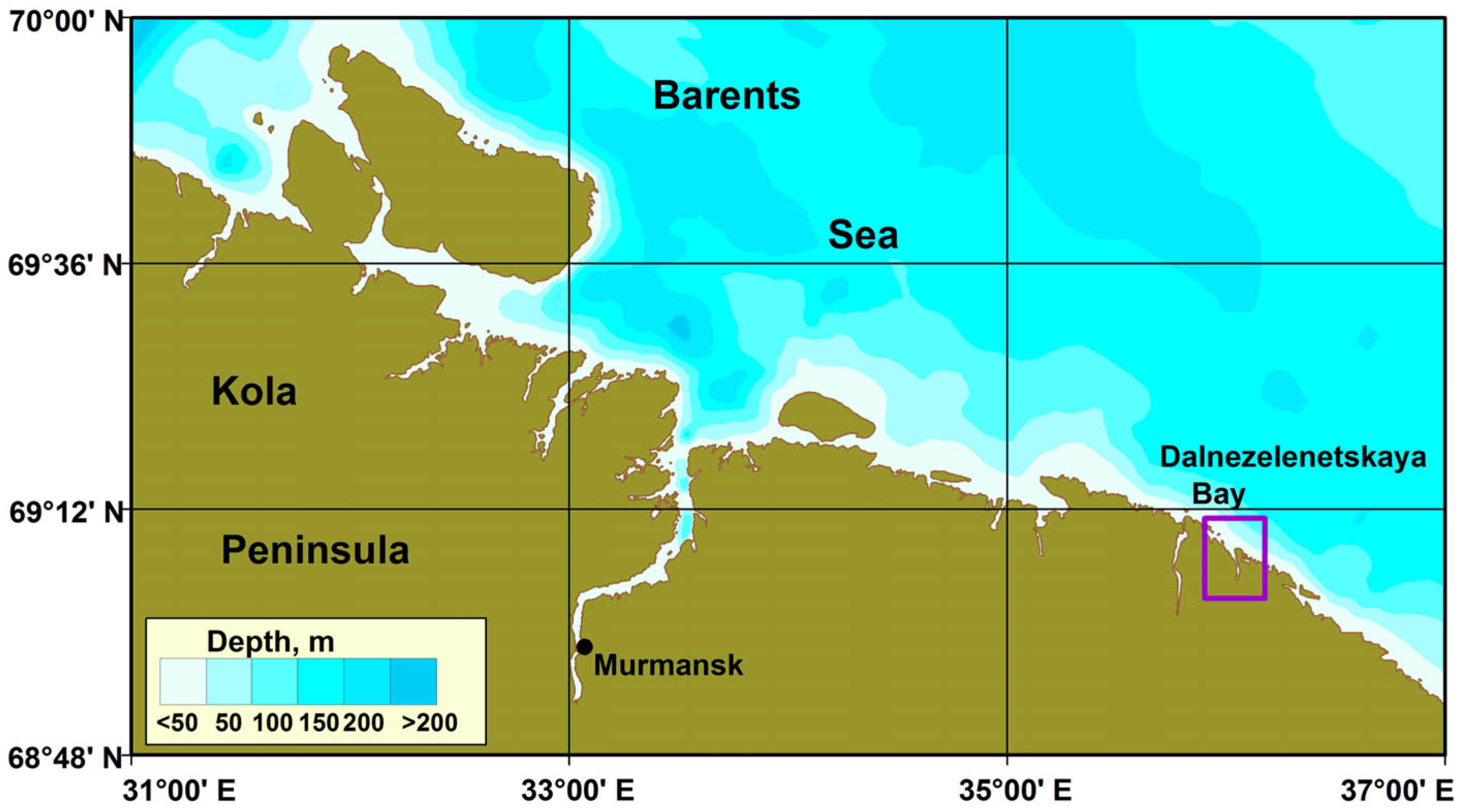
2.1. Study Area
2.2. Sampling and Processing
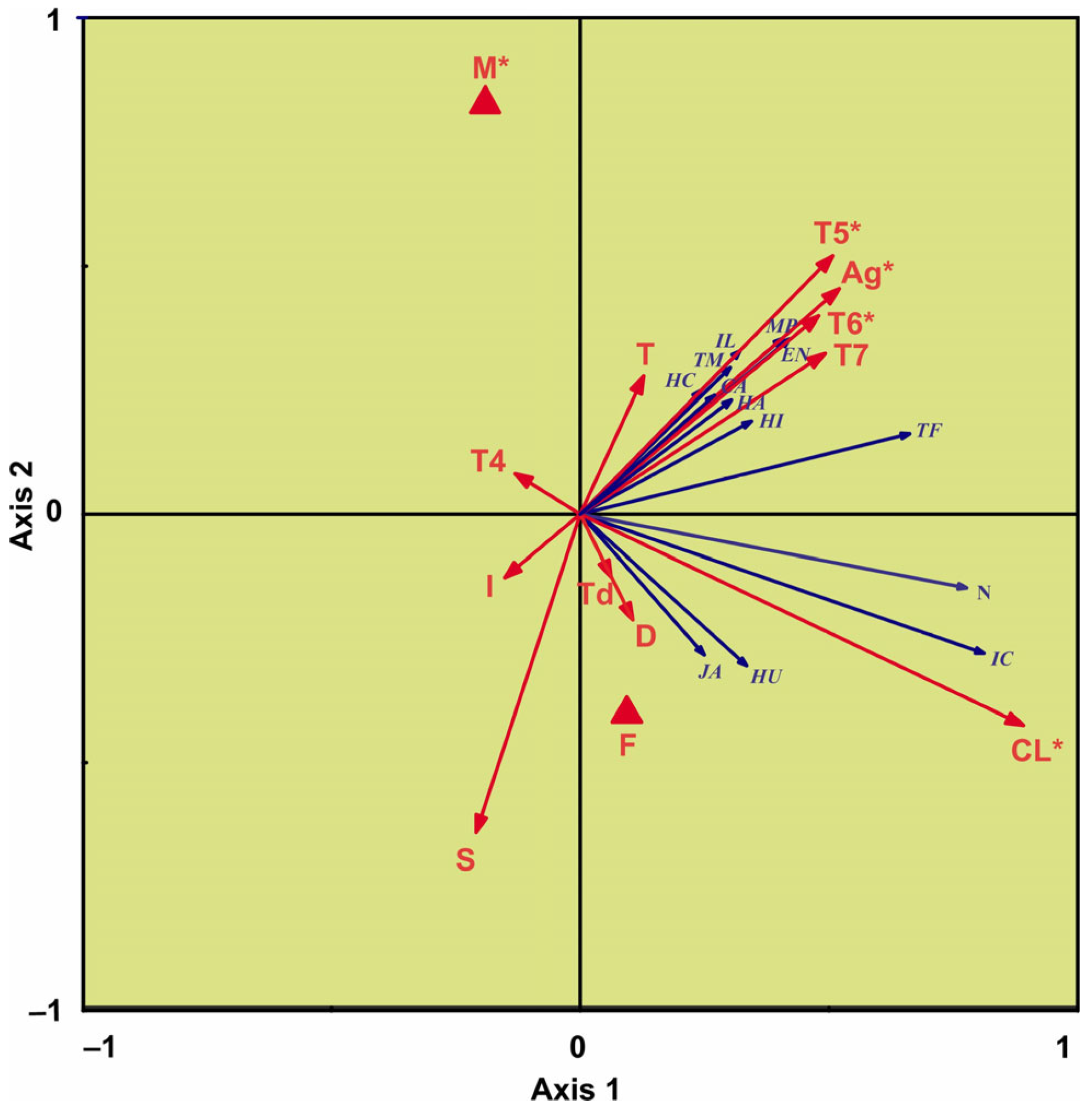
2.3. Statistical Analysis
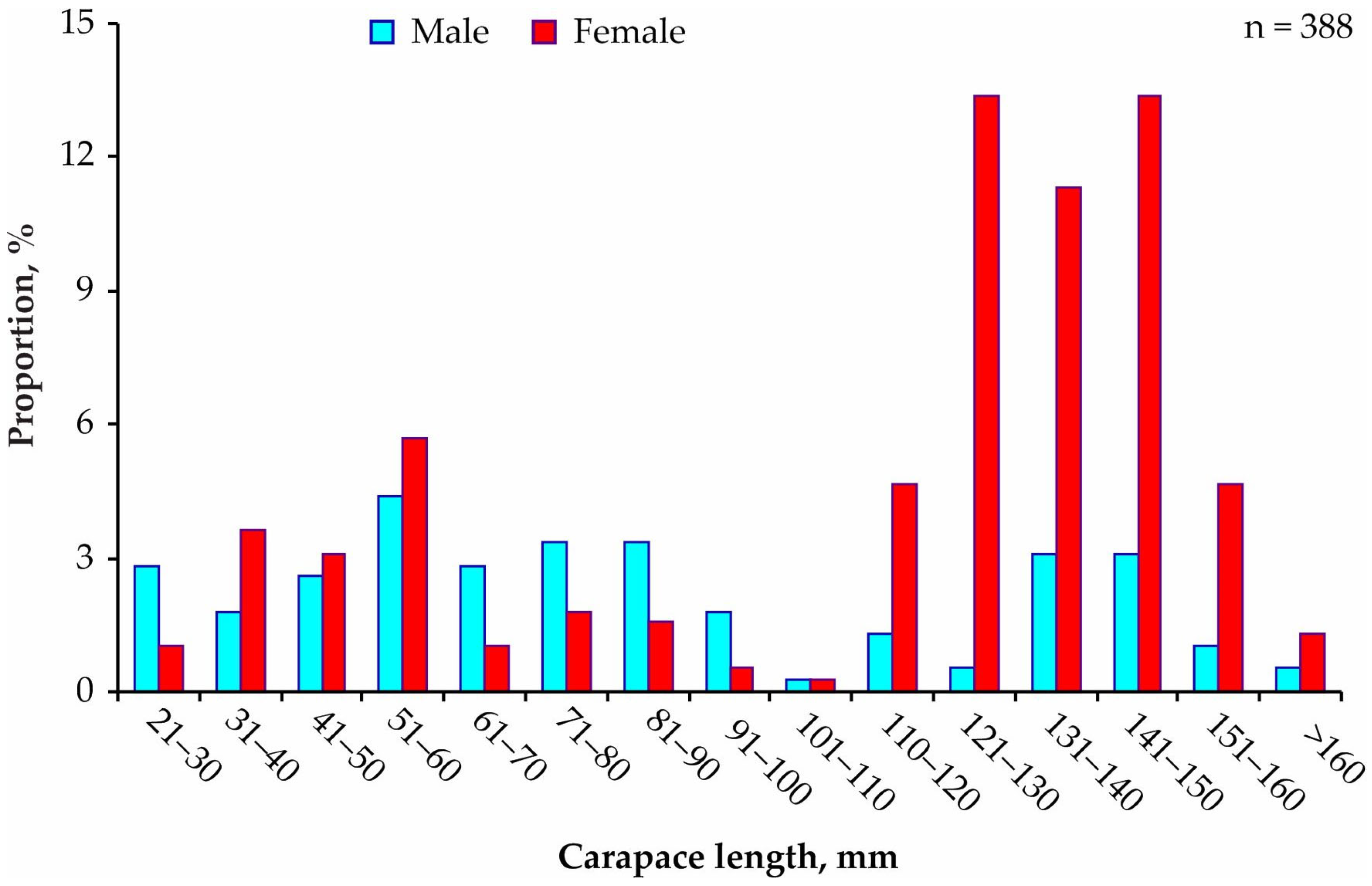
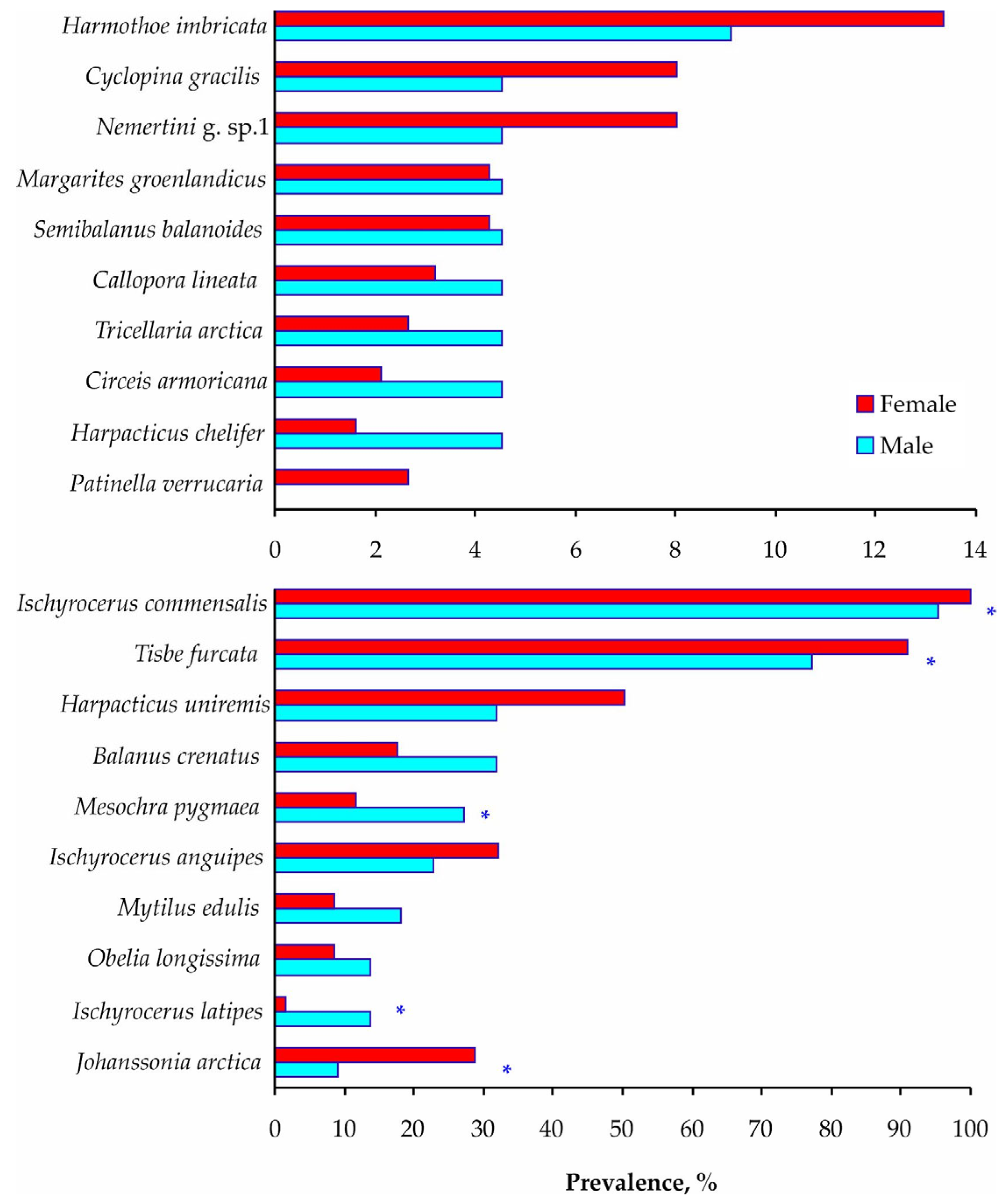
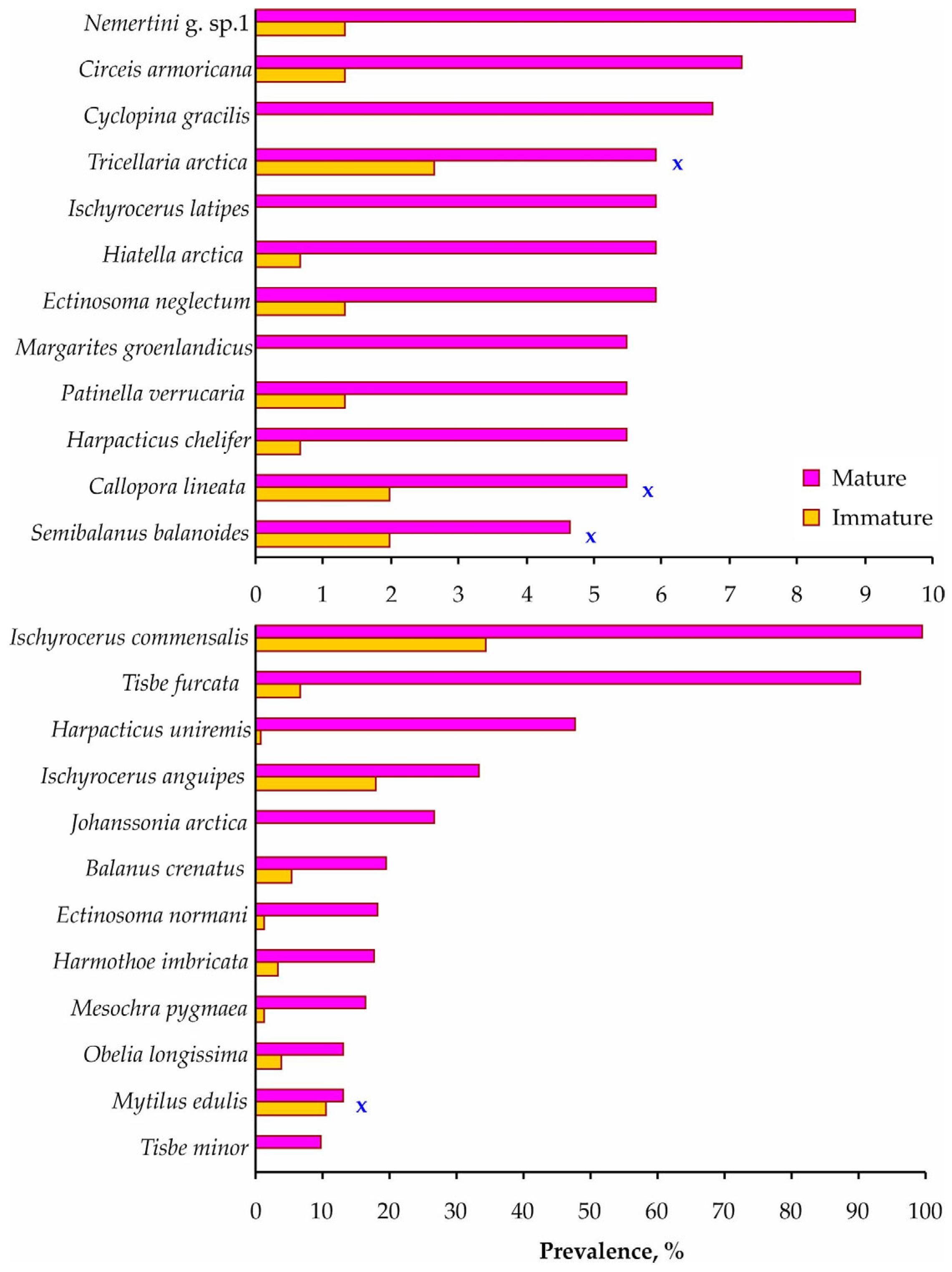
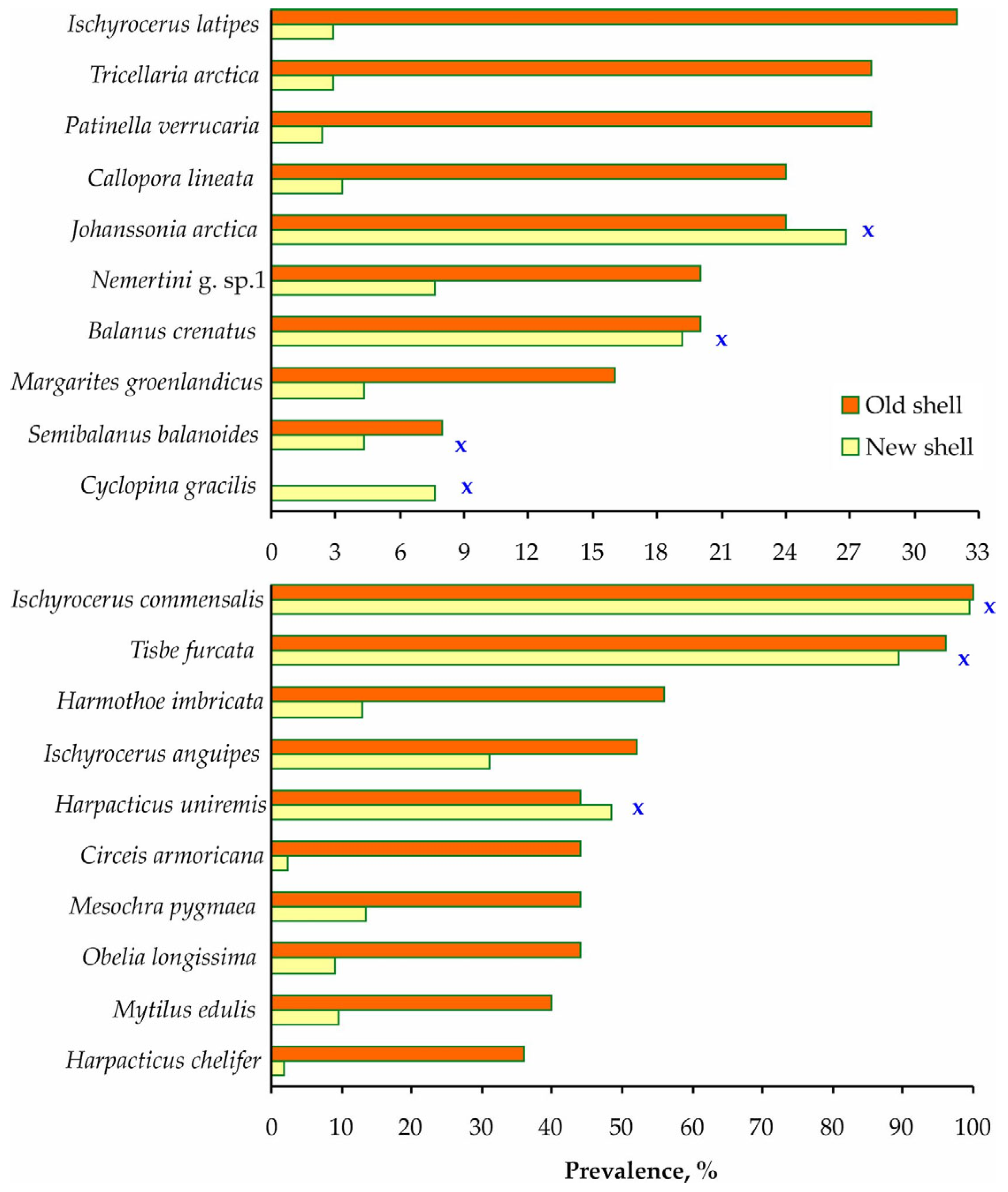
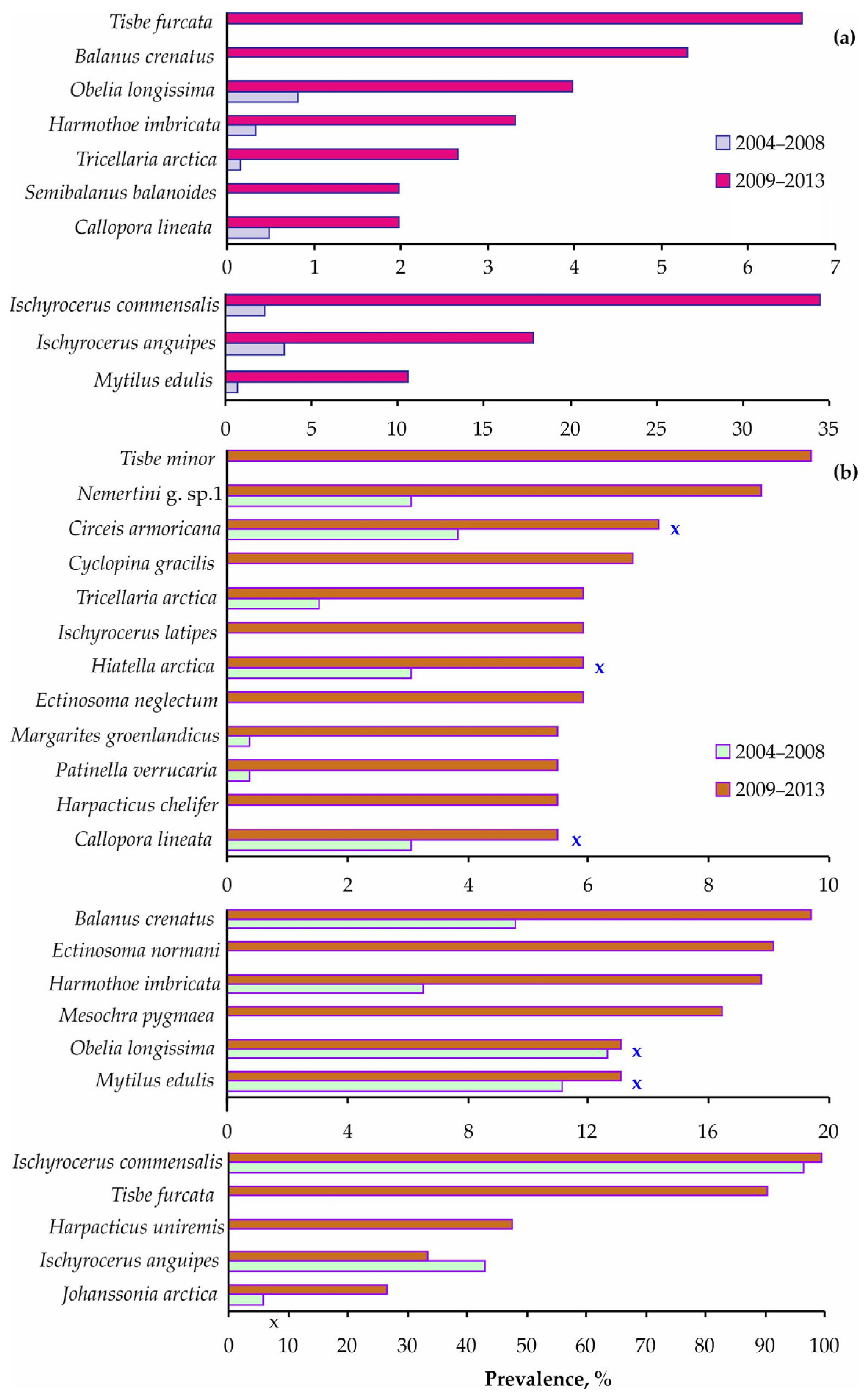
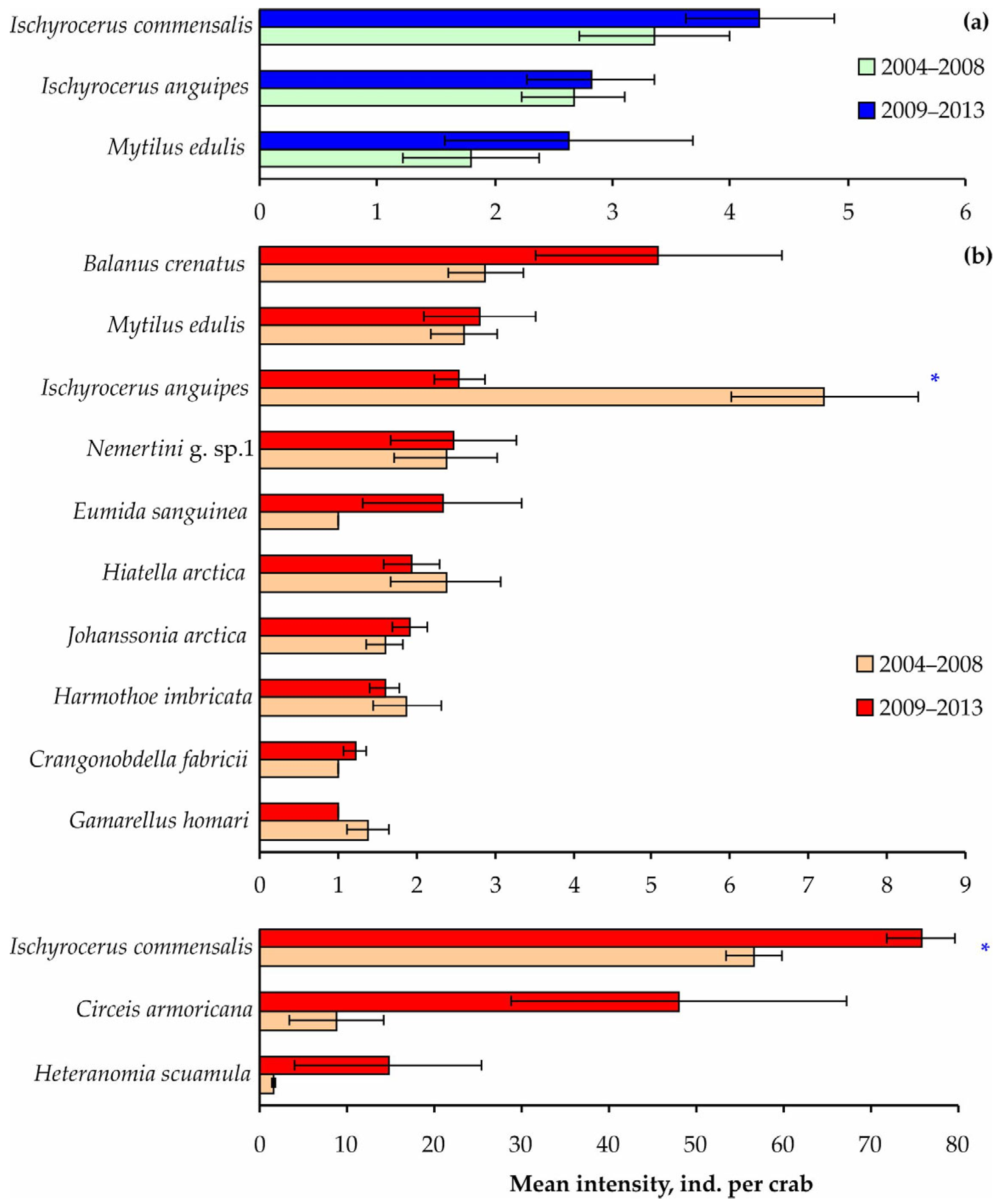
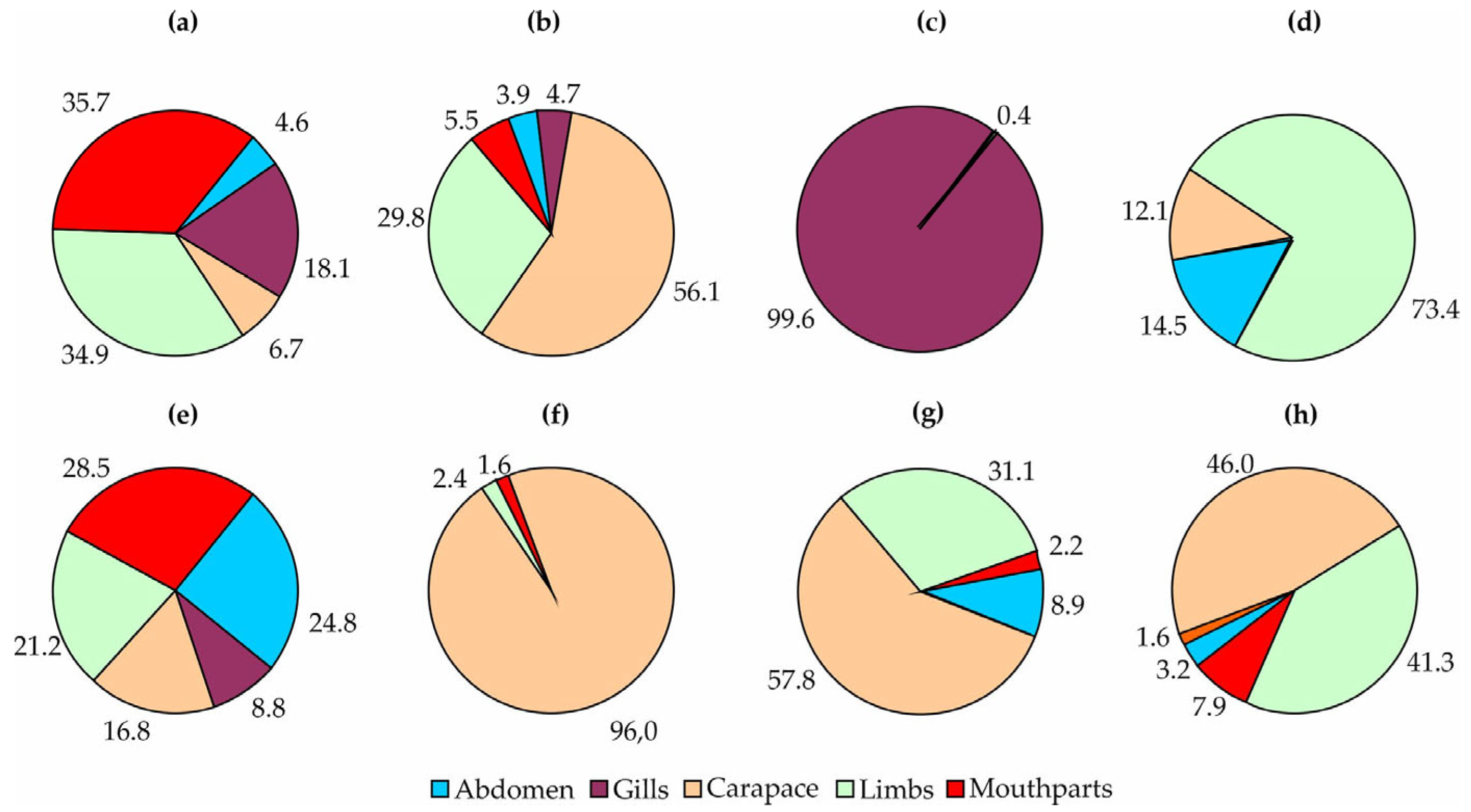
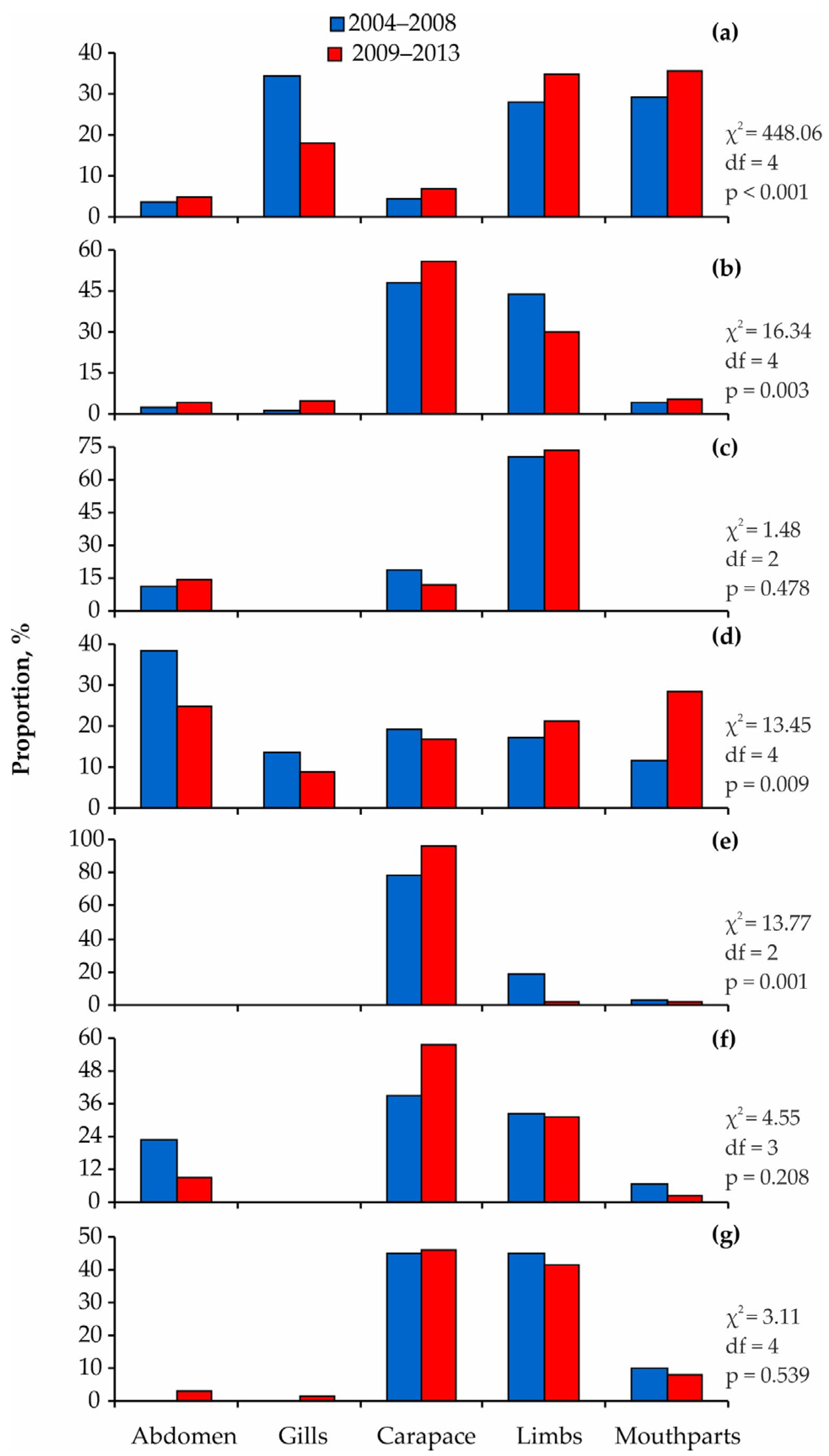
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wassmann, P.; Reigstad, M.; Haug, T.; Rudels, B.; Carroll, M.L.; Hop, H.; Gabrielsen, G.W.; Falk-Petersen, S.; Denisenko, S.G.; Arashkevich, E.; et al. Food webs and carbon flux in the Barents Sea. Prog. Oceanogr. 2006, 71, 232–287. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Copepod communities off Franz Josef Land (northern Barents Sea) in late summer of 2006 and 2007. Polar Biol. 2011, 34, 1231–1238. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Estimated copepod production rate and structure of mesozooplankton communities in the coastal Barents Sea during summer–autumn 2007. Polar Biol. 2012, 35, 1321–1342. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Cucumaria in Russian waters of the Barents Sea: Biological aspects and aquaculture potential. Front. Mar. Sci. 2021, 8, 613453. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Ecology and distribution of red king crab larvae in the Barents Sea: A review. Water 2022, 14, 2328. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Commercial fish and shellfish in the Barents Sea: Have introduced crab species affected the population trajectories of commercial fish? Rev. Fish Biol. Fish. 2015, 25, 297–322. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Red king crab (Paralithodes camtschaticus) fisheries in Russian waters: Historical review and present status. Rev. Fish Biol. Fish. 2018, 28, 331–353. [Google Scholar] [CrossRef]
- Fuglestad, J.L.; Benestad, R.; Ivanov, V.; Jørgensen, L.L.; Kovacs, K.M.; Nilssen, F.; Skjoldal, H.R.; Tchernova, J. Ecosystems of the Barents Sea Region. In Governing Arctic Seas: Regional Lessons from the Bering Strait and Barents Sea; Young, O., Berkman, P., Vylegzhanin, A., Eds.; Informed Decisionmaking for Sustainability; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Dvoretsky, V.G.; Dvoretsky, A.G. Coastal mesozooplankton assemblages during spring bloom in the eastern Barents Sea. Biology 2022, 11, 204. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer-fall macrozooplankton assemblages in a large Arctic estuarine zone (south-eastern Barents Sea): Environmental drivers of spatial distribution. Mar. Environ. Res. 2022, 173, 105498. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acid composition of the Barents Sea red king crab (Paralithodes camtschaticus) leg meat. J. Food Compos. Anal. 2021, 98, 103826. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Tipisova, E.V.; Elfimova, A.E.; Alikina, V.A.; Dvoretsky, V.G. Sex hormones in hemolymph of red king crabs from the Barents Sea. Animals 2021, 11, 2149. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acids in the circulatory system of an invasive king crab from the Barents Sea. J. Food Compos. Anal. 2022, 110, 104528. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Tipisova, E.V.; Alikina, V.A.; Elfimova, A.E.; Dvoretsky, V.G. Thyroid hormones in hemolymph of red king crabs from the Barents Sea. Animals 2022, 12, 379. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Inter-annual dynamics of the Barents Sea red king crab (Paralithodes camtschaticus) stock indices in relation to environmental factors. Polar Sci. 2016, 10, 541–552. [Google Scholar] [CrossRef]
- Bakanev, S.V.; Stesko, A.V. Red king crab. In Materials Justifying Changes to the Previously Approved Total Allowable Catch in Fishing Areas in Inland Seas of the Russian Federation, on the Continental Shelf of the Russian Federation, in the Exclusive Economical Zone of the Russian Federation, in the Azov and Caspian Seas in 2021; FGBUN VNIRO (Polar Branch): Murmansk, Russia, 2021; pp. 2–16. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Renewal of the recreational red king crab fishery in Russian waters of the Barents Sea: Potential benefits and costs. Mar. Policy 2022, 136, 104916. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Chan, F.T.; Stanislawczyk, K.; Sneekes, A.C.; Dvoretsky, A.; Gollasch, S.; Minchin, D.; David, M.; Jelmert, A.; Albretsen, J.; Bailey, S.A. Climate change opens new frontiers for marine species in the Arctic: Current trends and future invasion risks. Glob. Change Biol. 2019, 25, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Alekseev, V.R. Confusing invader: Acanthocyclops americanus (Copepoda: Cyclopoida) and its biological, anthropogenic and climate-dependent mechanisms of rapid distribution in Eurasia. Water 2021, 13, 1423. [Google Scholar] [CrossRef]
- Dimitriadis, C.; Fournari-Konstantinidou, I.; Sourbès, L.; Koutsoubas, D.; Katsanevakis, S. Long term interactions of native and invasive species in a marine protected area suggest complex cascading effects challenging conservation outcomes. Diversity 2021, 13, 71. [Google Scholar] [CrossRef]
- Falk-Petersen, J.; Renaud, P.; Anisimova, N. Establishment and ecosystem effects of the alien invasive red king crab (Paralithodes camtschaticus) in the Barents Sea—A review. ICES J. Mar. Sci. 2011, 68, 479–488. [Google Scholar] [CrossRef]
- Britayev, T.A.; Rzhavsky, A.V.; Pavlova, L.V.; Dvoretskij, A.G. Studies on impact of the alien Red King Crab (Paralithodes camtschaticus) on the shallow water benthic communities of the Barents Sea. J. Appl. Ichthyol. 2010, 26 (Suppl. 2), 66–73. [Google Scholar] [CrossRef]
- Oug, E.; Cochrane, S.K.J.; Sundet, J.H.; Norling, K.; Nilsson, H.C. Effects of the invasive red king crab (Paralithodes camtschaticus) on soft-bottom fauna in Varangerfjorden, northern Norway. Mar. Biodiv. 2011, 41, 467–479. [Google Scholar] [CrossRef]
- Pavlova, L.V.; Dvoretsky, A.G. Prey selectivity in juvenile red king crabs from the coastal Barents Sea. Diversity 2022, 14, 568. [Google Scholar] [CrossRef]
- Zaklan, S.D. Review of the Family Lithodidae (Crustacea: Anomura: Paguroidea): Distribution, biology and fisheries. In Crabs in Cold Water Regions: Biology, Management, and Economics Alaska Sea Grant College Program, AK-SG-02-01; MacIntosh, R.A., Ed.; Anchorage College: Anchorage, AK, USA, 2002; pp. 751–845. [Google Scholar]
- Klitin, A.K. The Red King Crab along the Shores of Sakhalin and the Kuril Islands: Biology, Distribution, and Functional Structure of the Area; Natsrybresursy: Moscow, Russia, 2003. (In Russian) [Google Scholar]
- Bernier, R.Y.; Locke, A.; Hanson, J.M. Lobsters and crabs as potential vectors for tunicate dispersal in the southern Gulf of St. Lawrence, Canada. Aquat. Invas. 2009, 4, 105–110. [Google Scholar] [CrossRef]
- Huys, R. Harpacticoid copepods—Their symbiotic associations and biogenic substrata: A review. Zootaxa 2016, 4174, 448–729. [Google Scholar] [CrossRef]
- Bhaduri, R.N.; Valentich-Scott, P.; Hilgers, M.; Singh, R. New host record for the California mussel Mytilus californianus (Bivalvia, Mytilidae), epibiotic on the pacific sand crab Emerita analoga (Decapoda, Hippidae) from Monterey Bay, California (U.S.A.). Crustaceana 2017, 90, 69–75. [Google Scholar] [CrossRef]
- Frizzera, A.; Bojko, J.; Cremonte, F.; Vázquez, N. Symbionts of invasive and native crabs, in Argentina: The most recently invaded area on the Southwestern Atlantic coastline. J. Invert. Pathol. 2021, 184, 107650. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epifauna associated with an introduced crab in the Barents Sea: A 5-year study. ICES J. Mar. Sci. 2010, 67, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Dvoretsky, A.G.; Dvoretsky, V.G. Copepods associated with the red king crab Paralithodes camtschaticus (Tilesius, 1815) in the Barents Sea. Zool. Stud. 2013, 52, 17. [Google Scholar] [CrossRef] [Green Version]
- Dvoretsky, A.G.; Dvoretsky, V.G. New echinoderm-crab epibiotic associations from the coastal Barents Sea. Animals 2021, 11, 917. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibiotic communities of common crab species in the coastal Barents Sea: Biodiversity and infestation patterns. Diversity 2022, 14, 6. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Effects of environmental factors on the abundance, biomass, and individual weight of juvenile red king crabs in the Barents Sea. Front. Mar. Sci. 2020, 7, 726. [Google Scholar] [CrossRef]
- Evseeva, O.Y.; Ishkulova, T.G.; Dvoretsky, A.G. Environmental drivers of an intertidal bryozoan community in the Barents Sea: A case study. Animals 2022, 12, 552. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Aquaculture of green sea urchin in the Barents Sea: A brief review of Russian studies. Rev. Aquac. 2020, 12, 1280–1290. [Google Scholar] [CrossRef]
- Donaldson, W.E.; Byersdorfer, S.E. Biological Field Techniques for Lithodid Crabs; Alaska Sea Grant College Program: Fairbanks, AK, USA; University of Alaska: Fairbanks, AK, USA, 2005. [Google Scholar]
- Kuzmin, S.A.; Gudimova, E.N. Introduction of the Kamchatka (Red King) Crab in the Barents Sea. Peculiarities of Biology, Perspectives of Fishery; KSC RAS Press: Apatity, Russia, 2002. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size-at-age of juvenile red king crab (Paralithodes camtschaticus) in the coastal Barents Sea. Cah. Biol. Mar. 2014, 55, 43–48. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size at maturity of female red king crab, Paralithodes camtschaticus, from the costal zone of Kola Peninsula (southern Barents Sea). Cah. Biol. Mar. 2015, 56, 49–54. [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Hemolymph molting hormone concentrations in red king crabs from the Barents Sea. Polar Biol. 2010, 33, 1293–1298. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Does spine removal affect molting process in the king red crab (Paralithodes camtschaticus) in the Barents Sea? Aquaculture 2012, 326–329, 173–177. [Google Scholar] [CrossRef]
- Lepš, J.; Smilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Anonymous. Kola Meridian Section. 2021. Available online: http://www.pinro.ru/labs/hid/kolsec22.php (accessed on 25 October 2021).
- Dvoretsky, V.G.; Dvoretsky, A.G. Mesozooplankton in the Kola Transect (Barents Sea): Autumn and winter structure. J. Sea Res. 2018, 142, 125–131. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer macrozooplankton assemblages of Arctic shelf: A latitudinal study. Cont. Shelf Res. 2019, 188, 103967. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest of southern Wisconsin. Ecol. Monogr. 1957, 27, 225–349. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Population dynamics of the invasive lithodid crab, Paralithodes camtschaticus, in a typical bay of the Barents Sea. ICES J. Mar. Sci. 2013, 70, 1255–1262. [Google Scholar] [CrossRef]
- Vader, W.; Tandberg, A.H.S. Amphipods as associates of other Crustacea: A survey. J. Crust. Biol. 2015, 35, 522–532. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.; Britayev, T.A. Symbiotic polychaetes revisited: An update of the known species and relationships (1998–2017). Oceanogr. Mar. Biol. Ann. Rev. 2018, 56, 371–448. [Google Scholar]
- Scheuring, L. Die Hydroiden. Untersuchungsfahrt des Reichsforschungsdampfer “Poseidon” in das Barentsmeer im Juni and Juli 1913. Wiss. Meeresuntersuch. Helgolandn. Ser. 1922, 13, 157–179. [Google Scholar]
- Dvoretsky, A.G. Epibionts of the great spider crab, Hyas araneus (Linnaeus, 1758), in the Barents Sea. Polar Biol. 2012, 35, 625–631. [Google Scholar] [CrossRef]
- Panteleeva, N.N. Hydroids (Cnidaria, Hydroidea) in the overgrowth of the red king crab from the coastal zone of the Barents Sea. In Proceedings of the International Symposium “Role of Climate and Fishery in the Structure Change of Shelf Zoobenthos”, Murmansk, Russia, 19–23 March 2003; Matishov, G.G., Ed.; MMBI KSC RAS: Murmansk, Russia, 2003; pp. 69–70. (In Russian). [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epifauna associated with the northern stone crab Lithodes maia in the Barents Sea. Polar Biol. 2008, 31, 1149–1152. [Google Scholar] [CrossRef]
- Steele, D.H.; Hooper, R.G.; Keats, D. Two corophioid amphipods commensal on spider crabs in Newfoundland. J. Crust. Biol. 1986, 6, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Dvoretsky, A.G.; Dvoretsky, V.G. Some aspects of the biology of the amphipods Ischyrocerus anguipes associated with the red king crab, Paralithodes camtschaticus, in the Barents Sea. Polar Biol. 2009, 32, 463–469. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Distribution of amphipods Ischyrocerus on the red king crab, Paralithodes camtschaticus: Possible interactions with the host in the Barents Sea. Estuar. Coast. Shelf Sci. 2009, 82, 390–396. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Fouling community of the red king crab, Paralithodes camtschaticus (Tilesius 1815), in a subarctic fjord of the Barents Sea. Polar Biol. 2009, 32, 1047–1054. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Population biology of Ischyrocerus commensalis, a crab-associated amphipod, in the southern Barents Sea: A multi-annual summer study. Mar. Ecol. 2011, 32, 498–508. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Checklist of fauna found in zooplankton samples from the Barents Sea. Polar. Biol. 2010, 33, 991–1005. [Google Scholar] [CrossRef]
- Jansen, P.A.; Mackenzie, K.; Hemmingsen, W. Some parasites and commensals of red king crab Paralithodes camtschaticus in the Barents Sea. Bull. Europ. Assoc. Fish Pathol. 1998, 18, 46–49. [Google Scholar]
- Hemmingsen, W.; Jansen, P.A.; MacKenzie, K. Crabs, leeches and trypanosomes: An unholy trinity? Mar. Pollut. Bull. 2005, 50, 336–339. [Google Scholar] [CrossRef]
- Harder, T. Marine epibiosis: Concepts, ecological consequences and host defence. In Marine and Industrial Biofouling; Flemming, H.C., Sriyutha Murthy, P., Venkatesan, R., Cooksey, K., Eds.; Springer: Berlin, Germany, 2009; pp. 219–231. [Google Scholar]
- Polyakov, I.V.; Alkire, M.B.; Bluhm, B.A.; Brown, K.A.; Carmack, E.C.; Chierici, M.; Danielson, S.L.; Ellingsen, I.; Ershova, E.A.; Gårdfeldt, K.; et al. Borealization of the Arctic Ocean in response to anomalous advection from sub-arctic seas. Front. Mar. Sci. 2020, 7, 491. [Google Scholar] [CrossRef]
- Bell, C.; McQuaid, C.D.; Porri, F. Barnacle settlement on rocky shores: Substratum preference and epibiosis on mussels. J. Exp. Mar. Biol. Ecol. 2015, 473, 195–201. [Google Scholar] [CrossRef]
- Yuvaraj, D.; Annushrie, A.; Niranjana, M.; Gnanasekaran, R.; Gopinath, M.; Iyyappan, J. A review on process and characterization of mussels and cirripeds for adhesive properties and applications thereof. Cur. Res. Green Sustain. Chem. 2021, 4, 100092. [Google Scholar] [CrossRef]
- Williams, J.D.; McDermott, J.J. Hermit crab biocoenoses; a worldwide review of the diversity and natural history of hermit crab associates. J. Exp. Mar. Biol. Ecol. 2004, 305, 1–128. [Google Scholar] [CrossRef]
- McGaw, I.J. Epibionts of sympatric species of Cancer crabs in Barkley sound, British Columbia. J. Crust. Biol. 2006, 26, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Di Camillo, C.; Bo, M.; Puce, S.; Tazioli, S.; Froglia, C.; Bavestrello, G. The epibiontic assemblage of Geryon longipes (Crustacea: Decapoda: Geryonidae) from the Southern Adriatic Sea. Ital. J. Zool. 2008, 75, 29–35. [Google Scholar] [CrossRef]
- Ibrahim, N.K. Epibiont communities of the two spider crabs Schizophrys aspera (H. Milne Edwards, 1834) and Hyastenus hilgendorfi (De Man, 1887) in Great Bitter Lakes, Suez Canal, Egypt. Egypt. J. Aquat. Biol. Fish. 2012, 16, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Dvoretsky, A.G.; Dvoretsky, V.G. Interspecific relationships of symbiotic amphipods on the red king crab in the Barents Sea. Dokl. Biol. Sci. 2010, 433, 279–281. [Google Scholar] [CrossRef]
- Kuris, A.M.; Blau, S.F.; Paul, A.J.; Shields, J.D.; Wickham, D.E. Infestation by brood symbionts and their impact on egg mortality of the red king crab, (Paralithodes camtschatica) in Alaska. Can. J. Fish. Aquat. Sci. 1991, 48, 559–568. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. The amphipod Ischyrocerus commensalis on the eggs of the red king crab Paralithodes camtschaticus: Egg predator or scavenger? Aquaculture 2010, 298, 185–189. [Google Scholar] [CrossRef]
- Hendrickx, M.E.; Ramírez-Félix, E. Settlement of the barnacle Balanus trigonus Darwin, 1854, on Panulirus gracilis Streets, 1871, in western Mexico. Nauplius 2019, 27, e2019020. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Biological aspects, fisheries, and aquaculture of Yesso scallops in Russian waters of the Sea of Japan. Diversity 2022, 14, 399. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Intraspecific relationships of the red king crab symbionts in the Barents Sea. Dokl. Biol. Sci. 2008, 422, 333–335. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Interspecific competition of symbiotic and fouling species of red king crab in the Barents Sea. Dokl. Biol. Sci. 2011, 440, 300–302. [Google Scholar] [CrossRef]
- Puccinelli, E.; McQuaid, C.D. Commensalism, antagonism or mutualism? Effects of epibiosis on the trophic relationships of mussels and epibiotic barnacles. J. Exp. Mar. Biol. Ecol. 2021, 540, 151549. [Google Scholar]
- Khan, R.A. Biology of marine piscicolid leech Johanssonia arctica (Johansson) from Newfoundland. Proc. Helmintol. Soc. Wash. 1982, 48, 266–278. [Google Scholar]
- Utevsky, S.Y.; Trontelj, P. Phylogenetic relationships of fish leeches (Hirudinea, Piscicolidae) based on mitochondrial DNA sequences and morphological data. Zool. Scr. 2004, 33, 375–385. [Google Scholar] [CrossRef]
- Savoie, L.; Miron, J.; Biron, M. Fouling community of the snow crab Chionoecetes opilio in Atlantic Canada. J. Crust. Biol. 2007, 27, 30–36. [Google Scholar] [CrossRef]
- Savoie, L.; Miron, J.; Biron, M. Fouling community of the snow crab Chionoecetes opilio in Sydney Bight, Canada: Preliminary observations in relation to sampling period and depth/geographical location. Cah. Biol. Mar. 2007, 48, 347–359. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Immature Crabs (CL < 90 mm) | Mature Crabs (CL > 90 mm) | PR Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| PR | IN | PR | IN | ||||||||
| Min | Max | X | SE | Min | Max | X | SE | ||||
| Algae | |||||||||||
| Alaria esculenta (Linnaeus) Greville, 1830 | – | – | – | – | – | 1.3 | 1 | 1 | 1 | 0 | 0.8 |
| Odonthalia dentata (Linnaeus) Lyngbye, 1819 | – | – | – | – | – | 0.8 | 1 | 1 | 1 | 0 | 0.5 |
| Palmaria palmata (Linnaeus) F.Weber & D.Mohr, 1805 | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Polysiphonia sp. | – | – | – | – | – | 0.8 | 1 | 1 | 1 | 0 | 0.5 |
| Saccharina latissima (Linnaeus) C.E.Lane, C.Mayes, Druehl & G.W.Saunders, 2006 | – | – | – | – | – | 2.1 | 1 | 1 | 1 | 0 | 1.3 |
| Hydrozoa | |||||||||||
| Campanularia groenlandica Levinsen, 1893 | – | – | – | – | – | 2.1 | 1 | 1 | 1 | 0 | 1.3 |
| Coryne hincksi Bonnevie, 1898 | 0.7 | – | – | – | – | 2.1 | 1 | 1 | 1 | 0 | 1.5 |
| Cuspidella sp. | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Gonothyraea loveni (Allman, 1859) | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Halecium beanii (Johnston, 1838) | 0.7 | 1 | 1 | 1 | 0 | 2.1 | 1 | 1 | 1 | 0 | 1.5 |
| Obelia geniculata (Linnaeus, 1758) | 0.7 | 1 | 1 | 1 | 0 | 1.7 | 1 | 1 | 1 | 0 | 1.3 |
| Obelia longissima (Pallas, 1766) | 4.0 | 1 | 1 | 1 | 0 | 13.1 | 1 | 1 | 1 | 0 | 9.5 |
| Symplectoscyphus tricuspidatus (Alder, 1856) | 0.7 | 1 | 1 | 1 | 0 | 1.3 | 1 | 1 | 1 | 0 | 1.0 |
| Nemertea | |||||||||||
| Nemertini g. sp.1 | 1.3 | 8 | 44 | 26 | 18 | 8.9 | 1 | 14 | 2.5 | 0.8 | 5.9 |
| Nemertini g. sp.2 | – | – | – | – | – | 2.1 | 1 | 6 | 3.4 | 1.1 | 1.3 |
| Polychaeta | |||||||||||
| Branchiomma infarctum (Krøyer, 1856) | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Bushiella (Jugaria) similis (Bush, 1905) | 0.7 | 1 | 1 | 1 | 0 | 0.8 | 1 | 1 | 1 | 0 | 0.8 |
| Chone infundibuliformis Krøyer, 1856 | – | – | – | – | – | 0.4 | 2 | 2 | 2 | 0 | 0.3 |
| Chone sp. | – | – | – | – | – | 1.3 | 1 | 2 | 1.3 | 0.3 | 0.8 |
| Circeis armoricana Saint-Joseph, 1894 | 1.3 | 1 | 1 | 1 | 0 | 7.2 | 1 | 295 | 47.9 | 19.2 | 4.9 |
| Eumida sanguinea (Örsted, 1843) | – | – | – | – | – | 3.8 | 1 | 10 | 2.3 | 1.0 | 2.3 |
| Harmothoe imbricata (Linnaeus, 1767) | 3.3 | 1 | 1 | 1 | 0 | 17.7 | 1 | 6 | 1.6 | 0.2 | 12.1 |
| Harmothoe impar impar (Johnston, 1839) sensu Malmgren, 1865 | – | – | – | – | – | 2.5 | 1 | 2 | 1.5 | 0.2 | 1.5 |
| Lepidonotus squamatus (Linnaeus, 1758) | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Polydora ciliata (Johnston, 1838) | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Syllidae g. sp. | – | – | – | – | – | 2.1 | 1 | 1 | 1 | 0 | 1.3 |
| Syllis armillaris (O.F. Müller, 1776) | 0.7 | 1 | 1 | 1 | 0 | 0.4 | 1 | 1 | 1 | 0 | 0.5 |
| Hirudinea | |||||||||||
| Johanssonia arctica (Johansson, 1898) | – | – | – | – | – | 26.6 | 1 | 12 | 1.9 | 0.2 | 16.2 |
| Platybdella fabricii (Malm, 1863) | 0.7 | 1 | 1 | 1 | 0 | 3.8 | 1 | 2 | 1.2 | 0.1 | 2.6 |
| Platybdella olriki (Malm, 1865) | – | – | – | – | – | 1.7 | 1 | 1 | 1 | 0 | 1.0 |
| Polyplacophora | |||||||||||
| Tonicella marmorea (O. Fabricius, 1780) | – | – | – | – | – | 2.1 | 1 | 1 | 1 | 0 | 1.3 |
| Gastropoda | |||||||||||
| Margarites groenlandicus (Gmelin, 1791) | – | – | – | – | – | 5.5 | 1 | 3 | 1.3 | 0.2 | 3.4 |
| Margarites helicinus (Phipps, 1774) | 1.3 | 1 | 1 | 1 | 0 | 2.5 | 1 | 2 | 1.2 | 0.2 | 2.1 |
| Rissoa parva (da Costa, 1778) | – | – | – | – | – | 3.8 | 1 | 2 | 1.1 | 0.1 | 2.3 |
| Bivalvia | |||||||||||
| Chlamys islandica (O. F. Müller, 1776) | – | – | – | – | – | 0.8 | 1 | 1 | 1 | 0 | 0.5 |
| Heteranomia squamula (Linnaeus, 1758) | – | – | – | – | – | 1.7 | 1 | 46 | 14.8 | 10.7 | 1.0 |
| Hiatella arctica (Linnaeus, 1767) | 0.7 | 1 | 1 | 1 | 0 | 5.9 | 1 | 5 | 1.9 | 0.4 | 3.9 |
| Musculus discors (Linnaeus, 1767) | 0.7 | 1 | 1 | 1 | 0 | 1.7 | 1 | 3 | 1.5 | 0.5 | 1.3 |
| Mytilus edulis Linnaeus, 1758 | 10.6 | 1 | 18 | 2.6 | 1.1 | 13.1 | 1 | 18 | 2.8 | 0.7 | 12.1 |
| Copepoda | |||||||||||
| Ameira tenuicornis Scott T., 1902 | – | – | – | – | – | 0.8 | 1 | 2 | 1.5 | 0.5 | 0.5 |
| Calanus finmarchicus (Gunnerus, 1770) | – | – | – | – | – | 1.7 | 1 | 1 | 1 | 0 | 1.0 |
| Cyclopina gracilis Claus, 1863 | – | – | – | – | – | 6.8 | 1 | 9 | 2.4 | 0.5 | 4.1 |
| Dactylopusia tisboides (Claus, 1863) | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Dactylopusia vulgaris Sars G.O., 1905 | – | – | – | – | – | 1.3 | 1 | 6 | 3.0 | 1.5 | 0.8 |
| Ectinosoma mixtum Sars G.O., 1904 | – | – | – | – | – | 0.8 | 1 | 4 | 2.5 | 1.5 | 0.5 |
| Ectinosoma normani Scott T. & Scott A., 1896 | 1.3 | 1 | 3 | 2 | 1 | 18.1 | 1 | 36 | 6.8 | 1.2 | 11.6 |
| Halectinosoma elongatum (Sars G.O., 1904) | – | – | – | – | – | 0.8 | 1 | 2 | 1.5 | 0.5 | 0.5 |
| Halectinosoma gracile (Scott T. & Scott A., 1896) | – | – | – | – | – | 1.3 | 1 | 5 | 3.3 | 1.2 | 0.8 |
| Halectinosoma neglectum (Sars G.O., 1904) | 1.3 | 1 | 144 | 72.5 | 71.5 | 5.9 | 1 | 12 | 3.4 | 0.9 | 4.1 |
| Halectinosoma sarsii (Boeck, 1872) | 0.7 | 1 | 1 | 1 | 0 | 0.4 | 1 | 1 | 1 | 0 | 0.5 |
| Harpacticus chelifer (Müller O.F., 1776) | – | – | – | – | – | 5.5 | 1 | 615 | 61.1 | 46.4 | 3.4 |
| Harpacticus flexus Brady & Robertson, 1873 | – | – | – | – | – | 2.5 | 1 | 9 | 3.2 | 1.3 | 1.5 |
| Harpacticus littoralis Sars G.O., 1910 | – | – | – | – | – | 2.1 | 1 | 4 | 1.8 | 0.6 | 1.3 |
| Harpacticus uniremis uniremis Krøyer in Gaimard, 1842–1845? | 0.7 | 3 | 3 | 3 | 0 | 47.7 | 1 | 90 | 6.7 | 1.0 | 29.4 |
| Mesochra pygmaea (Claus, 1863) | 1.3 | 1 | 3 | 2 | 1 | 16.5 | 1 | 29 | 7.4 | 1.2 | 10.6 |
| Microcalanus pusillus Sars G.O., 1903 | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Microsetella norvegica (Boeck, 1865) | – | – | – | – | – | 0.8 | 1 | 1 | 1 | 0 | 0.5 |
| Oithona similis Claus, 1866 | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Tisbe furcata (Baird, 1837) | 6.6 | 1 | 31 | 7.8 | 3.7 | 90.3 | 1 | 1607 | 106.3 | 14.7 | 57.7 |
| Tisbe minor (Scott T. & Scott A., 1896) | – | – | – | – | – | 9.7 | 1 | 13 | 3.0 | 0.6 | 5.9 |
| Tisbe tenera (Sars G.O., 1905) | – | – | – | – | – | 1.3 | 1 | 2 | 1.3 | 0.3 | 0.8 |
| Zaus abbreviatus Sars G.O., 1904 | – | – | – | – | – | 1.7 | 1 | 5 | 2.5 | 1.0 | 1.0 |
| Isopoda | |||||||||||
| Jaera albifrons Leach, 1814 | 0.7 | 1 | 1 | 1 | 0 | 0.4 | 1 | 1 | 1 | 0 | 0.5 |
| Amphipoda | |||||||||||
| Ampelisca sp. | 0.7 | 1 | 1 | 1 | 0 | 0.8 | 1 | 13 | 7 | 6 | 0.8 |
| Caprella septentrionalis Krøyer, 1838 | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Gammarellus homari (J.C. Fabricius, 1779) | – | – | – | – | – | 3.0 | 1 | 1 | 1 | 0 | 1.8 |
| Ischyrocerus anguipes Krøyer, 1838 | 17.9 | 1 | 14 | 2.8 | 0.5 | 33.3 | 1 | 15 | 2.5 | 0.3 | 27.3 |
| Ischyrocerus commensalis Chevreux, 1900 | 34.4 | 1 | 22 | 4.3 | 0.6 | 99.6 | 1 | 361 | 75.7 | 3.9 | 74.2 |
| Ischyrocerus latipes Krøyer, 1842 | – | – | – | – | – | 5.9 | 1 | 14 | 2.7 | 0.9 | 3.6 |
| Ischyrocerus megacheir (Boeck, 1871) | 0.7 | 1 | 1 | 1 | 0 | – | – | – | – | – | 0.3 |
| Cirripedia | |||||||||||
| Balanus balanus (Linnaeus, 1758) | – | – | – | – | – | 0.8 | 1 | 1 | 1 | 0 | 0.5 |
| Balanus crenatus Bruguière, 1789 | 5.3 | 1 | 7 | 2.0 | 0.7 | 19.4 | 1 | 69 | 5.1 | 1.6 | 13.9 |
| Semibalanus balanoides (Linnaeus, 1767) | 2.0 | 1 | 3 | 1.7 | 0.7 | 4.6 | 1 | 6 | 1.8 | 0.5 | 3.6 |
| Verruca stroemia (O.F. Müller, 1776) | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Bryozoa | |||||||||||
| Callopora lineata (Linnaeus, 1767) | 2.0 | 1 | 1 | 1 | 0 | 5.5 | 1 | 1 | 1 | 0 | 4.1 |
| Crisia denticulata (Lamarck, 1816) | – | – | – | – | – | 2.5 | 1 | 1 | 1 | 0 | 1.5 |
| Crisularia harmsworthi (Waters, 1900) | – | – | – | – | – | 1.7 | 1 | 1 | 1 | 0 | 1.0 |
| Disporella hispida (Fleming, 1828) | 1.3 | 1 | 1 | 1 | 0 | 1.7 | 1 | 1 | 1 | 0 | 1.5 |
| Oncousoecia diastoporides (Norman, 1869) | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Patinella verrucaria (Linnaeus, 1758) | 0.7 | 1 | 1 | 1 | 0 | 5.5 | 1 | 1 | 1 | 0 | 3.6 |
| Terminoflustra membranaceotruncata (Smitt, 1868) | 0.7 | 1 | 1 | 1 | 0 | 1.3 | 1 | 1 | 1 | 0 | 1.0 |
| Tricellaria arctica Busk, 1855 | 2.6 | 1 | 1 | 1 | 0 | 5.9 | 1 | 1 | 1 | 0 | 4.6 |
| Echinodermata | |||||||||||
| Asterias rubens Linnaeus, 1758 | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Ophiura robusta (Ayres, 1852) | – | – | – | – | – | 1.3 | 1 | 1 | 1 | 0 | 0.8 |
| Strongylocentrotus droebachiensis (O.F. Muller, 1776) | 0.7 | 1 | 1 | 1 | 0 | – | – | – | – | – | 0.3 |
| Acari | |||||||||||
| Acari g.sp. | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Thalassarachna basteri (Johnston, 1836) | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Pantopoda | |||||||||||
| Nyphon sp. | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Sipuncula | |||||||||||
| Phascolosoma sp. | – | – | – | – | – | 1.3 | 1 | 1 | 1 | 0 | 0.8 |
| Priapulida | |||||||||||
| Halicryptus spinulosus von Siebold, 1849 | – | – | – | – | – | 0.4 | 1 | 1 | 1 | 0 | 0.3 |
| Variable | Explained Variation, % | F-Value | p |
|---|---|---|---|
| Carapace length | 36 | 219.68 | <0.001 |
| Shell condition | 7 | 45.82 | <0.001 |
| Temperature in May | 4 | 31.47 | <0.001 |
| Temperature in June | 2 | 8.51 | 0.001 |
| Sex | 1 | 4.29 | 0.013 |
| Number of injured legs | 0 | 1.53 | 0.189 |
| Depth | 0 | 1.11 | 0.300 |
| Annual temperature | 0.01 | 0.97 | 0.358 |
| Annual salinity | 0 | 2.93 | 0.051 |
| Temperature on the day of sampling | 0 | 1.28 | 0.257 |
| Temperature in April | 0 | 0.09 | 0.908 |
| Index | Immature Crabs | Mature Crabs | Males | Females | ||||
|---|---|---|---|---|---|---|---|---|
| 2004–2008 | 2009–2013 | 2004–2008 | 2009–2013 | 2004–2008 | 2009–2013 | 2004–2008 | 2009–2013 | |
| Species richness | 12 | 34 | 43 | 88 | 34 | 77 | 35 | 75 |
| Shannon–Weaver index | 2.802 | 3.702 | 3.288 | 4.706 | 3.425 | 5.174 | 3.175 | 4.243 |
| Pielou evenness | 0.780 | 0.720 | 0.606 | 0.723 | 0.6732 | 0.8256 | 0.6191 | 0.6812 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dvoretsky, A.G.; Dvoretsky, V.G. Epibionts of an Introduced King Crab in the Barents Sea: A Second Five-Year Study. Diversity 2023, 15, 29. https://doi.org/10.3390/d15010029
Dvoretsky AG, Dvoretsky VG. Epibionts of an Introduced King Crab in the Barents Sea: A Second Five-Year Study. Diversity. 2023; 15(1):29. https://doi.org/10.3390/d15010029
Chicago/Turabian StyleDvoretsky, Alexander G., and Vladimir G. Dvoretsky. 2023. "Epibionts of an Introduced King Crab in the Barents Sea: A Second Five-Year Study" Diversity 15, no. 1: 29. https://doi.org/10.3390/d15010029