
Spatial Patterns and Determinants of Endemic Taxa Richness in the Genus Viburnum (Adoxaceae) in China
 , , and
, , and
Abstract
:1. Introduction
2. Methods
2.1. Plant Taxa and Distribution Data
2.2. Species Distribution Model
2.3. Environmental Variables
2.4. Data Analysis
3. Results
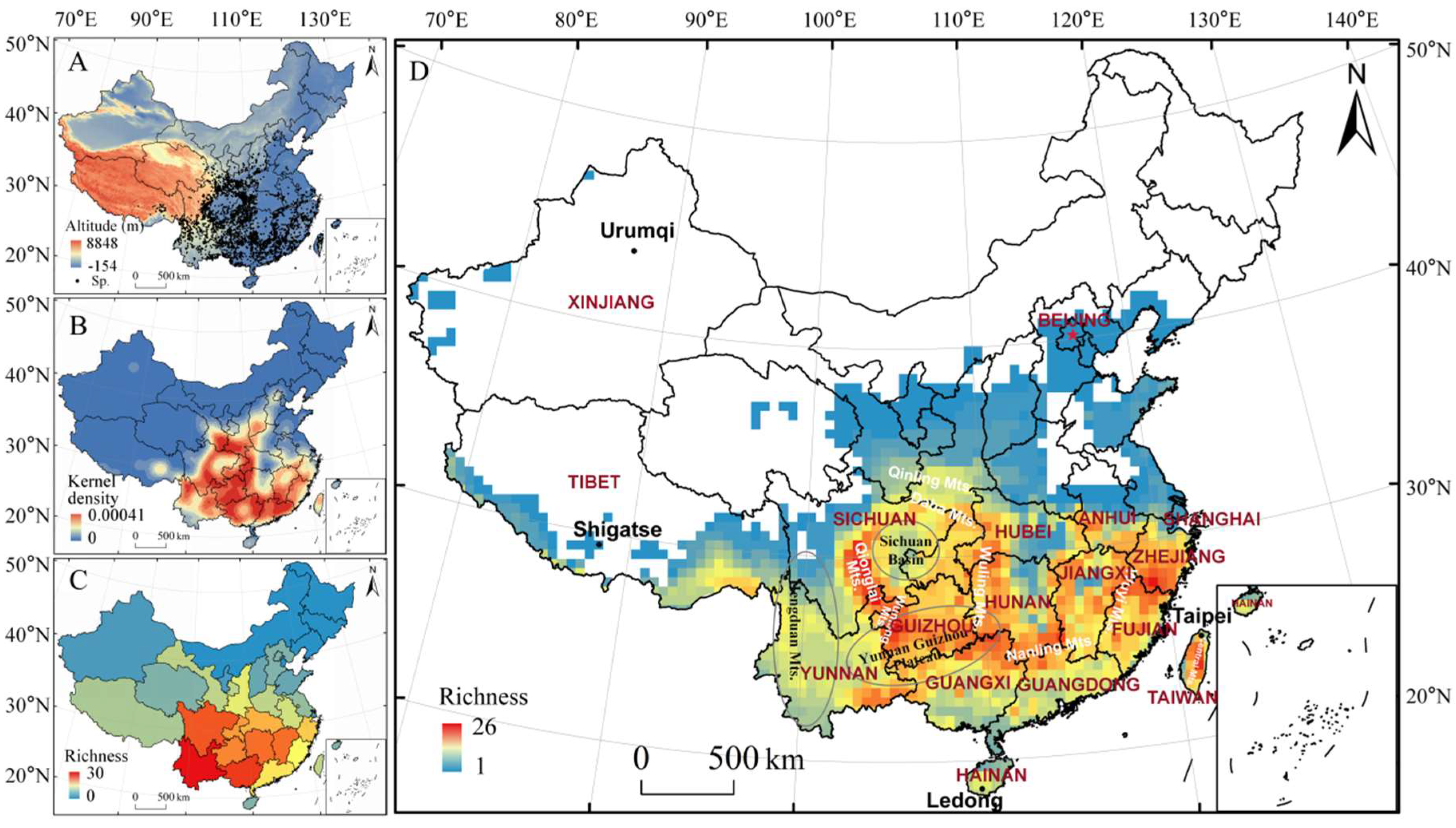
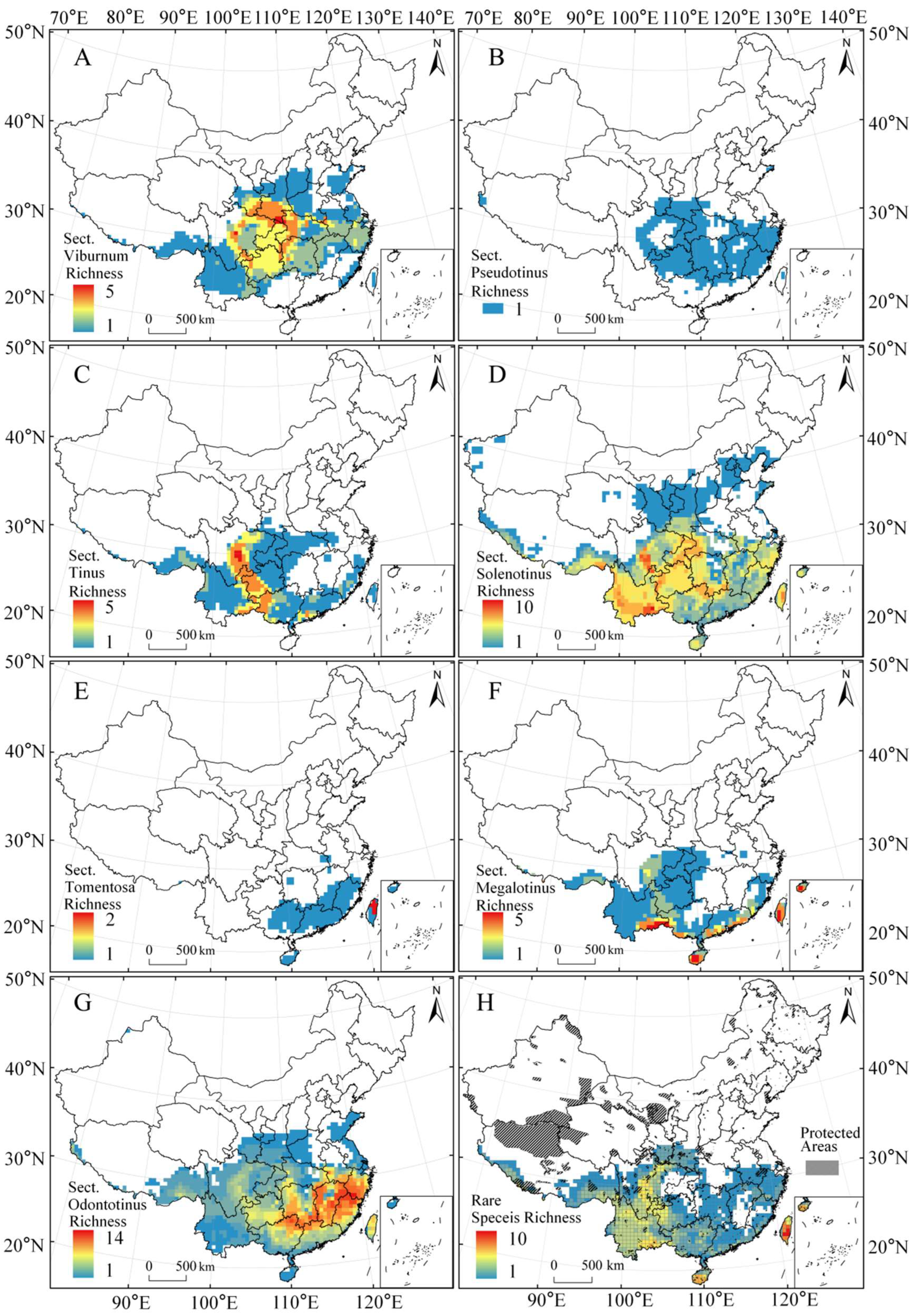
3.1. Endemic Richness of Genus Viburnum in China
3.2. Geographic Patterns of Endemic Richness of Genus Viburnum in China
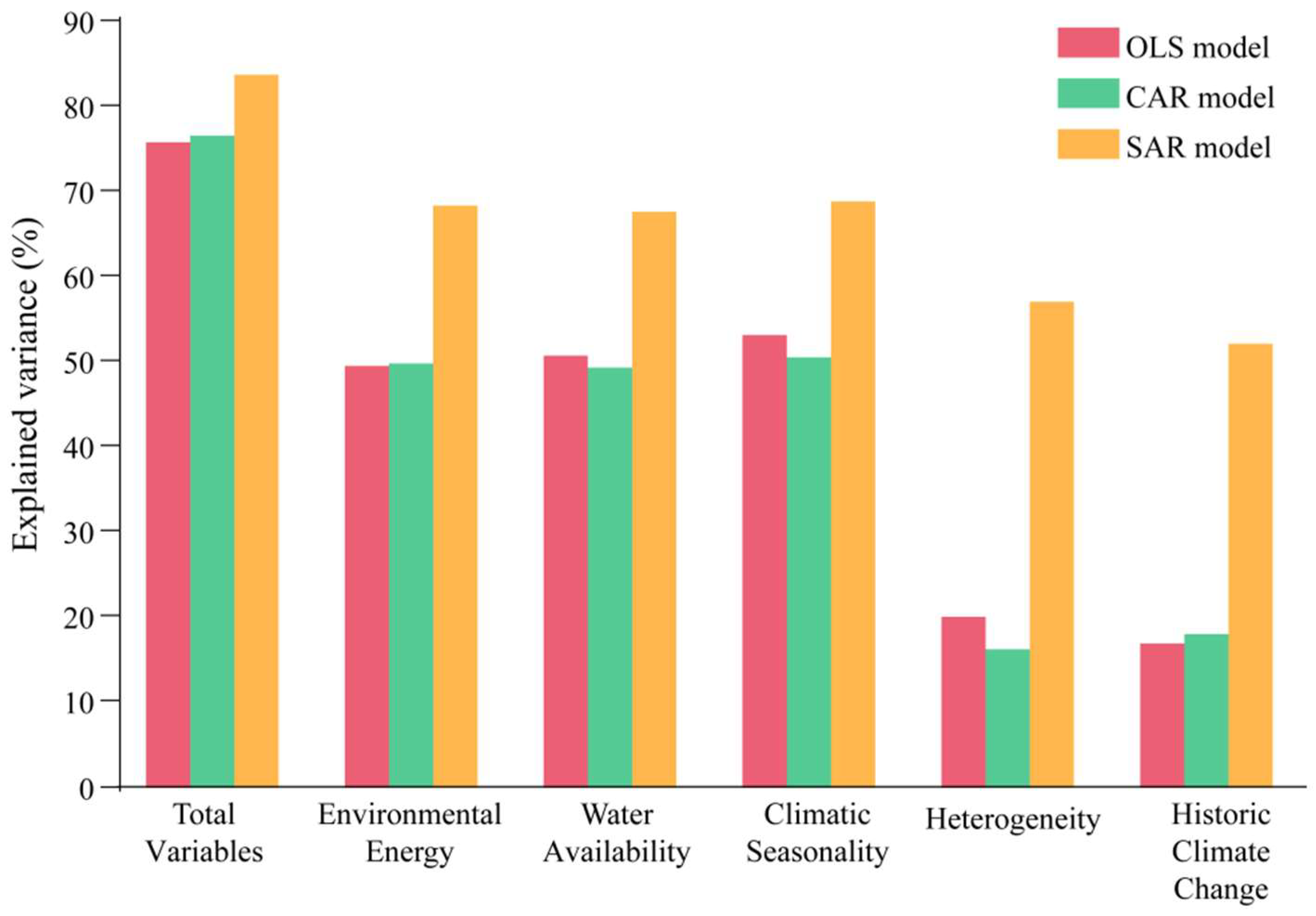
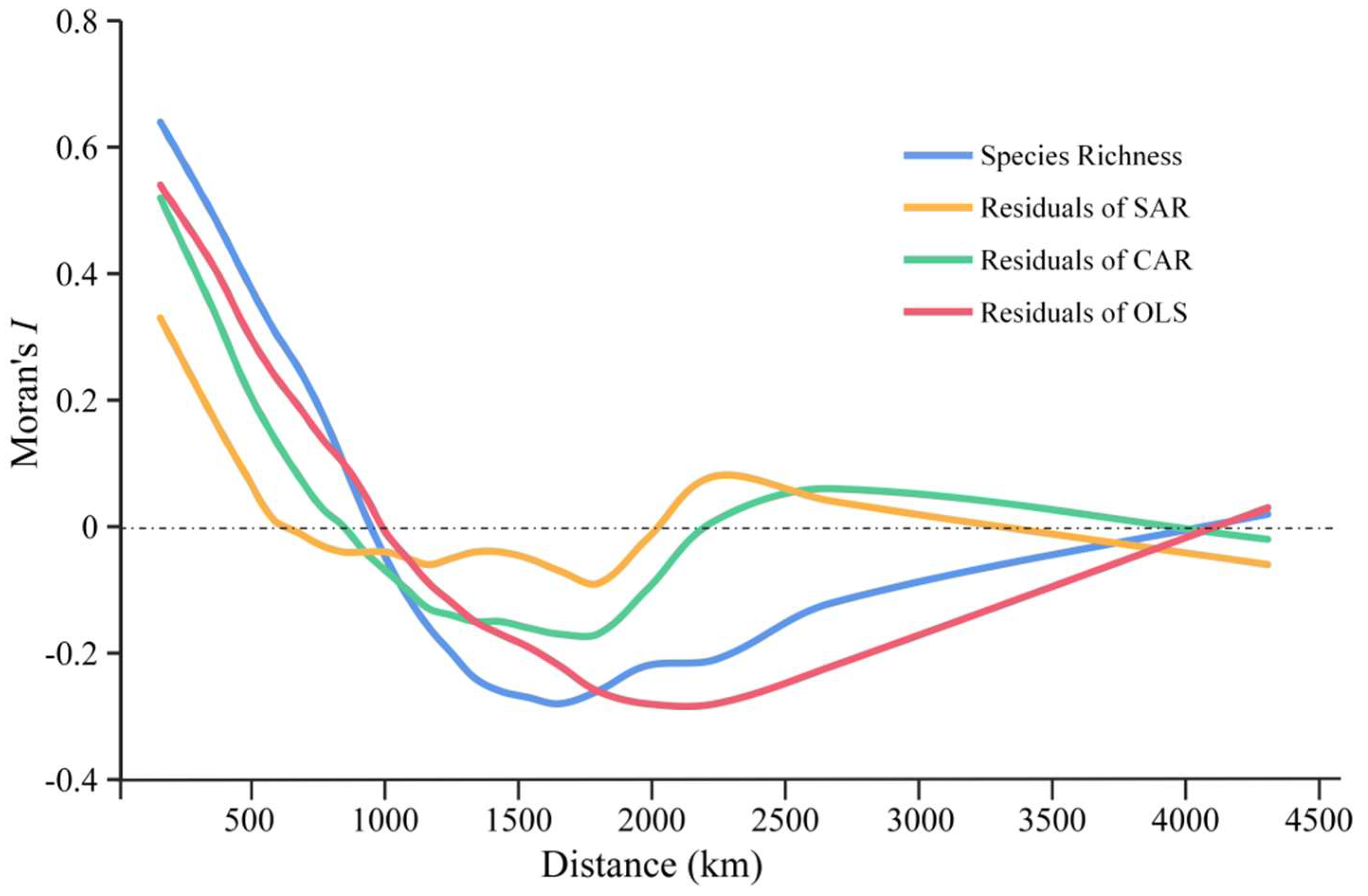
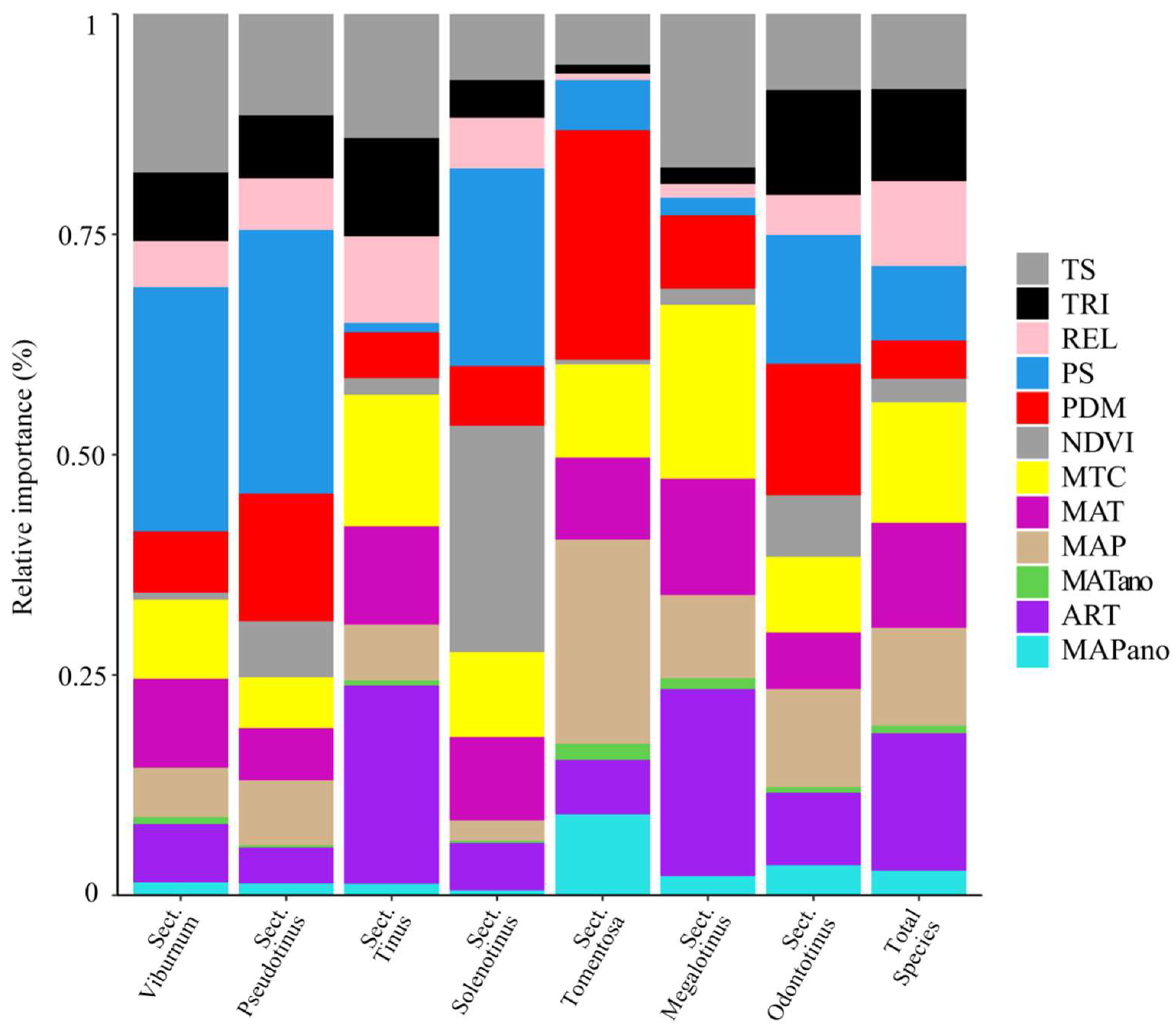
3.3. Assessment of Variables on Species Richness
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Ding, Z.; Lie, G.; Zhou, Z.; Singh, P.B.; Zhang, Z.; Hu, H. Species richness patterns of vascular plants and their drivers along an elevational gradient in the central Himalayas. Glob. Ecol. Conserv. 2020, 24, e01279. [Google Scholar] [CrossRef]
- Arvid Grytnes, J.; Vetaas, O.R. Species Richness and Altitude: A Comparison between Null Models and Interpolated Plant Species Richness along the Himalayan Altitudinal Gradient, Nepal. Am. Nat. 2002, 159, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-F.; Xu, X.; Cheng, X.-L.; Liu, Y.; Luo, A.; Lyu, T.; Wang, W.-L.; Nizamani, M.M.; Wang, Z. Spatial patterns and determinants of Moraceae richness in China. J. Plant Ecol. 2022, rtac025. [Google Scholar] [CrossRef]
- Dakhil, M.A.; Li, J.; Pandey, B.; Pan, K.; Liao, Z.; Olatunji, O.A.; Zhang, L.; Eid, E.M.; Abdelaal, M. Richness patterns of endemic and threatened conifers in south-west China: Topographic-soil fertility explanation. Environ. Res. Lett. 2021, 16, 034017. [Google Scholar] [CrossRef]
- Rahbek, C.; Graves, G.R. Multiscale assessment of patterns of avian species richness. Proc. Natl. Acad. Sci. USA 2001, 98, 4534–4539. [Google Scholar] [CrossRef]
- Xu, H.; Cao, M.; Wu, J.; Cai, L.; Ding, H.; Lei, J.; Wu, Y.; Cui, P.; Chen, L.; Le, Z.; et al. Determinants of Mammal and Bird Species Richness in China Based on Habitat Groups. PLoS ONE 2015, 10, e0143996. [Google Scholar]
- Chen, S.-B.; Jiang, G.-M.; Ouyang, Z.-Y.; Xu, W.-H.; Xiao, Y. Relative importance of water, energy, and heterogeneity in determining regional pteridophyte and seed plant richness in China. J. Syst. Evol. 2011, 49, 95–107. [Google Scholar] [CrossRef]
- Kerr, J.T.; Packer, L. Habitat heterogeneity as a determinant of mammal species richness in high-energy regions. Nature 1997, 385, 252–254. [Google Scholar] [CrossRef]
- Qian, H.; Kissling, W.D. Spatial scale and cross-taxon congruence of terrestrial vertebrate and vascular plant species richness in China. Ecology 2010, 91, 1172–1183. [Google Scholar] [CrossRef]
- Barajas-Barbosa, M.P.; Weigelt, P.; Borregaard, M.K.; Keppel, G.; Kreft, H. Environmental heterogeneity dynamics drive plant diversity on oceanic islands. J. Biogeogr. 2020, 47, 2248–2260. [Google Scholar] [CrossRef]
- O’Brien, E. Water-energy dynamics, climate, and prediction of woody plant species richness: An interim general model. J. Biogeogr. 1998, 25, 379–398. [Google Scholar] [CrossRef]
- Fang, W.-J.; Cai, Q.; Zhao, Q.; Ji, C.-J.; Zhu, J.-L.; Tang, Z.-Y.; Fang, J.-Y. Species richness patterns and the determinants of larch forests in China. Plant Divers. 2022. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Jetz, W.; Rahbek, C. Geographic Range Size and Determinants of Avian Species Richness. Science 2002, 297, 1548–1551. [Google Scholar] [CrossRef]
- Harrison, S.; Noss, R. Endemism hotspots are linked to stable climatic refugia. Ann. Bot. 2017, 119, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Svenning, J.-C.; Skov, F. Ice age legacies in the geographical distribution of tree species richness in Europe. Glob. Ecol. Biogeogr. 2007, 16, 234–245. [Google Scholar] [CrossRef]
- Chen, S.-Y.; Liu, C.-P.; Baskin, C.C.; Chien, C.-T. Deep complex morphophysiological dormancy in seeds of Viburnum plicatum var. formosanum (Adoxaceae) from subtropical mountains. Seed Sci. Res. 2021, 31, 236–242. [Google Scholar] [CrossRef]
- Giladi, I.; Ziv, Y.; May, F.; Jeltsch, F. Scale-dependent determinants of plant species richness in a semi-arid fragmented agro-ecosystem. J. Veg. Sci. 2011, 22, 983–996. [Google Scholar] [CrossRef]
- Leo, M.; Calleja, J.A.; Lara, F.; Garilleti, R.; Medina, N.G. Drivers of plant richness patterns of Mediterranean riparian forests at local and regional scales have bottom-up and top-down effects. J. Veg. Sci. 2019, 30, 485–497. [Google Scholar] [CrossRef]
- Huang, E.; Chen, Y.; Fang, M.; Zheng, Y.; Yu, S. Environmental drivers of plant distributions at global and regional scales. Glob. Ecol. Biogeogr. 2021, 30, 697–709. [Google Scholar] [CrossRef]
- Xie, W.-J.; Leus, L.; Wang, J.-H.; Van Laere, K. Fertility barriers in interspecific crosses within Viburnum. Euphytica 2017, 213, 34. [Google Scholar] [CrossRef]
- Ran, H.; Liu, Y.; Wu, C.; Cao, Y. Phylogenetic and Comparative Analyses of Complete Chloroplast Genomes of Chinese Viburnum and Sambucus (Adoxaceae). Plants 2020, 9, 1143. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Quispe, C.; Vergara, C.V.; Kitic, D.; Kostic, M.; Armstrong, L.; Shinwari, Z.K.; Khalil, A.T.; Brdar-Jokanovic, M.; Ljevnaic-Masic, B.; et al. Genus Viburnum: Therapeutic Potentialities and Agro-Food-Pharma Applications. Oxid. Med. Cell. Longev. 2021, 2021, 26. [Google Scholar] [CrossRef]
- Yang, Q.; Valéry, M. Flora of China. Volume 19: Viburnum L. through Adoxaceae; Science Press: Beijing, China, 2011; pp. 570–611. [Google Scholar]
- Backlund, A.; Bremer, B. Phylogeny of the Asteridae s.str. based on rbcL sequences with particular reference to the Dipsacales. Plant Syst. Evol. 1997, 207, 225–254. [Google Scholar] [CrossRef]
- Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Hara, H. A Revision of the Caprifoliaceae of Japan with Reference to Allied Plants in Other Districts and the Adoxaceae; Academia Scientific Books Inc.: Toyko, Japan, 1983. [Google Scholar]
- Winkworth, R.C.; Donoghue, M.J. Viburnum phylogeny based on combined molecular data: Implications for taxonomy and biogeography. Mol. Phylogenet. Evol. 2005, 92, 653–666. [Google Scholar] [CrossRef]
- Landis, M.J.; Eaton, D.A.R.; Clement, W.L.; Park, B.; Spriggs, E.L.; Sweeney, P.W.; Edwards, E.J.; Donoghue, M.J. Joint estimation of geographic movements and biome shifts during the global diversification of Viburnum. Syst. Biol. 2020, 70, 67–85. [Google Scholar] [CrossRef]
- Schmerler, S.B.; Clement, W.L.; Beaulieu, J.M.; Chatelet, D.S.; Sack, L.; Donoghue, M.J.; Edwards, E.J. Evolution of leaf form correlates with tropical-temperate transitions in Viburnum (Adoxaceae). Proc. Biol. Sci. 2012, 279, 3905–3913. [Google Scholar] [CrossRef]
- Lens, F.; Vos, R.; Charrier, G.; van der Niet, T.; Merckx, V.; Baas, P.; Aguirre, G.J.; Jacobs, B.; Chacon, D.L.; Smets, E.; et al. Scalariform-to-simple transition in vessel perforation plates triggered by differences in climate during the evolution of Adoxaceae. Ann. Bot. 2016, 118, 1043–1056. [Google Scholar] [CrossRef]
- Clement, W.L.; Donoghue, M.J. Dissolution of Viburnum Section Megalotinus (Adoxaceae) of Southeast Asia and Its Implications for Morphological Evolution and Biogeography. Int. J. Plant Sci. 2011, 172, 559–573. [Google Scholar] [CrossRef] [Green Version]
- Clement, W.; Arakaki, M.; Sweeney, P.; Edwards, E.J.; Donoghue, M.J. A chloroplast genome tree for Viburnum (Adoxaceae): Implications for character evolution and phylogenetic classification. Am. J. Bot. 2014, 101, 1029–1049. [Google Scholar] [CrossRef] [PubMed]
- Spriggs, E.L.; Clement, W.L.; Sweeney, P.W.; Madriñán, S.; Edwards, E.J.; Donoghue, M.J. Temperate radiations and dying embers of a tropical past: The diversification of Viburnum. New Phytol. 2015, 207, 340–354. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.-L.; Ye, L.-X.; Chen, F.; Xie, W.-Y.; Chen, Z.-H. An update of the data of the genus Viburnum in Zhejiang. J. Hangzhou Norm. Univ. Nat. Sci. Ed. 2020, 19, 261–266. [Google Scholar]
- Xu, B. Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1988; Volume 72. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Graham, C.H.; Elith, J.; Hijmans, R.J.; Guisan, A.; Townsend Peterson, A.; Loiselle, B.A. The influence of spatial errors in species occurrence data used in distribution models. J. Appl. Ecol. 2008, 45, 239–247. [Google Scholar] [CrossRef]
- Zhang, M.-G.; Slik, J.W.F.; Ma, K.-P. Using species distribution modeling to delineate the botanical richness patterns and phytogeographical regions of China. Sci. Rep. 2016, 6, 22400. [Google Scholar] [CrossRef]
- Wan, J.-N.; Mbari, N.J.; Wang, S.-W.; Liu, B.; Mwangi, B.N.; Rasoarahona, J.R.E.; Xin, H.-P.; Zhou, Y.-D.; Wang, Q.-F. Modeling impacts of climate change on the potential distribution of six endemic baobab species in Madagascar. Plant Divers. 2021, 43, 117–124. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, Y.; Musili, P.M.; Mwachala, G.; Hu, G.; Wang, Q. Inventory incompleteness and collecting priority on the plant diversity in tropical East Africa. Biol. Conserv. 2020, 241, 108313. [Google Scholar] [CrossRef]
- Hosmer, D.W. Wiley Series in Probability and Statistics; Chap. 2; Multiple logistic regression. Applied logistic regression; John Wiley & Sons, Inc.: New York, NY, USA, 2000; pp. 31–46. [Google Scholar]
- Javidan, N.; Kavian, A.; Pourghasemi, H.R.; Conoscenti, C.; Jafarian, Z.; Rodrigo-Comino, J. Evaluation of multi-hazard map produced using MaxEnt machine learning technique. Sci. Rep. 2021, 11, 6496. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Sandel, B.; Arge, L.; Dalsgaard, B.; Davies, R.G.; Gaston, K.J.; Sutherland, W.J.; Svenning, J.C. The Influence of Late Quaternary Climate-Change Velocity on Species Endemism. Science 2011, 334, 660–664. [Google Scholar] [CrossRef]
- Rangel, T.F.; Diniz-Filho, J.A.F.; Bini, L.M. SAM: A comprehensive application for Spatial Analysis in Macroecology. Ecography 2010, 33, 46–50. [Google Scholar] [CrossRef]
- Groemping, U. Relative importance for linear regression in R: The package relaimpo. J. Stat. Softw. 2006, 17, 1–27. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: http://www.R-project.org/ (accessed on 10 August 2021).
- Liu, Y.; Shen, Z.; Wang, Q.; Su, X.; Zhang, W.; Shrestha, N.; Xu, X.; Wang, Z. Determinants of richness patterns differ between rare and common species: Implications for Gesneriaceae conservation in China. Divers. Distrib. 2017, 23, 235–246. [Google Scholar] [CrossRef]
- Shrestha, N.; Su, X.; Xu, X.; Wang, Z. The drivers of high Rhododendron diversity in south-west China: Does seasonality matter? J. Biogeogr. 2018, 45, 438–447. [Google Scholar] [CrossRef]
- Pandey, B.; Khatiwada, J.R.; Zhang, L.; Pan, K.; Dakhil, M.A.; Xiong, Q.; Yadav, R.K.P.; Siwakoti, M.; Tariq, A.; Olatunji, O.A.; et al. Energy-water and seasonal variations in climate underlie the spatial distribution patterns of gymnosperm species richness in China. Ecol. Evol. 2020, 10, 9474–9485. [Google Scholar] [CrossRef]
- Panda, R.M.; Behera, M.D.; Roy, P.S.; Biradar, C. Energy determines broad pattern of plant distribution in Western Himalaya. Ecol. Evol. 2017, 7, 10850–10860. [Google Scholar] [CrossRef]
- Romdal, T.S.; Araujo, M.B.; Rahbek, C. Life on a tropical planet: Niche conservatism and the global diversity gradient. Glob. Ecol. Biogeogr. 2013, 22, 344–350. [Google Scholar] [CrossRef]
- Löwenberg-Neto, P.; de Carvalho, C.J.; Hawkins, B.A. Tropical niche conservatism as a historical narrative hypothesis for the Neotropics: A case study using the fly family Muscidae. J. Biogeogr. 2011, 38, 1936–1947. [Google Scholar] [CrossRef]
- Chien, C.-T.; Chen, S.-Y.; Tsai, C.-C.; Baskin, J.M.; Baskin, C.C.; Kuo-Huang, L.-L. Deep simple epicotyl morphophysiological dormancy in seeds of two Viburnum species, with special reference to shoot growth and development inside the seed. Ann. Bot. 2011, 108, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, Y.; Sun, X.; Xing, S.; Cong, W.; Liu, X. Study on Dormancy Mechanism and Breaking Dormancy Method of Viburnum sargentii Seeds. Am. J. Plant Sci. 2019, 10, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Currie, D.J.; Paquin, V. Large-scale biogeographical patterns of species richness of trees. Nature 1987, 329, 326–327. [Google Scholar] [CrossRef]
- Jimenez-Alfaro, B.; Girardello, M.; Chytry, M.; Svenning, J.-C.; Willner, W.; Gegout, J.-C.; Agrillo, E.; Antonio Campos, J.; Jandt, U.; Kacki, Z.; et al. History and environment shape species pools and community diversity in European beech forests. Nat. Ecol. Evo. 2018, 2, 483–490. [Google Scholar] [CrossRef]
- Liao, Z.; Zhang, L.; Nobis, M.P.; Wu, X.; Pan, K.; Wang, K.; Dakhil, M.A.; Du, M.; Xiong, Q.; Pandey, B.; et al. Climate change jointly with migration ability affect future range shifts of dominant fir species in Southwest China. Divers. Distrib. 2020, 26, 352–367. [Google Scholar] [CrossRef]
- Lu, L.-M.; Mao, L.-F.; Yang, T.; Ye, J.-F.; Liu, B.; Li, H.-L.; Sun, M.; Miller, J.T.; Mathews, S.; Hu, H.-H.; et al. Evolutionary history of the angiosperm flora of China. Nature 2018, 554, 234–238. [Google Scholar] [CrossRef]
- Ding, W.-N.; Ree, R.H.; Spicer, R.A.; Xing, Y.-W. Ancient orogenic and monsoon-driven assembly of the world’s richest temperate alpine flora. Science 2020, 369, 578–581. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OLS Models | CAR Models | SAR Models | |||||||
|---|---|---|---|---|---|---|---|---|---|
| t | R2 | AIC | t | R2 | AIC | t | R2 | AIC | |
| Environmental energy | |||||||||
| MAT | 26.498 | 0.285 | 913.599 | 21.825 | 0.300 | 875.970 | 12.282 | 0.496 | 295.932 |
| ART | −37.480 | 0.444 | 467.920 | −31.713 | 0.458 | 424.184 | −28.419 | 0.679 | −497.299 |
| MTC | 35.452 | 0.416 | 555.496 | 28.107 | 0.416 | 556.969 | 19.114 | 0.571 | 13.626 |
| Water availability | |||||||||
| MAP | 41.050 | 0.489 | 321.442 | 35.986 | 0.504 | 268.409 | 29.583 | 0.673 | −467.821 |
| PDM | 24.220 | 0.250 | 998.358 | 21.927 | 0.269 | 953.337 | 15.860 | 0.531 | 168.554 |
| Climatic seasonality | |||||||||
| TS | −26.890 | 0.291 | 898.774 | −22.896 | 0.316 | 836.727 | −24.031 | 0.639 | −289.754 |
| PS | −24.840 | 0.260 | 975.488 | −23.118 | 0.289 | 903.153 | −13.334 | 0.501 | 280.869 |
| Heterogeneity | |||||||||
| REL | 10.030 | 0.054 | 1407.314 | 13.530 | 0.109 | 1300.729 | 19.866 | 0.538 | 143.528 |
| TRI | 13.680 | 0.096 | 1327.069 | 16.096 | 0.146 | 1226.710 | 19.768 | 0.540 | 135.663 |
| NDVI | −10.880 | 0.063 | 1390.441 | −10.389 | 0.091 | 1337.344 | −5.611 | 0.444 | 468.735 |
| Historic climate change | |||||||||
| MATano | 9.466 | 0.048 | 1417.672 | 6.257 | 0.064 | 1388.166 | 4.388 | 0.447 | 462.120 |
| MAPano | 19.650 | 0.180 | 1155.614 | 15.310 | 0.168 | 1179.985 | 14.132 | 0.519 | 214.806 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, W.; Du, S.; Ying, J.; Ngumbau, V.M.; Huang, S.; Wang, S.; Liu, H. Spatial Patterns and Determinants of Endemic Taxa Richness in the Genus Viburnum (Adoxaceae) in China. Diversity 2022, 14, 744. https://doi.org/10.3390/d14090744
Lyu W, Du S, Ying J, Ngumbau VM, Huang S, Wang S, Liu H. Spatial Patterns and Determinants of Endemic Taxa Richness in the Genus Viburnum (Adoxaceae) in China. Diversity. 2022; 14(9):744. https://doi.org/10.3390/d14090744
Chicago/Turabian StyleLyu, Wenjun, Shenglan Du, Jiali Ying, Veronicah Mutele Ngumbau, Sheng Huang, Shengwei Wang, and Hongtao Liu. 2022. "Spatial Patterns and Determinants of Endemic Taxa Richness in the Genus Viburnum (Adoxaceae) in China" Diversity 14, no. 9: 744. https://doi.org/10.3390/d14090744