
Photoautotrophic Euendoliths and Their Complex Ecological Effects in Marine Bioengineered Ecosystems
by
 , ,
, ,
Alexia M. Dievart
1,* ,
,
Christopher D. McQuaid
1,
Gerardo I. Zardi
1,2,
Katy R. Nicastro
1,3,4 and
Pierre W. Froneman
1 1
Coastal Research Group, Department of Zoology and Entomology, Rhodes University, African Street, Grahamstown 6139, South Africa
2
Laboratoire Biologie des Organismes et Ecosystèmes Aquatiques, Unité Mixte de Recherche 8067 BOREA (CNRS, MNHN, UPMC, UCBN, IRD-207), Normandie Université, UNICAEN, 14000 Caen, France
3
Laboratoire d’Océanologie et de Géosciences, Unité Mixte de Recherche 8187 LOG (Univ. Lille, CNRS, ULCO, IRD), Université de Lille, 59000 Lille, France
4
CCMAR—Centro de Ciencias do Mar, CIMAR Laboratório Associado, Universidade do Algarve, Campus de Gambelas, 8005-139 Faro, Portugal
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(9), 737; https://doi.org/10.3390/d14090737
Submission received: 11 August 2022
/
Revised: 3 September 2022
/
Accepted: 4 September 2022
/
Published: 7 September 2022
(This article belongs to the Section Marine Diversity)
Abstract
:Photoautotrophic euendolithic microorganisms are ubiquitous where there are calcium carbonate substrates to bore into and sufficient light to sustain photosynthesis. The most diverse and abundant modern euendolithic communities can be found in the marine environment. Euendoliths, as microorganisms infesting inanimate substrates, were first thought to be ecologically irrelevant. Over the past three decades, numerous studies have subsequently shown that euendoliths can colonize living marine calcifying organisms, such as coral skeletons and bivalve shells, causing both sub-lethal and lethal damage. Moreover, under suitable environmental conditions, their presence can have surprising benefits for the host. Thus, infestation by photoautotrophic euendoliths has significant consequences for calcifying organisms that are of particular importance in the case of ecosystems underpinned by calcifying ecosystem engineers. In this review, we address the nature and diversity of marine euendoliths, as revealed recently through genetic techniques, their bioerosive mechanisms, how environmental conditions influence their incidence in marine ecosystems and their potential as bioindicators, how they affect live calcifiers, and the potential future of euendolithic infestation in the context of global climate change and ocean acidification.
1. Introduction
Autotrophic (cyanobacteria, and red and green microalgae) and heterotrophic (fungi) euendolithic microorganisms, discovered in the 19th century [1], actively penetrate the hard substrates in which they live [2]. Euendolithic organisms have been present on Earth since the Proterozoic, 2500 to 541 million years ago [3], coinciding with the appearance of oxygen in Earth’s atmosphere, and are thus thought to have played a major role in the development and evolution of life, as well as in the production and destruction of carbonates [4,5,6]. Euendoliths are ubiquitous, as they can be found in almost every environment, geographical location, or depth, where the appropriate substratum (e.g., relatively soluble carbonate and phosphate substrates) is available and their biological requirements are met [4,7,8,9,10,11]. For example, photoautotrophic euendoliths are restricted to the photic zone, where there is sufficient light to carry out photosynthesis, and are particularly diverse and ecologically significant in marine environments [7,8]. They colonize a wide variety of carbonate substrates, from inanimate and dead carbonates to the calcified structures of live animals, such as the skeletons of hard corals, bivalve shells, and the thalli of crustose coralline algae [12,13,14,15,16]. Since their discovery, numerous studies have been conducted on the euendoliths colonizing inanimate and dead calcium carbonates [8,17,18,19,20]. Initially, researchers speculated that the bioerosive activities of euendoliths had little to no effect on infested live calcifying organisms [21]. However, an increasing number of more recent studies suggest that euendolithic infestation has both negative and, surprisingly, beneficial effects on live calcifying organisms [22,23,24,25]. Bivalves, corals, and coralline algae create persistent physical structures that provide habitats for a wide array of associated species [26,27]. Thus, they are referred to as ‘ecosystem engineers’ and can form the basis of entire ecosystems [28,29]. Such marine ecosystems are under multiple anthropogenic pressures, including overexploitation, chronic pollution, and population fragmentation, and their responses to ongoing global climate change and ocean acidification remain uncertain [30,31]. Under such circumstances, the negative and beneficial effects of euendolithic infestation on individual live calcifying organisms have the potential to reverberate at the population, community, or ecosystem level, with complex, and potentially unexpected, ecological outcomes [32,33]. Understanding how euendolithic infestation interacts with other environmental stressors, and how these interactions will change under future environmental conditions is critical to predicting the long-term fate of such ecosystems.
Here, we focus on photoautotrophic euendoliths in the marine environment and their complex ecological effects on live calcifying organisms and the bioengineered ecosystems they create. This review discusses: (1) the nature of marine euendoliths and the techniques used for their observation, (2) the mechanisms involved in their bioerosive activity, (3) their incidence and distribution in marine ecosystems, in relation to various environmental parameters, and their potential as ecological indicators (sensu [34]), (4) how they affect today’s living calcifying organisms, with a focus on corals, coralline algae, and bivalves, and (5) how euendoliths are anticipated to respond to global climate change, and particularly to ocean acidification.
2. What Are Euendoliths and How Are They Observed?
Initially thought to be part of the substrate morphology [1], microborings observed in calcium carbonate substrates were later correctly attributed to the activities of microorganisms, such as algae and fungi [35,36,37], which became known as ‘endoliths’. The term ‘endolith’ refers to a morphologically and physiologically heterogenous group of microorganisms living within a rock or other stony matter, such as coral skeletons or animal shells [38], and more specifically, to organisms that actively bore into relatively soluble substrates, such as phosphate and carbonate substrates [2,8,39] (Figure 1).
While more detailed classifications exist (e.g., [40]), a broad distinction is made among:
- Epiliths that live on the surface of the substrate;
- Chasmoliths that adhere to the surface of fissures and cracks in the substrate;
- Cryptoendoliths that adhere to the surface of pre-existing cavities within porous rocks, including spaces produced and vacated by euendoliths, with no dissolution action;
- Euendoliths that actively penetrate carbonate (and phosphate) substrates and reside partially or completely inside cavities of their own making.
These distinctions are not mutually exclusive as some organisms can display more than a single boring habit or may alter their habits during their life cycles. For example, several boring algae mature on the surface of shells and penetrate the substrate with endolithic filaments, while part of their thallus remains epilithic [41,42]. Endolithic green algae from the genus Ostreobium Bornet and Flahault (1889) switch to a chasmolithic lifestyle after reaching larger cavities within the substrate [43]. Some red algae from the genera Porphyra C. Agardh (1824) and Bangia Lyngbye (1819) can exhibit an endolithic lifestyle during the early stages of their life cycle, called the conchocelis phase, while occurring in a wide variety of habitats as adults [44,45].
The first descriptions of euendoliths were derived from dead mollusk shells gathered from the coast [35,46]. Euendolithic green algae were observed through a thin shell fragment, forming a horizontal layer parallel to the surface, with an underlying network of ramifications into the substrate. Relatively inaccessible (Figure 1), euendolithic microorganisms and their microborings require basic but specific techniques to be studied [8,42,47].
These include:
- Isolation of endoliths. After fixing the sample (substrate and euendoliths of interest) in formaldehyde solution to prevent structural damage to enclosed euendolithic organisms, the surrounding carbonate substrate is usually dissolved using dilute acid. The released endolithic filaments can then be observed and identified under light microscopy or transmission electron microscopy (TEM) [8]. As the hard matrix supporting the euendolithic filaments is dissolved, the organic components of the boring collapse, making spatial relationships and growth arrangements difficult to reconstruct. This technique only allows a qualitative assessment of euendolithic communities but is still widely used to detect and identify euendoliths of interest [15,48,49].
- In situ observations. Microborings can be studied in standard petrographic thin sections or sufficiently transparent fragments of shells using light microscopy [50] or scanning electron microscopy (SEM). The 2D visualization of microborings by petrographic thin sections is of limited use for the study of 3D objects, such as empty or filled euendolithic microborings, but allows their observation and the estimation of their true penetration depth in association with the surrounding matrix [42].
- Cast-embedding of microboring networks [8,42,51]. Combined embedding and casting in polymerized resins preserves the spatial arrangements of boring tunnels (3D architecture) and the euendolithic organisms in situ [51], allowing proper examination after the dissolution of the surrounding substrate (routine protocol in [8], see Figure 2 in [42]). Such casts can then be observed using light microscopy or SEM. However, this technique is limited when microborings are filled by secondary carbonate precipitation following the death of the euendoliths [42].
- Cultivation [48,52,53]. Natural samples (e.g., shell fragments, ooids) harboring euendoliths are used to inoculate agar plates containing a chemically defined culture medium. Once single colonies grow enough to be mechanically isolated, each colony is transferred into a liquid medium and stored under specific environmental conditions. While cultivation selects heavily for fast-growing microorganisms, it is a valuable tool to confirm taxonomic identification and to investigate the life history and physiology of euendoliths [44,45,54]. Cultivation also represents a necessary step to build the databases used for the identification of environmental DNA sequences [52].
- X-ray computed tomography (CT) and micro-computed tomography (micro-CT) (reviewed in [55]). These non-destructive 3D-visualization tools are firmly established for paleontological investigations, with their highest resolution encompassing the spatial magnitude of microborings [56,57,58]. They are, however, still relatively expensive and time-consuming, and require a high level of technical skill [55,58]. More affordable micro-CT and associated analyses can underestimate the extent of bioerosion by photoautotrophic euendoliths [57], as microborings range between 1 and 100 µm in diameter [32]. Micro-CT does, nonetheless, allow the investigation of microborings (empty or filled) within substrates that cannot be altered or dissolved (e.g., type material).
Most techniques used to observe euendoliths focus on the characteristic pattern of their microborings (i.e., form, diameter, direction, length, and pattern of the tunnel), which allows taxonomic identification even in the absence of the organism itself. Different euendolithic species boring into the same substrate and under uniform ecological conditions will produce distinctive boring patterns, in terms of size, shape, and mode of branching of tunnels [59]. Both biological and mineralogical factors should be considered in the characterization of microborings. While the size, frequency, and branching patterns of microborings are dictated by the size and properties of the euendolithic organism, the directions of growth and the fine sculpture of the inner surface of the microborings are influenced by the mineralogical properties of the substrate [8,59]. Critically, a single euendolithic species can display a large variety of morphologically different patterns when boring into different substrates or under different ecological conditions of light and water supply, amongst others [8]. For example, the microborings produced by intertidal euendolithic cyanobacteria are shallower and composed of straight tunnels where desiccation is high, in the supratidal or upper intertidal, while being deeper and tortuous in areas with stagnant water or lower on the shore [8]. These factors reduce the accuracy of taxonomic identification using microborings. Moreover, euendolithic organisms often exhibit few distinguishing morphological features, making taxonomic identification via direct observation equally unreliable [60]. While morphological features of the organism or its traces (i.e., microborings) are useful in the initial discovery of unknown entities, taxonomic identification is best achieved using genetic and molecular techniques [60] and/or cultivation [52].
Used as a complement to morphological descriptions, single- and multi-marker genetic approaches allow the identification of cryptic clades and/or species within euendolithic species complexes and provide tools to determine the composition of natural euendolithic communities. For example, two species within the green algal genus Ostreobium were described using morphological features [35,61], whereas up to 95 delimited cryptic species have been identified within this genus using genetic and molecular techniques [62,63,64,65,66,67]. Environmental DNA (eDNA) metabarcoding allows the automated identification of multiple euendolithic species from a single sample, either containing entire organisms or a piece of substrate containing degraded DNA/RNA (e.g., coral skeleton, bivalve shell), by targeting the amplification and sequencing of one or several specific DNA/RNA regions using universal primers [68]. The composition of euendolithic communities can be investigated using multiple markers, including 16S rRNA and 23S rDNA targeting bacteria and cyanobacteria [64,67,69,70], and 18S rRNA, rbcL, and tufA targeting chlorophytes [54,63,65,67,71]. Moreover, the use of eDNA metabarcoding, in combination with other techniques, such as microscopy, spectrophotometry, and cultivation, has revealed previously undisclosed diversity of prokaryotic and eukaryotic endolithic organisms [52,64,67] and can help resolve their phylogenetic history [64]. For example, euendolithic green algae almost exclusively belong to the class Ulvophyceae, in which the ability to bore evolved independently over 20 times [64], while the cyanobacterium Acaryochloris marina Miyashita and Chihara (2003) has been recorded for the first time in the skeleton of live corals using eDNA [64] and produces chlorophyll-d, allowing it to use far-red light for photosynthesis and thus to occupy niches depleted of visible light [72]. Finally, the correct identification of the euendolithic species is fundamental in the study of euendolithic communities and their impacts, as different strains within the same species complex (e.g., Ostreobium quekettii Bornet and Flahault, 1889) can differ in their physiology [73,74].
3. How Do Euendoliths Erode Calcium Carbonate?
To understand biologically mediated carbonate dissolution, the basis of biotic carbonate precipitation in its simplest form needs to be revisited. Through their metabolic activity, small cells or organisms indirectly create a microenvironment where the concentrations in the chemical species present increase the local precipitation of carbonate, such as calcification by photosynthetic cyanobacterial communities [75,76].
At slightly alkaline or neutral pH, oxygenic photosynthesis can be expressed as:
Protons effectively consumed by oxygenic photosynthesis can then drive the thermodynamic equilibrium of carbonate dissolution–precipitation (with the equilibrium constant expressed as ) toward the solid phase (indicated by (s); here, calcium carbonate):
The free energy of dissolution is given by:
where is the ideal gas constant (8.314 J·K−1·mol−1); , the absolute temperature (in Kelvin); , the equilibrium constant of carbonate dissolution–precipitation; and is the ion activity product [77]. If > , carbonate precipitation will be thermodynamically favored whereas if < , dissolution will be favored. Through their autotrophic metabolism, cyanobacteria and microalgae promote the alkalinization of the medium, increasing local carbonate precipitation. Furthermore, in the case of marine euendoliths, seawater is supersaturated with respect to calcite and aragonite, making carbonate dissolution thermodynamically unfavorable [78]. This leads to an apparent geomicrobial paradox in photoautotrophic euendolithic organisms [78]: how do they achieve local carbonate dissolution, essential for boring, although their autotrophic nature and their surrounding environment should enhance local carbonate precipitation?
The exact mechanisms by which euendolithic organisms bore into carbonate substrates have long remained elusive [4,46,78,79,80,81], with several possible mechanisms being suggested for euendolithic cyanobacteria and later discarded [78,82]. Indeed, suggested boring mechanisms in cyanobacteria need to be consistent with observations and physiological and geochemical principles [78]. Specifically, they need to fulfill three necessary conditions:
- The dissolution process is thermodynamically unfavorable, as it mainly occurs in waters saturated with calcium carbonate (i.e., calcite and aragonite). Excavation then becomes an ATP-driven active process with an energetic cost;
- The carbonate-dissolving mechanisms must be localized at the “head” of the microborer (i.e., apical cells) as dissolution produces true tunnels, with the typical “negative” shape of the borer [59];
- The proposed mechanisms must allow for the conservation of mass and electrical charge as in chemical reactions, the mass and the electrical charge of the components before the reaction must be equal to the mass and the electrical charge of the components after the reaction.
Previously proposed mechanisms included the excretion of weak organic acids, the separation of antagonistic photosynthetic and boring activities, and enzymatic dissolution [79,80,83], but none fulfill all three conditions listed above [78,82].
Garcia-Pichel subsequently proposed an alternative boring mechanism, coined ’The Calcium Pump’, which relies on the active transport of Ca2+ through the cyanobacterial trichome, leading to low concentrations of free Ca2+ at the end of the borehole, which in turn decreases ion activity product below levels that would make dissolution thermodynamically favorable (i.e., < ; Figure 2) [78]. The participation of P-type Ca2+ ATPases and evidence for transcellular Ca2+ transport have been demonstrated for strains of one of the most common cyanobacterial euendoliths, Mastigocoleus testarum Lagerheim ex Bornet and Flahault (1886) [82,84]. Long-range Ca2+ transport within the cyanobacterial filament is made possible through two unique cellular adaptations discovered in this cyanobacterium: active pumping of Ca2+ is orchestrated by the preferential localization of P-type Ca2+ ATPases at one pole of the cell, while specialized cells, called calcicytes, allow fast Ca2+ transport at low, nontoxic concentrations throughout undifferentiated cells (Figure 2). Calcicytes also act as a buffer against excessive, detrimental Ca2+ concentrations before final excretion from the borehole [84]. Furthermore, ’The Calcium Pump’ model fulfills the three necessary conditions enumerated earlier: it is an active process, localized at the end of the borehole, which allows for the conservation of mass and electrical charge [78]. The model is also consistent with the observation of microetchings [79,85], which, in the absence of hard or moving parts in cyanobacteria, points towards carbonate dissolution by cation removal, and with the range of bored substrates, which share Ca2+ as a common denominator [78]. Finally, it alleviates reprecipitation of calcium carbonate within the borehole thanks to the intracellular transport of Ca2+ [78]. While these studies have shed light on the boring mechanism for euendolithic cyanobacteria [86], details regarding the boring mechanism carried out by other photoautotrophic euendoliths have still not been investigated in depth. However, Krause et al. have recently demonstrated the existence of active calcium uptake by the green alga Ostreobium quekettii, occurring at the apical tip of the siphonal thallus, in a process similar to that exhibited by cyanobacteria [87]. All microborers exclusively use chemical bioerosion [32,78] and with regards to these latest findings, further studies are required to reveal the exact boring mechanism of endolithic green and red algae.
As to why euendoliths bore, Garcia-Pichel et al. suggested that they are metabolically dependent on the substrate for dissolved carbon dioxide CO2 released during substrate excavation that can be used for photosynthesis [82]. Euendoliths preferentially fix carbon from the most readily available source, either from the atmospheric pool of dissolved CO2 or the mineral substrate they bore into [88]. When dissolved CO2 is limited, euendolithic microorganisms, such as Mastigocoleus testarum, grow preferentially as euendoliths and thus, derive most of their carbon from the mineral substrate they excavate [88]. By doing so, euendoliths can complete their geomicrobial action on the substrate and potentially gain a competitive advantage over photosynthetic epiliths, which may suffer from limited dissolved CO2 access as the epilithic biofilm thickens [88]. Considering a variety of possible selection pressures (e.g., nutrient acquisition, protection from UV and/or predators), Cockell and Herrera suggested that the boring behaviors of euendoliths could also have originated as mechanisms against entombment by mineralization, especially in substrates with high mineralization rates [81].
![Diversity 14 00737 g002]()
Figure 2.
(Right panel). Proposed ’Calcium Pump’ transport mechanism within the trichome of euendolithic cyanobacteria (both filamentous and pseudofilamentous), with the inferred distributions of transporter components (legend in the upper left corner) and the potential fates of the calcite dissolution products (Ca2+ and CO32−). The calcium transport unit consists of repeating individual bipolar cells, where one pole is specialized for calcium uptake and the opposite for calcium extrusion. Conversion of carbonate ions CO32− released from calcite into HCO3− is promoted by the counter-transported protons. This HCO3− is then actively transported within the cell, where it is converted into CO2 through the carbon-concentrating mechanisms and can then be used in photosynthesis [89]. (Left panel) Holistic calcium ion localization during boring, with relative calcium concentrations within each compartment indicated. Calcicytes allow a higher proportion of cells within the trichome to remain photosynthetically active by controlling intrafilamentous calcium flow (modified from [90]).
Figure 2.
(Right panel). Proposed ’Calcium Pump’ transport mechanism within the trichome of euendolithic cyanobacteria (both filamentous and pseudofilamentous), with the inferred distributions of transporter components (legend in the upper left corner) and the potential fates of the calcite dissolution products (Ca2+ and CO32−). The calcium transport unit consists of repeating individual bipolar cells, where one pole is specialized for calcium uptake and the opposite for calcium extrusion. Conversion of carbonate ions CO32− released from calcite into HCO3− is promoted by the counter-transported protons. This HCO3− is then actively transported within the cell, where it is converted into CO2 through the carbon-concentrating mechanisms and can then be used in photosynthesis [89]. (Left panel) Holistic calcium ion localization during boring, with relative calcium concentrations within each compartment indicated. Calcicytes allow a higher proportion of cells within the trichome to remain photosynthetically active by controlling intrafilamentous calcium flow (modified from [90]).

4. Incidence of Photoautotrophic Euendoliths in Marine Ecosystems
Photoautotrophic euendoliths have a cosmopolitan geographical distribution and have been recorded in a variety of habitats, including terrestrial [9,91], freshwater and volcanic lakes [11,92], brackish [93], and marine environments [4,94]. Euendoliths are ubiquitous in the marine environment, occurring in enclosed seas, such as the Adriatic Sea [16] and the Mediterranean Sea [95,96], in cold-temperate [97,98,99], tropical waters [13,100,101,102], as well as in the Arctic and Antarctic [94,103]. Present essentially anywhere, there is sufficient light to allow for photosynthesis and a carbonate substrate to bore into; euendolithic communities play an important role in ecological processes in the marine environment [4,104]. Although they appear to erode virtually all suitable substrates, the distribution of euendoliths and the composition of euendolithic communities are extremely variable and depend on light availability, the nature of the substrate, and a variety of abiotic and biotic environmental factors acting in synergy [19,52,105].
4.1. Light Availability
As photosynthetic organisms, light availability is the major determinant of euendolithic activity and distribution and has a strong influence on the composition of euendolithic communities [105], reflecting the specific light requirements of different species [106]. In most marine habitats, light availability is highly variable and is influenced by the topography of the area, the presence of 3D structures, the nature of the substrate, and water depth.
Euendoliths are more abundant, and erosion more severe, in microhabitats with high light availability, such as sun-exposed surfaces in the intertidal, mostly horizontal, and moderately inclined surfaces high on the shore [14,107,108,109], or in shallow waters [105], compared to microhabitats with low light availability, such as down-facing and shaded substrates [107,110,111]. Reduction of light availability at polluted sites [112,113] or in habitats at greater depths [101,105] similarly reduces euendolith abundance. Geographically, photoautotrophic euendoliths are more abundant, and erosion more severe, at lower latitudes than higher latitudes, in both the intertidal and underwater [33,111,114,115]. As boring by euendoliths is an active mechanism, it is often restricted to environmental conditions optimal for growth [116]. As solar irradiance increases and cloud cover decreases, the depth of the photic zone increases towards lower latitudes [114]. At higher latitudes, temperatures and light availability are generally low and highly seasonal [115], with a prolonged night and ice cover towards the poles [94,103,117] and a condensed photic zone [111,114]. Combined, these unfavorable environmental conditions slow down euendolithic infestation and limit photoautotrophic euendolithic communities to a few specialist species capable of coping with such adverse conditions.
While the composition of euendolithic communities shifts as light availability decreases with increasing depth, their bathymetric distribution is consistent around the world (see Tables 1 and 2 in [42]) [8,19,105,110,111,114,118,119,120,121,122,123]. Euendoliths are ubiquitous in the supratidal, intertidal, and wave spray zones [124], where assemblages are dominated by cyanobacteria and chlorophytes, referred to as the CyChlo-association [105], in sediments [101] as well as mollusk shells and coral skeletons [14,97,105]. In the shallow photic zones, the additional conchocelis stages of rhodophytes can be observed (CyChloRho-association) in the early stages of colonization [105]. In the disphotic zone or in shaded microhabitats, where light availability is dramatically reduced, only heterotrophs and low-light specialists amongst the photoautotrophs occur, forming the so-called OstPleHet-association [10,97,105]. These include the cyanobacterium Plectonema terebans Bornet and Flahault ex Gomont (1892) and the chlorophyte Ostreobium quekettii that have been recorded down to about 300 m [125,126,127]. Finally, heterotrophic organisms (i.e., fungi and bacteria) dominate the benthic assemblages of the deep, aphotic zone. At a finer scale, different clades within the same euendolithic species can be distributed along a depth gradient, suggesting different physiological traits [63]. Not only does the composition of euendolithic communities change with depth, but also with time, as mature euendolithic communities, even in shallow, clear waters, are dominated by the OstPleHet-association [105].
4.2. Nature of the Substrate
Photoautotrophic euendoliths colonize a wide range of carbonate substrates, from compact limestone and loose sediments to living calcifying organisms or their fragmented remains [8]. Euendoliths have been recorded in the skeletons of corals [13,32,128] and coralline algae [15,16,129], in the shells of mollusks [14,100,130] and brachiopods [131], in the tests of foraminifera [124], in the calcareous tubes of annelids [99] and the plates of barnacles [103], and in sclerosponges [132].
Colonization by and distribution of euendoliths is intrinsically influenced by the nature and physical properties of the substrate, such as its mineralogy, porosity, translucency, density, or architecture [39,52,101]. While most euendoliths appear to be generalists, substrate preferences are found in some, such as the cyanobacterium Mastigocoleus testarum that bores into calcium carbonate substrates but not into other carbonates [39,116]. Skeletal remains of calcifying organisms are more susceptible to euendolithic infestation than other carbonate substrates [43,50,101,133], with the highest levels of an infestation occurring in the densest and least porous substrates. Within skeletal remains, the high-Mg calcite skeleton of crustose coralline algae is more susceptible to dissolution than the skeletons of massive or branching corals or bivalve shells, which are mostly composed of aragonite [134,135]. In shallow waters, the depth of penetration of euendoliths in dead coral skeletons nearly approximates that in live bivalve shells [19,100,101,102]. This is a result of the higher porosity and translucency of the coral skeleton [32], and the presence of organic lamellae (i.e., conchiolin) within bivalve shells that slow down excavation by photoautotrophic euendoliths [50]. Only heterotrophic euendoliths, such as fungi, can penetrate and proliferate into organic lamellae, as they feed on the organic matter [50,100,136].
In live calcifying hosts (e.g., corals, coralline algae, bivalves), a wide range of defenses prevents the colonization of the calcified parts by euendoliths. Coral skeletons are protected by the polyp tissue [13,121,137], while coralline algae have the capacity of sloughing their protective epithelial cells to prevent biofouling [138]. Bivalve, brachiopod, and other mollusk shells have a protective layer, the periostracum, which deters fouling organisms [139,140,141,142,143]. Nonetheless, the incidence of euendolithic infestation in live calcifying organisms is high around the world, independent of the location or the substrate. Nearly all corals around the world have been recorded as infested by euendoliths [13,128,132], while up to 90% of bivalves shells are infested on rocky shores worldwide [14,96,107,108]. After the death of the calcifying organisms, colonization becomes more intense, as euendoliths do not have to overcome the active or passive defense mechanisms of the host or adjust their boring performances to carbonate accretion rates of living organisms [13,15,101,137]. In newly available dead carbonate substrates, a succession of microborer communities can be observed, boring from the surface down into the substrate [13,144,145,146]: (i) pioneer species, such as the large chlorophyte Phaeophila sp. Hauck (1876) and the cyanobacterium Mastigocoleus testarum, settle within the first three months, (ii) these are followed by an intermediate stage, between 3 and 6 months, where the chlorophyte Ostreobium sp. starts to dominate euendolithic communities, and (iii) the final stage, largely dominated by Ostreobium, after more than 6 months of exposure.
4.3. Biotic and Abiotic Environmental Factors
In addition to light, biotic, and abiotic environmental factors can act in synergy to influence the composition and density of euendolithic communities, as well as their rates of microbioerosion. In the intertidal, the abrasive effects of sand and other sediments carried by the winds or the waves favor the initial colonization of the substrate by euendoliths, and ultimately increase the severity of infestation in mussels [14]. Meanwhile, in shallow waters, nutrient concentration, epilithic cover, and the presence of macroborers and macrograzers interact to shape euendolithic communities [19,147,148,149]. This interaction operates through several mechanisms:
- Grazers are attracted to the substrate by the presence of photoautotrophic euendoliths, as these represent a renewable source of food [15,19,151]. The boring activity of euendoliths weakens the superficial layers of the substrate, which can facilitate the settlement of macroborers with their own bioerosive activity, as well as grazing;
- On the one hand, macrograzers constantly remove the superficial layers of the substrate, thus extending the depth to which the light can penetrate and, therefore, the depth to which the endoliths can bore, increasing microboring rates [150,152]. Grazing also reduces the settlement and growth of epilithic organisms that compete with euendoliths for space and diminish light availability [17]. On the other hand, macroborers excrete different waste products within the infested substrate, such as ammonium, phosphates, or CO2. Such waste products act as fertilizer for euendolithic communities, which increase in abundance, biomass, and productivity in the vicinity of macroborers [149,152,153].
Under intense grazing pressure, euendolithic growth and boring rates cannot keep pace with the rapid removal of the substratum by grazers, resulting in lower microboring rates [119]. If grazing exceeds euendolithic boring rates, the food resource is eventually exhausted, and the denuded surface must be recolonized: intense grazing impedes the development of euendolithic communities so that they rarely reach maturity [18,119]. Different groups of organisms are more or less efficient at grazing [113]. Sea urchins are efficient grazers compared to fish, and remove a larger portion of the carbonate substrate, resulting in a lower measurable boring activity of euendoliths in sites with higher densities of sea urchins than of fish [113,119,154,155].
Depending on its nature, the epilithic communities covering the bored substrate can also influence the abundance and composition of euendolithic communities, by diminishing light availability and attracting/deterring macrograzers [105,113,119,122,156]. With time, epiliths start to colonize the surface of the bored substrate and become denser, filtering out light for the underlying euendolithic communities. Interactions between epilithic cover and euendolithic communities are generally reported for substrates exposed for long periods of time, usually >6 months [105,122,157,158]. Under low grazing pressure, the pioneer species Mastigocoleus testarum dominates euendolithic communities on experimental substrates covered with algal turfs, joined by the low-light specialists Plectonema terebrans and Ostreobium quekettii on shaded substrates covered with crustose coralline algae or macroalgae [113,119]. Depending on its nature, the epilithic cover can influence the intensity of macrograzing on the bored substrate: algal turfs attract macrograzers while crustose coralline algae and macroalgae, which are unpalatable or inedible for most grazers, act as a deterrent [119,151,154].
Nutrient concentrations in the surrounding environment influence the abundance, species composition, and microbioerosion rates of euendolithic communities, as well as the density and species composition of the epilithic communities [112,119]. At sites subjected to eutrophication, microbioerosion rates by euendoliths are higher than in more oligotrophic waters, in association with either low [119] or high grazing pressure [152]. Conversely, nutrient-enriched turbid inshore waters are characterized by lower microbioerosion rates compared to clear oligotrophic offshore waters at the Great Barrier Reef, suggesting that increased turbidity resulting from the entrapment of sediments in the epilithic cover at inshore sites diminishes light availability and, thus, restricts euendolithic colonization, even at high nutrient concentrations [113]. Elevated concentrations of inorganic nutrients (e.g., nitrogen, phosphorus) drastically increase the severity of colonization, the depth of penetration, and ultimately the microbioerosion rates of euendolithic communities, especially in the absence of macrograzing [156,157,158]. Different euendolithic taxa display variable responses to the addition of nutrients, depending on the nature of the nutrient and the specific limitations of the euendoliths; while heterotrophic euendoliths increase in abundance when exposed to increased levels of organic matter, euendolithic green algae and cyanobacteria react to higher levels of nitrogen and phosphate, respectively [156,157,158]. In live calcifying substrates, the effects of nutrient addition on euendolithic communities can be mitigated by the host response to the same nutrients. For example, when phosphorus is added, the skeletal growth rates of hard corals increase and “dilute” euendolithic communities, as they are unable to keep up with increased coral growth [159]. Additionally, nutrient concentrations can influence the species composition and density of epilithic communities [112], with an indirect impact on the underlying euendolithic communities [105,119,122]. In coral reefs with low nutrient loads, the epilithic cover is dominated by algal turfs associated with pioneer euendolithic species that require high light intensities, while in reefs subjected to eutrophication, the dominance shifts to macroalgae and crustose coralline algae, diminishing light availability for an underlying euendolithic community abundant in low-light specialists [119].
4.4. Photoautotrophic Euendoliths as Bioindicators
With their specific niche specializations, in terms of nutrition, light, temperature, nutrient concentrations, and other physical parameters, and the characteristic microborings they produce, photoautotrophic and heterotrophic euendoliths can be used as present geographical, water quality, and paleobathymetric bioindicators at the species or community level [8,124]. Similarly, fossil euendoliths could potentially be used as indicators of past temperatures, salinity levels, or trophic dynamics, but more research is still needed [42]. Some euendoliths, such as members of the genus Ostreobium, display a complex biogeographical distribution. Ostreobium has been found in abundance on temperate coasts but becomes rare in tropical waters [50,133], showing a clear latitudinal pattern. Within the genus, Ostreobium quekettii dominates endolithic assemblages in nearly all corals, while O. constrictum K.J. Lukas (1974) and O. brabantium Weber Bosse (1932) have only been found in the Atlantic and Pacific Oceans, respectively [61]. More recently, del Campo et al. revealed a complex biogeographical distribution of three identified clades within the genus Ostreobium across coral reefs [160]. However, there is a need for more recent and detailed regional comparisons of the incidence of individual euendolithic species and the composition of euendolithic assemblages, as advances in molecular genetic techniques and the use of polyphasic approaches allow better detection and taxonomic identification of euendolithic specimens [52,64,67].
Photoautotrophic euendoliths have the potential to be employed as indicators of water quality and/or pollution [156,157,158,161,162]. The composition of euendolithic communities shifts depending on the concentrations of inorganic nutrients and organic matter in the surrounding environment [156,157,158]. In the Arabian Gulf, oil pollution is responsible for a dramatic decrease in the abundance of live euendoliths and microbioerosion rates on foraminifera [161], while in Italy, benthic foraminifera demonstrate increased levels of euendolithic infestation by photoautotrophic euendoliths in sites with heavy metal pollution [162,163].
Modern euendolithic bathymetric distribution can be reliably used as an indicator of the current depth, as it can be cross-checked by direct measurements of the physical parameters. Through their boring activity, microbial euendoliths leave traces, as an “instant fossil” in the sedimentary record, each with its own bathymetric range [124] and remarkable constancy through time. While single fossil traces can be used for the identification of individual ichnotaxa, the study of euendolithic trace communities (or ichnocoenoses) within the substrate allows for paleobathymetric and paleoecological reconstructions over geological time scales [8,20]. The light-dependence of photoautotrophic euendoliths thus reveals itself as a useful tool to determine the limit between past photic and aphotic zones [20]. The study of fossil trace assemblages in paleodepth reconstruction allows for a finer detection of past sea level variations than other commonly used methods [120], such as the analyses of the successive coralgal communities [164]. However, fossil microborings should be compared with modern microborings from similar substrate types [120,122] and the ichnocoenoses described must be directly compared with modern assemblages for interpretation [8]. Indeed, a bathymetric ichnocoenoses index could provide misleading paleobathymetric conclusions in shallow waters when 3D-underwater structures (e.g., reefs) create heterogeneous light conditions on the infested substrate (i.e., illuminated, shaded, and cryptic habitats) at the same depth [98,110,165]. Similarly, the investigated substrate could have been translocated from shallow depths to its deposition site [42]. Thus, fossil microborings should be viewed as semiquantitative rather than absolute indicators of paleophotic zones [165,166].
5. Photoautotrophic Euendoliths in Marine Bioengineered Ecosystems
While numerous studies have assessed euendolith-induced biodegradation of carbonate skeletal materials, until recently, severe harm to living host organisms was understood to be limited to the erosive activity of invertebrates or fungal borers [21]. Due to low light penetration within the substratum, photoautotrophic euendoliths were generally thought to be unable to inflict significant structural damage on live organisms, as they eroded only the uppermost layers of the carbonate substrate [21]. Over the last three decades, mounting evidence has shown that the eroding activity of photoautotrophic euendoliths can be the source of severe, often lethal, damage to living calcifying organisms [22,24,107]. However, the presence of euendoliths has also been observed to have beneficial effects [23,25,108].
Here, we focus on three groups of calcifying organisms, i.e., corals, crustose coralline algae, and bivalves, and describe the process of colonization by euendoliths in live hosts, their incidence, and their detrimental and beneficial effects on the individual host, as well as on the bioengineered ecosystem (summarized in Table 1), i.e., coral reefs, coralline algal mats, and bivalve beds.
5.1. Corals and Crustose Coralline Algae
In live corals, the colonization of the skeleton by euendoliths from the water column is prevented by the polyp tissue [121,137,181]. In young coral recruits, the entire corallite (i.e., part of the skeleton elaborated by a single polyp) is tightly covered with polyp tissue [128,167,185], which efficiently protects the underlying calcareous skeleton from infestation by other life stages of endolithic organisms (e.g., propagules, epilithic biofilms), thanks to its superficial mucus and cnidocysts. In some instances, the polyp tissue retracts towards the proximal portions of the coral colony, either temporarily when corals are under major thermal stress [186] or permanently in the case of solitary corals [187], leaving the skeleton unprotected. Therefore, photoautotrophic euendoliths have the ability to colonize the coral skeleton from its base, as soon as the larvae settle on an already-infested substrate [167], or to enter through the exposed skeleton through lateral fissures or when the polyp tissue retracts [186,187]. In live crustose coralline algae (Corallinophycidae), colonization by photoautotrophic euendoliths from the surface is prevented by the presence of live cells (as for corals), capable of ‘sloughing’ as a defense against biofouling [138,188]. With crustose coralline algae (CCA) growing apically, the live tissue remains in the upper part of the thallus, giving it a red color, leaving empty calcified cell walls at the basal layer of the thallus, which turns whitish-gray [15,16]. Without the protective live tissue, euendoliths colonize CCA from its basal layer, which can be in direct contact with an already-infested substrate [15,16]. Thereafter, photoautotrophic euendoliths have to match or exceed skeletal calcification rates, by positive phototrophic growth orientation (i.e., growing from the inside of the skeleton towards the surface) and high rates of growth and carbonate penetration [13,15]. Only a few euendolithic organisms have adapted to low-light conditions, such as the cyanobacterium Plectonema terebrans, the chlorophyte Ostreobium quekettii, and less frequently some conchocelis stages of Bangial rhodophytes, are known to keep pace with coral growth [13,61,63,137,181,189,190]. In addition, the cyanobacterium Mastigocoleus testarum can be found in live CCA crusts [15,16,128], and P. terebrans has been observed boring directly into CCA calcified cell walls [191]. Areas of dense euendolithic growth are often correlated with areas of slower growth in corals, as shown by the patterns of colored bands in massive, slow-growing corals [13,192,193,194]. In fast-growing corals, such patterns are less clear [195], with euendoliths being more abundant in the middle part of the skeleton [159]. The distribution of euendolithic organisms is, however, not limited to the colored bands as they can be present in most regions of the skeleton although in insufficient densities to be visible [13]. In the same fashion, euendolithic infestation in live CCA crusts forms a green band underneath the live tissue [15]. Photoautotrophic euendoliths are ubiquitous in live zooxanthellate corals (Symbiodiniaceae [196]), with 100% of Atlantic, Indian, and Pacific corals being infested [13,132,144,192,193]. In the southern Chilean fjords, 83% of the colonies of the azooxanthellate Desmophyllum dianthus Esper (1974) were infested in the euphotic zone [128]. Photoautotrophic euendoliths have been recorded from live crustose coralline algae in the Mediterranean Sea and the Pacific Ocean [15,16].
The negative effects of endolithic infestation on live corals have largely been demonstrated in azooxanthellate corals, which do not benefit from a symbiosis with zooxanthellae. However, given their similar calcification rates [197], it is reasonable to expect that azooxanthellate and zooxanthellate corals will display similar negative effects to infestation by photoautotrophic euendoliths. Euendolithic infestation affects apical growth and skeletal structure of adult coral colonies [24]; however, it does not slow the extension rates and fitness of coral recruits [167]. Heavily infested areas of the coral Desmophyllum dianthus display a thicker but more porous skeletal structure, reduced apical extension, and enhanced skeletal deformations [24]. Thus, euendolithic infestation alters the calcification pattern of the coral and unbalances the energetic budget towards the maintenance of skeletal integrity, at the expense of the vertical growth of the polyp and other essential biological processes, such as reproduction. Skeletal deformations, resulting from the secondary deposition of aragonite on the coral skeleton, can act as a mechanical barrier to endolithic infestation [24,181], but are linked to increased coral mortality and reduced coral growth and reproduction, thus reducing the overall fitness of the coral [198,199]. As a source of food for grazing organisms [124], euendoliths attract excavating invertebrates and vertebrates, enhancing overall bioerosion activity in coral reefs and leading to deeper damage to the substrate [19,113]. Finally, filamentous cyanobacteria associated with the Black Band Disease have been observed actively boring through both coral skeleton and tissue, indicating a potential relationship between endolithic organisms within the coral skeleton and coral diseases [200]. As euendoliths accelerate internal bioerosion rates in CCA, their infestation is likely to increase CCA porosity [15], with consequences similar to those in corals. As microbioerosion is lower in CCA than in corals (7.9% vs. 25% of the bored substrate) [13,15], its negative effects on CCA might also not be detectable or significant.
As early as the 1950s, Odum and Odum suspected another kind of relationship between photoautotrophic euendoliths and their live coral hosts, one that is positive and mutualistic rather than negative, which they termed ‘ectosymbiosis’ [144]. Forty years later, Schlichter et al. first demonstrated the translocation of photoassimilates from the euendolithic algae living in the coral skeleton to the coral tissue via the uptake mechanisms for dissolved organic substances of cnidarians in the azooxanthellate coral Tubastraea micranthus Ehrenberg (1834) [170]. This interaction increases the polyp’s fitness by increasing productivity and biomass, while in return, the metabolic end products of the coral can be used by the endolithic algae. Subsequently, Schlichter et al. highlighted the role of the green euendolithic alga Ostreobium quekettii in intra-colonial nutrient recycling in two zooxanthellate coral species, though this was considered to be minor compared to the contribution made by zooxanthellae [171].
In live coral skeletons, three main factors influence the availability of light to euendolithic organisms [193,201]:
- Light transmission is affected by the architecture of the coral’s skeleton. Corallites on the top of coral colonies guide light deeper into the coral skeleton, while for corallites on the side of the colony, light enters at an angle, reducing its penetration into the skeleton [193];
- Water depth.
Low light intensities within the coral skeleton suggest an adaptation to shaded environments for euendoliths and, thus, low euendolithic photosynthetic production and a low contribution to the metabolism of healthy zooxanthellate corals [171,190,201,205]. For example, respiration and photosynthetic rates of endolithic algae in the zooxanthellate coral Porites compressa Dana (1846) represent, respectively, only 1.4% and 6% of those of the zooxanthellae, corresponding to the amount of PAR effectively reaching the endoliths [171,205]. During bleaching events, however, corals lose their symbiotic zooxanthellae and their shading effect to the endolithic layer (Figure 3c,d). Photoautotrophic euendoliths then acclimate to the increased solar irradiance penetrating the skeleton [177] and start blooming [206]. Two to three weeks after the onset of bleaching, characteristic greenish (Ostreobium quekettii) and reddish (Plectonema terebrans) pigments of photoautotrophic euendoliths become visible through the now transparent coral tissue (Figure 3d) [23,98,128]. Following this increase in photosynthetic activity, increased quantities of photoassimilates are continuously translocated between the euendoliths and the coral tissue, thanks to the close contact between euendolithic filaments and coral tissue; the cytoplasmic compartments of corals and euendolithic algae are only separated by a cell membrane and by a cell wall and membrane, respectively [170]. On the one hand, due to the capacity of coral cells to absorb dissolved organic substances, such as sugars and amino-acids [207], these photosynthetic products can be readily used by the coral tissue when released by euendoliths, and contribute to the metabolic demands of the coral, especially during bleaching events [23,170,208]. On the other hand, euendoliths may benefit from the internal CO2 pool and nitrogen-containing metabolites of the coral, using these as a source of inorganic carbon for photosynthesis and nitrogen, respectively [209]. Although euendolithic sources of energy are nutritionally less important than those of zooxanthellae [171,208] and insufficient to support sexual reproduction by the coral [210], the bloom of euendolithic organisms in the coral skeleton can promote the survival of zooxanthellate corals during bleaching events until new zooxanthellae are recruited [206]. In the event of a rapid bleaching event, however, high light intensities coupled with high temperatures cause the photoinhibition of euendolithic activity [178]. Moreover, the increased abundance of euendoliths in the coral skeleton during bleaching events can result in a corresponding increase in bioerosion rates, further increasing the porosity of the coral skeleton [148]. Thus, photoautotrophic euendolithic communities can sustain coral survival through a critical supply of organic carbon and nutrients during periods of environmental change but slowly weaken the structural integrity of the coral skeleton, increasing its vulnerability to mechanical damage. Conversely, in live CCA crusts, euendolithic infestation can reduce the susceptibility of CCA skeletons to dissolution, especially under ocean acidification and global climate change [129,135]. Through their boring activity, photoautotrophic euendoliths remove highly soluble magnesium calcite from the skeleton, indirectly increasing internal interstitial pH [179], thereby promoting internal secondary precipitation of minerals [211] and increasing the relative abundance of dolomite in the skeleton [135]. CCA skeletons rich in dolomite are less susceptible to dissolution and skeletal disruption [134,135], as dolomite is a more stable form of carbonate and some euendoliths (such as Mastigocoleus testarum) are unable to bore through it [116]. Because live CCA crusts are only moderately bored compared to corals, it appears that the beneficial effects of euendolithic infestation could largely outweigh their negative effects, even under heat and acidification stress.
Coral reefs are amongst the most diverse and complex marine ecosystems in the world, providing a habitat for countless associated species [27], as well as providing many services to mankind, such as commercial fishing, tourism, and coastal protection against storms and cyclones [27,212]. In tropical reefs, CCA are critical biological and geological components [188], cementing together sand, dead corals, and debris to create a stable substratum [213]. This substratum facilitates the settlement of coral larvae and other invertebrates of commercial value, such as abalone and sea urchins [135,214,215]. Overall, the maintenance of healthy coral reef ecosystems is critically dependent on the balance between constructive forces, such as calcification by corals and encrusting coralline algae, and destructive forces, including bioerosion [216]. Severe euendolithic infestation of individual coral skeletons can fragilize whole coral reefs, increasing their susceptibility to damage by cyclones and storms [212], El Niño events [217,218], and predators, such as the crown-of-thorns starfish [12], and diminishing the coastal protection they offer to other ecosystems and mankind. However, during extreme heat waves, euendolithic infestation of corals can mitigate the effects of bleaching and promote coral survival [23,170,171,178]. In live CCA crusts, euendolithic infestation promotes long-term survival by altering the mineralogical composition of the skeleton [134,135]. Finally, by affecting the inanimate substrates composing the ecosystem as well, endolithic infestation plays a role in shore bioerosion in interaction with grazers [113]. For more detail about the role of boring microflora in modern coral reef ecosystems, see [32].
5.2. Bivalves
In live bivalves, the colonization of the shell by photoautotrophic euendoliths is prevented by the periostracum, a well-defined external proteinaceous coating of variable thickness that constitutes the outermost layer of the shell (Figure 3a) [14,22,93,219]. The early developmental stages of bivalves are largely free of endoliths [94,107], as their protective periostracum is still intact. Over time, the periostracum can be abraded, by contact with neighboring mussels within a bed and/or by sediment particles carried by waves [14,107] or by the activity of macrograzers [94]. Once the periostracum is removed, photoautotrophic euendoliths colonize the bivalve shell from the outside in, in a similar fashion to dead substrates. This involves a succession of microboring species through time [14,49,100]: (i) pioneer species within the first months, such as the cyanobacteria Mastigocoleus testarum and Plectonema terebrans, and the chlorophyte Phaeophila sp., (ii) an intermediate stage after the pioneer species are established, with the addition of the cyanobacterium Solentia stratosa Ercegovic (1927), and (iii) a late successional stage, after more than 6 months of exposure, with the addition of the cyanobacteria Hyella sp. Bornet and Flahault (1888) and/or Pleurocapsa sp. Thuret (1885), and the chlorophyte Ostreobium quekettii, which replaces Phaeophila. In some instances, infested bivalve shells take a greenish color when euendolithic infestation is dense [93,94]. The spatial distribution of cyanobacterial euendoliths in bivalve shells reflects this succession in time, as the last colonizers are only found towards the older parts of the shell, where they co-occur with pioneer species [49]. The succession of microborer communities does not necessarily involve the replacement of euendolithic species by others [100], but rather their addition [14,49]. Consequently, euendolithic infestation is stronger on the oldest parts of the shell (i.e., umbo and central shell), and with increasing shell length, as a proxy of age [22,33,94,96,100,172]. Most studies have focused on the succession of cyanobacterial euendolithic communities on bivalve shells [14,49], so less detail is available for chlorophytes and other photoautotrophic euendoliths [96,100,220]. Photoautotrophic euendoliths colonize intertidal bivalve shells around the world, independent of their location or the nature of the shore [14,94,221]. On South African rocky shores, the incidence of endolith-infested shells in the brown mussel Perna perna Linnaeus (1758) and the blue mussel Mytilus galloprovincialis Lamarck (1819) varied between 23% and 95% on sheltered shores within bays and on shores exposed to the open ocean, respectively [14,107,221]. In California, between 60% and 90% of the shells of the Californian mussels, Mytilus californianus Conrad (1837), were infested by euendoliths, depending on their position with respect to height on the shore [108], while 93% of the shells of the bearded horse mussel, Modiolus barbatus Linnaeus (1758), were infested in the Adriatic Sea [96].
As euendoliths bore into the shell, bivalves need to repair the shell and maintain carbonate structures through secondary carbonate deposition [22], an energetically demanding process requiring 25–50% of their energy budget [222,223]. The bivalve’s energetic budgeting then shifts towards self-repair with time, at the expense of growth, reproduction, attachment strength, and overall condition [22,107,168,173,220]. The severity of euendolithic infestation and its negative effects on bivalves is, however, species-specific. In South Africa, the invasive blue mussel, Mytilus galloprovincialis, exhibits greater infestation levels and negative effects than the native brown mussel, Perna perna [107,221]. Regardless of the rate of shell deposition in infested bivalves (up to two times that of non-infested bivalves), it is never enough to compensate for the degradation caused by euendoliths [22]. Ultimately, infestation becomes so severe that bivalves fracture their own shells with the force applied by their adductor muscles [14]. Thus, endolithic infestation can at times account for more than 50% of total mortality in highly infested populations [22].
Moreover, damage to the individual bivalve exceeds simple structural damage and has critical consequences at the population and community levels. Through their boring activity, euendoliths reduce shell strength and thickness, increasing vulnerability to predation, wave action [22], or trampling [175]. Similarly, shell microtopography and its antifouling characteristics [142] can be altered by euendoliths, which could lead to higher settlement of epibionts such as barnacles (as hypothesized in [172]). Infestations by larger epibionts increase a bivalve’s vulnerability to drag forces and predators [224], while these epibionts’ boring actions could further enhance euendolithic infestation [172,225]. In addition, by creating a more complex, rougher shell surface, euendolithic infestation influences the hydrodynamic properties of mussel beds, leading to greater ingestion rates of microplastics by mussels within infested mussel beds, and greater retention of microplastic on infested mussel shells and within infested mussel beds [184]. Moreover, infested mussels produce fewer byssal threads for attachment to the substratum, making them more vulnerable to drag forces [107,173] and creating an architecturally less complex mussel bed for the associated infauna. Gonad mass, and thus reproductive capacity, are also negatively affected by euendolithic infestation [22,107,173,220]. Larger mussels, responsible for the bulk of gamete output from the population, are the most susceptible to mortality through shell collapse and suffer the greatest decrease of gonad tissue due to endolithic infestation [22], thus reducing population reproductive output.
Shell-boring photoautotrophic euendoliths have, however, also been shown to have beneficial effects on the bivalve host under certain environmental conditions [25,33,108]. Infested mussels lose their dark color through the loss of the periostracum, which exposes the pale nacreous layer underneath and the deposition of excess calcium on the outside of the shell by euendoliths (Figure 3b) [14,25,49,108]. Infested mussel shells, being paler, absorb around 10% less sunlight energy than non-infested shells, resulting in lower body temperatures (up to 5 °C lower) and greater water retention [25,108]. During heat waves, infested mussels can experience lower short-term mortality rates [25,108], as they warm up slower and reach lower body temperatures than non-infested mussels during aerial exposure [115]. The beneficial effects of endolithic infestation can extend to neighboring mussels within a bed: mussel beds mostly composed of infested mussels exhibit lower temperatures and higher humidity in the interstitial space [33,115]. The beneficial effects of euendoliths are regulated by particular topographic and meteorological conditions [33,109,115]. Thermal buffering by euendoliths is greatest when bivalves occur on horizontal or moderately inclined, mostly sun-exposed, surfaces, and under conditions of high solar irradiance and air temperature, low wind speed, and moderate humidity [109,115]. Thus, thermal buffering by euendoliths is the strongest and most ecologically significant when (and where) bivalves suffer from stressful environmental conditions, particularly in the summer and at lower latitudes [33,109,115]. The protection offered to associated species by the bioengineering effects of mussel beds is thus likely to be enhanced by endolith-induced improvements in within-bed humidity and temperature, especially in the upper intertidal zone where mussel beds are typically mono-layered and thermal refuges are scarce [108].
Worldwide, bivalves are the most prominent ecosystem engineers on intertidal rocky shores [226], providing a functional habitat and refuge to a wide array of associated species in a physiologically stressful environment characterized by high temperature and low humidity during emersion [226,227,228,229]. Bivalve beds offer a refuge from thermal and desiccation stresses [229,230], and predators [227,231], a habitat for epibionts [226], and participate in the transport of solutes and organic particles in the near-shore environment [26,232]. Bivalve beds provide many ecosystem services, such as fishing, other resources, protection against the waves, and stabilization of the coastline [233,234]. Euendolithic infestation of bivalve beds increases the susceptibility of these ecosystems to damage by waves and anthropogenic stressors, such as trampling and plastic pollution [175,184], while reducing their vulnerability to heat waves [25,115].
5.3. Other Groups
While limited information is available on the incidence and distribution of photoautotrophic euendoliths in calcifying organisms, the physiological, biological, and ecological impacts of endolithic infestation have not been documented outside of commercially or structurally important calcifiers, such as corals, CCA, and bivalves (see [32]).
In soft corals, the chlorophytes Ostreobium quekettii and Ulvella endozoica (Goldberg, Makemson, and Colley) R. Nielsen, C.J. O’Kelly, and B. Wysor (2013) are associated with lower tensile strength and elasticity of the endoskeleton [174], and necrosis of the surrounding live tissue in two species of gorgonian corals [182,183]. In brachiopods, the presence of conchocelis stages of the red algae Porphyra or Bangia recorded in the outer layer of the shell [131,169,235] was linked to the potential disruption of growth and shell weakening, as biomineralization processes are disturbed and ‘blisters’ appeared at the areas of shell repair, as in bivalves [169]. Limpet and snail shells are infested by euendolithic cyanobacteria, such as Mastigocoleus testarum or Hyella caespitosa Bornet and Flahault (1888) [143], and green and red algae, such as Phaeophila or Porphyra [94,176,236], and become thinner and more fragile due to bioerosion (similar to bivalves). Infested limpets and snails suffer higher mortality rates, especially in adverse environments (i.e., open coast) [176,237]. In barnacles, the chlorophyte O. quekettii, the cyanobacterium M. testarum, Hyella sp. and Plectonema terebrans, and conchocelis stages of Bangia red algae were found in the plates [44,103,117,238]. Euendolithic infestation starts from the apex of the shell down to the base, weakening the barnacle shell and channeling its energy towards self-repair [238]. In foraminifera, the shell is infested by the same euendolithic species as barnacles, which increases their susceptibility to mechanical breakage and fragmentation and can potentially attract grazers [239]. By contrast, euendoliths at the test surface can promote the formation of a micritic crust around infested tests through secondary carbonate deposition [240], potentially increasing the potential of the test for preservation [241].
While the effects of euendolithic infestation on live calcifying organisms, such as foraminifera, limpets, brachiopods, or barnacles, could be simply underexplored, the sparsity of studies could also be an artifact of lower levels of euendolithic infestation in these organisms, which could translate in less detectable negative effects on the host.
6. Photoautotrophic Euendoliths in the Anthropocene
Marine calcifiers and their future relationship with photoautotrophic euendoliths will be influenced by global climate change (GCC) [32,242,243,244]. GCC is caused by the increase in the emission of anthropogenic greenhouse gases, such as CO2, and manifests as an increase in air and sea surface temperatures (SST), and changes in rainfall regimes [245]. As air temperatures rise, cloud cover is predicted to decrease at the planetary scale, which increases the amount of solar radiation reaching the surface, increasing temperatures even more [245]. For the past 200 years, the world’s oceans have absorbed about one-third of the CO2 released into the atmosphere, which has led to ocean acidification (OA), with a reduction of the pH of surface seawater of 0.1 units [246,247,248]. If global anthropogenic CO2 emissions continue to rise in line with current trends, the average pH of the ocean’s surface could further decrease by 0.7 units in the next millennium, with a rate of change one hundred times greater than in the past 300 M years [247,249]. For marine calcifiers, rising SST, ocean acidification, and the increase in solar radiation will negatively impact calcification, survival, growth, and reproduction, and diminish their resistance to other environmental stressors, such as pollution [30,250,251,252,253]. With decreasing calcification and a weakening of existing calcified structures due to passive dissolution in a more acidic ocean, marine calcifying organisms will become more susceptible to bioerosion [244]. Understanding how photoautotrophic euendoliths will behave in our changing world thus becomes important for understanding the fate of many bioengineered marine ecosystems [32,244].
Euendolithic infestation by photoautotrophs of carbonate substrates, especially those of live calcifying organisms, is expected to increase in prevalence with increased SST, solar radiation, and OA [109,115,129,179,254,255]. Surveys at sites with natural pH gradients, such as volcanic vents [162,254,255], and a series of mesocosm experiments controlling SST and pH [135,179,256,257,258,259,260] confirm this trend in experimental and dead substrates. Photoautotrophic euendoliths are expected to increase in biomass and cover, and to penetrate deeper and faster into the substrate, thus promoting microbioerosion, with a shift in communities under predicted future oceanic conditions of increased SST and reduced pH. The increase in frequency and intensity of marine heat waves in response to GCC will likely be associated with mass mortality events in bivalves [107,252], and coral and CCA reefs [135]. Such mortality events ultimately increase the availability of dead carbonate substrates for photoautotrophic euendoliths. Although manipulative experiments are scarce [259], hypotheses can be formulated on the future of photoautotrophic euendoliths that colonize live marine calcifiers by capitalizing on the knowledge compiled in this review. In a similar fashion to dead substrates, photoautotrophic euendoliths are expected to increase in abundance and biomass in live calcifying organisms with increasing SST and solar radiation. Under OA and GCC, marine calcifiers are more vulnerable to microbioerosion [242] and heat waves [135,252]. With favorable environmental change and the weakening of calcifying organisms, the negative effects of euendolithic infestation [22,24,107,167] are expected to be magnified in the future. In the worst-case scenario, the combined influence of GCC and euendolithic infestation is expected to increase microbioerosion rates by up to 150% in reefs by 2100 [129,156,157,158,179]. This equates to about two-thirds of reef carbonate deposited per year being dissolved by the action of microborers by 2100 [258].
As the negative effects of euendolithic infestation on live calcifying organisms are expected to increase in intensity under future oceanic conditions, so might the beneficial effects. Photoautotrophic euendoliths can contribute to host survival under OA and heat waves. In corals, photoassimilates are translocated directly from the euendoliths to the host until symbiotic zooxanthellae recolonize the coral tissue [23,170,171,210]. However, this mutualistic relationship is limited in the case of rapid heat wave-induced bleaching events, as high light intensities coupled with high temperatures inhibit euendolithic photosynthetic activity [178]. In bivalves, photoautotrophic euendoliths indirectly enhance the albedo of the shell, thus reducing the overall body temperature and the mortality rates experienced by infested bivalves [25,108,115]. The beneficial effects of euendoliths can extend to neighboring mussels, further increasing the thermal buffering provided by mussel beds to associated species on rocky shores [33,115]. In CCA crusts, photoautotrophic euendoliths preferentially remove the highly dissoluble fraction of the carbonate skeleton, thus increasing its resistance to bioerosion, either due to OA or photoautotrophic euendoliths themselves [116,129,134,135]. Both detrimental and beneficial effects of euendolithic infestation in live calcifying organisms are expected to increase in intensity with the ongoing GCC and OA. Nevertheless, in the long term, euendolithic infestation is detrimental to its calcifying hosts, ultimately leading to their death.
7. Conclusions
As discussed, photoautotrophic euendoliths are significant biological components of marine ecosystems. Photoautotrophic euendoliths, which are relatively inaccessible without specific observational techniques, are ubiquitous, found wherever there is a calcium carbonate substrate and suitable environmental conditions for survival. Through their boring activity, they leave long-lasting traces in the substrate that allow their identification. As each species has its own ecological requirements, the presence and abundance of euendolithic species and the composition of euendolithic communities are influenced by abiotic (e.g., light availability, nature of the substrate, water quality) and biotic parameters (e.g., grazers, epibiotic communities). Therefore, euendolithic species or entire communities can be employed as geographical, water quality, or paleobathymetric indicators. Photoautotrophic euendoliths colonize both inanimate calcium carbonate substrates and the calcified structures of live organisms. In live calcifying organisms, endolithic infestation has both detrimental and beneficial effects: while euendoliths bore into the calcified structure, the host’s energy is allocated to secondary structural repair at the expenses of growth, survival, and reproduction; but endolithic infestation can mitigate temperature stress of the host or provide an alternative source of nutrition in the case of corals. However, in the long term, endolithic infestation ultimately leads to the host’s death. In the future, the prevalence and severity of endolithic infestation are predicted to increase, which could lead to higher levels of microbioerosion in marine carbonate substrates, either inanimate or alive, and thus profoundly modify the dynamics of ecosystems founded on marine calcifiers.
Infestation by photoautotrophic euendoliths in live calcifying ecosystem engineers can significantly influence the phenotype, physiology, and biology of individual host specimens, and thus the quality of the habitat they collectively provide (e.g., mussel beds, coral reefs). Research on photoautotrophic euendoliths, especially in live substrates, could beneficiate from the following directions in the future:
- Update and standardize the taxonomy of known photoautotrophic euendolithic species, including morphological descriptions, ecological requirements, and molecular-based approaches to taxonomy;
- Determine the boring mechanisms for euendolithic species, such as red and green algae, and how these processes may be affected by environmental change, including OA and GCC;
- Investigate euendolithic communities (e.g., species composition, abundance, biomass), and the prevalence and severity of euendolithic infestation in less-investigated marine calcifiers (e.g., brachiopods, barnacles, limpets, snails), under different environmental conditions and timescales, and using a combination of approaches;
- Develop a standardized experimental framework, including ex situ and in situ experiments, to evaluate and compare the modalities of euendolithic infestation and how infestation may be influenced by various abiotic and biotic parameters (e.g., nutrient concentrations, light availability, presence of grazers and predators);
- Determine the detrimental and beneficial effects of euendolithic infestation on live marine calcifiers, using manipulative experiments when possible;
- Centralize available information on euendoliths (including heterotrophs) in a publicly accessible database (e.g., Ocean Acidification International Coordination Centre).
When researching marine calcifiers, the presence of euendolithic communities and their effects on host species and ecosystems should be acknowledged and, if possible, quantified. As euendolithic infestation has significant effects on marine calcifiers, failing to consider photoautotrophic euendoliths could lead to biases in the results or their interpretation, even when they are not the main research focus.
Author Contributions
Conceptualization, A.M.D., C.D.M., G.I.Z., K.R.N.; funding acquisition, C.D.M.; project administration, A.M.D.; supervision, C.D.M., G.I.Z., K.R.N., P.W.F.; visualization, A.M.D.; writing—original draft, A.M.D.; writing—review and editing, A.M.D., C.D.M., G.I.Z., K.R.N., P.W.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação para a Ciência e a Tecnologia (FCT–MEC, Portugal, grant number: UIDB/04326/2020, EXPL/BIA-BMA/0682/2021, and the National Research Foundation of South Africa, grant number: 64801).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Writing—review of early draft, Kevin C. K. Ma (ORCID: 0000-0003-1678-077X).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the writing of the manuscript or in the decision to publish.
References
- Carpenter, W. On the Microscopic Structure of Shells. Rep. Br. Assoc. Adv. Sci. 1845, 14, 1–24. [Google Scholar]
- Golubic, S.; Friedmann, I.; Schneider, J. The Lithobiontic Ecological Niche, with Special Reference to Microorganisms. J. Sediment. Petrol. 1981, 51, 475–478. [Google Scholar]
- Zhang, Y.; Golubic, S. Endolithic Microfossils (Cyanophyta) from Early Proterozoic Stromatolites, Hebei, China. Acta Micropalaeontologica Sin. 1987, 4, 1–3. [Google Scholar]
- Schneider, J.; Le Campion-Alsumard, T. Construction and Destruction of Carbonates by Marine and Freshwater Cyanobacteria. Eur. J. Phycol. 1999, 34, 417–426. [Google Scholar] [CrossRef]
- Knoll, A.H. Cyanobacteria and Earth History. In The Cyanobacteria: Molecular Biology, Genomics, and Evolution; Herrero, A., Flores, E., Eds.; Caister Academic Press: Poole, UK, 2008; pp. 1–19. ISBN 978-1-904455-15-8. [Google Scholar]
- Sánchez-Baracaldo, P.; Bianchini, G.; Wilson, J.D.; Knoll, A.H. Cyanobacteria and Biogeochemical Cycles through Earth History. Trends Microbiol. 2022, 30, 143–157. [Google Scholar] [CrossRef]
- Frémy, P. Contribution à La Physiologie Des Thallophytes Marines Perforant et Cariant Les Roches Calcaires et Les Coquilles. Ann. Inst. Océan. 1945, 22, 107–144. [Google Scholar]
- Golubic, S.; Perkins, R.D.; Lukas, K.J. Boring Microorganisms and Microborings in Carbonate Substrates. In The Study of Trace Fossils; Frey, R.W., Ed.; Springer: Berlin/Heidelberg, Germany, 1975; pp. 229–259. ISBN 978-3-642-65925-6. [Google Scholar]
- Friedmann, I.E.; Hua, M.; Ocampo-Friedmann, R. Cryptoendolithic Lichen and Cyanobacterial Communities of the Ross Desert, Antarctica. Polarforschung 1988, 58, 251–259. [Google Scholar] [PubMed]
- Golubic, S.; Schneider, J. Microbial Endoliths as Internal Biofilms. In Fossil and Recent Biofilms: A Natural History of Life on Earth; Krumbein, W.E., Paterson, D.M., Zavarzin, G.A., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 249–263. ISBN 978-90-481-6412-7. [Google Scholar]
- Gaylarde, P.M.; Jungblut, A.-D.; Gaylarde, C.C.; Neilan, B.A. Endolithic Phototrophs from an Active Geothermal Region in New Zealand. Geomicrobiol. J. 2006, 23, 579–587. [Google Scholar] [CrossRef]
- Musso, B. Internal Bioerosion In Situ Living and Dead Corals on the Great Barrier Reef. Ph.D. Thesis, James Cook University, Townsville, Australia, 1994. [Google Scholar]
- Le Campion-Alsumard, T.; Golubic, S.; Hutchings, P. Microbial Endoliths in Skeletons of Live and Dead Corals: Porites Lobata (Moorea, French Polynesia). Mar. Ecol. Prog. Ser. 1995, 117, 149–157. [Google Scholar] [CrossRef]
- Kaehler, S. Incidence and Distribution of Phototrophic Shell-Degrading Endoliths of the Brown Mussel Perna Perna. Mar. Biol. 1999, 135, 505–514. [Google Scholar] [CrossRef]
- Tribollet, A.; Payri, C. Bioerosion of the Coralline Alga Hydrolithon Onkodes by Microborers in the Coral Reefs of Moorea, French Polynesia. Oceanol. Acta 2001, 24, 329–342. [Google Scholar] [CrossRef]
- Ghirardelli, L.A. Endolithic Microorganisms in Live and Dead Thalli of Coralline Red Algae (Corallinales, Rhodophyta) in the Northern Adriatic Sea. Acta Geol. Hisp. 2002, 37, 53–60. [Google Scholar]
- Golubic, S.; Schneider, J. Carbonate Dissolution. Stud. Environ. Sci. 1979, 3, 107–129. [Google Scholar]
- Kiene, W.E.; Hutchings, P.A. Bioerosion Experiments at Lizard Island, Great Barrier Reef. Coral Reefs 1994, 13, 91–98. [Google Scholar] [CrossRef]
- Chazottes, V.; Le Campion-Alsumard, T.; Peyrot-Clausade, M. Bioerosion Rates on Coral Reefs: Interactions between Macroborers, Microborers and Grazers (Moorea, French Polynesia). Palaeogeogr. Palaeoclimatol. Palaeoecol. 1995, 113, 189–198. [Google Scholar] [CrossRef]
- Glaub, I.; Vogel, K.; Gektidis, M. The Role of Modern and Fossil Cyanobacterial Borings in Bioerosion and Bathymetry. Ichnos 2001, 8, 185–195. [Google Scholar] [CrossRef]
- Laukner, G. Diseases of Mollusca: Bivalvia. In Diseases of Marine Animals; Kinne, O., Ed.; Biologische Anstalt Helgoland: Hamburg, Germany, 1983; Volume II, pp. 477–961. [Google Scholar]
- Kaehler, S.; McQuaid, C.D. Lethal and Sub-Lethal Effects of Phototrophic Endoliths Attacking the Shell of the Intertidal Mussel Perna Perna. Mar. Biol. 1999, 135, 497–503. [Google Scholar] [CrossRef]
- Fine, M.; Loya, Y. Endolithic Algae: An Alternative Source of Photoassimilates during Coral Bleaching. Proc. R. Soc. Lond. B Biol. Sci. 2002, 269, 1205–1210. [Google Scholar] [CrossRef] [PubMed]
- Hassenrück, C.; Jantzen, C.; Försterra, G.; Häussermann, V.; Willenz, P. Rates of Apical Septal Extension of Desmophyllum Dianthus: Effect of Association with Endolithic Photo-Autotrophs. Mar. Biol. 2013, 160, 2919–2927. [Google Scholar] [CrossRef]
- Zardi, G.I.; Nicastro, K.R.; McQuaid, C.D.; Ng, T.P.T.; Lathlean, J.; Seuront, L. Enemies with Benefits: Parasitic Endoliths Protect Mussels against Heat Stress. Sci. Rep. 2016, 6, 31413. [Google Scholar] [CrossRef] [PubMed]
- Crooks, J.A.; Khim, H.S. Architectural vs. Biological Effects of a Habitat-Altering, Exotic Mussel, Musculista Senhousia. J. Exp. Mar. Biol. Ecol. 1999, 240, 53–75. [Google Scholar] [CrossRef]
- Knowlton, N.; Brainard, R.E.; Fisher, R.; Moews, M.; Plaisance, L.; Caley, M.J. Coral Reef Biodiversity. In Life in the World’s Oceans; McIntyre, A.D., Ed.; Wiley-Blackwell: Oxford, UK, 2010; pp. 65–78. ISBN 978-1-4443-2550-8. [Google Scholar]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as Ecosystem Engineers. Oikos 1994, 69, 373. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Positive and Negative Effects of Organisms as Physical Ecosystem Engineers. Ecology 1997, 78, 1946–1957. [Google Scholar] [CrossRef]
- Helmuth, B.; Mieszkowska, N.; Moore, P.; Hawkins, S.J. Living on the Edge of Two Changing Worlds: Forecasting the Responses of Rocky Intertidal Ecosystems to Climate Change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 373–404. [Google Scholar] [CrossRef]
- Wild, C.; Hoegh-Guldberg, O.; Naumann, M.S.; Colombo-Pallotta, M.F.; Ateweberhan, M.; Fitt, W.K.; Iglesias-Prieto, R.; Palmer, C.; Bythell, J.C.; Ortiz, J.-C.; et al. Climate Change Impedes Scleractinian Corals as Primary Reef Ecosystem Engineers. Mar. Freshw. Res. 2011, 62, 205. [Google Scholar] [CrossRef]
- Tribollet, A. The Boring Microflora in Modern Coral Reef Ecosystems: A Review of Its Roles. In Current Developments in Bioerosion; Wisshak, M., Tapanila, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 67–94. ISBN 978-3-540-77597-3. [Google Scholar]
- Lourenço, C.R.; Nicastro, K.R.; McQuaid, C.D.; Sabour, B.; Zardi, G.I. Latitudinal Incidence of Phototrophic Shell-Degrading Endoliths and Their Effects on Mussel Bed Microclimates. Mar. Biol. 2017, 164, 129–139. [Google Scholar] [CrossRef]
- Fedor, P.J.; Spellerberg, I.F. Shannon–Wiener Index. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2013; p. 4. ISBN 978-0-12-409548-9. [Google Scholar]
- Bornet, M.E.; Flahault, C. Sur Quelques Plantes Vivant Dans Le Test Calcaire Des Mollusques. Bull. Soc. Bot. Fr. 1889, 36, CXLVII–CLXXVI. [Google Scholar] [CrossRef]
- Kölliker, A.V. On the Frequent Occurrence of Vegetable Parasites in the Hard Structures of Animals. Proc. R. Soc. Lond. 1860, 10, 95–99. [Google Scholar] [CrossRef]
- Wedl, C. On the Significance of the Canals Found in Many Mollusc and Gastropod Shells. Sitzungsberichte Kais. Akad. Wiss. 1859, 33, 451–472. [Google Scholar]
- Gary, M.; McAfee, R.; Wolf, C.L. Glossary of Geology; American Geological Institute: Washington, WA, USA, 1973. [Google Scholar]
- Couradeau, E.; Roush, D.; Guida, B.S.; Garcia-Pichel, F. Diversity and Mineral Substrate Preference in Endolithic Microbial Communities from Marine Intertidal Outcrops (Isla de Mona, Puerto Rico). Biogeosciences 2017, 14, 311–324. [Google Scholar] [CrossRef]
- Amarelle, V.; Carrasco, V.; Fabiano, E. The Hidden Life of Antarctic Rocks. In The Ecological Role of Micro-organisms in the Antarctic Environment; Castro-Sowinski, S., Ed.; Springer Polar Sciences; Springer International Publishing: Cham, Switzerland, 2019; pp. 221–237. ISBN 978-3-030-02785-8. [Google Scholar]
- Ercegović, A. Études Écologiques et Sociologiques Des Cyanophycées Lithophytes de La Côte Yougoslave de l’Adriatique. Bull. Int. Acad. Yougosl. Sci. B-Arts 1932, 26, 33–56. [Google Scholar]
- Wisshak, M. Microbioerosion. In Developments in Sedimentology; Elsevier: Amsterdam, The Netherlands, 2012; Volume 64, pp. 213–243. ISBN 978-0-444-53813-0. [Google Scholar]
- Schroeder, J.H. Calcified Filaments of an Endolithic Alga in Recent Bermuda Reefs. Neues Jahrb. Geol. Palaontol. Mon. 1972, 1972, 16–33. [Google Scholar]
- Drew, K.M. Studies in the Bangioideae. III. The Life-History of Porphyra Umbilicalis (L.) Kütz. Var. Laciniata (Lighf.) J. Ag. A. The Conchocelis-Phase in Culture. Ann. Bot. 1954, XVIII, 184–209. [Google Scholar]
- Drew, K.M. Studies in the Bangiophycidae. IV. The Conchocelis-Phase of Bangia Fuscopurpurea (Dillw.) Lyngbye in Culture. Publ. Sta. Zool. Napoli. 1958, 30, 358–372. [Google Scholar]
- Lagerheim, G. Note Sur Le Mastigocoleus, Nouveau Genre Des Algues Marines de l’Ordre Des Phycochromacées. Notarisia 1886, 1, 65–69. [Google Scholar]
- Golubic, S.; Schneider, J.; Le Campion-Alsumard, T.; Campbell, S.E.; Hook, J.E.; Radtke, G. Approaching Microbial Bioerosion. Facies 2019, 65, 25. [Google Scholar] [CrossRef]
- Al-Thukair, A.A.; Golubic, S.; Rosen, G. New Euendolithic Cyanobacteria from the Bahama Bank and the Arabian Gulf: Hyella Racemus Sp. Nov. 1. J. Phycol. 1994, 30, 764–769. [Google Scholar] [CrossRef]
- Ndhlovu, A.; McQuaid, C.D.; Nicastro, K.R.; Zardi, G.I. Community Succession in Phototrophic Shell-Degrading Endoliths Attacking Intertidal Mussels. J. Molluscan Stud. 2021, 87, eyaa036. [Google Scholar] [CrossRef]
- Rooney, W.S.J.; Perkins, R.D. Distribution and Geologic Significance of Microboring Organisms within Sediments of the Arlington Reef Complex, Australia. Geol. Soc. Am. Bull. 1972, 83, 1139–1150. [Google Scholar] [CrossRef]
- Golubic, S.; Brent, G.; Le Campion, T. Scanning Electron Microscopy of Endolithic Algae and Fungi Using a Multipurpose Casting-embedding Technique. Lethaia 1970, 3, 203–209. [Google Scholar] [CrossRef]
- Chacón, E.; Berrendero, E.; Garcia-Pichel, F. Biogeological Signatures of Microboring Cyanobacterial Communities in Marine Carbonates from Cabo Rojo, Puerto Rico. Sediment. Geol. 2006, 185, 215–228. [Google Scholar] [CrossRef]
- Al-Thukair, A.A. Calculating Boring Rate of Endolithic Cyanobacteria Hyella Immanis under Laboratory Conditions. Int. Biodeterior. Biodegrad. 2011, 65, 664–667. [Google Scholar] [CrossRef]
- Pasella, M.M.; Lee, M.-F.E.; Marcelino, V.R.; Willis, A.; Verbruggen, H. Ten Ostreobium (Ulvophyceae) Strains from Great Barrier Reef Corals as a Resource for Algal Endolith Biology and Genomics. Phycologia 2022, 61, 452–458. [Google Scholar]
- Cunningham, J.A.; Rahman, I.A.; Lautenschlager, S.; Rayfield, E.J.; Donoghue, P.C.J. A Virtual World of Paleontology. Trends Ecol. Evol. 2014, 29, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Sutton, M.D. Tomographic Techniques for the Study of Exceptionally Preserved Fossils. Proc. R. Soc. B Biol. Sci. 2008, 275, 1587–1593. [Google Scholar] [CrossRef] [PubMed]
- Silbiger, N.; Guadayol, Ò.; Thomas, F.; Donahue, M. Reefs Shift from Net Accretion to Net Erosion along a Natural Environmental Gradient. Mar. Ecol. Prog. Ser. 2014, 515, 33–44. [Google Scholar] [CrossRef]
- Wisshak, M.; Titschack, J.; Kahl, W.-A.; Girod, P. Classical and New Bioerosion Trace Fossils in Cretaceous Belemnite Guards Characterised via Micro-CT. Foss. Rec. 2017, 20, 173–199. [Google Scholar] [CrossRef]
- Golubic, S. Distribution, Taxonomy, and Boring Patterns of Marine Endolithic Algae. Am. Zool. 1969, 9, 747–751. [Google Scholar] [CrossRef]
- Verbruggen, H. Morphological Complexity, Plasticity, and Species Diagnosability in the Application of Old Species Names in DNA-Based Taxonomies. J. Phycol. 2014, 50, 26–31. [Google Scholar] [CrossRef]
- Lukas, K.J. Two Species of the Chlorophyte Genus Ostreobium from Skeletons of Atlantic and Caribbean Reef Corals. J. Phycol. 1974, 10, 331–335. [Google Scholar]
- Verbruggen, H.; Ashworth, M.; LoDuca, S.T.; Vlaeminck, C.; Cocquyt, E.; Sauvage, T.; Zechman, F.W.; Littler, D.S.; Littler, M.M.; Leliaert, F. A Multi-Locus Time-Calibrated Phylogeny of the Siphonous Green Algae. Mol. Phylogenet. Evol. 2009, 50, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Gutner-Hoch, E.; Fine, M. Genotypic Diversity and Distribution of Ostreobium Quekettii within Scleractinian Corals. Coral Reefs 2011, 30, 643–650. [Google Scholar] [CrossRef]
- Marcelino, V.R.; Verbruggen, H. Multi-Marker Metabarcoding of Coral Skeletons Reveals a Rich Microbiome and Diverse Evolutionary Origins of Endolithic Algae. Sci. Rep. 2016, 6, 31508. [Google Scholar] [CrossRef]
- Sauvage, T.; Schmidt, W.E.; Suda, S.; Fredericq, S. A Metabarcoding Framework for Facilitated Survey of Endolithic Phototrophs with TufA. BMC Ecol. 2016, 16, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Zapata, F.L.; Gómez-Osorio, S.; Sánchez, J.A. Conspicuous Endolithic Algal Associations in a Mesophotic Reef-Building Coral. Coral Reefs 2018, 37, 705–709. [Google Scholar] [CrossRef]
- Ricci, F.; Fordyce, A.; Leggat, W.; Blackall, L.L.; Ainsworth, T.; Verbruggen, H. Multiple Techniques Point to Oxygenic Phototrophs Dominating the Isopora Palifera Skeletal Microbiome. Coral Reefs 2021, 40, 275–282. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Brochmann, C.; Willerslev, E. Towards Next-Generation Biodiversity Assessment Using DNA Metabarcoding. Mol. Ecol. 2012, 21, 2045–2050. [Google Scholar] [CrossRef]
- Yang, S.-H.; Tandon, K.; Lu, C.-Y.; Wada, N.; Shih, C.-J.; Hsiao, S.S.-Y.; Jane, W.-N.; Lee, T.-C.; Yang, C.-M.; Liu, C.-T.; et al. Metagenomic, Phylogenetic, and Functional Characterization of Predominant Endolithic Green Sulfur Bacteria in the Coral Isopora Palifera. Microbiome 2019, 7, 3. [Google Scholar] [CrossRef]
- Roush, D.; Giraldo-Silva, A.; Garcia-Pichel, F. Cydrasil 3, a Curated 16S rRNA Gene Reference Package and Web App for Cyanobacterial Phylogenetic Placement. Sci. Data 2021, 8, 230. [Google Scholar] [CrossRef]
- Tandon, K.; Pasella, M.M.; Iha, C.; Ricci, F.; Hu, J.; O’Kelly, C.J.; Medina, M.; Kühl, M.; Verbruggen, H. Every Refuge Has Its Price: Ostreobium as a Model for Understanding How Algae Can Live in Rock and Stay in Business. Semin. Cell Dev. Biol. 2022, S1084952122000775. [Google Scholar] [CrossRef]
- Behrendt, L.; Larkum, A.W.; Norman, A.; Qvortrup, K.; Chen, M.; Ralph, P.; Sørensen, S.J.; Trampe, E.; Kühl, M. Endolithic Chlorophyll d-Containing Phototrophs. ISME J. 2011, 5, 1072–1076. [Google Scholar] [CrossRef] [PubMed]
- Massé, A.; Tribollet, A.; Meziane, T.; Bourguet-Kondracki, M.; Yéprémian, C.; Sève, C.; Thiney, N.; Longeon, A.; Couté, A.; Domart-Coulon, I. Functional Diversity of Microboring Ostreobium Algae Isolated from Corals. Environ. Microbiol. 2020, 22, 4825–4846. [Google Scholar] [CrossRef]
- Iha, C.; Dougan, K.E.; Varela, J.A.; Avila, V.; Jackson, C.J.; Bogaert, K.A.; Chen, Y.; Judd, L.M.; Wick, R.; Holt, K.E.; et al. Genomic Adaptations to an Endolithic Lifestyle in the Coral-Associated Alga Ostreobium. Curr. Biol. 2021, 31, 1393–1402. [Google Scholar] [CrossRef]
- Golubic, S.; Seong-Joo, L.; Browne, K.M. Cyanobacteria: Architects of Sedimentary Structures. In Microbial Sediments; Riding, R.E., Awramik, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 57–67. ISBN 978-3-642-08275-7. [Google Scholar]
- Merz-Preiβ, M. Calcification in Cyanobacteria. In Microbial Sediments; Riding, R.E., Awramik, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 50–56. ISBN 978-3-642-08275-7. [Google Scholar]
- Stumm, W.; Morgan, J.J. Aquatic Chemistry. Chemical Equilibria and Rates in Natural Waters, 3rd ed.; Environmental Science and Technology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1996. [Google Scholar]
- Garcia-Pichel, F. Plausible Mechanisms for the Boring on Carbonates by Microbial Phototrophs. Sediment. Geol. 2006, 185, 205–213. [Google Scholar] [CrossRef]
- Alexanderson, T.E. Marks of Unknown Carbonate-Decomposing Organelles in Cyanophyte Borings. Nature 1975, 254, 236–238. [Google Scholar] [CrossRef]
- Hatch, W.I. The Implication of Carbonic Anhydrase in the Physiological Mechanism of Penetration of Carbonate Substrata by the Marine Burrowing Sponge Cliona Celata (Demospongiae). Biol. Bull. 1980, 159, 135–147. [Google Scholar] [CrossRef]
- Cockell, C.S.; Herrera, A. Why Are Some Microorganisms Boring? Trends Microbiol. 2008, 16, 101–106. [Google Scholar] [CrossRef]
- Garcia-Pichel, F.; Ramirez-Reinat, E.; Gao, Q. Microbial Excavation of Solid Carbonates Powered by P-Type ATPase-Mediated Transcellular Ca2+ Transport. Proc. Natl. Acad. Sci. USA 2010, 107, 21749–21754. [Google Scholar] [CrossRef] [PubMed]
- Stal, L.J.; Krumbein, W.E. Temporal Separation of Nitrogen Fixation and Photosynthesis in the Filamentous, Non-Heterocystous Cyanobacterium Oscillatoria Sp. Arch. Microbiol. 1987, 149, 76–80. [Google Scholar] [CrossRef]
- Guida, B.S.; Garcia-Pichel, F. Extreme Cellular Adaptations and Cell Differentiation Required by a Cyanobacterium for Carbonate Excavation. Proc. Natl. Acad. Sci. USA 2016, 113, 5712–5717. [Google Scholar] [CrossRef]
- Fredd, C.N.; Fogler, H.S. The Influence of Chelating Agents on the Kinetics of Calcite Dissolution. J. Colloid Interface Sci. 1998, 204, 187–197. [Google Scholar] [CrossRef]
- Ramírez-Reinat, E.L.; Garcia-Pichel, F. Prevalence of Ca2+-ATPase-Mediated Carbonate Dissolution among Cyanobacterial Euendoliths. Appl. Environ. Microbiol. 2012, 78, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Krause, S.; Liebetrau, V.; Nehrke, G.; Damm, T.; Büsse, S.; Leipe, T.; Vogts, A.; Gorb, S.N.; Eisenhauer, A. Endolithic Algae Affect Modern Coral Carbonate Morphology and Chemistry. Front. Earth Sci. 2019, 7, 304. [Google Scholar] [CrossRef]
- Guida, B.S.; Bose, M.; Garcia-Pichel, F. Carbon Fixation from Mineral Carbonates. Nat. Commun. 2017, 8, 1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinfelder, J.R. Carbon Concentrating Mechanisms in Eukaryotic Marine Phytoplankton. Annu. Rev. Mar. Sci. 2011, 3, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Guida, B.S. Unique Cellular, Physiological, and Metabolic Adaptations to the Euendolithic Lifestyle in a Boring Cyanobacterium. Ph.D. Thesis, Arizona State University, Tempe, AZ, USA, 2016. [Google Scholar]
- Ascaso, C.; Wierzchosb, J.; Castelloa, R. Study of the Biogenic Weathering of Calcareous Litharenite Stones Caused by Lichen and Endolithic Microorganisms. Int. Biodeterior. Biodegrad. 1998, 42, 29–38. [Google Scholar] [CrossRef]
- Huber, J.; Jadin, F. Sur Une Algue Perforante d’Eau Douce. C R. Hebd. Séances L’Académie Sci. Paris 1892, 115, 262–264. [Google Scholar]
- Akpan, E.B. Bioerosion of Oyster Shells in Brackish Modern Mangrove Swamps, Nigeria. Ichnos 1990, 1, 125–132. [Google Scholar] [CrossRef]
- Cerrano, C.; Bavestrello, G.; Calcinai, B.; Cattaneo-Vietti, R.; Chiantore, M.; Guidetti, M.; Sarà, A. Bioerosive Processes in Antarctic Seas. Polar Biol. 2001, 24, 790–792. [Google Scholar] [CrossRef]
- Pantazidou, A.; Louvrou, I.; Economou-Amilli, A. Euendolithic Shell-Boring Cyanobacteria and Chlorophytes from the Saline Lagoon Ahivadolimni on Milos Island, Greece. Eur. J. Phycol. 2006, 41, 189–200. [Google Scholar] [CrossRef]
- Ćurin, M.; Peharda, M.; Calcinai, B.; Golubić, S. Incidence of Damaging Endolith Infestation of the Edible Mytilid Bivalve Modiolus Barbatus. Mar. Biol. Res. 2014, 10, 179–189. [Google Scholar] [CrossRef]
- Akpan, E.B.; Farrow, G.E. Shell-Boring Algae on the Scottish Continental Shelf: Identification, Distribution, Bathymetric Zonation. Trans. R. Soc. Edinb. Earth Sci. 1984, 75, 1–12. [Google Scholar] [CrossRef]
- Försterra, G.; Beuck, L.; Häussermann, V.; Freiwald, A. Shallow-Water Desmophyllum Dianthus (Scleractinia) from Chile: Characteristics of the Biocoenoses, the Bioeroding Community, Heterotrophic Interactions and (Paleo)-Bathymetric Implications. In Cold-Water Corals and Ecosystems; Freiwald, A., Roberts, J.M., Eds.; Erlangen Earth Conference Series; Springer-Verlag: Berlin/Heidelberg, Germany, 2005; pp. 937–977. ISBN 978-3-540-24136-2. [Google Scholar]
- Wilkinson, M.; Burrows, E.M. The Distribution of Marine Shell-Boring Green Algae. J. Mar. Biol. Assoc. UK 1972, 52, 59–65. [Google Scholar] [CrossRef]
- Mao Che, L.; Le Campion-Alsumard, T.; Boury-Esnault, N.; Payri, C.; Golubic, S.; Bézac, C. Biodegradation of Shells of the Black Pearl Oyster, Pinctada Margaritifera Var. Cumingii, by Microborers and Sponges of French Polynesia. Mar. Biol. 1996, 126, 509–519. [Google Scholar] [CrossRef]
- Perry, C.T. Grain Susceptibility to the Effects of Microboring: Implications for the Preservation of Skeletal Carbonates. Sedimentology 1998, 45, 39–51. [Google Scholar] [CrossRef]
- Tribollet, A. Dissolution of Dead Corals by Euendolithic Microorganisms Across the Northern Great Barrier Reef (Australia). Microb. Ecol. 2008, 55, 569–580. [Google Scholar] [CrossRef]
- Meyer, N.; Wisshak, M.; Freiwald, A. Ichnodiversity and Bathymetric Range of Microbioerosion Traces in Polar Barnacles of Svalbard. Polar Res. 2020, 39, 3766. [Google Scholar] [CrossRef]
- Campbell, S.E. The Modern Distribution and Geological History of Calcium Carbonate Boring Microorganisms. In Biomineralization and Biological Metal Accumulation; Westbroek, P., de Jong, E.W., Eds.; Springer: Dordrecht, The Netherlands, 1983; pp. 99–104. ISBN 978-94-009-7946-8. [Google Scholar]
- Gektidis, M. Development of Microbial Euendolithic Communities: The Influence of Light and Time. Bull. Geol. Soc. Den. 1999, 45, 147–150. [Google Scholar] [CrossRef]
- Tribollet, A.; Golubic, S.; Radtke, G.; Reitner, J. On Microbiocorrosion. In Advances in Stromatolite Geobiology; Lecture Notes in Earth Sciences; Springer: Berlin/Heidelberg, Germany, 2011; Volume 131, pp. 265–276. ISBN 978-3-642-10414-5. [Google Scholar]
- Zardi, G.I.; Nicastro, K.R.; McQuaid, C.D.; Gektidis, M. Effects of Endolithic Parasitism on Invasive and Indigenous Mussels in a Variable Physical Environment. PLoS ONE 2009, 4, e6560. [Google Scholar] [CrossRef]
- Gehman, A.M.; Harley, C.D.G. Symbiotic Endolithic Microbes Alter Host Morphology and Reduce Host Vulnerability to High Environmental Temperatures. Ecosphere 2019, 10, e02683. [Google Scholar] [CrossRef]
- Monsinjon, J.R.; McQuaid, C.D.; Nicastro, K.R.; Seuront, L.; Oróstica, M.H.; Zardi, G.I. Weather and Topography Regulate the Benefit of a Conditionally Helpful Parasite. Funct. Ecol. 2021, 35, 2691–2706. [Google Scholar] [CrossRef]
- Gektidis, M.; Dubinsky, Z.; Goffredo, S. Microendoliths of the Shallow Euphotic Zone in Open and Shaded Habitats at 30°N–Eilat, Israel – Paleoecological Implications. Facies 2007, 53, 43–55. [Google Scholar] [CrossRef]
- Wisshak, M.; Tribollet, A.; Golubic, S.; Jakobsen, J.; Freiwald, A. Temperate Bioerosion: Ichnodiversity and Biodiversity from Intertidal to Bathyal Depths (Azores). Geobiology 2011, 9, 492–520. [Google Scholar] [CrossRef]
- Le Bris, S.; Le Campion-Alsumard, T.; Romano, J.-C. Caractéristiques du feutrage algal des récifs coralliens de Polynésie française soumis à différentes intensités de bioérosion. Oceanol. Acta 1998, 21, 695–708. [Google Scholar] [CrossRef]
- Tribollet, A.; Golubic, S. Cross-Shelf Differences in the Pattern and Pace of Bioerosion of Experimental Carbonate Substrates Exposed for 3 Years on the Northern Great Barrier Reef, Australia. Coral Reefs 2005, 24, 422–434. [Google Scholar] [CrossRef]
- Wisshak, M.; Gektidis, M.; Freiwald, A.; Lundälv, T. Bioerosion along a Bathymetric Gradient in a Cold-Temperate Setting (Kosterfjord, SW Sweden): An Experimental Study. Facies 2005, 51, 93–117. [Google Scholar] [CrossRef]
- Zardi, G.I.; Monsinjon, J.R.; McQuaid, C.D.; Seuront, L.; Orostica, M.; Want, A.; Firth, L.B.; Nicastro, K.R. Foul-weather Friends: Modelling Thermal Stress Mitigation by Symbiotic Endolithic Microbes in a Changing Environment. Glob. Change Biol. 2021, 27, 2549–2560. [Google Scholar] [CrossRef]
- Ramírez-Reinat, E.L.; Garcia-Pichel, F. Characterization of a Marine Cyanobacterium That Bores into Carbonates and the Redescription of the Genus Mastigocoleus. J. Phycol. 2012, 48, 740–749. [Google Scholar] [CrossRef]
- Meyer, N.; Wisshak, M.; Freiwald, A. Bioerosion Ichnodiversity in Barnacles from the Ross Sea, Antarctica. Polar Biol. 2021, 44, 667–682. [Google Scholar] [CrossRef]
- Hutchings, P.A. Biological Destruction of Coral Reefs. Coral Reefs 1986, 4, 239–252. [Google Scholar] [CrossRef]
- Chazottes, V.; Le Campion-Alsumard, T.; Peyrot-Clausade, M.; Cuet, P. The Effects of Eutrophication-Related Alterations to Coral Reef Communities on Agents and Rates of Bioerosion (Reunion Island, Indian Ocean). Coral Reefs 2002, 21, 375–390. [Google Scholar] [CrossRef]
- Chazottes, V.; Cabioch, G.; Golubic, S.; Radtke, G. Bathymetric Zonation of Modern Microborers in Dead Coral Substrates from New Caledonia—Implications for Paleodepth Reconstructions in Holocene Corals. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 280, 456–468. [Google Scholar] [CrossRef]
- Bentis, C.; Kaufman, L.; Golubic, S. Endolithic Fungi in Reef-Building Corals (Order: Scleractinia) Are Common, Cosmopolitan, and Potentially Pathogenic. Biol. Bull. 2000, 198, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Vogel, K.; Gektidis, M.; Golubic, S.; Kiene, W.E.; Radtke, G. Experimental Studies on Microbial Bioerosion at Lee Stocking Island, Bahamas and One Tree Island, Great Barrier Reef, Australia: Implications for Paleoecological Reconstructions. Lethaia 2000, 33, 190–204. [Google Scholar] [CrossRef]
- Radtke, G.; Golubic, S. Microborings in Mollusk Shells, Bay of Safaga, Egypt: Morphometry and Ichnology. Facies 2005, 51, 118–134. [Google Scholar] [CrossRef]
- Golubic, S.; Campbell, S.E.; Drobne, K.; Cameron, B.; Balsam, W.L.; Cimerman, F.; Duboiss, L. Microbial Endoliths: A Benthic Overprint in the Sedimentary Record, and a Paleobathymetric Cross-Reference with Foraminifera. J. Paleontol. 1984, 58, 12. [Google Scholar]
- Kiene, W.E.; Radtke, G.; Gektidis, M.; Golubic, S.; Vogel, K. Factors Controlling the Distribution of Microborers in Bahamian Reef Environments. Facies 1995, 32, 176–188. [Google Scholar]
- Le Campion-Alsumard, T.; Campbell, S.E.; Golubic, S. Endoliths and the Depth of the Photic Zone: Discussion. J. Sediment. Petrol. 1982, 52, 1333–13338. [Google Scholar] [CrossRef]
- Lukas, K.J. Depth Distribution and Form among Common Microboring Algae from the Florida Continental Shelf. In Proceedings of the Abstract with Programs; Boulder: Toronto, ON, Canada, 1978; Volume 10, pp. 1–448. [Google Scholar]
- Försterra, G.; Häussermann, V. Unusual Symbiotic Relationships between Microendolithic Phototrophic Organisms and Azooxanthellate Cold-Water Corals from Chilean Fjords. Mar. Ecol. Prog. Ser. 2008, 370, 121–125. [Google Scholar] [CrossRef]
- Reyes-Nivia, C.; Diaz-Pulido, G.; Dove, S. Relative Roles of Endolithic Algae and Carbonate Chemistry Variability in the Skeletal Dissolution of Crustose Coralline Algae. Biogeosciences Discuss. 2014, 11, 2993–3021. [Google Scholar] [CrossRef]
- Tribollet, A.; Veinott, G.; Golubic, S.; Dart, R. Infestation of the North American Freshwater Mussel Elliptio Complanata (Head Lake, Canada) by the Euendolithic Cyanobacterium Plectonema Terebrans Bornet et Flahault. Algol. Stud. 2008, 128, 65–77. [Google Scholar] [CrossRef]
- Gaspard, D. Endolithic Algae, Fungi and Bacterial Activity in Holocene and Cretaceous Brachiopod Shells—Diagenetic Consequences. Mem. Assoc. Australas. Palaeontol. 2011, 41, 327–337. [Google Scholar]
- Lukas, K.J. Taxonomy and Ecology of the Endolithic Microflora of Reef Corals with a Review of the Literature on Endolithic Microphytes. Ph.D. Thesis, University of Rhode Island, Kingston, RI, USA, 1973. [Google Scholar]
- Perkins, R.D.; Halsey, S.D. Geologic Significance of Microboring Fungi and Algae in Carolina Shelf Sediments. J. Sediment. Res. 1971, 41, 843–853. [Google Scholar] [CrossRef]
- Nash, M.C.; Opdyke, B.N.; Troitzsch, U.; Russell, B.D.; Adey, W.H.; Kato, A.; Diaz-Pulido, G.; Brent, C.; Gardner, M.; Prichard, J.; et al. Dolomite-Rich Coralline Algae in Reefs Resist Dissolution in Acidified Conditions. Nat. Clim. Chang. 2013, 3, 268–272. [Google Scholar] [CrossRef]
- Diaz-Pulido, G.; Nash, M.C.; Anthony, K.R.N.; Bender, D.; Opdyke, B.N.; Reyes-Nivia, C.; Troitzsch, U. Greenhouse Conditions Induce Mineralogical Changes and Dolomite Accumulation in Coralline Algae on Tropical Reefs. Nat. Commun. 2014, 5, 3310. [Google Scholar] [CrossRef] [PubMed]
- Golubic, S.; Radtke, G.; Campion-Alsumard, T.L. Endolithic Fungi in Marine Ecosystems. Trends Microbiol. 2005, 13, 229–235. [Google Scholar] [CrossRef]
- Gutiérrez-Isaza, N.; Espinoza-Avalos, J.; León-Tejera, H.P.; González-Solís, D. Endolithic Community Composition of Orbicella Faveolata (Scleractinia) underneath the Interface between Coral Tissue and Turf Algae. Coral Reefs 2015, 34, 625–630. [Google Scholar] [CrossRef]
- Keats, D.W.; Groener, A.; Chamberlain, Y.M. Cell Sloughing in the Littoral Zone Coralline Alga, Spongites Yendoi (Foslie) Chamberlain (Corallinales, Rhodophyta). Phycologia 1993, 32, 143–150. [Google Scholar] [CrossRef]
- Owen, G.; Williams, A. The Caecum of Articulate Brachiopoda. Proc. R. Soc. Lond. B Biol. Sci. 1969, 172, 187–201. [Google Scholar]
- Scardino, A.; De Nys, R.; Ison, O.; O’Connor, W.; Steinberg, P. Microtopography and Antifouling Properties of the Shell Surface of the Bivalve Molluscs Mytilus Galloprovincialis and Pinctada Imbricata. Biofouling 2003, 19, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Scardino, A.; de Nys, R. Fouling Deterrence on the Bivalve Shell Mytilus Galloprovincialis: A Physical Phenomenon? Biofouling 2004, 20, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Bers, A.V.; Díaz, E.R.; da Gama, B.A.P.; Vieira-Silva, F.; Dobretsov, S.; Valdivia, N.; Thiel, M.; Scardino, A.J.; McQuaid, C.D.; Sudgen, H.E.; et al. Relevance of Mytilid Shell Microtopographies for Fouling Defence—A Global Comparison. Biofouling 2010, 26, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Prusina, I.; Peharda, M.; Ezgeta-Balic, D.; Puljas, S.; Glamuzina, B.; Golubic, S. Life-History Trait of the Mediterranean Keystone Species Patella Rustica: Growth and Microbial Bioerosion. Mediterr. Mar. Sci. 2015, 16, 393. [Google Scholar] [CrossRef]
- Odum, H.T.; Odum, E.P. Trophic Structure and Productivity of a Windward Coral Reef Community on Eniwetok Atoll. Ecol. Monogr. 1955, 25, 291–320. [Google Scholar] [CrossRef]
- Fine, M.; Roff, G.; Ainsworth, T.D.; Hoegh-Guldberg, O. Phototrophic Microendoliths Bloom during Coral “White Syndrome”. Coral Reefs 2006, 25, 577–581. [Google Scholar] [CrossRef]
- Grange, J.S.; Rybarczyk, H.; Tribollet, A. The Three Steps of the Carbonate Biogenic Dissolution Process by Microborers in Coral Reefs (New Caledonia). Environ. Sci. Pollut. Res. 2015, 22, 13625–13637. [Google Scholar] [CrossRef]
- Le Campion-Alsumard, T. Les Cyanophycées endolithes marines. Systématique, ultrastructure, écologie et biodestruction. Oceanol. Acta 1979, 2, 143–156. [Google Scholar]
- Tribollet, A.; Golubic, S. Reef Bioerosion: Agents and Processes. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 435–450. ISBN 978-94-007-0113-7. [Google Scholar]
- Fordyce, A.J.; Ainsworth, T.D.; Leggat, W. Microalgae, a Boring Bivalve and a Coral—A Newly Described Association Between Two Coral Reef Bioeroders Within Their Coral Host. Integr. Org. Biol. 2020, 2, obaa035. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Torunski, H. Biokarst on Limestone Coasts, Morphogenesis and Sediment Production. Mar. Ecol. 1983, 4, 45–63. [Google Scholar] [CrossRef]
- Nicholson, G.M.; Clements, K.D. Resolving Resource Partitioning in Parrotfishes (Scarini) Using Microhistology of Feeding Substrata. Coral Reefs 2020, 39, 1313–1327. [Google Scholar] [CrossRef]
- Zubia, M.; Peyrot-Clausade, M. Internal Bioerosion of Acropora Formosa in Réunion (Indian Ocean): Microborer and Macroborer Activities. Oceanol. Acta 2001, 24, 251–262. [Google Scholar] [CrossRef]
- Rice, M.M.; Maher, R.L.; Correa, A.M.S.; Moeller, H.V.; Lemoine, N.P.; Shantz, A.A.; Burkepile, D.E.; Silbiger, N.J. Macroborer Presence on Corals Increases with Nutrient Input and Promotes Parrotfish Bioerosion. Coral Reefs 2020, 39, 409–418. [Google Scholar] [CrossRef]
- Pari, N.; Peyrot-Clausade, M.; Le Campion-Alsumard, T.; Hutchings, P.; Chazottes, V.; Golubic, S.; Le Campion, J.; Fontaine, M. Bioerosion of Experimental Substrates on High Islands and on Atoll Lagoons (French Polynesia) after Two Years of Exposure. Mar. Ecol. Prog. Ser. 1998, 166, 119–130. [Google Scholar] [CrossRef]
- Pari, N.; Peyrot-Clausade, M.; Hutchings, P.A. Bioerosion of Experimental Substrates on High Islands and Atoll Lagoons (French Polynesia) during 5 Years of Exposure. J. Exp. Mar. Biol. Ecol. 2002, 276, 109–127. [Google Scholar] [CrossRef]
- Carreiro-Silva, M.; McClanahan, T.R.; Kiene, W.E. The Role of Inorganic Nutrients and Herbivory in Controlling Microbioerosion of Carbonate Substratum. Coral Reefs 2005, 24, 214–221. [Google Scholar] [CrossRef]
- Carreiro-Silva, M.; McClanahan, T.; Kiene, W. Effects of Inorganic Nutrients and Organic Matter on Microbial Euendolithic Community Composition and Microbioerosion Rates. Mar. Ecol. Prog. Ser. 2009, 392, 1–15. [Google Scholar] [CrossRef]
- Carreiro-Silva, M.; Kiene, W.E.; Golubic, S.; McClanahan, T.R. Phosphorus and Nitrogen Effects on Microbial Euendolithic Communities and Their Bioerosion Rates. Mar. Pollut. Bull. 2012, 64, 602–613. [Google Scholar] [CrossRef]
- Godinot, C.; Tribollet, A.; Grover, R.; Ferrier-Pagès, C. Bioerosion by Euendoliths Decreases in Phosphate-Enriched Skeletons of Living Corals. Biogeosciences Discuss. 2012, 9, 2425–2444. [Google Scholar] [CrossRef]
- del Campo, J.; Pombert, J.-F.; Šlapeta, J.; Larkum, A.; Keeling, P.J. The ‘Other’ Coral Symbiont: Ostreobium Diversity and Distribution. ISME J. 2017, 11, 296–299. [Google Scholar] [CrossRef]
- Al-Thukair, A.A. Effect of Oil Pollution on Euendolithic Cyanobacteria of the Arabian Gulf. Environ. Microbiol. 2002, 4, 125–129. [Google Scholar] [CrossRef]
- Cherchi, A.; Buosi, C.; Zuddas, P.; De Giudici, G. Bioerosion by Microbial Euendoliths in Benthic Foraminifera from Heavy Metal-Polluted Coastal Environments of Portovesme (South-Western Sardinia, Italy). Biogeosciences 2012, 9, 4607–4620. [Google Scholar] [CrossRef]
- Cherchi, A.; Da Pelo, S.; Ibba, A.; Mana, D.; Buosi, C.; Floris, N. Benthic Foraminifera Response and Geochemical Characterization of the Coastal Environment Surrounding the Polluted Industrial Area of Portovesme (South-Western Sardinia, Italy). Mar. Pollut. Bull. 2009, 59, 281–296. [Google Scholar] [CrossRef]
- Cabioch, G.; Montaggioni, L.F.; Faure, G.; Ribaud-Laurenti, A. Reef Coralgal Assemblages as Recorders of Paleobathymetry and Sea Level Changes in the Indo-Pacific Province. Quat. Sci. Rev. 1999, 18, 1681–1695. [Google Scholar] [CrossRef]
- Perry, C.T.; Macdonald, I.A. Impacts of Light Penetration on the Bathymetry of Reef Microboring Communities: Implications for the Development of Microendolithic Trace Assemblages. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 186, 101–113. [Google Scholar] [CrossRef]
- Swinchatt, J.P. Algal Boring: A Possible Depth Indicator in Carbonate Rocks and Sediments. Geol. Soc. Am. Bull. 1969, 80, 1391. [Google Scholar] [CrossRef]
- Massé, A.; Domart-Coulon, I.; Golubic, S.; Duché, D.; Tribollet, A. Early Skeletal Colonization of the Coral Holobiont by the Microboring Ulvophyceae Ostreobium Sp. Sci. Rep. 2018, 8, 2293. [Google Scholar] [CrossRef] [PubMed]
- Ndhlovu, A.; McQuaid, C.D.; Monaco, C.J. Ectoparasites Reduce Scope for Growth in a Rocky-Shore Mussel (Perna Perna) by Raising Maintenance Costs. Sci. Total Environ. 2021, 753, 142020. [Google Scholar] [CrossRef]
- Curry, G.B. Microborings in Recent Brachiopods and the Functions of Caeca. Lethaia 1983, 16, 119–127. [Google Scholar] [CrossRef]
- Schlichter, D.; Zscharnack, B.; Krisch, H. Transfer of Photoassimilates from Endolithic Algae to Coral Tissue. Naturwissenschaften 1995, 82, 1–564. [Google Scholar] [CrossRef]
- Schlichter, D.; Kampmann, H.; Conrady, S. Trophic Potential and Photoecology of Endolithic Algae Living within Coral Skeletons. Mar. Ecol. 1997, 18, 299–317. [Google Scholar] [CrossRef]
- Marquet, N.; Nicastro, K.R.; Gektidis, M.; McQuaid, C.D.; Pearson, G.A.; Serrão, E.A.; Zardi, G.I. Comparison of Phototrophic Shell-Degrading Endoliths in Invasive and Native Populations of the Intertidal Mussel Mytilus Galloprovincialis. Biol. Invasions 2013, 15, 1253–1272. [Google Scholar] [CrossRef]
- Ndhlovu, A.; McQuaid, C.D.; Nicastro, K.R.; Zardi, G.I. Parasitism by Endolithic Cyanobacteria Reduces Reproductive Output and Attachment Strength of Intertidal Ecosystem Engineers. Mar. Biol. 2022, 169, 37. [Google Scholar] [CrossRef]
- Goldberg, W.M.; Makemson, J.C.; Colley, S.B. Entocladia Endozoica Sp. Nov., a Pathogenic Chlorophyte: Structure, Life History, Physiology, and Effect on Its Coral Host. Biol. Bull. 1984, 166, 368–383. [Google Scholar] [CrossRef]
- Nicastro, K.R.; McQuaid, C.D.; Zardi, G.I. Between a Rock and a Hard Place: Combined Effect of Trampling and Phototrophic Shell-Degrading Endoliths in Marine Intertidal Mussels. Mar. Biodivers. 2019, 49, 1581–1586. [Google Scholar] [CrossRef]
- Nolan, C.P. Size, Shape and Shell Morphology in the Antarctic Limpet Nacella Concinna at Signy Island, South Orkney Islands. J. Molluscan Stud. 1991, 57, 225–238. [Google Scholar] [CrossRef]
- Fine, M.; Steindler, L.; Loya, Y. Endolithic Algae Photoacclimate to Increased Irradiance during Coral Bleaching. Mar. Freshw. Res. 2004, 55, 115. [Google Scholar] [CrossRef]
- Fine, M.; Meroz-Fine, E.; Hoegh-Guldberg, O. Tolerance of Endolithic Algae to Elevated Temperature and Light in the Coral Montipora Monasteriata from the Southern Great Barrier Reef. J. Exp. Biol. 2005, 208, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Nivia, C.; Diaz-Pulido, G.; Kline, D.; Guldberg, O.-H.; Dove, S. Ocean Acidification and Warming Scenarios Increase Microbioerosion of Coral Skeletons. Glob. Change Biol. 2013, 19, 1919–1929. [Google Scholar] [CrossRef] [PubMed]
- Risk, M.J.; Sammarco, P.W.; Edinger, E.N. Bioerosion in Acropora across the Continental Shelf of the Great Barrier Reef. Coral Reefs 1995, 14, 79–86. [Google Scholar] [CrossRef]
- Le Campion-Alsumard, T.; Golubic, S.; Priess, K. Fungi in Corals: Symbiosis or Disease? Interaction between Polyps and Fungi Causes Pearl-like Skeleton Biomineralization. Mar. Ecol. Prog. Ser. 1995, 117, 137–147. [Google Scholar] [CrossRef]
- Morse, D.E.; Morse, A.; Duncan, H.; Trench, R.K. Algal Tumors in the Caribbean Octocorallian, Gorgonia Ventalina: II. Biochemical Characterization of the Algae, and First Epidemiological Observations. Bull. Mar. Sci. 1981, 31, 399–409. [Google Scholar]
- Morse, D.E.; Morse, A.; Duncan, H. Algal Tumors in the Caribeean Sea-Fan, Gorgonia Ventalina. In Proceeding of Third International Coral Reef Symposium; Taylor, D.L., Ed.; Rosenstiel School of Marine and Atmospheric Science: Miami, FL, USA, 1977; Volume 1, pp. 623–629. [Google Scholar]
- Nicastro, K.R.; Seuront, L.; McQuaid, C.D.; Zardi, G.I. Symbiont-Induced Intraspecific Phenotypic Variation Enhances Plastic Trapping and Ingestion in Biogenic Habitats. Sci. Total Environ. 2022, 826, 153922. [Google Scholar] [CrossRef] [PubMed]
- Stolarski, J. Gardineria—A Scleractinian Living Fossil. Acta Palaeontol. Pol. 1996, 41, 339–367. [Google Scholar]
- Brown, B.E.; Tudhope, A.W.; Le Tissier, M.D.A.; Scoffin, T.P. A Novel Mechanism for Iron Incorporation into Coral Skeletons. Coral Reefs 1991, 10, 211–215. [Google Scholar] [CrossRef]
- Lazier, A.V.; Smith, J.E.; Risk, M.J.; Schwarcz, H.P. The Skeletal Structure of Desmophyllum Cristagalli: The Use of Deep-Water Corals in Sclerochronology. Lethaia 1999, 32, 119–130. [Google Scholar] [CrossRef]
- Steneck, R.S. The Ecology of Coralline Algal Crusts: Convergent Patterns and Adaptative Strategies. Annu. Rev. Ecol. Syst. 1986, 17, 3–303. [Google Scholar] [CrossRef]
- Laborel, J.; Le Campion-Alsumard, T. Infestation Massive Du Squelette de Coraux Vivants Par Des Rhodophycées de Type Conchocelis. Comptes Rendus Acad. Sci. Paris 1979, 288, 1575–1577. [Google Scholar]
- Ralph, P.J.; Larkum, A.W.D.; Kühl, M. Photobiology of Endolithic Microorganisms in Living Coral Skeletons: 1. Pigmentation, Spectral Reflectance and Variable Chlorophyll Fluorescence Analysis of Endoliths in the Massive Corals Cyphastrea Serailia, Porites Lutea and Goniastrea Australensis. Mar. Biol. 2007, 152, 395–404. [Google Scholar] [CrossRef]
- Ghirardelli, L.A. An Endolithic Cyanophyte in the Cell Wall of Calcareous Algae. Bot. Mar. 1998, 41, 367–373. [Google Scholar] [CrossRef]
- Duerden, J.E. Boring Algae as Agents in the Disintegration of Corals. Bull. Am. Mus. Nat. Hist. 1902, 16, 323–332. [Google Scholar]
- Highsmith, R.C. Lime-Boring Algae in Hermatypic Coral Skeletons. J. Exp. Mar. Biol. Ecol. 1981, 55, 267–281. [Google Scholar] [CrossRef]
- Delvoye, L. Endolithic Algae in Living Stony Corals: Algal Concentrations under Influence of Depth-Dependent Light Conditions and Coral Tissue Fluorescence in Agaricia agaricites (L.) and Meandrina meandrites (L.) (Scleractinia, Anthozoa). Stud. Nat. Hist. Caribb. Reg. 1992, 71, 24–41. [Google Scholar]
- Tribollet, A.; Pica, D.; Puce, S.; Radtke, G.; Campbell, S.E.; Golubic, S. Euendolithic Conchocelis Stage (Bangiales, Rhodophyta) in the Skeletons of Live Stylasterid Reef Corals. Mar. Biodivers. 2018, 48, 1855–1862. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.T. Calcification in Hermatypic and Ahermatypic Corals. Science 1996, 271, 637–639. [Google Scholar] [CrossRef]
- Aeby, G.S.; Williams, G.J.; Franklin, E.C.; Haapkyla, J.; Harvell, C.D.; Neale, S.; Page, C.A.; Raymundo, L.; Vargas-Ángel, B.; Willis, B.L.; et al. Growth Anomalies on the Coral Genera Acropora and Porites Are Strongly Associated with Host Density and Human Population Size across the Indo-Pacific. PLoS ONE 2011, 6, e16887. [Google Scholar] [CrossRef] [PubMed]
- McClanahan, T.R.; Weil, E.; Maina, J. Strong Relationship between Coral Bleaching and Growth Anomalies in Massive Porites. Glob. Chang. Biol. 2009, 15, 1804–1816. [Google Scholar] [CrossRef]
- Miller, A.; Blackwelder, P.; Al-Sayegh, H.; Richardson, L. Fine-Structural Analysis of Black Band Disease-Infected Coral Reveals Boring Cyanobacteria and Novel Bacteria. Dis. Aquat. Organ. 2011, 93, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Kanwisher, J.W.; Wainwright, S.A. Oxygen Balance in Some Reef Corals. Biol. Bull. 1967, 133, 378–390. [Google Scholar] [CrossRef]
- Halldal, P. Photosynthetic Capacities and Photosynthetic Action Spectra of Endozoic Algae of the Massive Coral Favia. Biol. Bull. 1968, 134, 411–424. [Google Scholar] [CrossRef]
- Magnusson, S.; Fine, M.; Kühl, M. Light Microclimate of Endolithic Phototrophs in the Scleractinian Corals Montipora Monasteriata and Porites Cylindrica. Mar. Ecol. Prog. Ser. 2007, 332, 119–128. [Google Scholar] [CrossRef]
- Shibata, K.; Haxo, F.T. Light Transmission and Spectral Distribution through Epi- and Endozoic Algal Layers in the Brain Coral, Favia. Biol. Bull. 1969, 136, 461–468. [Google Scholar] [CrossRef]
- Shashar, N.; Stambler, N. Endolithic Algae within Corals—Life in an Extreme Environment. J. Exp. Mar. Biol. Ecol. 1992, 163, 277–286. [Google Scholar] [CrossRef]
- Galindo-Martínez, C.T.; Weber, M.; Avila-Magaña, V.; Enríquez, S.; Kitano, H.; Medina, M.; Iglesias-Prieto, R. The Role of the Endolithic Alga Ostreobium Spp. during Coral Bleaching Recovery. Sci. Rep. 2022, 12, 2977. [Google Scholar] [CrossRef] [PubMed]
- Schlichter, D. Cnidaria: Permability, Epidermal Transport and Related Phenoma. In Biology of the Integument; Bereiter-Hahn, J., Matoltsy, A.G., Richards, K.S., Eds.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 79–95. ISBN 978-3-642-51595-8. [Google Scholar]
- Kühl, M.; Holst, G.; Larkum, A.W.D.; Ralph, P.J. Imaging of Oxygen Dynamics within the Endolithic Algal Community of the Massive Coral Porites Lobata. J. Phycol. 2008, 44, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Titlyanov, E.A.; Kiyashko, S.I.; Titlyanova, T.V.; Kalita, T.L.; Raven, J.A. δ13C and δ15N Values in Reef Corals Porites Lutea and P. Cylindrica and in Their Epilithic and Endolithic Algae. Mar. Biol. 2008, 155, 353–361. [Google Scholar] [CrossRef]
- Fine, M.; Zibrowius, H.; Loya, Y. Oculina Patagonica: A Non-Lessepsian Scleractinian Coral Invading the Mediterranean Sea. Mar. Biol. 2001, 138, 1195–1203. [Google Scholar] [CrossRef]
- Nothdurft, L.D.; Webb, G.E. Earliest Diagenesis in Scleractinian Coral Skeletons: Implications for Palaeoclimate-Sensitive Geochemical Archives. Facies 2009, 55, 161–201. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M.L. The Effects of Storms and Cyclones on Coral Reefs: A Review. J. Coast. Res. Spec. Issue 1994, 12, 211–231. [Google Scholar]
- Littler, M.M.; Littler, D.S. Impact of CLOD Pathogen on Pacific Coral Reefs. Science 1995, 267, 1356–1360. [Google Scholar] [CrossRef]
- Harrington, L.; Fabricius, K.; De’ath, G.; Negri, A. Recognition and Selection of Settlement Substrata Determine Post-Settlement Survival in Corals. Ecology 2004, 85, 3428–3437. [Google Scholar] [CrossRef]
- Hayakawa, J.; Kawamura, T.; Ohashi, S.; Horii, T.; Watanabe, Y. Habitat Selection of Japanese Top Shell (Turbo Cornutus) on Articulated Coralline Algae: Combination of Preferences in Settlement and Post-Settlement Stage. J. Exp. Mar. Biol. Ecol. 2008, 363, 118–123. [Google Scholar] [CrossRef]
- Scoffin, T.P.; Alexandersson, E.T.; Bowes, G.E.; Clokie, J.J.; Farrow, G.E.; Milliman, J.D. Recent, Temperate, Sub-Photic, Carbonate Sedimentation: Rockall Bank, Northeast Atlantic. J. Sediment. Petrol. 1980, 50, 331–356. [Google Scholar]
- Eakin, C.M. Where Have All the Carbonates Gone? A Model Comparison of Calcium Carbonate Budgets before and after the 1982-1983 El Niño at Uva Island in the Eastern Pacific. Coral Reefs 1996, 15, 109–119. [Google Scholar]
- Glynn, P.W. State of Coral Reefs in the Galápagos Islands: Natural vs Anthropogenic Impacts. Mar. Pollut. Bull. 1994, 29, 131–140. [Google Scholar] [CrossRef]
- Raghukumar, C.; Sharma, S.; Lande, V. Distribution and Biomass Estimation of Shell-Boring Algae in the Intertidal at Goa, India. Phycologia 1991, 30, 303–309. [Google Scholar] [CrossRef]
- Alfaro, A.C.; Webb, S.C.; Barnaby, C. Variability of Growth, Health, and Population Turnover within Mussel Beds of Perna Canaliculus in Northern New Zealand. Mar. Biol. Res. 2008, 4, 376–383. [Google Scholar] [CrossRef]
- Ndhlovu, A.; McQuaid, C.D.; Nicastro, K.; Marquet, N.; Gektidis, M.; Monaco, C.J.; Zardi, G. Biogeographical Patterns of Endolithic Infestation in an Invasive and an Indigenous Intertidal Marine Ecosystem Engineer. Diversity 2019, 11, 75. [Google Scholar] [CrossRef] [Green Version]
- Gardner, J.; Thomas, M. Growth, Mortality and Production of Organic Matter by a Rocky Intertidal Population of Mytilus Edulis in the Quoddy Region of the Bay of Fundy. Mar. Ecol. Prog. Ser. 1987, 39, 31–36. [Google Scholar] [CrossRef]
- Griffiths, C.L.; King, J.A. Energy Expended on Growth and Gonad Output in the Ribbed Mussel Aulacomya Ater. Mar. Biol. 1979, 53, 217–222. [Google Scholar] [CrossRef]
- Wahl, M. Ecological Lever and Interface Ecology: Epibiosis Modulates the Interactions between Host and Environment. Biofouling 2008, 24, 427–438. [Google Scholar] [CrossRef]
- Thieltges, D.W.; Buschbaum, C. Vicious Circle in the Intertidal: Facilitation between Barnacle Epibionts, a Shell Boring Polychaete and Trematode Parasites in the Periwinkle Littorina Littorea. J. Exp. Mar. Biol. Ecol. 2007, 340, 90–95. [Google Scholar] [CrossRef]
- Gutiérrez, J.L.; Jones, C.G.; Strayer, D.L.; Iribarne, O.O. Mollusks as Ecosystem Engineers: The Role of Shell Production in Aquatic Habitats. Oikos 2003, 101, 79–90. [Google Scholar] [CrossRef]
- Witman, J.D. Refuges, Biological Disturbance, and Rocky Subtidal Community Structure in New England. Ecol. Monogr. 1985, 55, 421–445. [Google Scholar] [CrossRef]
- Suchanek, T.H. Mussels and Their Role in Structuring Rocky Shore Communities. In The Ecology of Rocky Coasts; Moore, P.J., Seed, R., Eds.; Colombia Univ. Press: New York, NY, USA, 1986; pp. 70–96. [Google Scholar]
- Stephens, E.G.; Bertness, M.D. Mussel Facilitation of Barnacle Survival in a Sheltered Bay Habitat. J. Exp. Mar. Biol. Ecol. 1991, 145, 33–48. [Google Scholar] [CrossRef]
- Jurgens, L.J.; Gaylord, B. Physical Effects of Habitat-Forming Species Override Latitudinal Trends in Temperature. Ecol. Lett. 2018, 21, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Thiel, M.; Dernedde, T. Recruitment of Shore Crabs Carcinus Maenas on Tidal Flats: Mussel Clumps as an Important Refuge for Juveniles. Helgoländer Meeresunters. 1994, 48, 321–332. [Google Scholar] [CrossRef]
- Tsuchiya, M.; Nishihira, M. Islands of Mytilus as a Habitat for Small Intertidal Animals: Effect of Island Size on Community Structure. Mar. Ecol. Prog. Ser. 1985, 25, 71–81. [Google Scholar] [CrossRef]
- Piazza, B.P.; Banks, P.D.; La Peyre, M.K. The Potential for Created Oyster Shell Reefs as a Sustainable Shoreline Protection Strategy in Louisiana. Restor. Ecol. 2005, 13, 499–506. [Google Scholar] [CrossRef]
- Borsje, B.W.; van Wesenbeeck, B.K.; Dekker, F.; Paalvast, P.; Bouma, T.J.; van Katwijk, M.M.; de Vries, M.B. How Ecological Engineering Can Serve in Coastal Protection. Ecol. Eng. 2011, 37, 113–122. [Google Scholar] [CrossRef]
- Peck, L.S.; Clarke, A.; Holmes, L.J. Size, Shape and the Distribution of Organic Matter in the Recent Antarctic Brachiopod Liothyrella Uva. Lethaia 1987, 20, 33–40. [Google Scholar] [CrossRef]
- Walker, S.E. Endobionts on Modern and Fossil Turritella from the Northern Gulf of California Region. Ichnos 1998, 6, 99–115. [Google Scholar] [CrossRef]
- Day, E.G.; Branch, G.M.; Viljoen, C. How Costly Is Molluscan Shell Erosion? A Comparison of Two Patellid Limpets with Contrasting Shell Structures. J. Exp. Mar. Biol. Ecol. 2000, 243, 185–208. [Google Scholar] [CrossRef]
- Parke, M.W.; Moore, H.B. The Biology of Balanus Balanoides. II. Algal Infection of the Shell. J. Mar. Biol. Assoc. UK 1935, 20, 49–56. [Google Scholar] [CrossRef]
- Peebles, M.W.; Lewis, R.D. Differential Infestation of Shallow-Water Benthic Foraminifera by Microboring Organisms: Possible Biases in Preservation Potential. PALAIOS 1988, 3, 345. [Google Scholar] [CrossRef]
- Kobluk, D.R.; Risk, M.J. Calcification of Exposed Filaments of Endolithic Algae, Micrite Envelope Formation and Sediment Production. SEPM J. Sediment. Res. 1977, 47, 517–528. [Google Scholar] [CrossRef]
- Bathurst, R.G.C. Boring Algae, Micrite Envelopes and Lithification of Molluscan Biosparites. Geol. J. 1966, 5, 15–32. [Google Scholar] [CrossRef]
- Raven, J.A. Ocean Acidification Due to Increasing Atmospheric Carbon Dioxide; The Royal Society: London, UK, 2005. [Google Scholar]
- Andersson, A.J. Coastal Ocean and Carbonate Systems in the High CO2 World of the Anthropocene. Am. J. Sci. 2005, 305, 875–918. [Google Scholar] [CrossRef]
- Schönberg, C.H.L.; Fang, J.K.H.; Carreiro-Silva, M.; Tribollet, A.; Wisshak, M. Bioerosion: The Other Ocean Acidification Problem. ICES J. Mar. Sci. 2017, 74, 895–925. [Google Scholar] [CrossRef]
- Mendoza, V.; Pazos, M.; Garduño, R.; Mendoza, B. Thermodynamics of Climate Change between Cloud Cover, Atmospheric Temperature and Humidity. Sci. Rep. 2021, 11, 21244. [Google Scholar] [CrossRef]
- Gruber, N.; Sarmiento, J.L.; Stocker, T.F. An Improved Method for Detecting Anthropogenic CO2 in the Oceans. Glob. Biogeochem. Cycles 1996, 10, 809–837. [Google Scholar] [CrossRef]
- Caldeira, K.; Wickett, M.E. Anthropogenic Carbon and Ocean PH. Nature 2003, 425, 365. [Google Scholar] [CrossRef] [PubMed]
- Sabine, C.L.; Feely, R.A.; Gruber, N.; Key, R.M.; Lee, K.; Bullister, J.L.; Wanninkhof, R.; Wong, C.S.; Wallace, D.W.R.; Tilbrook, B.; et al. The Oceanic Sink for Anthropogenic CO2. Science 2004, 305, 367–371. [Google Scholar] [CrossRef]
- Key, R.M.; Kozyr, A.; Sabine, C.L.; Lee, K.; Wanninkhof, R.; Bullister, J.L.; Feely, R.A.; Millero, F.J.; Mordy, C.; Peng, T.-H. A Global Ocean Carbon Climatology: Results from Global Data Analysis Project (GLODAP). Glob. Biogeochem. Cycles 2004, 18, GB4031. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O. Climate Change, Coral Bleaching and the Future of the World’s Coral Reefs. Mar. Freshw. Res. 1999, 50, 839–866. [Google Scholar] [CrossRef]
- Gattuso, J.-P.; Allemand, D.; Frankignoulle, M. Photosynthesis and Calcification at Cellular, Organismal and Community Levels in Coral Reefs: A Review on Interactions and Control by Carbonate Chemistry. Am. Zool. 1999, 39, 160–183. [Google Scholar] [CrossRef]
- Petes, L.E.; Menge, B.A.; Murphy, G.D. Environmental Stress Decreases Survival, Growth, and Reproduction in New Zealand Mussels. J. Exp. Mar. Biol. Ecol. 2007, 351, 83–91. [Google Scholar] [CrossRef]
- O’Donnell, M.J.; George, M.N.; Carrington, E. Mussel Byssus Attachment Weakened by Ocean Acidification. Nat. Clim. Chang. 2013, 3, 587–590. [Google Scholar] [CrossRef]
- Enochs, I.C.; Manzello, D.P.; Tribollet, A.; Valentino, L.; Kolodziej, G.; Donham, E.M.; Fitchett, M.D.; Carlton, R.; Price, N.N. Elevated Colonization of Microborers at a Volcanically Acidified Coral Reef. PLoS ONE 2016, 11, e0159818. [Google Scholar] [CrossRef]
- Marcelino, V.R.; Morrow, K.M.; Oppen, M.J.H.; Bourne, D.G.; Verbruggen, H. Diversity and Stability of Coral Endolithic Microbial Communities at a Naturally High pCO2 Reef. Mol. Ecol. 2017, 26, 5344–5357. [Google Scholar] [CrossRef]
- Tribollet, A.; Atkinson, M.J.; Christopher, L. Effects of Elevated pCO2 on Epilithic and Endolithic Metabolism of Reef Carbonates. Glob. Chang. Biol. 2006, 12, 2200–2208. [Google Scholar] [CrossRef]
- Tribollet, A.; Godinot, C.; Atkinson, M.; Langdon, C. Effects of Elevated pCO2 on Dissolution of Coral Carbonates by Microbial Euendoliths. Glob. Biogeochem. Cycles 2009, 23, 3008. [Google Scholar] [CrossRef]
- Tribollet, A.; Chauvin, A.; Cuet, P. Carbonate Dissolution by Reef Microbial Borers: A Biogeological Process Producing Alkalinity under Different pCO2 Conditions. Facies 2019, 65, 9. [Google Scholar] [CrossRef]
- Diaz-Pulido, G.; Anthony, K.R.N.; Kline, D.I.; Dove, S.; Hoegh-Guldberg, O. Interactions between Ocean Acidification and Warming on the Mortality and Dissolution of Coralline Algae. J. Phycol. 2012, 48, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Leggat, W.P.; Camp, E.F.; Suggett, D.J.; Heron, S.F.; Fordyce, A.J.; Gardner, S.; Deakin, L.; Turner, M.; Beeching, L.J.; Kuzhiumparambil, U.; et al. Rapid Coral Decay Is Associated with Marine Heatwave Mortality Events on Reefs. Curr. Biol. 2019, 29, 2723–2730.e4. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Types of endolithic organisms in relation to a hard rocky substrate (modified from [40]).
Figure 1.
Types of endolithic organisms in relation to a hard rocky substrate (modified from [40]).

Figure 3.
Signs of infestation by euendolithic microborers in live marine calcifiers. (a) Non-infested Perna perna bed. (b) Perna perna mussels showing extremely eroded shells due to the action of euendolithic cyanobacteria. (c) Bleached coral with evident Ostreobium bloom (indicated by white arrows). (d) Cross-section of Paragoniastrea australensis coral showing Ostreobium that inhabits the skeleton. The inset shows Ostreobium filaments after skeletal decalcification. Copyright: © Alexia DIEVART (a,b), © Alexander Fordyce (c). Images (c,d) from [74].
Figure 3.
Signs of infestation by euendolithic microborers in live marine calcifiers. (a) Non-infested Perna perna bed. (b) Perna perna mussels showing extremely eroded shells due to the action of euendolithic cyanobacteria. (c) Bleached coral with evident Ostreobium bloom (indicated by white arrows). (d) Cross-section of Paragoniastrea australensis coral showing Ostreobium that inhabits the skeleton. The inset shows Ostreobium filaments after skeletal decalcification. Copyright: © Alexia DIEVART (a,b), © Alexander Fordyce (c). Images (c,d) from [74].


{kind=link}
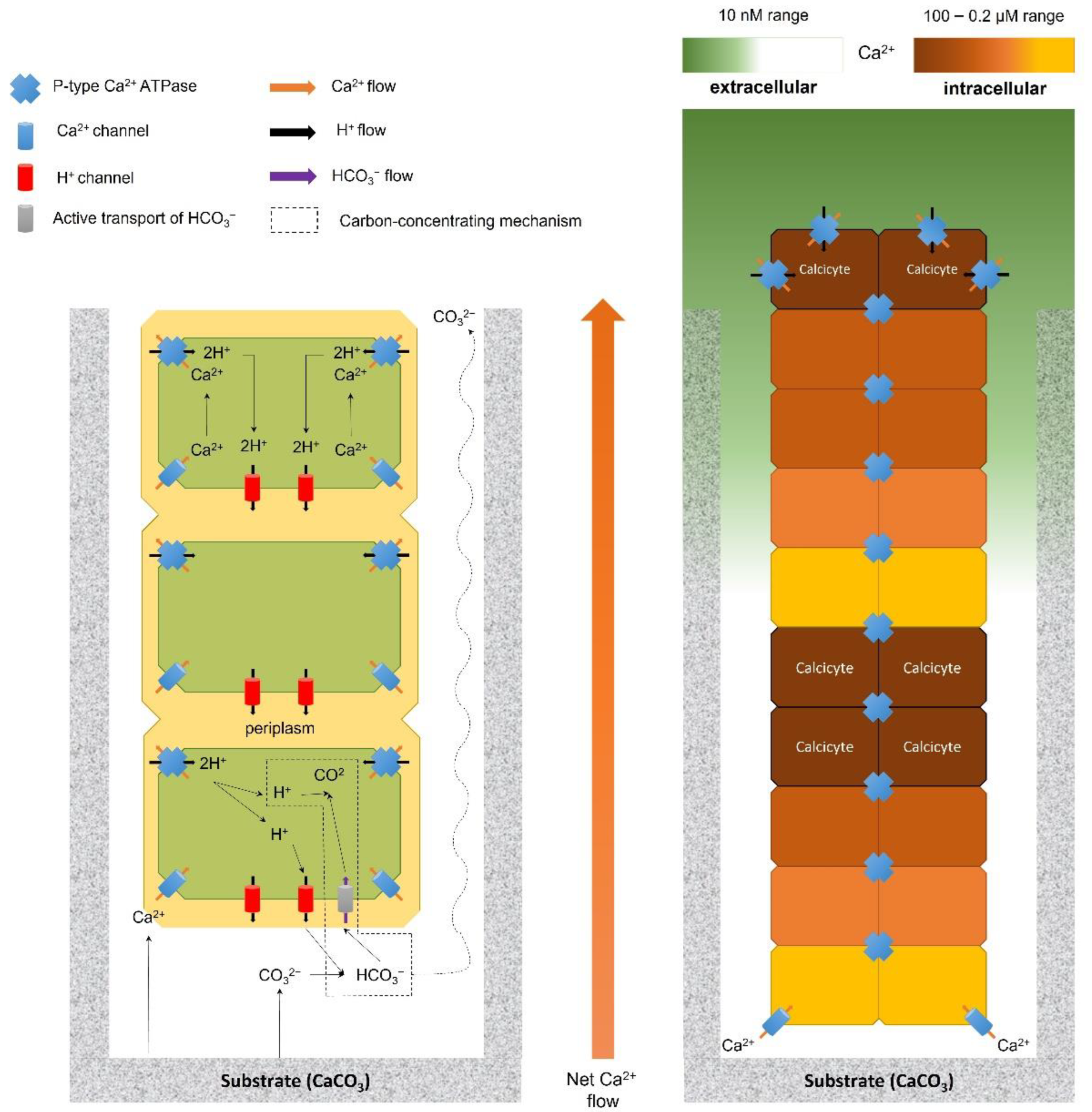{kind=link}
{kind=link}
Table 1.
Summary of the negative and positive effects observed and suspected (underlined) of euendolithic infestation on the physiological parameters, calcified structures, biological interactions, and bioengineering qualities of main live calcifying hosts.
Table 1.
Summary of the negative and positive effects observed and suspected (underlined) of euendolithic infestation on the physiological parameters, calcified structures, biological interactions, and bioengineering qualities of main live calcifying hosts.
| Responses to Endolithic Infestation | Live Calcifying Hosts | References | |||
|---|---|---|---|---|---|
| Corals | Coralline Algae | Bivalves | Others | ||
| Physiological Parameters | |||||
| Growth | ↓ = | ↓ | ↓ | [22,24,143,167,168,169] | |
| General condition | = | ↓ | [22,96,107,167,170,171,172] | ||
| Reproduction | ↓ | = | [22,168,173,174] | ||
| Attachment strength | ↓ | [107,172,173] | |||
| General survival | ↑ = | ↓ ✞ | ↓ | [14,22,73,143,144,170,171,173,175,176] | |
| Individual survival to heat stress | ↑ (lim) | ↑ (lim) | [23,25,33,108,109,115,177,178] | ||
| Calcified structures | |||||
| Microbioerosion | ↑ | ↑↓ | ↑ | ↑ | [13,15,22,108,129,143,169,179] |
| Thickness | ↑ | ↓ ✞ | ↓ | [22,24,98,143] | |
| Strength | ↓ | ↓ ✞ | ↓ | [22,107,143,169,172,174,175,176,180] | |
| Porosity | ↑ | ↑ | ↑ | [13,15,24,108] | |
| Deformations | ↑ | ↑ ✞ | ↑ | [14,24,96,174,181,182,183] | |
| Maintenance costs | ↑ | ↑ | ↑ | [22,24,96,98,143,168,181] | |
| Mineralogy | ~ | ~ | [96,135] | ||
| Biological interactions | |||||
| Epibionts | ↑ | [172] | |||
| Predators | ↑ | ↑ | [107] | ||
| Grazers | ↑ | ↑ | ↑ | [151,176] | |
| Photoautotrophic euendoliths | ↔ | ↔ | ↔ | [15,24,108,144,170,171] | |
| Bioengineered ecosystems | |||||
| Architectural complexity | ↑↓ | ↓ | [129,135] | ||
| Coastal protection from waves and other stressors | ↓ | ↑↓ | ↓ | ↓ | [107,129,135,174,175,179] |
| Mitigation of environmental stressors for associated species | ↑ | [33,109,115] | |||
| Resistance to anthropogenic stressors | ↓ | [175,184] | |||
Symbols for effects: (=)—no effect; (↑)—positive effect, reinforcement; (↓)—negative effect, reduction; (↓↑)—variable responses depending on the host species and/or environmental conditions; (~)—alteration in the composition of the parameter; (✞)—mortality observed in the host species; (↔)—mutualistic relationship; (lim)—effect observed during unusual harsh environmental conditions (i.e., heatwaves). Please note that some effects presented in this table are based on observations of a single species or different life cycle stages of the host species.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dievart, A.M.; McQuaid, C.D.; Zardi, G.I.; Nicastro, K.R.; Froneman, P.W. Photoautotrophic Euendoliths and Their Complex Ecological Effects in Marine Bioengineered Ecosystems. Diversity 2022, 14, 737. https://doi.org/10.3390/d14090737
AMA Style
Dievart AM, McQuaid CD, Zardi GI, Nicastro KR, Froneman PW. Photoautotrophic Euendoliths and Their Complex Ecological Effects in Marine Bioengineered Ecosystems. Diversity. 2022; 14(9):737. https://doi.org/10.3390/d14090737
Chicago/Turabian StyleDievart, Alexia M., Christopher D. McQuaid, Gerardo I. Zardi, Katy R. Nicastro, and Pierre W. Froneman. 2022. "Photoautotrophic Euendoliths and Their Complex Ecological Effects in Marine Bioengineered Ecosystems" Diversity 14, no. 9: 737. https://doi.org/10.3390/d14090737
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.