
Genome Insight and Description of Previously Uncultured N2-Fixing Bacterium Rhizobium terricola sp. nov., Isolated from Forest Rhizospheric Soil by Using Modified Culture Method



Abstract
:1. Introduction
2. Material and Methods
2.1. Isolation and Ecology
2.2. 16S rRNA Phylogeny
2.3. Genome Features
2.4. Physiology and Chemotaxonomy
3. Results and Discussion
3.1. Isolation of Uncultured Strain
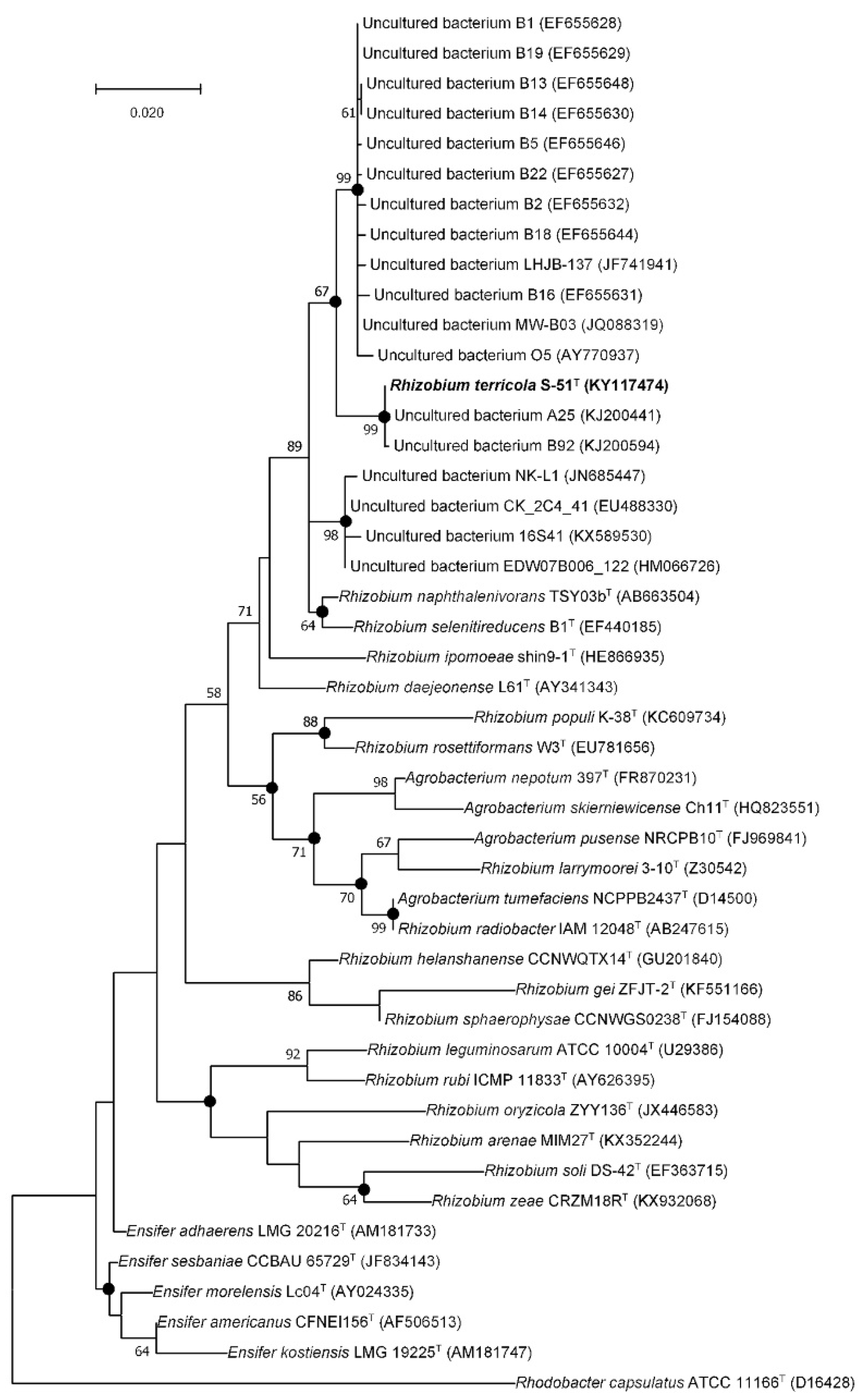
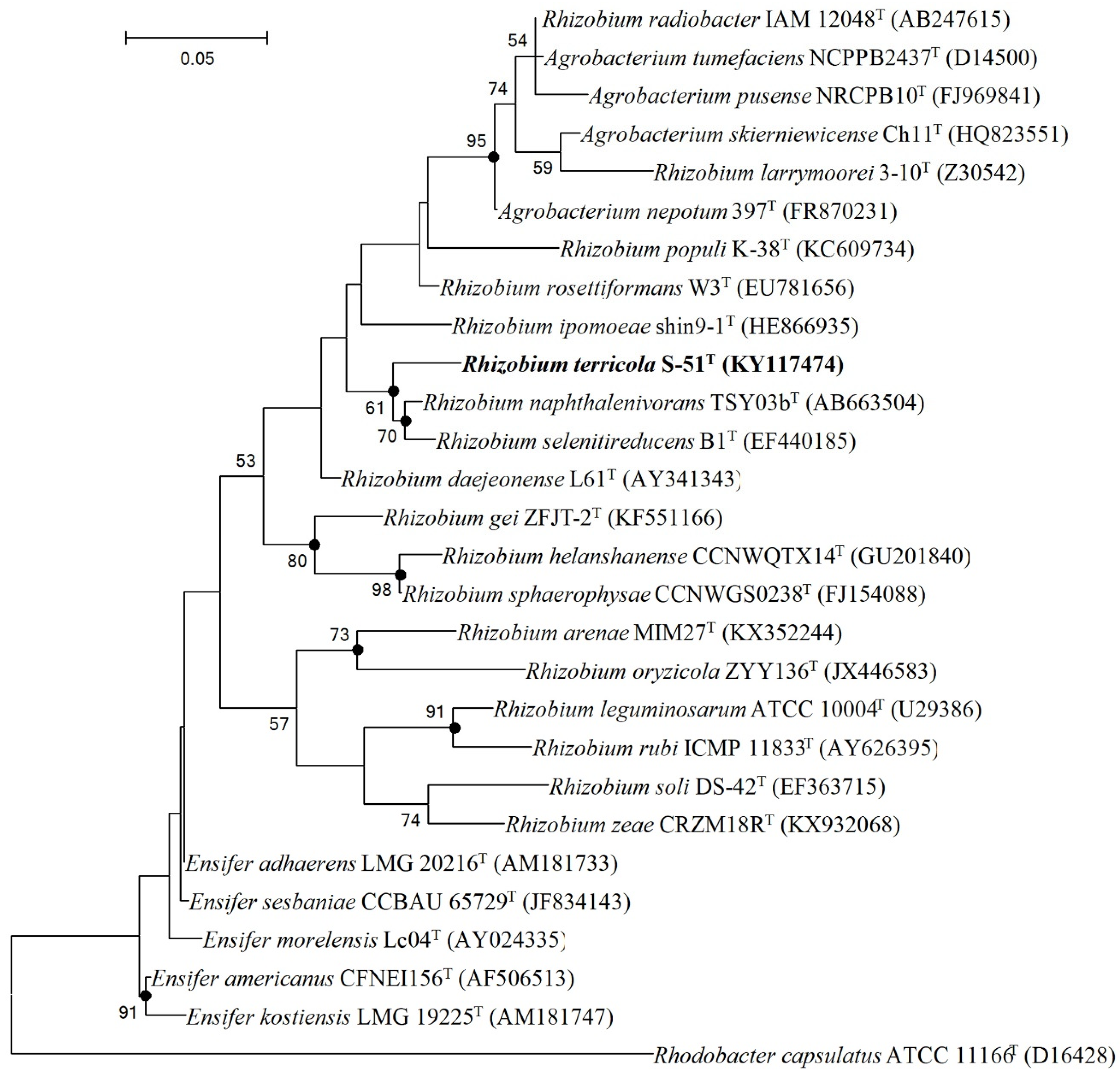
3.2. Phylogenetic Analysis
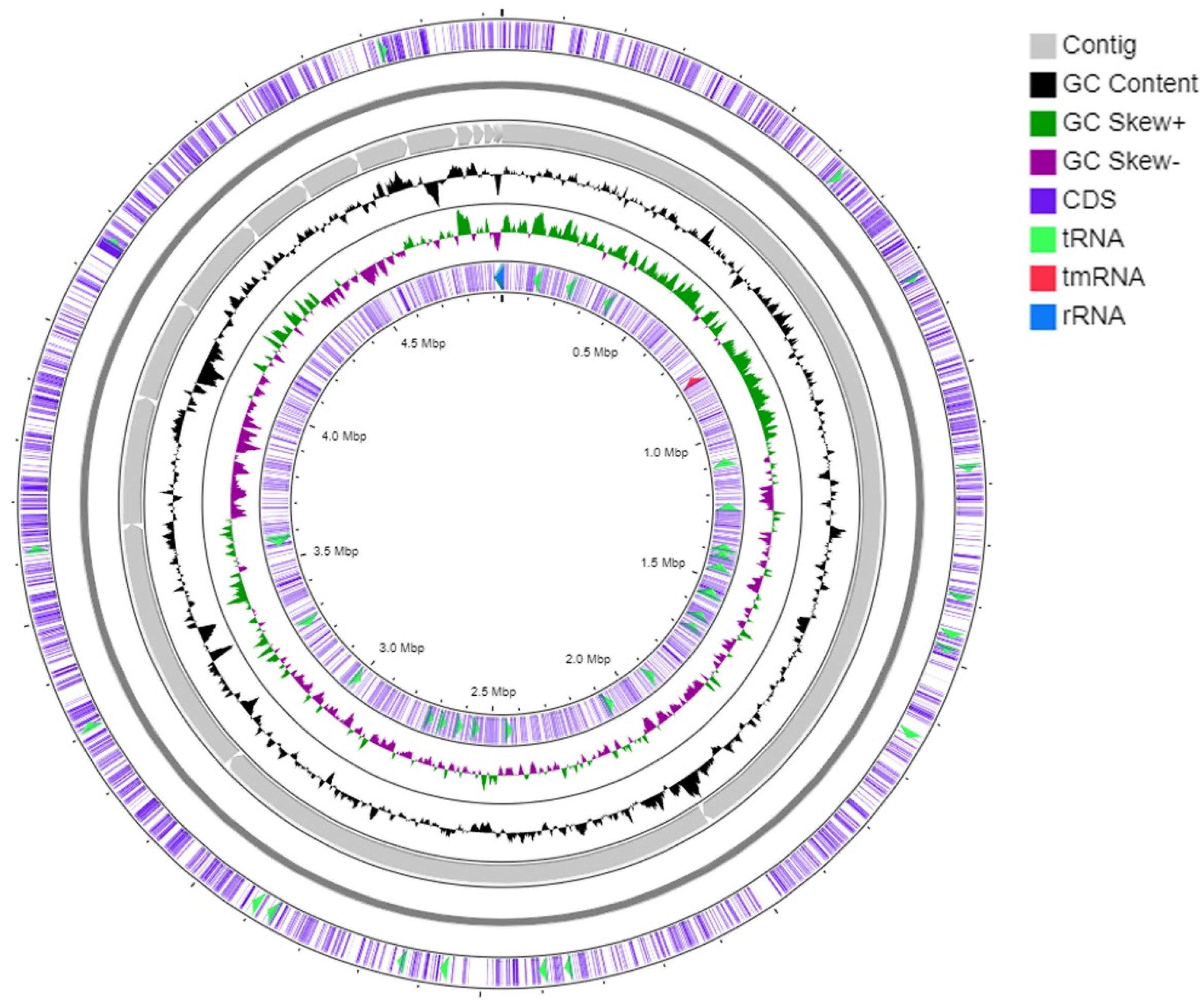
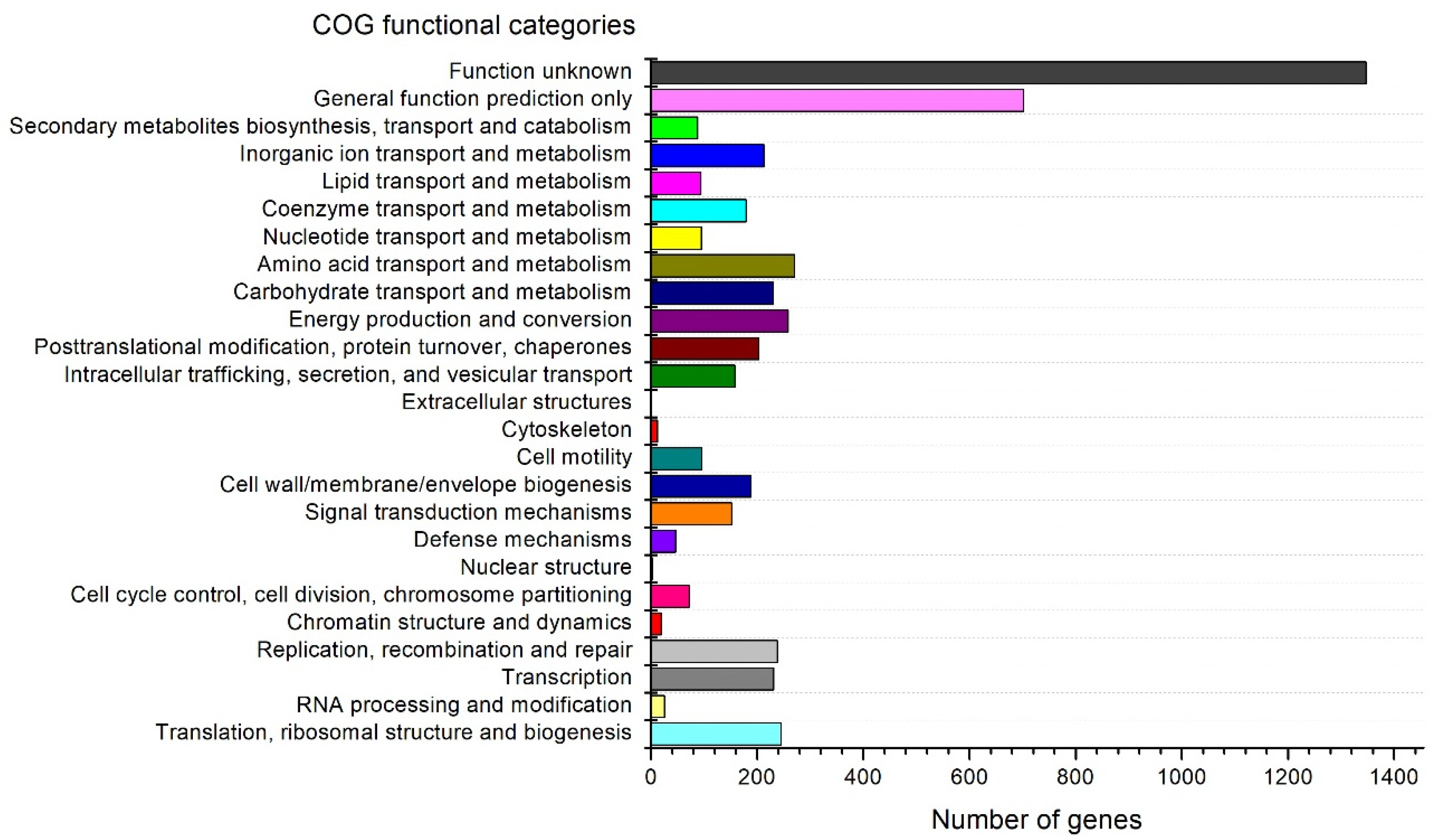
3.3. Genome Analysis
3.4. Physiology and Chemotaxonomic Characteristics
4. Conclusions
Description of Rhizobium terricola sp. nov.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Chaudhary, D.K.; Khulan, A.; Kim, J. Development of a novel cultivation technique for uncultured soil bacteria. Sci. Rep. 2019, 9, 6666. [Google Scholar] [CrossRef] [PubMed]
- Vartoukian, S.R.; Palmer, R.M.; Wade, W.G. Strategies for culture of “unculturable” bacteria. FEMS Microbiol. Lett. 2010, 309, 1–7. [Google Scholar] [CrossRef]
- Pulschen, A.A.; Bendia, A.G.; Fricker, A.D.; Pellizari, V.H.; Galante, D.; Rodrigues, F. Isolation of uncultured bacteria from Antarctica using long incubation periods and low nutritional media. Front. Microbiol. 2017, 8, 1346. [Google Scholar] [CrossRef] [PubMed]
- Frank, B. Über die Pilzsymbiose der Leguminosen. Ber. Dtsch. Bot. Ges. 1889, 7, 332–346. [Google Scholar]
- Young, J.M.; Kuykendall, L.D.; Martínez-Romero, E.; Kerr, A.; Sawada, H. A revision of Rhizobium Frank 1889, with an emended description of the genus, and the inclusion of all species of Agrobacterium Conn 1942 and Allorhizobium undicola de Lajudie et al. 1998 as new combinations: Rhizobium radiobacter, R. rhizogenes, R. rubi, R. undicola and R. vitis. Int. J. Syst. Evol. Microbiol. 2001, 51, 89–103. [Google Scholar] [PubMed]
- Mohapatra, B.; Sarkar, A.; Joshi, S.; Chatterjee, A.; Kazy, S.K.; Maiti, M.K.; Satyanarayana, T.; Sar, P. An arsenate-reducing and alkane-metabolizing novel bacterium, Rhizobium arsenicireducens sp. nov., isolated from arsenic-rich groundwater. Arch. Microbiol. 2017, 199, 191–201. [Google Scholar] [CrossRef]
- Zhang, X.; Harper, R.; Karsisto, M.; Lindstrom, K. Diversity of Rhizobium bacteria isolated from the root nodules of leguminous trees. Int. J. Syst. Evol. Microbiol. 1991, 41, 104–113. [Google Scholar] [CrossRef]
- Kaiya, S.; Rubaba, O.; Yoshida, N.; Yamada, T.; Hiraishi, A. Characterization of Rhizobium naphthalenivorans sp. nov. with special emphasis on aromatic compound degradation and multilocus sequence analysis of housekeeping genes. J. Gen. Appl. Microbiol. 2012, 58, 211–224. [Google Scholar] [CrossRef]
- Quan, Z.X.; Bae, H.S.; Baek, J.H.; Chen, W.F.; Im, W.T.; Lee, S.T. Rhizobium daejeonense sp. nov. isolated from a cyanide treatment bioreactor. Int. J. Syst. Evol. Microbiol. 2005, 55, 2543–2549. [Google Scholar] [CrossRef]
- Kaur, J.; Verma, M.; Lal, R. Rhizobium rosettiformans sp. nov., isolated from a hexachlorocyclohexane dump site, and reclassification of Blastobacter aggregatus Hirsch and Müller 1986 as Rhizobium aggregatum comb. nov. Int. J. Syst. Evol. Microbiol. 2011, 61, 1218–1225. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, R.P.; Ren, C.; Lai, Q.L.; Zeng, R.Y. Rhizobium marinum sp. nov., a malachitegreen-tolerant bacterium isolated from seawater. Int. J. Syst. Evol. Microbiol. 2015, 65, 4449–4454. [Google Scholar] [CrossRef] [PubMed]
- Sheu, S.Y.; Huang, H.W.; Young, C.C.; Chen, W.M. Rhizobium alvei sp. nov., isolated from a freshwater river. Int. J. Syst. Evol. Microbiol. 2015, 65, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Ramana, C.V.; Parag, B.; Girija, K.R.; Raghu Ram, B.; Venkata Ramana, V.; Sasikala, C. Rhizobium subbaraonis sp. nov., an endolithic bacterium isolated from beach sand. Int. J. Syst. Evol. Microbiol. 2013, 63, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Hunter, W.J.; Kuykendall, L.D.; Manter, D.K. Rhizobium selenireducens sp. nov.: A selenite-reducing α- Proteobacteria isolated from a bioreactor. Curr. Microbiol. 2007, 55, 455–460. [Google Scholar] [CrossRef]
- Mus, F.; Crook, M.B.; Garcia, K.; Costas, A.G.; Geddes, B.A.; Kouri, E.D.; Paramasivan, P.; Ryu, M.H.; Oldroyd, G.E.D.; Poole, P.S.; et al. Symbiotic nitrogen fixation and the challenges to its extension to nonlegumes. Appl. Environ. Microbiol. 2016, 82, 3698–3710. [Google Scholar] [CrossRef] [PubMed]
- Zahran, H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [CrossRef]
- Dahal, R.H.; Kim, J. Fluviicola kyonggii sp. nov., a bacterium isolated from forest soil and emended description of the genus Fluviicola. Int. J. Syst. Evol. Microbiol. 2018, 68, 1885–1889. [Google Scholar] [CrossRef]
- Dahal, R.H.; Chaudhary, D.K.; Kim, J. Pinisolibacter ravus gen. nov., sp. nov., isolated from pine forest soil and allocation of the genera Ancalomicrobium and Pinisolibacter to the family Ancalomicrobiaceae fam. nov., and emendation of the genus Ancalomicrobium Staley 1968. Int. J. Syst. Evol. Microbiol. 2018, 68, 1955–1962. [Google Scholar] [CrossRef]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Fitch, W.M. Toward defining the course of evolution: Minimum change for a specific tree topology. Syst. Zool. 1971, 20, 406. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Lee, I.; Chalita, M.; Ha, S.-M.; Na, S.-I.; Yoon, S.-H.; Chun, J. ContEst16S: An algorithm that identifies contaminated prokaryotic genomes using 16S RNA gene sequences. Int. J. Syst. Evol. Microbiol. 2017, 67, 2053–2057. [Google Scholar] [CrossRef]
- Seppey, M.; Manni, M.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness. Methods Mol. Biol. 2019, 1962, 227–245. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Ha, S.-M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Grant, J.R.; Stothard, P. The CGView Server: A comparative genomics tool for circular genomes. Nucleic Acids Res. 2008, 36, W181–W184. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A web server for whole-genome comparison and annotation of orthologous clusters across multiple species. Nucleic Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef]
- Doetsch, R.N. Determinative Methods of Light Microscopy. In Manual of Methods for General Bacteriology; Gerdhardt, P., Murray, R.G.E., Costilow, R.N., Nester, E.W., Wood, W.A., Krieg, N.R., Phillips, G.B., Eds.; American Society for Microbiology: Washington, DC, USA, 1981; pp. 21–33. [Google Scholar]
- Breznak, J.A.; Costilow, R.N. Physicochemical factors in growth. In Methods for General and Molecular Bacteriology; Reddy, C.A., Beveridge, T.J., Breznak, J.A., Marzluf, G.A., Schmidt, T.M., Snyder, L.R., Eds.; American Society of Microbiology: Washington, DC, USA, 2007; pp. 309–329. [Google Scholar]
- Smibert, R.M.; Krieg, N.R. Phenotypic characterization. In Methods for General and Molecular Bacteriology; Gerhardt, P., Murray, R.G.E., Wood, W.A., Krieg, N.R., Eds.; American Society for Microbiology: Washington, DC, USA, 1994; pp. 607–654. [Google Scholar]
- Dahal, R.H.; Kim, J. Dyadobacter flavus sp. nov. and Dyadobacter terricola sp. nov., two novel members of the family Cytophagaceae isolated from forest soil. Arch. Microbiol. 2018, 200, 1067–1074. [Google Scholar] [CrossRef]
- Miladiarsi; Mubarik, N.R.; Widyastuti, R. Selection, characterization and application of rhizobacteria and its effect on chili (Capsicum annuum L.) Plant Growth. Res. J. Microbiol. 2017, 12, 161–169. [Google Scholar] [CrossRef]
- Sasser, M. Bacterial Identification by Gas Chromatographic Analysis of Fatty Acid Methyl Esters (GC-FAME); MIDI Technical Note 101; MIDI Inc.: Newark, DE, USA, 1990. [Google Scholar]
- Minnikin, D.E.; O’Donnell, A.G.; Goodfellow, M.; Alderson, G.; Athalye, M.; Schaal, A.; Parlett, J.H. An integrated procedure for the extraction of bacterial isoprenoid quinones and polar lipids. J. Microbiol. Methods 1984, 2, 233–241. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidis, K.T.; Tiedje, J.M. Towards a genome-based taxonomy for prokaryotes. J. Bacteriol. 2005, 187, 6258–6264. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genome Features | Value |
|---|---|
| Genome size (bp) | 4,930,044 |
| G+C content (mol%) | 63.1 |
| No. of contigs | 19 |
| N50 | 1,092,225 |
| No. of subsystem | 327 |
| No. of proteins | 4589 |
| Total genes | 4682 |
| CDSs (total) | 4629 |
| Protein-coding genes | 4589 |
| Genes (RNA) | 53 |
| Complete rRNAs (5S, 16S, 23S) | 3 (1, 1, 1) |
| tRNAs | 46 |
| ncRNAs | 4 |
| Pseudo genes (total) | 40 |
| CDSs (without protein) | 4589 |
| Genome coverage | 254.0× |
| CRISPR repeats | 1 |
| Cas cluster | 0 |
| Complete and single-copy BUSCOs (S) | 143 (96.62%) |
| Complete and duplicated BUSCOs (D) | 0 (0%) |
| Fragmented BUSCOs (F) | 0 (0%) |
| Missing BUSCOs (M) | 5 (3.38%) |
| Total BUSCO groups searched) | 148 (100%) |
| Strains | S-51T | ||
|---|---|---|---|
| ANI | AAI | dDDH | |
| Rhizobium naphthalenivorans NBRC 107585T | 80.7 | 81.5 | 23.0 |
| Rhizobium selenitireducens ATCC BAA-1503T | 82.6 | 83.6 | 25.3 |
| Rhizobium daejeonense CCBAU 10050T | 78.8 | 78.8 | 21.9 |
| Rhizobium rubi NBRC 13261T | 74.3 | 53.3 | 19.7 |
| Rhizobium pusense NRCPB10T | 75.2 | 68.3 | 20.9 |
| Characteristic | 1 | 2 | 3 |
|---|---|---|---|
| Isolation source | Forest soil | (Sediment) | (Bioreactor) |
| Maximum growth temperature (°C) | 37 | 37 | 40 |
| pH range | 5.5–11.0 | 6.0–9.0 | 6.0–9.5 |
| Highest salt tolerance (%, w/v) | 5.0 | 3.0 | 1.5 |
| Nitrate reduction | + | – | + |
| Hydrolysis of | |||
| Urea | + | + | – |
| CM-cellulose | – | – | + |
| Enzyme activities | |||
| Acid phosphatase | w | + | + |
| Alkaline phosphatase | w | + | + |
| Esterase (C4) | w | + | + |
| Naphthol-AS-BI-phosphohydrolase | – | + | – |
| α-glucosidase | + | + | – |
| β-galactosidase | – | + | + |
| β-glucosidase | + | – | – |
| Assimilation from: | |||
| Adipic acid | w | – | – |
| Capric acid | – | + | + |
| d-maltose | + | + | – |
| d-mannitol | + | – | + |
| d-mannose | + | + | – |
| d-sorbitol | + | – | + |
| l-alanine | w | + | + |
| l-fucose | + | – | + |
| l-histidine | – | – | + |
| l-rhamnose | + | – | + |
| Malic acid | + | + | – |
| Potassium gluconate | – | + | – |
| Propionic acid | w | – | – |
| DNA G + C content (mol%) | 63.1 | 61.2 | 63.5 |
| Fatty Acid | 1 | 2 | 3 |
|---|---|---|---|
| Saturated | |||
| C16:0 | 1.2 | 1.7 | 1.9 |
| C18:0 | 6.5 | 10.2 | 7.9 |
| C19:0 cycloω8c | 7.9 | 6.9 | 4.6 |
| C19:0 10-methyl | tr | 0.9 | tr |
| Unsaturated | |||
| C18:1ω9c | 2.3 | – | 1.1 |
| C20:1ω7c | 2.0 | 0.6 | 1.3 |
| Hydroxy | |||
| C18:0 3OH | 2.0 | 3.3 | 2.9 |
| Summed features * | |||
| 2† | 4.4 | – | 3.9 |
| 3 | 0.6 | 3.1 | 1.3 |
| 8 | 71.9 | 72.3 | 73.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dahal, R.H.; Chaudhary, D.K.; Kim, J.; Kim, D.-U.; Kim, J. Genome Insight and Description of Previously Uncultured N2-Fixing Bacterium Rhizobium terricola sp. nov., Isolated from Forest Rhizospheric Soil by Using Modified Culture Method. Diversity 2022, 14, 733. https://doi.org/10.3390/d14090733
Dahal RH, Chaudhary DK, Kim J, Kim D-U, Kim J. Genome Insight and Description of Previously Uncultured N2-Fixing Bacterium Rhizobium terricola sp. nov., Isolated from Forest Rhizospheric Soil by Using Modified Culture Method. Diversity. 2022; 14(9):733. https://doi.org/10.3390/d14090733
Chicago/Turabian StyleDahal, Ram Hari, Dhiraj Kumar Chaudhary, Jungmin Kim, Dong-Uk Kim, and Jaisoo Kim. 2022. "Genome Insight and Description of Previously Uncultured N2-Fixing Bacterium Rhizobium terricola sp. nov., Isolated from Forest Rhizospheric Soil by Using Modified Culture Method" Diversity 14, no. 9: 733. https://doi.org/10.3390/d14090733