
Effects of Growth Conditions on Phenolic Composition and Antioxidant Activity in the Medicinal Plant Ageratina petiolaris (Asteraceae)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Approach and Experimental Design
Sampling of Plants for Analysis
2.3. Determination of Phenolic Compounds and Antioxidant Activity by Spectrophotometry
2.3.1. Sample Preparation
2.3.2. Total Polyphenols
2.3.3. Flavonoids
2.3.4. Antioxidant Activity by DPPH and FRAP
2.4. Determination of Phenolic Acids and Flavonoids by HPLC-DAD
2.4.1. Sample Preparation
2.4.2. HPLC-DAD Analysis
2.5. Statistical Analysis
3. Results
3.1. Total Phenolic Compounds and Antioxidant Activity
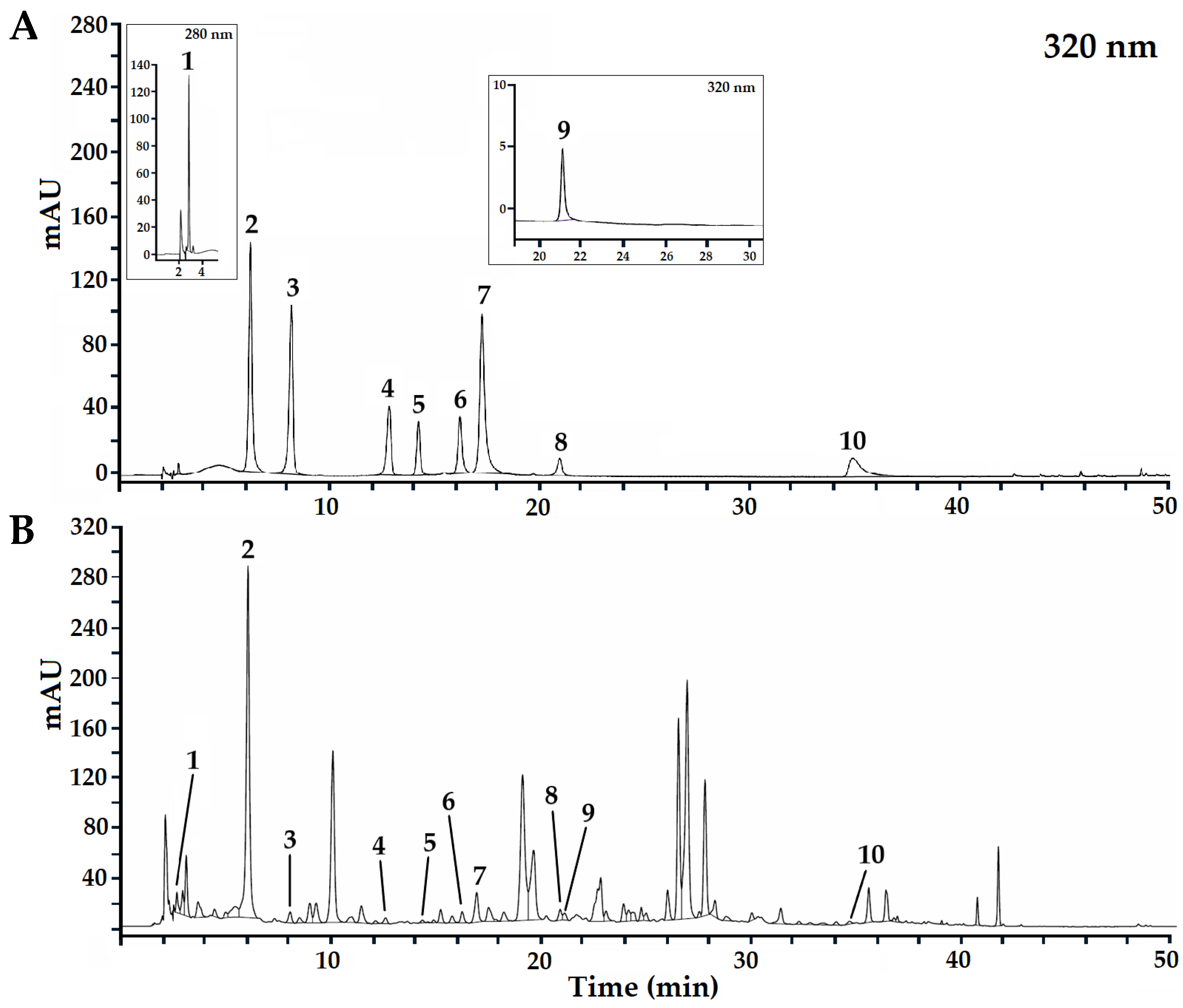
3.2. Profile of Phenolic Acids and Flavonoids
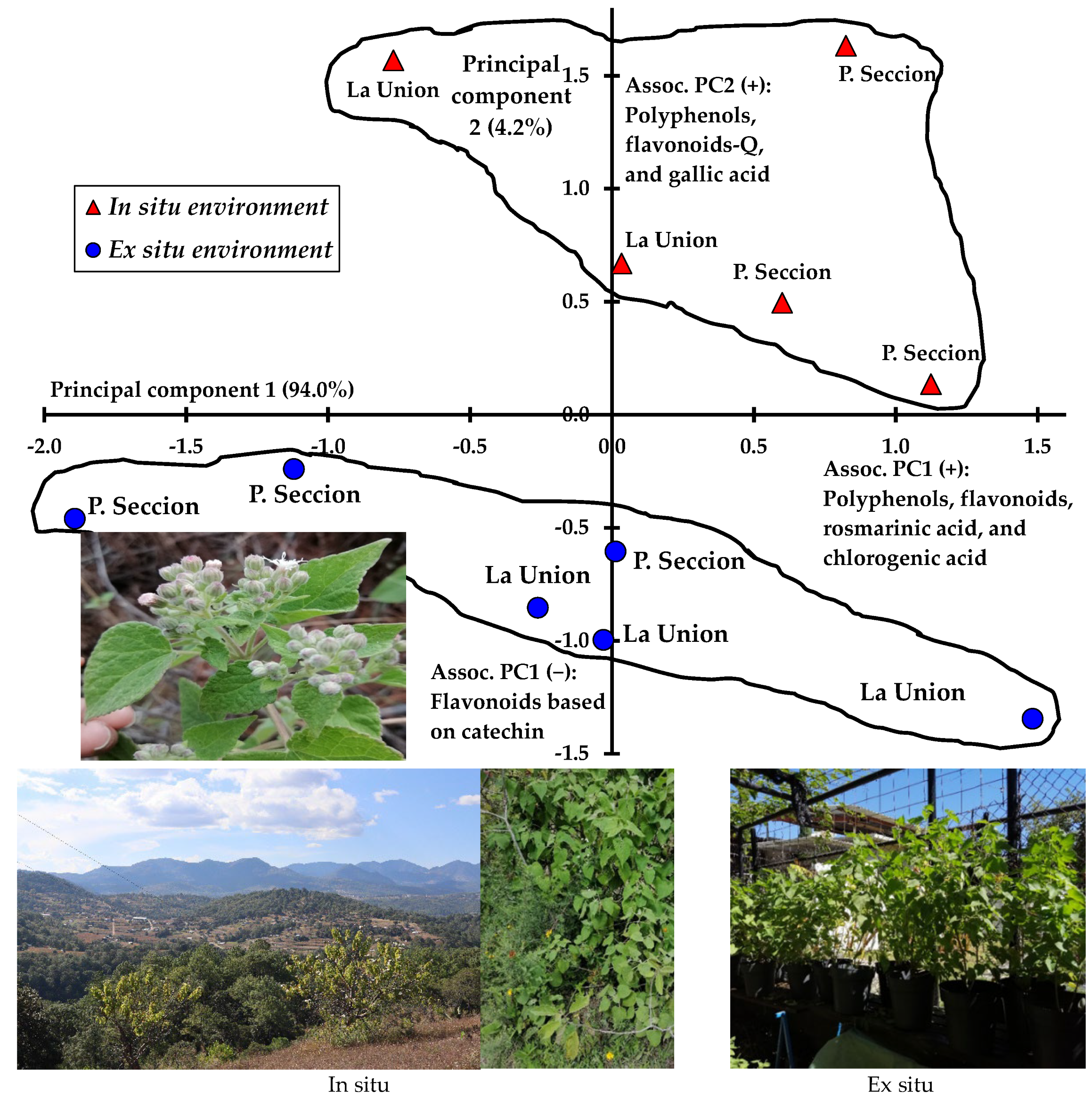
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Ochoa, M.L.; Chávez-Servia, J.L.; Vera-Guzmán, A.M.; Aquino-Bolaños, E.N.; Carrillo-Rodríguez, J.C. Medicinal plants used by indigenous communities of Oaxaca, Mexico, to treat gastrointestinal disorders. In Pharmacognosy—Medicinal Plants; Perveen, S., Al-Taweel, A., Eds.; IntechOpen: London, UK, 2019; pp. 1–37. [Google Scholar]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant. Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Nisar, A. Medicinal plants and phenolic compounds. In Phenolic Compounds—Chemistry, Synthesis, Diversity, Non-Conventional Industrial Pharmaceutical and Therapeutic Applications; Badria, F.A., Ed.; IntechOpen: London, UK, 2022; pp. 1–11. [Google Scholar] [CrossRef]
- Banothu, V.; Uma, A. Effect of biotic and abiotic stresses on plant metabolic pathways. In Phenolic Compounds—Chemistry, Synthesis, Diversity, Non-Conventional Industrial Pharmaceutical and Therapeutic Applications; Badria, F.A., Ed.; IntechOpen: London, UK, 2022; pp. 1–11. [Google Scholar] [CrossRef]
- Sampaio, B.L.; Edrada-Ebel, R.; Batista-Da Costa, F. Effect of the environment on the secondary metabolic profile of Tithonia diversifolia: A model for environmental metabolomics of plants. Sci. Rep. 2016, 6, 29265. [Google Scholar] [CrossRef] [Green Version]
- Bautista, I.; Boscaiu, M.; Lidón, A.; Linares, J.V.; Lull, C.; Donat, M.P.; Mayoral, O.; Vicente, O. Environmentally induced changes in antioxidant phenolic compounds levels in wild plants. Acta Physiol. Plant. 2016, 38, 9. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-Y.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef]
- Verma, N.; Shukla, S. Impact of various factors responsible for fluctuation in plant secondary metabolites. J. Appl. Res. Med. Aromat. Plants 2015, 2, 105–113. [Google Scholar] [CrossRef]
- Sharma, A.; Flores-Vallejo, R.C.; Cardoso-Taketa, A.; Villarreal, M.L. Antibacterial activities of medicinal plants used in Mexican traditional medicine. J. Ethnopharmacol. 2017, 208, 264–329. [Google Scholar] [CrossRef] [PubMed]
- Bustos-Brito, C.; Andrade-Cetto, A.; Giraldo-Aguirre, J.D.; Moreno-Vargas, A.D.; Quijano, L. Acute hypoglycemic effect and phytochemical composition of Ageratina petiolaris. J. Ethnopharmacol. 2016, 185, 341–346. [Google Scholar] [CrossRef]
- Mata-Torres, G.; Andrade-Cetto, A.; Espinoza-Hernández, F.A.; Cárdenas-Vázquez, R. Hepatic glucose output inhibition by Mexican plants used in the treatment of type 2 diabetes. Front. Pharmacol. 2020, 11, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo-Juárez, I.; González, V.; Jaime-Aguilar, H.; Martínez, G.; Linares, E.; Bye, R.; Romero, I. Anti-Helicobacter pylori activity of plants used in Mexican traditional medicine for gastrointestinal disorders. J. Ethnopharmacol. 2009, 122, 402–405. [Google Scholar] [CrossRef]
- De Rzedowski, G.C.; Rzedowski, J. Flora Fanerogámica del Valle de México, 2nd ed.; Instituto de Ecología, A.C., Comisión Nacional para el Conocimiento, Uso de la Biodiversidad: Pátzcuaro, México, 2005; pp. 786–800. [Google Scholar]
- UNAM. Biblioteca Digital de la Medicina Tradicional Mexicana; Universidad Nacional Autónoma de México: Ciudad de México, Mexico, 2009; Available online: http://www.medicinatradicionalmexicana.unam.mx/index.php (accessed on 25 January 2022).
- Borges, L.L.; Alves, S.F.; Sampaio, B.L.; Conceição, E.C.; Bara, M.T.F.; Paula, J.R. Environmental factors affecting the concentration of phenolic compounds in Myrcia tomentosa leaves. Rev. Bras. Farmacogn.-Braz. J. Pharmacogn. 2013, 23, 230–238. [Google Scholar] [CrossRef] [Green Version]
- INEGI (Instituto Nacional de Estadística y Geografía). Compendio de Información Geográfica Municipal 2010. San Martín Huamelúlpam, Oaxaca; INEGI: Aguascalientes, Mexico, 2005.
- NOM (Norma Oficial Mexicana). NOM-021-RECNAT-2000; Especificaciones de Fertilidad, Salinidad y Clasificación de Suelos. Estudios, Muestreo y Análisis. Diario Oficial, 31 December 2002; 73. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Lin, J.-Y.; Tang, C.-Y. Determination of total phenolic and flavonoid contents in selected fruits and vegetables, as well as their stimulatory effects on mouse splenocyte proliferation. Food Chem. 2007, 101, 140–147. [Google Scholar] [CrossRef]
- Pękal, A.; Pyrzynska, K. Evaluation of aluminium complexation reaction for flavonoid content assay. Food Anal. Methods 2014, 7, 1776–1782. [Google Scholar] [CrossRef] [Green Version]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999, 299, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Pająk, P.; Socha, R.; Broniek, J.; Królikowska, K.; Fortuna, T. Antioxidant properties, phenolic and mineral composition of germinated chia, golden flax, evening primrose, phacelia and fenugreek. Food Chem. 2019, 275, 69–76. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute Inc. (SAS). Base SAS® 9.1.3 Procedures Guide, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2006; Volumes 1–4. [Google Scholar]
- Gui, F.-R.; Wan, F.-H.; Guo, J.-Y. Population genetics of Ageratina adenophora using inter-simple sequences repeat (ISSR) molecular markers in China. Plant Biosyst. 2008, 142, 255–263. [Google Scholar] [CrossRef]
- Arbona, V.; Manzi, M.; de Ollas, C.; Gómez-Cárdenas, A. Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef]
- Nurzyńska-Wierdak, R. Does mineral fertilization modify essential oil content and chemical composition in medicinal plants? Acta Sci. Pol. 2013, 12, 3–16. [Google Scholar]
- Krishnaiah, D.; Sarbatly, R.; Mithyanandam, R. A review of the antioxidant potential of medicinal plant species. Food Bioprod. Process. 2011, 89, 217–233. [Google Scholar] [CrossRef]
- Arciniegas, A.; Pérez-Castorena, A.L.; Meléndez-Aguirre, M.; Ávila, J.G.; García-Bores, A.M.; Villaseñor, J.L.; de Vivar, A.R. Chemical composition and antimicrobial activity of Ageratina deltoidea. Chem. Biodivers. 2018, 15, e1700529. [Google Scholar] [CrossRef]
- Sánchez-Ramos, M.; Marquina-Bahena, S.; Alvarez, L.; Román-Guerrero, A.; Bernabé-Antonio, A.; Cruz-Sosa, F. Phytochemical, Pharmacological, and Biotechnological Study of Ageratina pichinchensis: A Native Species of Mexico. Plants 2021, 10, 2225. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Peralta, L.; Ocampo-Acuña, Y.D.; Rios, M.Y. Secondary metabolites from two varieties of Ageratina espinosarum and their chemophenetic significance. Biochem. Syst. Ecol. 2022, 102, 104409. [Google Scholar] [CrossRef]
- Romero-Cerecero, O.; Zamilpa, A.; González-Cortazar, M.; Alonso-Cortés, D.; Jiménez-Ferrer, E.; Nicasio-Torres, P.; Aguilar-Santamaría, L.; Tortoriello, J. Pharmacological and chemical study to identify wound-healing active compounds in Ageratina pichinchensis. Planta Med. 2013, 79, 622–627. [Google Scholar] [CrossRef]
- Hodaei, M.; Rahimmalek, M.; Arzani, A.; Talebi, M. The effect of water stress on phytochemical accumulation, bioactive compounds and expression of key genes involved in flavonoid biosynthesis in Chrysanthemum morifolium L. Ind. Crops. Prod. 2018, 120, 295–304. [Google Scholar] [CrossRef]
- Bettaieb, I.; Hamrouni-Sellami, I.; Bourgou, S.; Limam, F.; Marzouk, B. Drought effects on polyphenol composition and antioxidant activities in aerial parts of Salvia officinalis L. Acta Physiol. Plant. 2011, 33, 1103–1111. [Google Scholar] [CrossRef]
- Singh, G.; Passsari, A.K.; Leo, V.V.; Mishra, V.K.; Subbarayan, S.; Singh, B.P.; Kumar, B.; Kumar, S.; Gupta, V.K.; Lalhlenmawia, H.; et al. Evaluation of phenolic content variability along with antioxidant, antimicrobial, and cytotoxic potential of selected traditional medicinal plants from India. Front. Plant Sci. 2016, 7, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Geographic and Environmental Growing Conditions of A. petiolaris | Natural In Situ Conditions | Ex Situ Cultivation | |
|---|---|---|---|
| San Martin Huamelulpam | Santa Cruz Xoxocotlan | ||
| Primera Seccion | La Union | ||
| Location descriptors: | |||
| Altitude (m) | 2221 | 2235 | 1530 |
| Latitude (N) | 17°23′42.7″ | 17°23′29.8″ | 17°02′64.6″ |
| Longitude (W) | 97°36′41.3″ | 97°37′26.6″ | 96°71′95.3″ |
| Annual average temperature (°C) a | 14–18 | 18–20 | |
| Annual average rainfall (mm) | 700–1000 | 600–700 | |
| Solar radiation (W/m2, maximum) | 1155–1308 | 1297–1349 | |
| Climate predominant | Temperate sub-humid with precipitation from June to November | Semi-dry to semi-warm | |
| Chemical analyses of soil b: | |||
| Organic matter (%) | 1.98 | 7.67 | 9.32 |
| pH (in H2O) | 8.06 | 7.72 | 6.99 |
| P-Olsen (mg kg−1) | 2.89 | 11.60 | 101.50 |
| K (cmol kg−1) | 0.72 | 0.75 | 2.66 |
| Ca (cmol kg−1) | 47.8 | 43.9 | 29.95 |
| Mg (cmol kg−1) | 0.86 | 1.62 | 4.35 |
| Na (cmol kg−1) | 0.10 | 0.05 | 0.50 |
| Fe (mg kg−1) | 2.29 | 6.26 | 12.38 |
| Zn (mg kg−1) | 0.77 | 1.15 | 60.34 |
| Mn (mg kg−1) | 8.44 | 26.53 | 38.16 |
| Cu (mg kg−1) | nd | 0.62 | 1.12 |
| Inorganic N (mg kg−1) | 3.71 | 7.78 | 125.80 |
| Electric conductivity | 0.25 | 0.38 | 1.20 |
| Cation exchange capacity (cmol kg−1) | 50.00 | 47.00 | 38.50 |
| Sources of Variation | Total Polyphenols | Flavonoids | Ant. Activity | ||
|---|---|---|---|---|---|
| Quercetin Equiv. | Catechin Equiv. | DPPH | FRAP | ||
| Growth environment (E) | 1389.8 ** | 9.13 ** | 1.29 ns | 102.1 ** | 224.1 ** |
| Origin locations of samples (Po) | 81.9 ns | 0.008 ns | 0.25 ns | 15.5 ns | 3.2 ns |
| E × Po | 4199.1 ** | 11.5 ** | 36.52 ** | 139.3 ** | 383.4 ** |
| Sampling (S) | 58.0 ns | 0.001 ns | 0.16 ns | 1.7 ns | 4.1 ns |
| Replica of Lab./S 1 | 9.6 ns | 0.006 ns | 0.02 ns | 0.2 ns | 0.4 ns |
| Error | 69.5 | 0.2 | 0.5 | 4.5 | 6.9 |
| Coeff. variation (%) | 20.9 | 16.4 | 16.0 | 16.2 | 16.9 |
| Study Factors | Levels (Locations of Oaxaca, Mexico) | Total Polyphenols (mg GAE g−1 dw) | Flavonoids (mg g−1 dw) | Antiox. Activ. (µmol TE g−1 dw) | ||
|---|---|---|---|---|---|---|
| QE 1 | CE 1 | DPPH | FRAP | |||
| Growth environments (E) | In situ | 46.8 a 2 | 11.2 a | 23.3 a | 220.7 a | 338.3 a |
| Ex situ | 37.9 b | 7.2 b | 20.8 a | 165.2 b | 231.3 b | |
| Origin locations of samples (Po) | P. Seccion | 35.5 a 2 | 7.3 a | 18.1 a | 154.0 a | 223.3 a |
| La Union | 44.6 a | 9.0 a | 24.9 a | 202.6 a | 288.6 a | |
| Environment–location (E × Po) interactions: | ||||||
| Primera Seccion | In situ | 51.8 a 2 | 13.0 a | 27.8 a | 241.3 a | 394.4 a |
| Ex situ | 30.1 c | 5.4 c | 14.8 b | 124.9 c | 166.3 c | |
| La Union | In situ | 39.3 b | 8.5 b | 16.6 b | 189.7 b | 254.2 b |
| Ex situ | 45.8 a | 9.1 b | 26.7 a | 205.5 ab | 296.3 b | |
| Sources of Variation | Origin Locations (Po) | Growth Environment (E) | Po × E | Sampling (S) | Rep./S 1 | Error | CV (%) |
|---|---|---|---|---|---|---|---|
| Gallic acid | 3862.8 ** | 54,618.0 * | 3873.8 ** | 2257.7 ** | 506.9 ** | 38.5 | 16.5 |
| Chlorogenic acid | 1.47 ns | 13.66 ns | 449.20 ** | 15.99 ns | <0.01 ns | 8.4 | 18.4 |
| Caffeic acid | 0.32 ns | 0.14 ns | 18.77 ** | 0.40 ns | <0.01 ns | 0.34 | 14.5 |
| ρ-Coumaric acid | <0.001 ns | 0.454 * | 3.609 ** | 0.314 * | <0.001 ns | 0.11 | 10.3 |
| Robinin | 8.72 ** | 26.9 ** | 1.29 * | 11.76 ** | <0.01 ns | 0.92 | 26.1 |
| Rutin | 5813.5 ** | 55,236.9 ** | 5829.8 ** | 3928.0 ** | 2.2 ns | 1.9 | 4.0 |
| Luteolin-7-glucoside | 4,269,150 ** | 13,116,425 ** | 3,327,901 ** | 544,166 ** | 1501 ns | 2220 | 5.0 |
| Apigenin-7-glucoside | 0.2 ns | 164,364 ** | 789.9 ** | 18,493.4 ** | 23.7 ns | 34.4 | 6.2 |
| Rosmarinic acid | 149.6 ns | 48,836.8 ** | 7205.7 ** | 866.7 ** | 37.0 ns | 55.9 | 15.5 |
| Kaempferol | 20.69 ** | 2.08 ** | 6.58 ** | 1.19 ** | <0.01 ns | 0.18 | 12.7 |
| Compounds Evaluated (µg g−1 dw) | Origin Locations of Plants (Po), Oaxaca | Growth Environments (E) | Loc.–Environment Interactions (Po × E) | |||||
|---|---|---|---|---|---|---|---|---|
| Primera Seccion | La Union | |||||||
| P. Seccion | La Union | In Situ | Ex Situ | In Situ | Ex Situ | In Situ | Ex Situ | |
| Gallic acid | 30.3 b 1 | 45.0 a | 94.1 | nd | 75.7 b | nd | 112.4 a | nd |
| Chlorogenic acid | 4228.0 a | 5336.5 a | 5006.9 a | 4632.5 a | 7434 a | 2091 b | 2580 b | 7174 a |
| Caffeic acid | 53.6 a | 74.2 a | 61.9 a | 65.2 a | 90.5 a | 28.9 b | 33.3 b | 101.4 a |
| ρ-Coumaric acid | 24.0 a | 24.2 a | 27.6 a | 21.7 b | 38.4 a | 14.4 c | 16.8 c | 29.1 b |
| Robinin | 114.6 a | 44.4 b | 157.8 a | 27.3 b | 221.9 a | 43.0 bc | 93.6 b | 11.6 c |
| Rutin | 43.0 a | 25.2 b | 85.3 | nd | 107.6 a | nd | 63.0 b | nd |
| Luteolin-7-glucoside | 1229.7 a | 633.3 b | 1454.3 a | 408.8 b | 2015.8 a | 443.7 c | 892.7 b | 373.9 d |
| Apigenin-7-glucoside | 90.2 a | 101.5 a | 156.9 a | 45.9 b | 161.1 a | 43.0 d | 152.7 b | 50.2 c |
| Rosmarinic acid | 48.4 a | 47.7 a | 89.3 a | 23.4 b | 99.4 a | 14.4 d | 69.1 b | 37.0 c |
| Kaempferol | 9.5 b | 69.1 a | 21.1 b | 51.4 a | 12.0 b | 7.8 b | 30.3 b | 94.9 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Ochoa, M.L.; Vera-Guzmán, A.M.; Mondragón-Chaparro, D.M.; Sandoval-Torres, S.; Carrillo-Rodríguez, J.C.; Chávez-Servia, J.L. Effects of Growth Conditions on Phenolic Composition and Antioxidant Activity in the Medicinal Plant Ageratina petiolaris (Asteraceae). Diversity 2022, 14, 595. https://doi.org/10.3390/d14080595
Pérez-Ochoa ML, Vera-Guzmán AM, Mondragón-Chaparro DM, Sandoval-Torres S, Carrillo-Rodríguez JC, Chávez-Servia JL. Effects of Growth Conditions on Phenolic Composition and Antioxidant Activity in the Medicinal Plant Ageratina petiolaris (Asteraceae). Diversity. 2022; 14(8):595. https://doi.org/10.3390/d14080595
Chicago/Turabian StylePérez-Ochoa, Mónica L., Araceli M. Vera-Guzmán, Demetria M. Mondragón-Chaparro, Sadoth Sandoval-Torres, José C. Carrillo-Rodríguez, and José L. Chávez-Servia. 2022. "Effects of Growth Conditions on Phenolic Composition and Antioxidant Activity in the Medicinal Plant Ageratina petiolaris (Asteraceae)" Diversity 14, no. 8: 595. https://doi.org/10.3390/d14080595