3.1. Microbiota of Corn Silage
In the control samples, lactic acid bacteria ranged from 2.69 log cfu/g on APT to 5.11 log cfu/g on MRS, the numbers of coliform bacteria and enterococci were under the detection limit, the total number of microorganisms was 3.54 ± 0.43 log cfu/g and microscopic filamentous fungi was 2.73 ± 0.33 log cfu/g. In the silage with the additive on the base of lactic acid bacteria, lactic acid bacteria ranged from 2.83 log cfu/g on MSE to 5.05 log cfu/g on MRS, the numbers of coliform bacteria and enterococci were under the detection limit, total counts of microorganisms ranged from 3.23 to 3.51 log cfu/g and microscopic filamentous fungi ranged from 2.13 to 2.21 log cfu/g. In the silage with the nutritional addition, urea, lactic acid bacteria ranged from 4.14 to 4.21 log cfu/g, numbera of coliform bacteria and enterococci were under the detection limit, total number of microorganisms was 3.26 ± 0.22 log cfu/g, and microscopic filamentous fungi was 3.38 ± 0.45 log cfu/g (
Table 2). Generally, the fermentation quality is largely influenced by the characteristic of the raw material and the epiphytic microorganisms on its surface [
28]. Final feed quality is largely influenced by the species and numbers of dominant microorganisms in the fermentation process [
29].
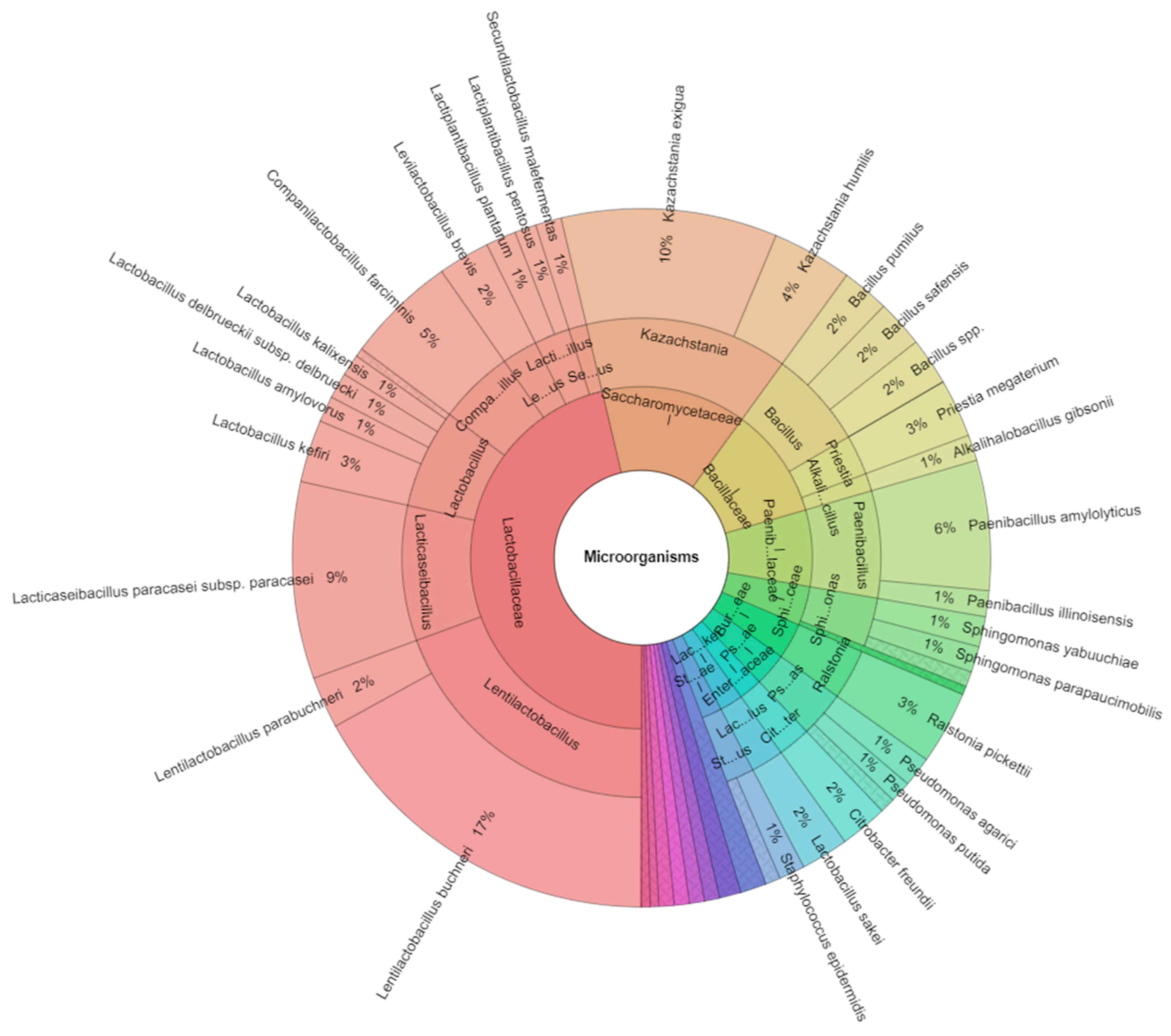
In our study, a total of 43 species of bacteria and yeast were identified (
Table 3). The 17 families included Bacillaceae, Burkholderiaceae, Clostridiaceae, Cryptococcaceae, Enterobacteriaceae, Lachnospiraceae, Lactobacillaceae, Moraxellaceae, Micrococaceae, Paenibacillaceae, Promicromonosporaceae, Pseudomonadaceae, Rhizobiaceae, Saccharomycetaceae, Shewanellaceae, Sphingomonadaceae, and Staphylococcaceae, and 25 genera included
Acinetobacter, Arthrobacter, Alkalihalobacillus, Priestia, Bacillus, Blastomonas, Cellulosimicrobium, Citrobacter, Clostridium, Lacrimispora, Cryptococcus, Kazachstania, Lactobacillus, Levilactobacillus, Lentilactobacillus, Companilactobacillus, Secundilactobacillus, Lacticaseibacillus, Paenibacillus, Pseudomonas, Ralstonia, Rhizobium, Shewanella, Sphingomonas, and
Staphylococcus that were isolated from all samples of corn silage (
Table 4). The most isolated family from all corn silage was Lactobacillaceae. The most isolated species was
Lentilactobacillus buchneri (17%) (
Figure 1). Bacteria, yeast, and fungi with detoxification abilities were isolated from different sources, and LAB are the preferred candidates for eliminating mycotoxins in silages because they play a critical role in the ensiling fermentation.
Lactiplantibacillus plantarum and
Lentilactobacillus buchneri are known to enhance lactic acid (LA) fermentation and acetic acid (ACA) production to improve fermentation quality and inhibit aerobic spoilage [
23].
In the study of Wang et al. [
30] they examined the effects of lactic acid bacteria (LAB)
Lactiplantibacillus plantarum subsp. ZA3 and
Artemisia argyi (AA) on the fermentation characteristics, microbial community, and mycotoxins. The results showed that corn silage has microbial communities,
Acetobacter and
Enterobacter, which were inhibited in all AA groups, while a higher abundance of lactobacilli was maintained; moreover,
Candida,
Pichia, and
Kazachstania abundances were decreased in both groups. In our study, different results were found, the most isolated species were from the family Lactobacillaceae and yeast
Kazachastania exigua (10%). Our results did not confirm previous research. We can assume that the increased number of LAB increases the production of lactic acid, which affects the growth of yeast. The relatively lower pH values in inoculated silages, combining activities of acidification and antagonistic activity towards other bacteria, promotes the reduction in bacterial and fungi diversities, and ultimately improves feed quality. This observation indicated that the fungi community can change when the environment changes from anaerobic to aerobic. It is possible that acid-tolerant bacteria still dominate the bacterial community in the early period of aerobic exposure, and the variation in microbial community.
In our study, microscopic fungi in each group of samples were isolated and they were of the genera Alternaria, Aspergillus, Fusarium, Mucor, Rhizopus, and Penicillium. The most isolated genera 25% resp. 25% in control samples were Aspergillus and Penicillium. Similar results were found in samples treated with the addition of LAB with an incidence of Aspergillus of 12.5% and Penicillium of 18%, and in samples with nutritional additive urea with an incidence of Aspergillus of 15% and Penicillium of 20%.
In the study of Krustev and Khristov [
31], eight species of microscopic fungi were demonstrated in the sampled corn silage:
Mucor,
Penicillium,
Aspergillus,
Alternaria, and
Trichoderma, similar as in our study. It was established that the number of species in the surface layer of the ensilaged mass was the highest.
Isolation of novel LAB strains for application in silage has been a common practice over the years but it is still an activity with current importance around the globe [
32,
33,
34], due to the interest in collecting diverse strains for future applications not only as silage inoculants but also in other plant-based food for animal and human uses [
35].
Lentilactobacillus buchneri is presently the gold standard to promote aerobic stability in corn silage [
36,
37]. Acetic acid is one of the main organic acids produced by heterofermentative LAB and it has the capacity of promoting aerobic stability when silos are opened [
38].
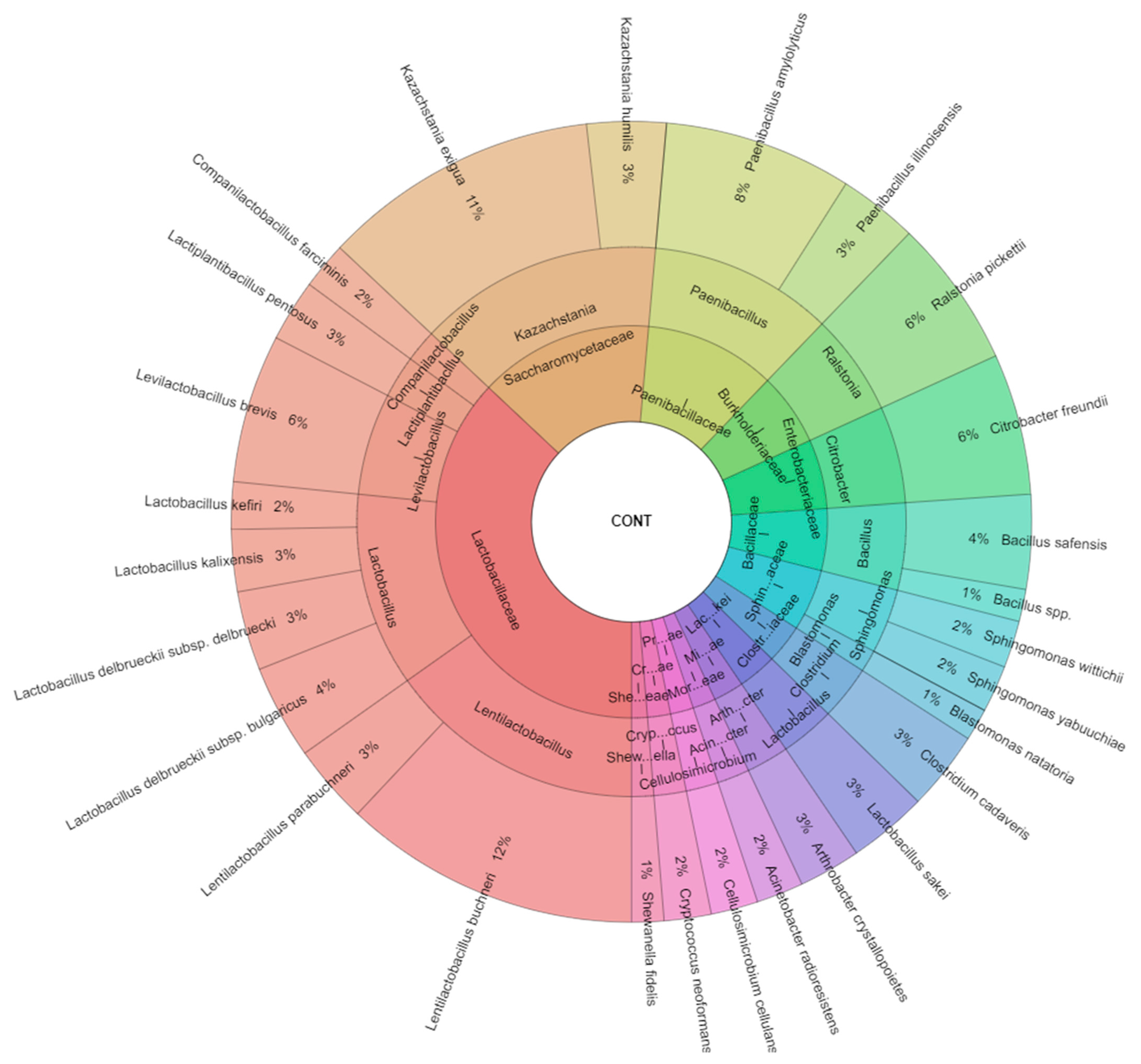
All isolated species in control samples were 27 species and the most isolated species was from the family Lactobacillaceae (9 species,
Figure 2). The most isolated species from control samples were
Lentilactobacillus buchneri (12%) and
Kazachstania exigua (11%), following with
Paenibacillus amylolyticus (8%),
Citrobacter freundii, and
Ralstonia picketii (6%).
L. buchneri Ls141 and 463 were used as external reference strains.
L. buchneri Ls141 had been isolated from corn silage in a previous study [
39].
Candida ethanolica,
Saccharomyces bulderi,
Pichia anomala,
Kazachstania unispora, and
Saccharomyces cerevisiae were the predominant yeasts.
Pichia anomala,
Issatchenkia orientalis,
S. cerevisiae, and
Pichia fermentans were the prevalent species in high moisture corn [
36].
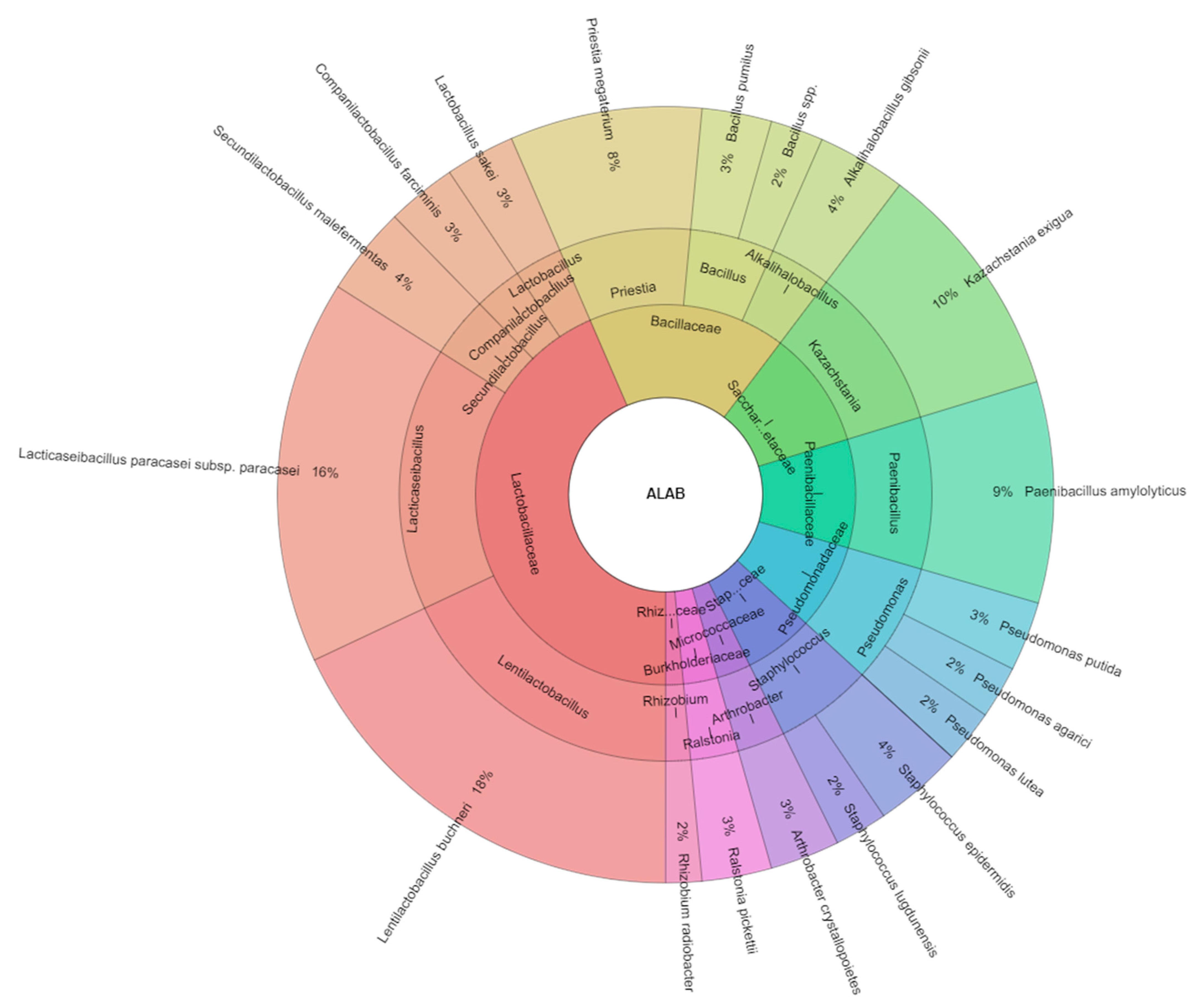
All isolated species in corn samples with the addition of LAB were 19 species and the most isolated species were from the families Lactibacillaceae (5 species) and Bacillaceae (4 species,
Figure 3). The most isolated species from corn samples with the addition of LAB were
Lentilactobacillus buchneri (18%) and
Lacticaseibacillus paracasei subsp.
paracasei (16%), following with
Kazachstania exiqua (10%),
Paenibacillus amylolyticus (9%), and
Priestia megaterium (8%). Driehuis et al. [
40] observed that strains of
Lentilactobacillus buchneri (
Lactobacillus buchneri) [
41] were able to degrade lactic acid into acetic acid and 1,2-propanediol [
42], which could then be metabolized into propionic acid [
43]. Since both acetate and propionate are strong yeast inhibitors [
44,
45], these modifications positively improve the aerobic stability of silage. More recently, co-inoculation with
L. buchneri NCIMB 40788 and
Lentilactobacillus hilgardii CNCM-I-4785 (
Lactobacillus hilgardii) was reported to increase the stability of different silages [
46,
47]. While microbial dynamics during fermentation were recently characterized in corn silage inoculated with these two microorganisms [
48], little research has been undertaken to characterize microbial succession and mycotoxin production in inoculated vs. uninoculated silages during the feed-out phase [
49,
50].
Kazachstania exiqua was isolated from corn silage in the study of Santos et al. [
36].
Ruminal microorganisms are capable of transforming nitrogen from NPN compounds into protein of high nutritional value. However, if the release of ammonia promoted by NPN exceeds the use capacity by ruminal microbiota, there will be an excretion of this excess with a consequent loss of energy. If the ammonia concentration extrapolates the excretion capacity, the intoxication of the animal may occur [
51]. Nitrogen from NPN compounds can be converted by ruminal microbes into protein with a high nutritional value [
52]. However, there will be the expulsion of this surplus and a resulting loss of energy if the release of ammonia encouraged by NPN exceeds the capacity for utilization by ruminal bacteria [
53].
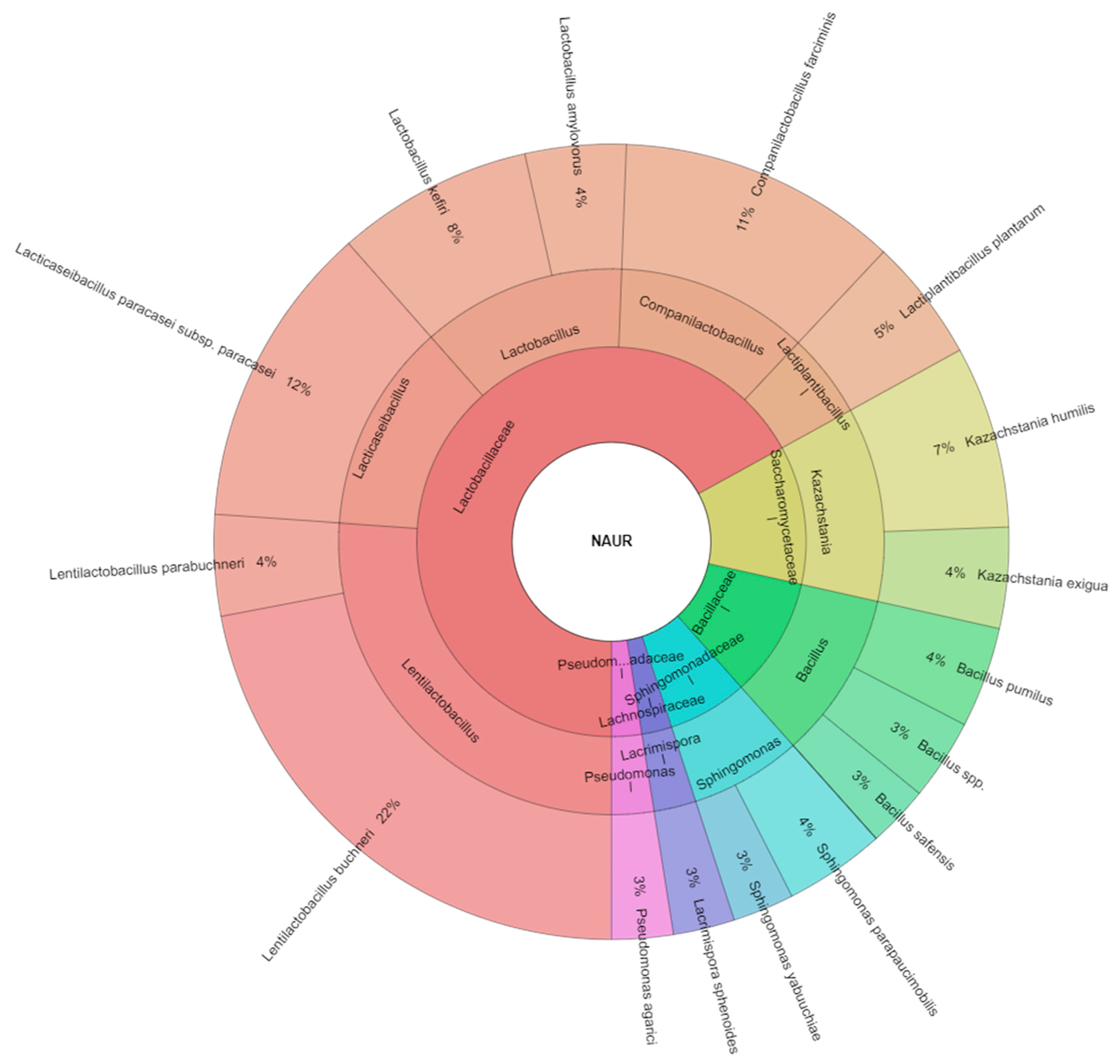
All isolated species from samples with nutritional additive urea were 16 species and most isolated species were from the Lactibacillaceae family similar to 7 species (
Figure 4). The most isolated species from the treated samples with nutritional additive urea were
Lentilactobacillus buchneri (22%) and
Lacticaseibacillus paracasei subsp.
paracasei (12%), following with
Companilactobacillus farciminis (11%) and
Lactobacillus kefiri (8%). Pang et al. [
54] reported that most of the bacterial community in silage belonged to the phylum Firmicutes and the genera
Lactobacillus, Pedicoccus, and
Weissella. These results showed that the dominant phyla in the measured samples were Proteobacteria and Firmicutes. This result was different from that of other researchers [
54,
55], who found that most bacteria involved in lactic acid fermentation of silage belonged to the genera
Lactobacillus,
Pedicoccus,
Weissella, and
Leuconostoc. Metagenomic analysis revealed that urea addition in the sheep diet significantly increased the relative abundance of genera involved in nitrogen metabolism especially. Increasing nitrogen sources by urea addition may be beneficial to microbial protein production. This could improve microbial utilization of additional N sources during ruminal fermentation. Therefore, the synchronization between ruminal ammonia nitrogen release and carbohydrate availability resulted in greater microbial protein synthesis [
56]. The bacterial composition was also altered by lysine supplementation to support energy metabolism, in which the microbial diversity was unchanged [
57].
3.2. Mycotoxin Composition of Corn Silage
Several mycotoxin species were identified in corn silage samples.
Table 5 and
Table 6 provide an overview on detected mycotoxin concentrations as well as on the level of significance. In this study, there were no statistically significant differences between variants of each mycotoxin species. Penagos-Tabares et al. [
58] confirmed the occurrence of the same mycotoxins as detected in this experiment. When mycotoxins are present, there are few ways to avoid unwanted problems and therefore prevention is essential. Some silage additives can reduce growth of fungi and hence mycotoxin formation [
14,
59]. Whitlow and Hagler [
59] found that these can be additives such as ammonia, propionic acid, sorbic acid, and bacterial or enzymatic additives. Dong et al. [
60] examined the interactions between the harvest stage and the dose of inoculant in corn silages and found that inoculant lowered (
p > 0.05) concentrations of deoxynivalenol at the milk stage. Furthermore, the inoculant significantly decreased or increased deoxynivalenol content with its different dosing at the dough stage [
60]. Aflatoxins and ochratoxins were not identified neither in control variant nor in experimental variants. In contrast, Kalúzová et al. [
16] determined these types of mycotoxins and decreased them with urea and inoculant addition. The effect of silage additives was manifested in reducing zearalenone content in experimental variants (
Table 5). Corn silage with urea achieved a significantly (
p < 0.05) higher concentration of zearalenone but its concentration after inoculant addition was even higher [
16]. An increased concentration of zearalenone was also reported by Drouin et al. [
61] when inoculant was added to the corn silage. The application of inoculant to corn silage resulted in increased fumonisin production, and Bakri [
62] registered the same effect in his experiment. Gallo et al. [
63] also detected fumonisins and found a greater level of fumonisin B1 (
p > 0.05) and fumonisin B2 (
p < 0.05) in inoculated corn. Corn silage NAUR resulted in a more than two times higher mean value of fumonisin B1 compared to CONT. Kalúzová et al. [
16] found a similar result with this nutritional additive but the differences were not significant. Teller et al. [
64] studied the effect of various additives on mycotoxin concentration in corn silage. A higher content of deoxynivalenol and fumonisin B1 was observed after inoculant treatment. As for zearalenone, the microbial additive lowered its content in silage samples [
64]. The highest value of all mycotoxins reached nivalenol in both CONT and ALAB variants. In comparison with the control, a higher content of nivalenol was observed in variants with inoculant addition. Wang et al. [
65] found a significant tendency in differences in nivalenol content depending on storage temperature and type/use of lactic acid bacteria. Eckard et al. [
66] detected nivalenol in 8 of 19 samples of corn silage between a range of 190 and 760 µg/kg. A lowering trend (
p > 0.05) of this mycotoxin in inoculated corn silage was noticed by Bakri [
62]. Variant NAUR was higher in nivalenol content than CONT, but lower compared to variant ALAB. Although type A trichothecenes and ergot alkaloids are present in cereal crops, they were not detected by the HPLC-MS/MS method in this experiment. Type A trichothecenes include, for instance, mycotoxins such as T-2 toxin and HT-2 toxin, which are quite often found in silage. Some authors [
16,
65,
67] confirmed a change in T-2 toxin concentration by using urea and various microbial additives in corn silages. Contamination by ergot alkaloids is mostly seen in forages such as tall fescue, sorghum, and ryegrass, but their presence in other forages is not refuted [
68,
69]. Zhang et al. [
69] noted that ergot alkaloids are rarely present in corn silage in China, but still, they occurred in one of their samples of corn silage at a concentration of 15.3 µg/kg.
Besides free
Fusarium mycotoxins (deoxynivalenol, fumonisin B1, nivalenol, and zearalenone), the presence of some other
Fusarium contaminants known as emerging mycotoxins (beauvericin, enniatins, and moniliformin) were also found. Beauvericin was detected only in corn silage with the addition of urea. An average concentration of 47 µg/kg of beauvericin by Zachariasova et al. [
70] was determined in corn silage samples. Reisinger et al. [
71] found 120 samples of corn silage positive on beauvericin, and this mycotoxin belonged to five most frequently detected mycotoxins in their study. In corn silage samples, Sørensen et al. [
72] did not detect the presence of enniatin A and A1 but confirmed the presence of enniatin B and B1. In contrast to Sørensen et al. [
72], the mean value of enniatin B was 28.01 µg/kg lower and that of enniatin B1 was 9.50 µg/kg lower in our corn silage samples without additive. On the other hand, neither enniatin A nor B1 were confirmed in the corn silage from Rasmussen et al. [
73], but the mean value of enniatin B (44.00 µg/kg) was lower compared to our study of silage without additive. Similarly, Storm et al. [
74] only reported the occurrence of enniatin B with an average value higher (53.00 µg/kg) than that reported in
Table 6 for the silage without additive. In whole-plant corn silage, enniatins did not appear in any of the samples or were detected at very low concentrations [
75]. However, Shimshoni et al. [
76] detected the occurrence of enniatin A (0.3 µg/kg), enniatin A1 (0.8 µg/kg), enniatin B (0.2 µg/kg), as well as enniatin B1 (0.9 µg/kg). Identical to the previous author, McElhinney et al. [
77] also found the presence of these mycotoxins, but in grass silage. Moreover, the presence of enniatin B was confirmed by Wambacq et al. [
78] in 82 corn silage samples with concentrations up to 5000 µg/kg. As in the present study, these values were not statistically significant in studies, which all the authors mentioned. Moniliformin was present in all samples; however, lower concentrations were obtained in both treated variants. Zhang et al. [
69] reported relatively low concentrations of moniliformin, and the highest concentration of 116 µg/kg was found. On the contrary, Gräfenhan et al. [
79] did not detect moniliformin in red clover silage (control and treated variant), but after addition of soil, moniliformin concentration jumped up to 222 µg/kg. As Kalúzová et al. [
14] mentioned, the effect of urea on mycotoxin concentrations has not been widely monitored so far and the effect on chemical composition and fermentation parameters was more closely monitored in this case. Some studies confirmed a positive suppression effect of silage additives on mycotoxin concentrations [
23,
61,
62,
63]. However, other studies reveal the increasing mycotoxin content after the application of silage additives [
17,
65]. Contradictory results are caused probably by many factors such as temperature, water activity, and pH value [
80,
81,
82], which are affecting the environment of microscopic fungi producing the mycotoxins in stressful conditions; thus other experiments are necessary in this research field.
3.3. Relationship between the Mycotoxin Concentrations and Microscopic Fungi and Yeasts, Harmful Effects, and Mycotoxin Limits in Feeds
The main correlation characteristics between the mold and yeast populations and detected mycotoxins in corn silages were not statistically significant; however, the coefficient of determination between the May and FUMB1 (
r2) was relatively high (0.79%) (
Table 7). This was also confirmed by Barug et al. [
83], where a direct relationship between the microscopic fungi and yeast and certain mycotoxins in silage was not observed. Similarly, no correlations were found between fungal DNA and mycotoxin concentrations [
84]. According to Schenck et al. [
85], a correlation between the presence of
Fusarium toxins (NIV, DON, 3-ACDON, HT-2, T-2, BEAU, and ENNB) and the presence of
Fusarium culmorum,
F. equiseti,
F. graminearum, or
F. poae could not be proved. However, there were negative significant correlations between the nivalenol and enniatin A1 (
p < 0.01) and moniliformin (
p < 0.05). On the other side, the positive correlation between the enniatin A1 and moniliformin (
p < 0.05) was observed.
Aspergillus, Fusarium, and
Penicillium species comprise a well-known group of microscopic filamentous fungi that are infamous for their ability to make many potent mycotoxins. Mycotoxins play a significant role in the defensive strategies of mycotoxigenic fungi. The fungal species more frequently identified in this work have been previously reported in silage [
86]. The presence of mycotoxins, which are produced by
Aspergillus and
Penicilium species, was not evaluated in corn silages in this study.
Aflatoxins are mainly produced by toxigenic strains of
Aspergillus molds. In ruminants, reduced milk production in dairy cows, decreased milk quality and safety due to carry-over of toxins from contaminated feed, liver malfunctions, decreased feed efficiency and rate of gain in beef cows, and compromised immune and ruminal functions were observed [
86]. The maximum content of aflatoxin B1 for complete foodstuffs for cattle, sheep, and goats is 0.02 mg/kg with the exception of complete foodstuffs for dairy animals (0.005 mg/kg) and for calves and lambs (0.01 mg/kg). The maximum content of aflatoxin B1 for complementary foodstuffs for cattle, sheep, and goats is 0.02 mg/kg (except complementary foodstuffs for dairy animals, calves, and lambs: 0.005 mg/kg) [
87].
Ochratoxins are produced by several
Penicilium and
Aspergillus species, have hepatotoxic and nephrotoxic effects, causing poor feed conversion and limiting weight gains in ruminants [
88]. The guidance limit of ochratoxin A for feed materials (cereals and cereal products) is 0.25 mg/kg, and for complementary and complete foodstuffs are limited only for poultry and pigs [
89].
In analyzed corn silages, only fusarium mycotoxins were found. Deoxynivalenol causes feed refusal and lower weight gains, diarrhea, lower milk production, hepatotoxicity (in young preruminants), and immune alterations [
86,
88,
90,
91]. The regulatory level (guidance value) of deoxynivalenol for complementary and complete foodstuffs for ruminants is 5 mg/kg. For feed materials, guidance values are 8 mg/kg (for cereals and cereal products) and 12 mg/kg (for maize by-products) [
89].
Clinical signs in ruminants caused by fumonisins are decreased feed intake, milk production, and mild liver diseases [
86,
92]. The results of Roberts et al. [
93] revealed that exposure to deoxynivalenol and fumonisins was detrimental to the welfare of finishing steers and may compromise their ability to withstand other stressors such as disease, heat stress, or other toxins. The regulatory level (guidance value) of fumonisins (B1 + B2) for complementary and complete foodstuffs for adult ruminants (>4 months) is 50 mg/kg, and for feed materials (maize and maize products) is 60 mg/kg [
89].
Higher zearalenone contamination is linked with a risk of vaginal and rectal prolapses, infertility, hyperestrogenism, swelling of mammary glands, and milk production reduction in dairy cows [
88,
94]. The regulatory level (guidance value) of zearalenone for complementary and complete foodstuffs for calves, dairy cattle, sheep (including lambs), and goats (including kids) is 0.5 mg/kg. For feed materials, guidance values are 2 mg/kg (for cereals and cereal products) and 3 mg/kg (for maize by-products) [
89].
Nivalenol belongs to the B group of trichothecene mycotoxins along with deoxynivalenol, 3-acetyl-deoxynivalenol, 15-acetyl-deoxynivalenol, and fusarenon-X [
95]. Generally, the toxicity of deoxynivalenol and nivalenol is similar [
88,
96] and effects of nivalenol include immunotoxicity and hematotoxicity. With the exception of forage maize (and maize silage), levels of nivalenol in forages are generally low. For lactating dairy cows and beef cattle, the estimated lower-bound and upper-bound exposures to nivalenol are between 0.077 and 0.69 µg/kg of live body weight per day, except for maize-silage-based diets (1.9 and 4.6 µg/kg body weight per day) [
95]. The concentrations of regulated mycotoxins in our experiment did not exceed the maximum permitted, guidance, and daily tolerable intake according to the limits [
87,
89,
95,
97].
Animal exposure to enniatins and beauvericin is primarily from feed intake of cereal grains and cereal by-products [
97]. The primary toxic action of beauvericin and enniatins is related to their ability to form ion channels and transport NH
4+ or K
+ ions across the cell membrane, resulting in disturbance of the ion homeostasis and eventually cell death [
98,
99].
Enniatins are mutagenic and embryotoxic for animals [
100,
101]. For the sum of enniatins, the calculated lower-bound and upper-bound (UB) chronic exposures for ruminants ranged from 3.30 to 8.26 μg/kg body weight per day and estimated acute UB exposure is 32.6 μg/kg body weight per day, for ruminants [
97].
Moniliformin is mainly detected in cereal grains and cereal-based feed [
102]. The main pathological change observed in sheep was the degeneration of the proximal tubules of the kidneys after moniliformin intake [
103]. No toxicity data suitable for hazard characterization of moniliformin were identified for ruminants, farmed rabbits, horses, farmed fish, dogs, and cats. Therefore, the EFSA Panel on Contaminants in the Food Chain (CONTAM) [
104] considered of 0.20 mg moniliformin/kg body weight identified for pigs as an indicative reference point [
102].


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}