
Taxonomic Uncertainty and Its Conservation Implications in Management, a Case from Pyrus hopeiensis (Rosaceae)


1
College of Ecology and Nature Conservation, Beijing Forestry University, Beijing 100083, China
2
College of Biological Sciences and Technology, Beijing Forestry University, Beijing 100083, China
3
Centre of Pear Engineering Technology Research, State Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing Agricultural University, Nanjing 210095, China
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(6), 417; https://doi.org/10.3390/d14060417
Submission received: 6 May 2022
/
Revised: 22 May 2022
/
Accepted: 23 May 2022
/
Published: 24 May 2022
(This article belongs to the Special Issue Ecology, Evolution and Diversity of Plants)
Abstract
:Improved taxonomies and phylogenies are essential for understanding the evolution of organisms, the development of conservation plans, and the allocation of funds and resources, especially for threatened species with uncertain identities. Pears are an economically and nutritionally important fruit, and wild pear species are highly valued and protected because of their utility for the development of cultivars. Pyrus hopeiensis is an endangered species endemic to North China, which is sympatric with and difficult to distinguish from the widely distributed and morphologically similar species P. ussuriensis. To clarify its taxonomic identity, principal coordinate analysis was performed using 14 quantitative and qualitative characters from P. hopeiensis, P. ussuriensis, and P. phaeocarpa, and phylogenomic analysis was performed based on whole-genome resequencing and whole plastome data. Pyrus hopeiensis was synonymized with P. ussuriensis based on morphological and phylogenetic evidence, as well as our long-term field studies. Pyrus hopeiensis is proposed to be excluded from the list of local key protected wild plants. Given that the holotype of P. ussuriensis was not designated, a lectotype was designated in this work. Integrative evidence-based taxonomic study including museomics is suggested for organisms with uncertain identities, which will contribute to biodiversity conservation.
1. Introduction
The intensification of human activities, especially since the Industrial Revolution, has negatively affected global climate and biodiversity and is responsible for the Sixth Mass Extinction [1]. Rare and endangered plants are important components of biodiversity, and many of them are facing high extinction pressure because of inadequate conservation management [2], as well as taxonomic uncertainty [3,4]. Accurately identifying plants is a huge challenge, and taxonomic stability is an elusive goal [5,6]. Although studies of the evolution and taxonomic identity of species can greatly contribute to our knowledge of the organisms that we are interested in protecting, taxonomic problems might occur because of improvements in our knowledge of the phylogeny and evolution of organisms, as well as the recognition of previously made nomenclatural errors [7]. More than half of economically important tropical African ginger specimens from 40 herbaria in 21 countries are likely to be wrongly named [8], which can affect the conservation status assessment of related species. Haematocarpus subpeltatus Merr. (Menispermaceae), a misidentified critically endangered species that has received much conservation focus, has been identified as a new record in China, but the real entity that should be given priority for conservation, Eleutharrhena macrocarpa (Diels) Forman, has been clarified by molecular phylogenetic analyses [9]. These findings highlight the significance of taxonomy and molecular phylogeny in biodiversity conservation, especially for species with taxonomic uncertainty.
Pears are an economically and nutritionally important temperate fruit that have been cultivated for more than 3000 years [10]. The genus Pyrus L. (Rosaceae) is geographically divided into oriental and occidental pears [11]. Considerable morphological variation and extensive hybridization have been documented in the genus [12,13]. The number of species recognized in Pyrus varies from 21 to more than 80 [11,14], and 73 of them are abundant in Eurasia [15]. Although remarkable advances have been made in the genetics and breeding of Pyrus in recent years, and whole genomes of several species have been published, e.g., [10,13,16,17,18], the taxonomy, diversification and phylogeny of Pyrus remain unclear, several currently accepted species such as P. caucasica Fed., P. pyraster (L.) Burgsd. and P. spinosa Forssk. are not monophyletic in the phylogenetic trees [12,19], suggesting the necessity of further taxonomic and phylogenetic studies. One reason for the lack of robustness of the molecular trees of Pyrus in previous studies is the few numbers of molecular markers employed. Whole nuclear genome sequences can provide vital information for reconstruction species’ phylogenies [20], and complete plastomes can also provide key information for phylogenetic studies as well as for detecting hybridization events [21,22]. A hybrid usually neighbors one of its parents in the nuclear tree and group with another parent in the plastome tree, and phylogenetic discordance (cytonuclear discordance) is usually detected for hybrid species. Hence, extensive phylogenomic analyses are necessary for advancing our understanding of the phylogeny of Pyrus. This will shed light on the taxonomy and evolution of pear species and enhance pear production.
China is a diversity center for oriental pears, as 14 native species, including 5 primary wild species and over 2000 cultivars, have been reported to date [13,23,24]. Among them, Pyrus hopeiensis Yü, a taxonomically controversial species endemic to North China, is thought to be a hybrid of P. ussuriensis Maxim. and P. phaeocarpa Rehd. [25]. It is classified as an endangered species and is listed as a key protected wild plant of Hebei Province; it has thus received much conservation attention, e.g., [26,27,28,29]. Pyrus hopeiensis can be distinguished from P. phaeocarpa from several morphological characteristics, such as its persistent calyx and spiny serrate leaf margin (vs. caducous calyx and serrate leaf margin in P. phaeocarpa). However, differentiating P. hopeiensis and P. ussuriensis is more challenging. Pyrus ussuriensis is an important germplasm resource that is widely distributed in northern China, the Russian Far East, North Korea, and Japan, and more than 150 cultivars have been obtained from this species in China [30]. Pyrus hopeiensis has been documented in Beijing, Hebei, and Shandong provinces, and its type locality is Jieshi Mountain, a small hill with an altitude of 695 m in Changli County, Hebei Province, which is in the eastside of Yan Mountain in North China [23,25,31]. Based on the original description [25], P. hopeiensis can be distinguished from P. ussuriensis by its brown fruit and obvious spots on its surface (vs. yellow fruit with fewer spots). However, numerous individuals with intermediate morphological characteristics have been identified in the field and herbaria. Taxonomic ambiguity should be clarified, as this would aid conservation efforts for endangered and regional protected species. Hence, further morphological and phylogenomic studies should be performed to unravel the identity of P. hopeiensis.
In the present study, we aim to: (1) assess morphological differences among P. hopeiensis, P. phaeocarpa, and P. ussuriensis, (2) reconstruct the molecular phylogenetic tree and test the hybrid identity of P. hopeiensis based on whole genome resequencing data, and (3) clarify the taxonomic identity of P. hopeiensis. This study provides novel insight for the conservation management of Pyrus species and highlight the importance of plant taxonomy and phylogenomic analysis for biodiversity conservation.
2. Materials and Methods
2.1. Morphological Study
All specimens of P. hopeiensis (including type specimens) deposited in the herbarium of the Institute of Botany, Chinese Academy of Science (PE) and hundreds of specimens of P. ussuriensis and P. phaeocarpa in PE were examined (Appendix A). Fourteen qualitative and quantitative characters were selected (Table 1), which included 1 binary and 13 continuous characters. Given differences in leaf morphology during the flowering and fruiting period in Pyrus, morphological analyses were performed for both periods. Aside from one specimen collected from an individual cultivated at PE (Ren 2, 21 August 1962), only specimens of P. hopeiensis identified by Yü and Ku were used to avoid misidentification. Two data matrices were made for 45 flower specimens and 61 fruit specimens, and each was treated as an operational taxonomic unit. Following Wang [32], the data matrix was standardized using a zero-mean normalization method. The formula X* = (X − m)/s was used, where “X” is the sample, “m” is the arithmetic mean, and “s” is the standard deviation. A principal coordinate analysis was performed based on the Gower general similarity coefficient analysis for mixed data sets using MVSP-Version 3.13b software.
2.2. Phylogenomic Inference
In this study, whole-genome resequencing data of 27 samples of Pyrus representing 13 species from [33,34] were used for phylogenomic inference. Specifically, a total of 22 samples from Wu et al. [33] were employed, which included three individuals of P. phaeocarpa, five P. ussuriensis, and two P. hopeiensis. Five samples of P. hopeiensis (PWH8, PWH11, PWH18, PWH19, and PWH20) were used from Li et al. [34]. All samples were numbered following [33,34]. Malus × domestica L. was used as the outgroup, and its whole-genome sequence [35] was downloaded from NCBI.
Raw reads were filtered by removing several types of low-quality paired reads, including reads with adapters, paired reads with N content greater than 10%, and low-quality (Q < 10) paired reads contained in single-end sequencing reads that exceeded 50% of the length of the read. The accessions were then mapped to the apple genome using the MEM algorithm of Burrows–Wheeler Aligner in BWA software [36], and single nucleotide polymorphisms (SNPs) were called using the HaplotypeCaller module in GATK [37] and filtered using the following parameters: QD < 2.0‖MQ < 40.0‖FS > 60.0‖QUAL < −12.5‖ReadPosRankSum < −8.0—clusterSize 2—clusterWindowSize 5. The obtained SNPs were further filtered to construct a high-quality data matrix for phylogenetic inference with the minor allele frequency greater than 0.05, and less than 0.8 missing rates of the confirmed credible genotype from all accessions for each site and biallelic SNPs. Finally, a maximum likelihood (ML) phylogenetic tree was generated using IQTREE [38], and ultrafast bootstrap support values (BSML) were estimated with 1000 replicates [39].
A total of sixteen complete plastomes of Pyrus species were downloaded from NCBI, including one for each of the three species. Malus kansuensis (Batal.) Schneid. was used as the outgroup. Four samples of P. hopeiensis (PWH8, PWH11, PWH18, and PWH20) were selected from [34], one P. ussuriensis (1) was selected from [33]. The whole chloroplast genomes of these five samples were generated from the whole-genome resequencing data using the software GetOrganelle [40]. Plastomes of all samples were aligned by MAFFT [41]. After cutting both sides of the aligned sequences, a final data matrix with a length of 162,337 bp was obtained and used for phylogenetic analysis in IQTREE.
3. Results
3.1. Morphological Study
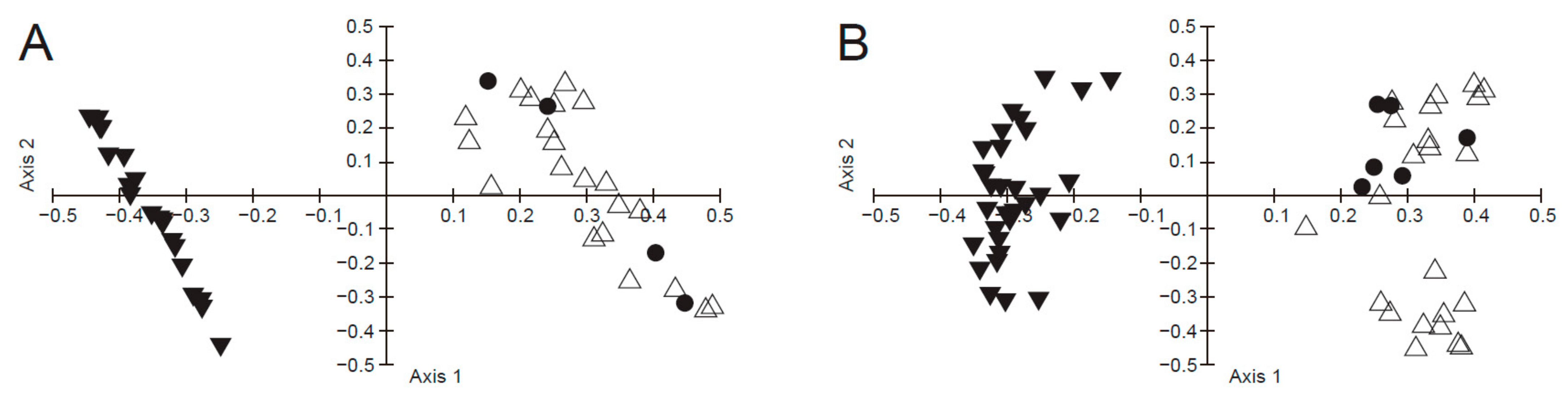
The results of principal coordinate analysis based on multiple characters showed that the four flower specimens of P. hopeiensis clustered with P. ussuriensis but were clearly separated from the P. phaeocarpa cluster (Figure 1A). Likewise, the six fruit specimens of P. hopeiensis were nested within the P. ussuriensis cluster, and this mixed group was distinct from that of P. phaeocarpa (Figure 1B).
3.2. Phylogenomic Inference
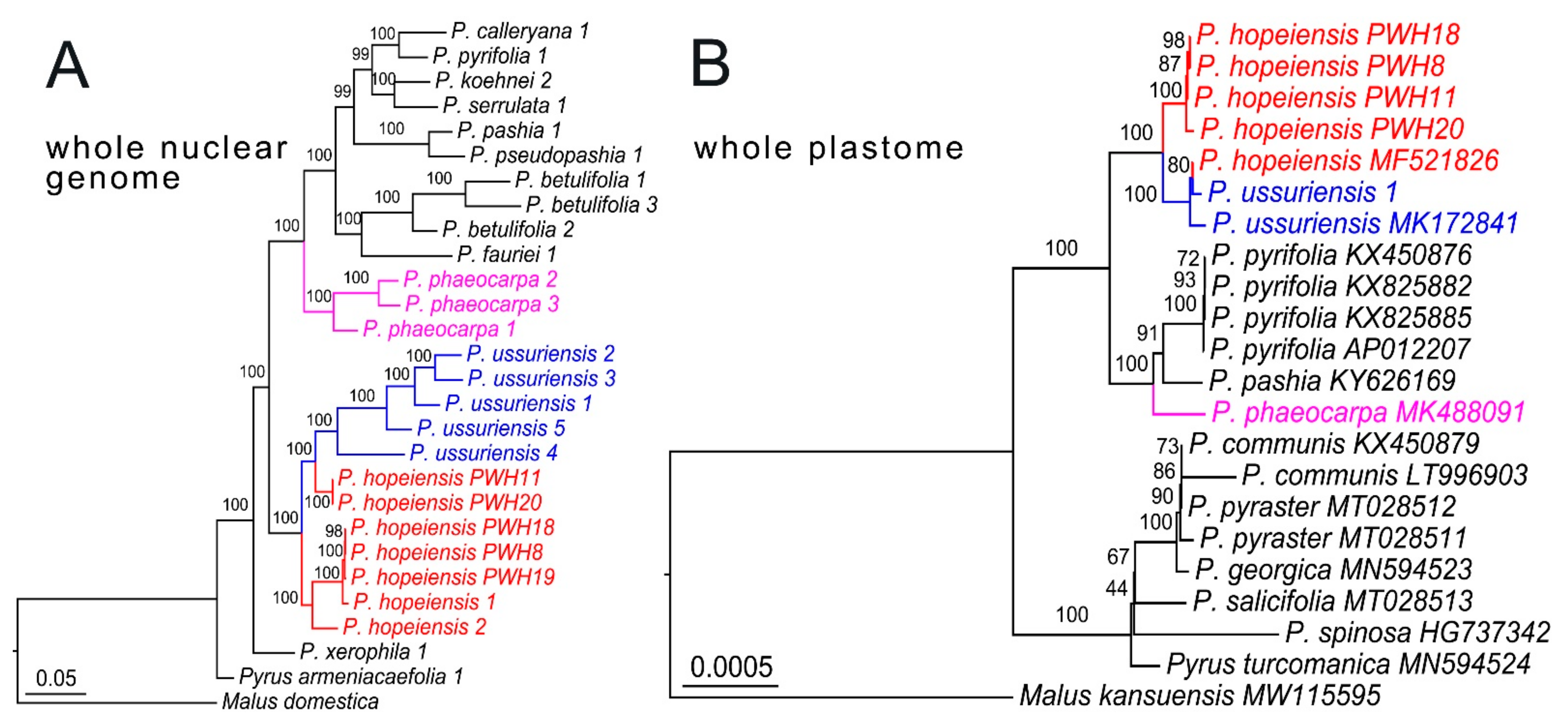
After filtration, a total of 204.87 G clean data were obtained from whole-genome resequencing data, and a high-quality SNP data matrix with a length of 1,340,483 bp was generated. The ML tree was well resolved, and the seven samples of P. hopeiensis were divided into two groups. Two of them (PWH11 and PWH20) were grouped with five samples of P. ussuriensis, forming a monophyletic clade with full support (BSML = 100, Figure 2A); this clade was sister to another fully supported monophyletic clade that comprised the rest of the P. hopeiensis samples (PWH8, PWH18, PWH19, 1 and 2). The three samples of P. phaeocarpa comprised a fully supported monophyletic clade (BSML = 100, Figure 2A), and this clade was distinct from the P. ussuriensis–P. hopeiensis clade. Similar results were obtained in the plastome tree, with samples of P. hopeiensis and P. ussuriensis combined into a fully supported monophyletic clade (BSML = 100, Figure 2B). A sample of P. hopeiensis from Shandong Province [28] was embedded within the subclade of P. ussuriensis. The clade of P. phaeocarpa was distinct from the P. ussuriensis–P. hopeiensis clade, and no phylogenetic discordance was detected among these three species.
4. Discussion
4.1. Integrative Evidence for Reappraisal of the Identity of Pyrus hopeiensis
Morphological characters are important for species identification, and some morphological characteristics vary greatly among habitats and at different growth stages; this can result in inconsistencies in the designation of the taxonomic identity of species. The color of fruit and the spots on the surface of fruit are key morphological characteristics for discriminating between P. hopeiensis and P. ussuriensis [25]. The holotype (PE00020671, Figure 3A), isotype (PE00020670), and paratypes (FS Chang 238, 269, 276) of P. hopeiensis were collected in September 1953; this species was described 10 years later [25]. Although the fruit was described as brown in the original paper (Pomum …, fuscum, …) [25] (p. 232), we found no fruit in these type specimens. However, the fruit is described as “mud-like yellow” (in Chinese) in the collection record of paratype FS Chang 269 (Figure S1). Several brown or black fruits are present on two specimens collected in Shandong Province (Zhou et al. 1178, 1590) on 6 and 7 June 1959 and identified as P. hopeiensis by Yü. Although “green fruit” is written clearly in Chinese on the record of Zhou et al. 1178, these immature pomes turned black in herbaria (Figure S2). Hence, the fruit color of P. hopeiensis recorded in previous studies (brown) was not consistent with that of individuals observed in the field (yellow).
We conducted a long-term field investigation in North China since 2010. The fruit color of P. ussuriensis recorded in previous studies (yellow) is unstable. A dried fruit preserved in the lectotype of P. ussuriensis (Figure 3B) is pale brown. The color of mature fruit varies from greenish yellow to reddish yellow in the field (Figure 3C–E). One specimen collected by P. Zhang (742) on 13 September 1956 from Changli County, Hebei Province (Figure S3A) has a mature fruit that is reddish in color. Another specimen (P. Zhang 766, Figure S3B) presents three densely spotted fruits, and the fruit color (brown) is clearly indicated on the collection record (in Chinese). These two specimens were both identified by Yü as P. hopeiensis on 29 April 1960. The density of spots on the fruits both varies greatly within and among individuals; for example, there are several spots in Figure 3C (Dongling Mountain, in the north end of Taihang Mountain) and 3D (Yudu Mountain, in the west of Yan Mountain), but almost none in Figure 3E (Song Mountain, neighboring Yudu Mountain). Furthermore, persistent and caducous calyxes associated with mature fruits were observed in the same trees in the field (e.g., Figure 3B). Carpel number (4 vs. 5) and the length of the fruit pedicel (long vs. short) are sometimes used to differentiate P. hopeiensis from P. ussuriensis. However, our morphological analyses of specimens (Figure 1) and our field observations (Figure 3C–I) indicate that this trait has no taxonomic relevance.
We conducted two field investigations at the type locality—Jieshi Mountain, Xingshuyuan Village, Changli County, Hebei Province—at the end of August in 2015 and 2017. This is a small hill like the others located in the Yan Mountains: its altitude is 695 m, and it is 15 km from the Bohai Sea. Besides P. hopeiensis and P. ussuriensis documented by Li et al. [34], we found several individuals of P. phaeocarpa and P. betulifolia. The calyx was absent in mature fruit of these two species; fruits of the former are approximately 2 cm in diameter (vs. <1 cm in the latter), and the carpel number is 3–4 (vs. 2–3) (Figure 4). In 2015, we met an employee of the local forestry bureau who identified an individual as the endemic species, P. hopeiensis (Figure 4A). In Figure 4, fruits Aa, Bb, and C were collected from this individual. Key morphological characters (such as calyx on fruit, carpel number and leaf margin) indicate that this individual is instead P. phaeocarpa. Aside from a large number of wild Pyrus individuals, the Jieshi Mountain type locality includes a large orchard that contains Pyrus cultivars. One of the P. betulifolia trees growing here has fruits with an unusual green color (Figure 4B(c),D). This special germplasm resource has also been discovered in Shanxi Province [42]. Unfortunately, this individual in the Jieshi Mountain died because of the growth of Pueraria montana (Loureiro) Merrill (Fabaceae) during our visit in 2017.
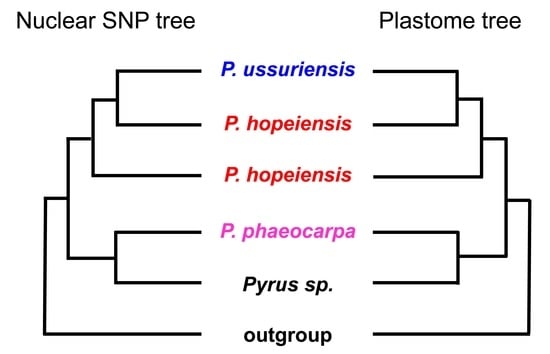
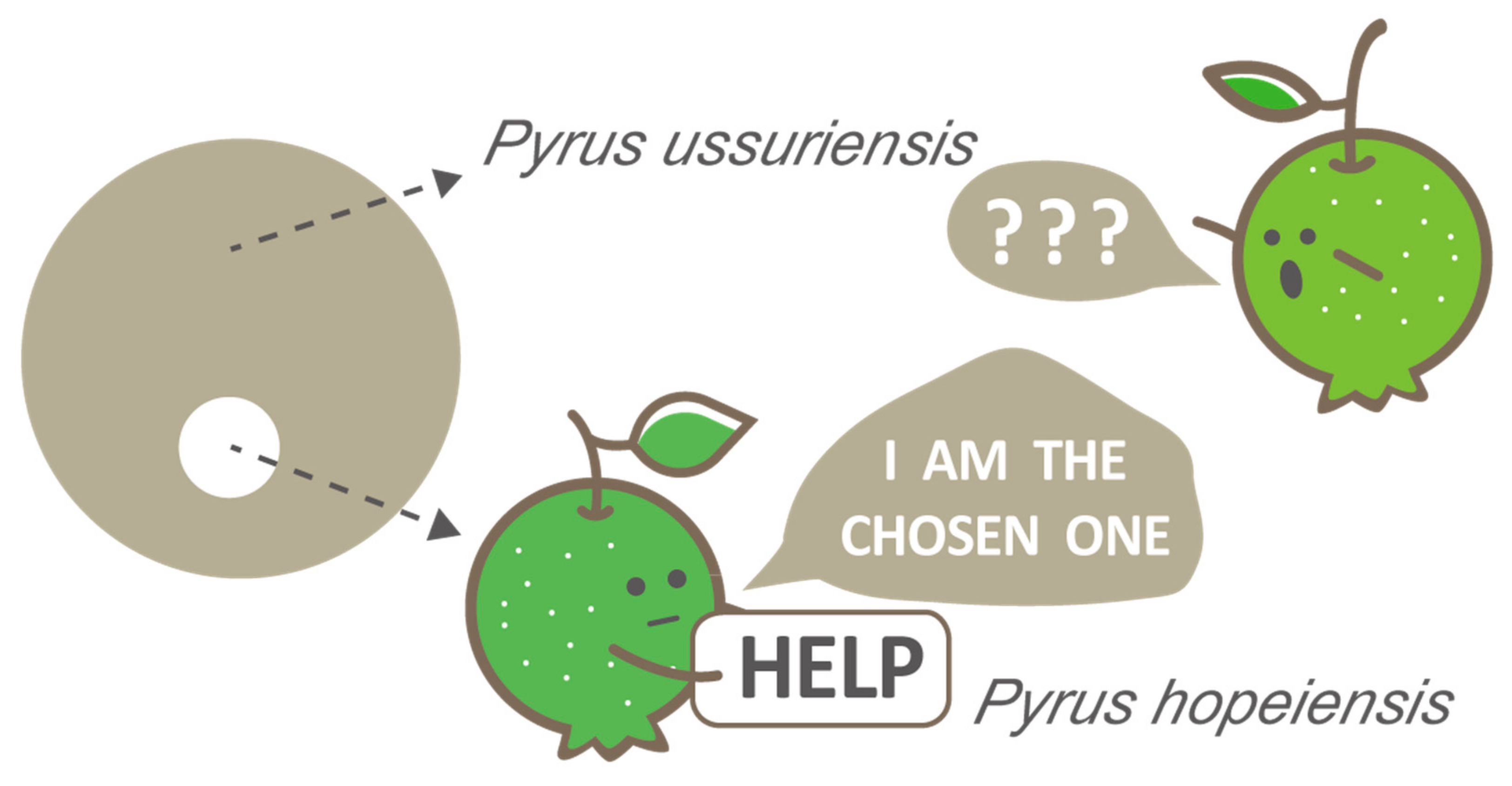
Given the uncertain morphological differences between P. hopeiensis and P. ussuriensis, a phylogenetic study provides a powerful tool for clarifying the relationships between these taxa. However, the high level of genetic exchange and hybridization in Rosaceae at both the species and genus level [34,43,44,45] precludes the resolution of phylogenetic relationships among Pyrus species when traditional molecular markers are used, e.g., [12,19,46]. With whole-genome resequencing data, a robust molecular tree of Pyrus was constructed based on the SNP data matrix [33]. Two individuals of P. hopeiensis collected from the Zhengzhou Fruit Research Institute, Chinese Academy of Agricultural Sciences (CAAS), and Research Institute of Pomology, CAAS, were sampled. Both were embedded within a fully supported monophyletic clade of P. ussuriensis, and they were distinct from the P. phaeocarpa clade. We performed phylogenomic inference in this study based on the whole-genome resequencing data of Wu et al. [33] and Li et al. [34]. Our results demonstrate that all the P. hopeiensis samples were mixed with P. ussuriensis, and they together comprised one fully supported monophyletic clade (Figure 2). However, no cytonuclear discordance was detected for these three pear species between the nuclear and plastid genome data. Hence, a hybrid origin of P. hopeiensis from P. ussuriensis and P. phaeocarpa was not supported in this study. Both the co-existence of P. ussuriensis and P. hopeiensis at the same locations and their synchronous flowering and fruiting phenology suggest that there is no niche differentiation between them. Hence, evidence from morphological comparisons, phylogenomic inference, and our extensive field studies indicate that the locally endemic and protected P. hopeiensis is a synonym of the widely distributed P. ussuriensis. Consequently, ‘the chosen one’ of wild pear species (P. hopeiensis), which has received multiple conservation resources, should be excluded from the list of local key protected wild plants (Figure 5).
4.2. Integrative Evidence-Based Taxonomy for Biodiversity Conservation
Climate change and biodiversity loss are two critical events of the Anthropocene, and the existence and distribution of biodiversity need to be better documented prior to proposing conservation strategies [47,48]. Although changes in the names of plants can be frustrating for conservationists, taxonomy is a science that involves classifying continuous variation in discrete categories and clarifying the phylogenetic relationships among taxa; taxonomists also have the responsibility of removing unnatural taxa that conflict with evolutionary and phylogenetic theory [49]. Species’ names should change when a more rational and testable hypothesis of the species is produced based on more evidence [50]. For example, Juglans hopeiensis Hu (Juglandaceae), a species endemic to North China and listed as a key protected wild plant of Hebei Province, is thought to be a hybrid of J. regia L. and J. mandshurica Maxim. [51]. Whole plastome data suggested that it was closely related to J. mandshurica, whereas the reduced-representation genomic data indicated a close relationship with J. regia [21]. Based on dense sampling whole-genome data, Zhang et al. [52] confirmed its hybrid identity and demonstrated that only the first generation of J. hopeiensis is viable, which precludes further speciation. On the one hand, the accurate identification of endangered species is a prerequisite before conservation action can be taken. On the other hand, species incorrectly listed as endangered species can result in a waste of valuable resources, and the prime of this are P. hopeiensis and E. macrocarpa [9]. Thus, integrative evidence is needed to assess the identity of controversial species, and these approaches should figure prominently in conservation management.
Because of limited resources, setting priorities for the conservation of threatened species is critically important. Species with unresolved taxonomies can pose problems for endangerment status assessments and impede conservation management; genetic data in the genomic era can provide information that can help clarify the taxonomic status of species [53]. In addition to living organisms, specimens preserved in museums and herbaria can provide valuable morphological and ecological data as well as genomic resources that can be used to resolve taxonomic uncertainties, reconstruct accurate phylogenies, and assess the mechanisms of ecological adaptation [54]. In some cases, type specimen may be the only credible remnant available for species identification. Hence, genetic resources from voucher specimens and even type specimens are vital for accurate identification, taxonomic designations, and conservation [55]. Although extracting DNA from museum specimens can be a major challenge and potentially be destructive [56,57,58], the growth of museomics, the study of DNA sequences obtained from museum specimens, combined with phenetic data collected from key specimens, will provide new taxonomic, evolutionary, and conservation insights.
4.3. Taxonomic Treatment
Based on the morphological comparison of specimens, phylogenomic inference, and our field studies, P. hopeiensis is a synonym of P. ussuriensis. The holotype of P. ussuriensis was not previously designated. Although several specimens are deposited in herbaria (e.g., GH, K and P), one of them preserved in LE that fit the original description of P. ussuriensis was designated as the lectotype.
Pyrus ussuriensis Maxim. ex. Rupr., Bull. Acad. Imp. Sci. Saint-Pétersbourg, sér. 2. 15: 132. 1857.—Lectotype (designated here): Russia, CJ Maximowicz s.n. (LE, barcode LE01009639, [photo!]; isolectotype: LE barcode LE01009638, [photo!]). = Pyrus hopeiensis Yü, Acta Phytotax. Sin. 8: 232. 1963. syn. nov.—Type: China, Hebei Province, Changli County, Xingshuyuan village, 21 September 1953, FS Chang 268 (PE barcode 00020671!; isolectotype: PE barcode 00020670!).
Other Specimens Examined
Pyrus ussuriensis Maxim. ex. Rupr.: RUSSIA. Ussuri: CJ Maximowicz 96 (K barcode K000758073 [photo]), CJ Maximowicz s.n. (GH barcode GH00032508 [photo], LE barcode LE01009633 [photo], LE01009634 [photo]). CHINA. Beijing: Changping, 4 May 2001, Changping Investigation Team 173 (PE barcode 01274862!), Haidian, June 1960, Anonymous s.n. (PE barcode 01274861!), 21 August 1962, G Ren 2 (PE barcode 01608659!), Jiangou, 25 April 1959, Hebei Investigation First Team 116 (PE barcode 00548756!), Mentougou, Xiaolongmen, July, 1978, Beijing Normal University Team s.n. (PE barcode 01274865!), Shangfangshan, 21 May 1959, Hebei Investigation First Team 341 (PE barcode 00548750!); Hebei Province: Changli County, Xingshuyuan village, 1 Sept. 1953, FS Chang 238 (PE barcode 00548753!), 21 Sept. 1953, FS Chang 269 (PE barcode 00548751!), 24 Sept. 1953, FS Chang 276 (PE barcode 00548755!), 13 September 1956, P Zhang 742 (PE barcode 01448598!), Chicheng County, Zhenanpu town, 7 September 1959, Anonymous 4100 (PE barcode 00548752!, 01274860!), Funing County, 24 April 1959, Anonymous 58 (PE barcode 00548760!), Laiyuan County, 26 July 1959, Anonymous 4-3743 (PE barcode 00548754!), Yi County, 24 July 1959, Anonymous 3246 (PE barcode 01274859!), 25 July 1959, Anonymous 3287 (PE barcode 01274858!), Yu County, Xiaowutai Mountain, 27 August 1950, YW Cui 2706 (PE barcode 01274864!), 28 August 1950, YW Cui 2864 (PE barcode 01274863!); Inner Mongolia Autonomous Region: Zhaomengkaqi, Xiaoxigoumen, 4 June 1962, Meng-Ning Comprehensive investigation Team 252 (PE barcode PE00548749!), 4 Aug. 1962, Meng-Ning Comprehensive investigation Team 1292 (PE barcode PE00548748!); Ningxia Autonomous Region: Helan Mountain, 25 May 1959, YQ He 02424 (PE barcode 00548759!); Shandong Province: Qingdao City, Laoshan, 7 June 1959, TY Zhou et al. 1590 (PE barcode 00548758!), 8 June 1959, TY Zhou et al. 1178 (PE barcode 00548757!); Shanxi Province: Xing County, Yellow River Investigation Second Team 2695 (PE barcode 00548761!).
5. Conclusions
Plant names may change because of the development of taxonomic studies, which can aid biodiversity conservation. Because of frequent gene exchange and hybridization among species, the phylogenetic relationships among Pyrus species remain unclear, and the taxonomic identities of several species still require examination, such as P. hopeiensis, a regional key protected and potential hybrid species endemic to North China. Comprehensive morphological studies were performed based on flowering and fruiting specimens of P. hopeiensis and its potential parent, P. ussuriensis, and P. phaeocarpa. Extensive phylogenomic inferences were performed based on a high-quality SNP data matrix that was generated from whole-genome resequencing and the whole plastome data. A long-term field investigation of Pyrus species in North China was also conducted. Both morphological and phylogenomic studies indicate a close relationship between samples of P. hopeiensis and P. ussuriensis, and this mixed cluster consists of a fully supported monophyletic clade and is distinct from other species. Hence, P. hopeiensis is a synonym of P. ussuriensis. The lectotype of P. ussuriensis was designated here. Our work provides information for both the taxonomic study and conservation management of Pyrus and highlights the significance of integrative evidence-based taxonomy in biodiversity conservation.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d14060417/s1, Figure S1. Paratype of Pyrus hopeiensis. FS Chang 269, PE barcode: 00548751. Figure S2. Specimen of Pyrus hopeiensis. TY Zhou et al., 1178, PE barcode: 00548757. Figure S3. Specimens of Pyrus hopeiensis. A: P Zhang 742 (PE barcode: 01448598), B: P Zhang 766 (PE barcode: 01449638).
Author Contributions
X.-Y.M. conceived the work, performed field investigation, and prepared, wrote, and revised the manuscript; J.W. (Jiang Wu) carried out specimens’ morphological statistics and the principal coordinate analysis; J.W. (Jun Wu) provided the original resequencing data of Pyrus species described in the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Science Foundation of China (grant number 32070235), the Project of the Second Investigation of the National Key Protected Wild Plants of Beijing, and the Project of Agricultural Wild Plant Resources Investigation of Beijing (grant number 13200346).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data are available in this article.
Acknowledgments
We thank Irina Illarionova (LE) for providing photos of the type specimen of Pyrus ussuriensis, and the curator and staff at PE for their help. We also thank Qin-Wen Lin, Yi-Xuan Zhu, Xue-Li Shen, Ling Tong, Wan-Jie Jiang, and De-Huai Zhang for their help during our field investigation.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
The information of specimens used for principal coordinate analysis given in the Appendix is taken from specimens deposited in PE.
Pyrus hopeiensis Yü: CHINA. Beijing: Changping District, 4 May 2001, Changping Investigation Team 173 (PE barcode 01274862), Haidian, June 1960, Anonymous s.n. (PE barcode 01274861), 21 August 1962, G Ren 2 (PE barcode 01608659), Jiangou, 25 April 1959, Hebei Investigation First Team 116 (PE barcode 00548756), Mentougou, Xiaolongmen, July, 1978, Beijing Normal University Team s.n. (PE barcode 01274865); Hebei Province: Changli County, Xingshuyuan village, 13 September 1956, P Zhang 742 (PE barcode 01448598), 19 September 1956, P Zhang 766 (PE barcode 01449638), 22 September 1956, P Zhang 782 (PE barcode 01449639, 01449640), Funing County, 24 April 1959, Anonymous 58 (PE barcode 00548760).
Pyrus phaeocarpa Rehd.: CHINA. Beijing: Haidian District, Cheying village, 22 August 1972, Anonymous Hai-676 (PE barcode 01204095), Mentougou District, Dongyangtuo village, 19 April 1954, ZL Yan 167 (PE barcode 01448580), Mengwu village, 20 April 1954, ZL Yan 171 (PE barcode 01448530); 20 August 1954, ZL Yan 168 (PE barcode 01448527); Hebei Province: Changli County, Fenghuangshan, 21 April 1953, TT Yü 35 (PE barcode 01448549, 01448550), near Railway Station Office, 27 April 1953, TT Yü 092 (PE barcode 01448535), Xishan, Anonymous s.n. (PE barcode 01448554), Xingshuyuan, 17 April 1953, TT Yü 13 (PE barcode 01448538), 20 April 1953, TT Yü 30 (PE barcode 01448555), TT Yü 227 (PE barcode 01448609, 01448611), West Zhanggezhuang, Anonymous 14 (PE barcode 01448534), 16 September 1953, Anonymous 6 (PE barcode 01448532), Anonymous 9 (PE barcode 01448540), 24 April 1956, P Zhang 647 (PE barcode 01448561, 01448562), 13 September 1956, P Zhang 738 (PE barcode 01448567), P Zhang 739 (PE barcode 01448560), P Zhang 739 (PE barcode 01448569), P Zhang 740 (PE barcode 01448569), P Zhang 741 (PE barcode 01449634), Liugezhuang, 6 October 1953, FS Chang 284 (PE barcode 00549049), Dongling, 22 April 1936, DF Jin K-901 (PE barcode 00549045, 00549062), Funing County, Jiejia village, 22 September 1956, FS Chang 776 (PE barcode 01448557, 01448558), FS Chang 777 (PE barcode 01448559), FS Chang 780 (PE barcode 01448575), Liugezhuang, 6 October 1953, FS Chang 283 (PE barcode 00549046), 19 September 1956, P Zhang 766 (PE barcode 01449638), Shibeigou, 24 April 1953, TT Yü 58 (PE barcode 00549044), Yuanjiagou, 23 April 1953, TT Yü 53 (PE barcode 01448543, 01448544), Qinghuangdao City, chitushan, 4 May 1953, TT Yü 141 (PE barcode 01448545); Henan Province: Song County, 15 September 1956, Bureau of Henan Forestry 986 (PE barcode 00549057), Bureau of Henan Forestry 1259 (PE barcode 00549055, 00549058), 26 August 1959, Anonymous 35149 (PE barcode 00549054), 1 October 1959, Bureau of Henan Forestry 1259 (PE barcode 00549056); Gansu Province: Wushan County, 4 June 1956, Yellow River Investigation Team 4480 (PE barcode 00549059); Jiangxi Province: Guangchang County, 7 October 1958, QM Hu 5398 (PE barcode 00549061), Suichuan County, 22 September 1963, JS Yue et al. 4168 (PE barcode 00549060); Shaanxi Province: Muhuguan, 22 June 1960, Anonymous 0496 (PE barcode 01274917, 01274918), Shangzhou City, 4 July 1960, Anonymous 1074 (PE barcode 01274921), Shanyang County, 12 May 1963, ZY Zhang 15906 (PE barcode 01274919), 12 May 1964, JX Yang 2656 (PE barcode 01274923), 22 May 1964, JX Yang 2733 (PE barcode 01274920), 10 June 1964, JX Yang 2929 (PE barcode 01274916), Banmiao, 6 May 1964, JX Yang 2558 (PE barcode 01274922). USA. 10 May 1918, HH Chung 4255 (PE barcode 01682167), 6 May 1930, CEK and FPM 17501 (PE barcode 01682166).
Pyrus ussuriensis Maxim. ex Rupr.: CHINA. Beijing: Changping District, Nankou, 29 May 1956, Herbarium Team 1198 (PE barcode 00549475), Fangshan District, 24 August 1956, Herbarium Team 3529 (PE barcode 00549474), Haidian District, Xishan, Biyunsi, 22 April 1955, Herbarium Team 561 (PE 00549442, 00549478), 10 July 1955, Wofosi Investigation Team 153 (PE barcode 01448240), 1 August 1957, YJ Zhang 327 (PE barcode 01448238), Western Park, 15 April 1951, FZ Wang 11 (PE barcode 01449781), Mentougou District, Baihuashan, 22 July 1956, CJ Liu and DY Xing 170 (PE barcode 00549483, 00549498), Dajuesi, April 1936, DF Jin 12008 (PE barcode 01449776, 01449777), Zhoujiaxiang, April 1936, Y Liu 12013 (PE barcode 01449775), 2 August 1936, DF Jin 181 (PE barcode 01449774), DF Jin 188 (PE 01449773), Miaofengshan, 23 May 1930, HF Chow 40274 (PE barcode 00549481), 6 May 1953, F Zhao 71 (PE barcode 00549506), Miyun County, Wulingshan, 4 May 1951, Y Liu and J Zhang 15067 (PE barcode 00549448, 00549451, 00549487), Pinggu District, Beiyang Bridge, 23 May 1972, Anonymous Ping-188 (PE barcode 01204098); Hebei Province: 1930, HF Chow 40336 (PE barcode 00549496), 21 April 1936, Anonymous 379 (PE barcode 00549493), Changli County, West Zhanggezhuang, 26 April 1956, P Zhang 651 (PE barcode 01449433), Dingzhou City, Xinxingzhuang, 15 April 1956, JY Zhong 529 (PE barcode 01449727), JY Zhong 530 (PE barcode 01449735), Zhaozhuang, 22 May 1956, JY Zhong 551 (PE barcode 01449737), Dongling, 23 April 1930, HF Chow 40325 (PE barcode 00549479), 1 May 1932, HF Chow 41931 (PE barcode 00549480), 22 April 1936, DF Jin K-900 (PE barcode 00549445), 20 June 1956, JX Duan 27 (PE barcode 00549441, 00549446), 25 June 1956, JX Duan 180 (PE barcode 00549450), Funing County, 30 April 1956, P Zhang 668 (PE barcode 01448400), Jiejiagou, 22 September 1956, FS Chang 778 (PE barcode 01448651), Neiqiu County, Fuanmugou, XY Liu and F Zhao 431 (PE barcode 01449778), Yu County, Xiaowutaishan, 13 September 1956, Herbarium Team 2417 (PE barcode 00549476), Zhangjiakou City, 25 April 1956, JX Duan 180 (PE barcode 00549447, 00549484, 00549485), Xiling, Huangtupo, 8 June 1953, F Zhao 222 (PE barcode 00549488); Tianjin: Ji County, Panshan, 4 July 1956, Herbarium Team 1941 (PE barcode 00549444); Precise locality unknown, Anonymous 12049 (PE barcode 01449783), Anonymous K-919 (PE barcode 01449782).
References
- Cowie, R.H.; Bouchet, P.; Fontaine, B. The Sixth Mass Extinction: Fact, fiction or speculation? Biol. Rev. 2022, 97, 640–663. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Chen, G.; Edward Grumbine, R.; Dao, Z.; Sun, W.; Guo, H. Conserving plant species with extremely small populations (PSESP) in China. Biodivers. Conserv. 2013, 22, 803–809. [Google Scholar] [CrossRef]
- Lu, Z.; Qin, H.; Jin, X.; Zhang, Z.; Yang, Q.; Hong, D.; Li, D.; Li, K.; Yuan, L.; Zhou, Z. On the necessity, principle, and process of updating the List of National Key Protected Wild Plants. Biodivers. Sci. 2021, 29, 1577–1582. [Google Scholar] [CrossRef]
- Yang, Y.; Tan, C.; Yang, Z. Conservation of gymnosperms in China: Perspectives from the List of National Key Protected Wild Plants. Biodivers. Sci. 2021, 29, 1591–1598. [Google Scholar] [CrossRef]
- Heywood, V.H.; Davis, P.H. Principles of Angiosperm Taxonomy; Oliver and Boyd: Edinburgh, UK; London, UK, 1963. [Google Scholar]
- Dubois, A. Phylogeny, taxonomy, and nomenclature: The problem of taxonomic categories and nomenclatural ranks. Zootaxa 2007, 1519, 27–58. [Google Scholar] [CrossRef]
- Morrison, W.R.; Lohr, J.L.; Duchen, P.; Wilches, R.; Trujillo, D.; Mair, M.; Renner, S.S. The impact of taxonomic change on conservation: Does it kill, can it save, or is it just irrelevant? Biol. Conserv. 2009, 142, 3201–3206. [Google Scholar] [CrossRef]
- Goodwin, Z.; Harris, D.J.; Filer, D.; Wood, J.R.I.; Scotland, R.W. Widespread mistaken identity in tropical plant collections. Curr. Biol. 2015, 25, R1066–R1067. [Google Scholar] [CrossRef] [Green Version]
- Lian, L.; Shen, J.Y.; Ortiz, R.D.C.; Yu, S.X.; Chen, Z.D.; Wang, W. Phylogenetic analyses and modelling distributions guide conservation of a critically endangered liana species, Eleutharrhena macrocarpa (Menispermaceae). Taxon 2021, 70, 931–945. [Google Scholar] [CrossRef]
- Zhang, M.; Xue, C.; Hu, H.; Li, J.; Xue, Y.; Wang, R.; Fan, J.; Zou, C.; Tao, S.; Qin, M.; et al. Genome-wide association studies provide insights into the genetic determination of fruit traits of pear. Nat. Commun. 2021, 12, 1144. [Google Scholar] [CrossRef]
- Rubtsov, G.A. Geographical distribution of the genus Pyrus and trends and factors in its evolution. Am. Nat. 1944, 78, 358–366. [Google Scholar] [CrossRef]
- Korotkova, N.; Parolly, G.; Khachatryan, A.; Ghulikyan, L.; Sargsyan, H.; Akopian, J.; Borsch, T.; Gruenstaeudl, M. Towards resolving the evolutionary history of Caucasian pears (Pyrus, Rosaceae)-Phylogenetic relationships, divergence times and leaf trait evolution. J. Syst. Evol. 2018, 56, 35–47. [Google Scholar] [CrossRef]
- Dong, X.; Wang, Z.; Tian, L.; Zhang, Y.; Qi, D.; Huo, H.; Xu, J.; Li, Z.; Liao, R.; Shi, M.; et al. De novo assembly of a wild pear (Pyrus betuleafolia) genome. Plant Biotechnol. J. 2020, 18, 581–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browicz, K. Concept and chorology of the genus Pyrus L. Arboret. Kórnick. 1993, 38, 17–33. [Google Scholar]
- Robertson, K.R.; Rohrer, J.R.; Rohrer, J.B.; Smith, P.G. A synopsis of genera in subfamily Maloideae (Rosaceae). Syst. Bot. 1991, 16, 376–394. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Z.; Shi, Z.; Zhang, S.; Ming, R.; Zhu, S.; Khan, M.A.; Tao, S.; Korban, S.S.; Wang, H.; et al. The genome of the pear (Pyrus bretschneideri Rehd.). Genome Res. 2013, 23, 396–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Singh, J.; Qin, M.; Li, S.; Zhang, X.; Zhang, M.; Khan, A.; Zhang, S.; Wu, J. Development of an integrated 200K SNP genotyping array and application for genetic mapping, genome assembly improvement and genome wide association studies in pear (Pyrus). Plant Biotechnol. J. 2019, 17, 1582–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirasawa, K.; Itai, A.; Isobe, S. Chromosome-scale genome assembly of Japanese pear (Pyrus pyrifolia) variety ‘Nijisseiki’. DNA Res. 2021, 28, b1. [Google Scholar] [CrossRef]
- Zheng, X.; Cai, D.; Potter, D.; Postman, J.; Liu, J.; Teng, Y. Phylogeny and evolutionary histories of Pyrus L. revealed by phylogenetic trees and networks based on data from multiple DNA sequences. Mol. Phylogenet. Evol. 2014, 80, 54–65. [Google Scholar] [CrossRef]
- Feng, S.; Bai, M.; Rivas-González, I.; Li, C.; Liu, S.; Tong, Y.; Yang, H.; Chen, G.; Xie, D.; Sears, K.E.; et al. Incomplete lineage sorting and phenotypic evolution in marsupials. Cell 2022, 185, 1646–1660.e18. [Google Scholar] [CrossRef]
- Mu, X.Y.; Sun, M.; Yang, P.F.; Lin, Q.W. Unveiling the identity of wenwan walnuts and phylogenetic relationships of Asian Juglans species using restriction site-associated DNA-sequencing. Front. Plant Sci. 2017, 8, 1708. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.B.; Liu, G.N.; Hong, D.Y.; Wen, J. Eriobotrya belongs to Rhaphiolepis (Maleae, Rosaceae): Evidence from chloroplast genome and nuclear ribosomal DNA data. Front. Plant Sci. 2020, 10, 1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yü, T.T.; Ku, T.C. Pyrus. In Flora Reipublicae Popularis Sinicae; Yü, T.T., Ed.; Science Press: Beijing, China, 1974; Volume 36, pp. 354–372. [Google Scholar]
- Wuyun, T.; Ma, T.; Uematsu, C.; Katayama, H. A phylogenetic network of wild Ussurian pears (Pyrus ussuriensis Maxim.) in China revealed by hypervariable regions of chloroplast DNA. Tree Genet. Genomes 2013, 9, 167–177. [Google Scholar] [CrossRef]
- Yü, T.T.; Kuan, K.C. Taxa nova Rosacearum Sinicarum (I). Acta Phytotaxon. Sin. 1963, 8, 202–236. [Google Scholar]
- Liang, T.T.; Ma, Y.; Guo, J.; Zang, D.K. Transcriptome sequencing and analysis of wild pear (Pyrus hopeiensis) using the Illumina Platform. Arab. J. Sci. Eng. 2016, 41, 45–53. [Google Scholar] [CrossRef]
- Zang, R.G.; Dong, M.; Li, J.Q.; Chen, X.Y.; Zeng, S.J.; Jiang, M.X.; Li, Z.Q.; Huang, J.H. Conservation and restoration for typical critically endangered wild plants with extremely small population. Acta Ecol. Sin. 2016, 36, 7130–7135. [Google Scholar] [CrossRef]
- Li, W.; Lu, Y.; Xie, X.; Li, B.; Han, Y.; Sun, T.; Xian, Y.; Yang, H.; Liu, K. Development of chloroplast genomic resources for Pyrus hopeiensis (Rosaceae). Conserv. Genet. Resour. 2018, 10, 511–513. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.; Li, L.; Gao, L.; Xu, J.; Yang, M. Structural and comparative analysis of the complete chloroplast genome of Pyrus hopeiensis “Wild Plants with a Tiny Population” and three other Pyrus species. Int. J. Mol. Sci. 2018, 19, 3262. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Tian, L.; Gao, Y.; Liu, F. Genetic diversity of cultivated and wild Ussurian Pear (Pyrus ussuriensis Maxim.) in China evaluated with M13-tailed SSR markers. Genet. Resour. Crop Evol. 2012, 59, 9–17. [Google Scholar] [CrossRef]
- Liu, Q.; Kang, M.; Jiang, Y. Some new recorded plants from Beijing and Hebei (II). J. Beijing Norm. Univ. 2003, 39, 674–676. [Google Scholar]
- Wang, Q. Identity of Microtoena coreana (Lamiaceae), a doubtful species from Korea. Phytotaxa 2015, 195, 79–85. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Y.; Xu, J.; Korban, S.S.; Fei, Z.; Tao, S.; Ming, R.; Tai, S.; Khan, A.M.; Postman, J.D.; et al. Diversification and independent domestication of Asian and European pears. Genome Biol. 2018, 19, 77. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.; Wang, S.; Zhang, Y.; Yang, M. The distribution and origins of Pyrus hopeiensis-“Wild Plant With Tiny Population” using whole genome resequencing. Front. Plant Sci. 2021, 12, 668796. [Google Scholar] [CrossRef] [PubMed]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus × domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; DePamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Qu, Y.; Wang, R.; Ma, C.; Li, D.; Song, J. Genetic diversity analysis of pear genus in Shanxi Province by SSR markers. North. Hort. 2015, 39, 11–17. [Google Scholar]
- Xiang, Y.; Huang, C.; Hu, Y.; Wen, J.; Li, S.; Yi, T.; Chen, H.; Xiang, J.; Ma, H. Evolution of Rosaceae fruit types based on nuclear phylogeny in the context of geological times and genome duplication. Mol. Biol. Evol. 2017, 34, 262–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.D.; Jin, J.J.; Chen, S.Y.; Chase, M.W.; Soltis, D.E.; Li, H.T.; Yang, J.B.; Li, D.Z.; Yi, T.S. Diversification of Rosaceae since the Late Cretaceous based on plastid phylogenomics. New Phytol. 2017, 214, 1355–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.B.; Ren, C.; Kwak, M.; Hodel, R.G.J.; Xu, C.; He, J.; Zhou, W.B.; Huang, C.H.; Ma, H.; Qian, G.Z.; et al. Phylogenomic conflict analyses in the apple genus Malus s.l. reveal widespread hybridization and allopolyploidy driving diversification, with insights into the complex biogeographic history in the Northern Hemisphere. J. Integr. Plant Biol. 2022, 64, 1020–1043. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Hu, C.; Spooner, D.; Liu, J.; Cao, J.; Teng, Y. Molecular evolution of Adh and LEAFY and the phylogenetic utility of their introns in Pyrus (Rosaceae). BMC Evol. Biol. 2011, 11, 255. [Google Scholar] [CrossRef] [Green Version]
- Corlett, R.T. Safeguarding our future by protecting biodiversity. Plant Divers. 2020, 42, 221–228. [Google Scholar] [CrossRef]
- Raven, P.; Wackernagel, M. Maintaining biodiversity will define our long-term success. Plant Divers. 2020, 42, 211–220. [Google Scholar] [CrossRef]
- Bremer, K.; Bremer, B.; Karis, P.; Kallersjo, M. Time for change in taxonomy. Nature 1990, 343, 202. [Google Scholar] [CrossRef]
- Raposo, M.A.; Stopiglia, R.; Brito, G.R.R.; Bockmann, F.A.; Kirwan, G.M.; Gayon, J.; Dubois, A. What really hampers taxonomy and conservation? A riposte to Garnett and Christidis (2017). Zootaxa 2017, 4317, 179–184. [Google Scholar] [CrossRef]
- Hu, H.H. Notulae systematicae ad Florem Sinensium V. Bull. Fan Mem. Inst. Biol. 1934, 5, 305–306. [Google Scholar]
- Zhang, W.P.; Cao, L.; Lin, X.R.; Ding, Y.M.; Liang, Y.; Zhang, D.Y.; Pang, E.L.; Renner, S.S.; Bai, W.N. Dead-end hybridization in walnut trees revealed by large-scale genomic sequence data. Mol. Biol. Evol. 2021, 39, msab308. [Google Scholar] [CrossRef]
- Krofel, M.; Hatlauf, J.; Bogdanowicz, W.; Campbell, L.A.D.; Godinho, R.; Jhala, Y.V.; Kitchener, A.C.; Koepfli, K.P.; Moehlman, P.; Senn, H.; et al. Towards resolving taxonomic uncertainties in wolf, dog and jackal lineages of Africa, Eurasia and Australasia. J. Zool. 2022, 316, 155–168. [Google Scholar] [CrossRef]
- Albani Rocchetti, G.; Armstrong, C.G.; Abeli, T.; Orsenigo, S.; Jasper, C.; Joly, S.; Bruneau, A.; Zytaruk, M.; Vamosi, J.C. Reversing extinction trends: New uses of (old) herbarium specimens to accelerate conservation action on threatened species. New Phytol. 2021, 230, 433–450. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-C., D.; Richardson, J.E.; Hart, M.; Serrano, J.; Cárdenas, D.; Gonzalez, M.A.; Cortés-B., R. A plea to DNA barcode type specimens: An example from Micropholis (Sapotaceae). Taxon 2022, 71, 154–167. [Google Scholar] [CrossRef]
- Hart, M.L.; Forrest, L.L.; Nicholls, J.A.; Kidner, C.A. Retrieval of hundreds of nuclear loci from herbarium specimens. Taxon 2016, 65, 1081–1092. [Google Scholar] [CrossRef]
- Silva, C.; Besnard, G.; Piot, A.; Razanatsoa, J.; Oliveira, R.P.; Vorontsova, M.S. Museomics resolve the systematics of an endangered grass lineage endemic to north-western Madagascar. Ann. Bot. 2017, 119, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Raxworthy, C.J.; Smith, B.T. Mining museums for historical DNA: Advances and challenges in museomics. Trends Ecol. Evol. 2021, 36, 1049–1060. [Google Scholar] [CrossRef]
Figure 1.
Principal coordinate analysis of Pyrus hopeiensis, P. ussuriensis and P. phaeocarpa based on morphological characters. (A) specimens of flowering period, (B) specimens of fruiting period. Black triangles: P. phaeocarpa, white triangles: P. ussuriensis, black circles: P. hopeiensis.
Figure 1.
Principal coordinate analysis of Pyrus hopeiensis, P. ussuriensis and P. phaeocarpa based on morphological characters. (A) specimens of flowering period, (B) specimens of fruiting period. Black triangles: P. phaeocarpa, white triangles: P. ussuriensis, black circles: P. hopeiensis.

Figure 2.
Maximum likelihood phylogenetic tree inferred from the SNP data matrix based on the whole-genome resequencing data (A) and whole plastid genome sequences (B). The numbers above the nodes indicate bootstrap values generated from maximum likelihood analysis, scale bar indicates substitutions per site. Whole plastome sequences of species downloaded from NCBI are given their GenBank numbers following species’ name in the plastome tree (B). Samples of P. hopeiensis are presented with red, P. ussuriensis with blue, and P. phaeocarpa with pink.
Figure 2.
Maximum likelihood phylogenetic tree inferred from the SNP data matrix based on the whole-genome resequencing data (A) and whole plastid genome sequences (B). The numbers above the nodes indicate bootstrap values generated from maximum likelihood analysis, scale bar indicates substitutions per site. Whole plastome sequences of species downloaded from NCBI are given their GenBank numbers following species’ name in the plastome tree (B). Samples of P. hopeiensis are presented with red, P. ussuriensis with blue, and P. phaeocarpa with pink.

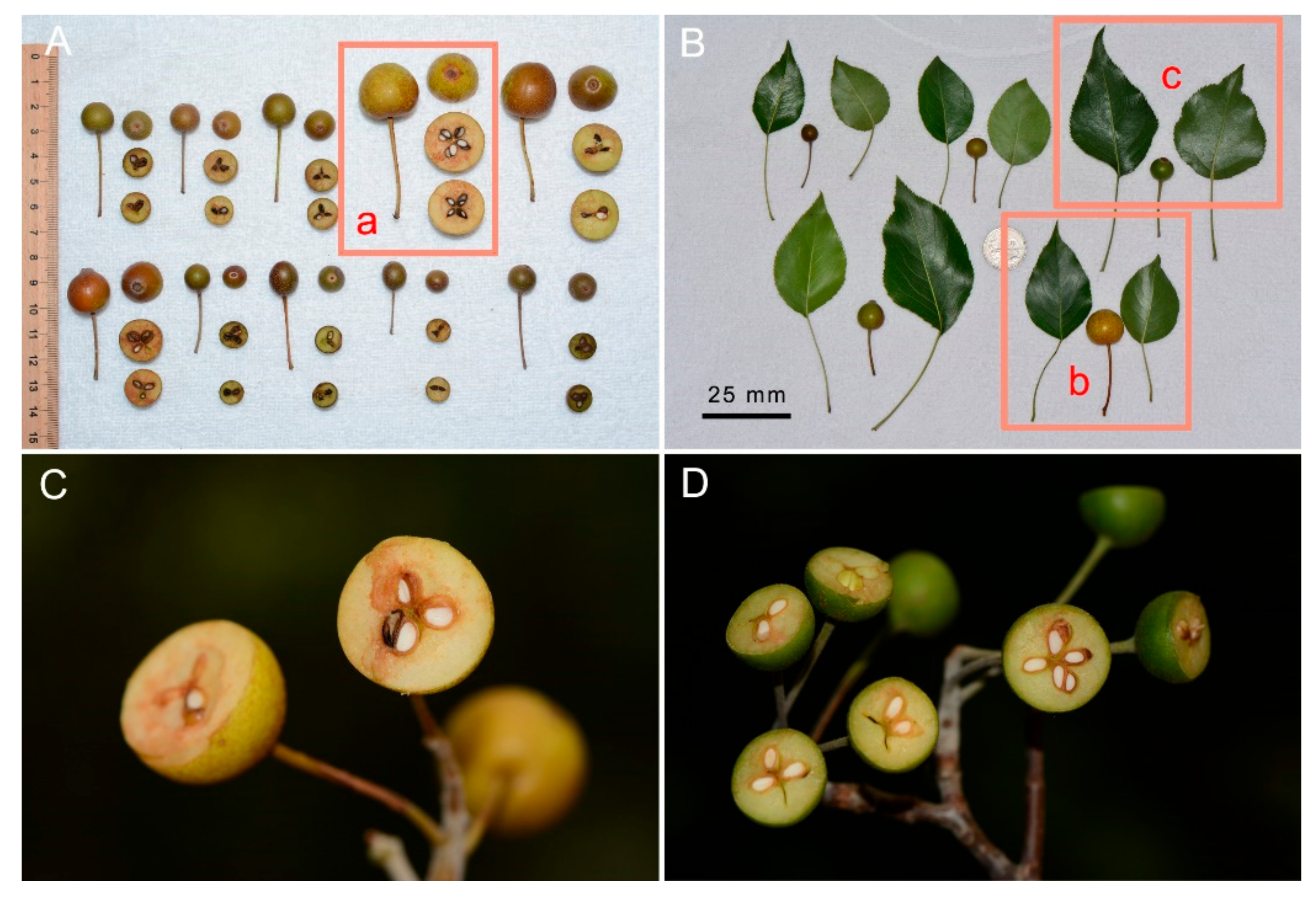
Figure 3.
Type specimens of Pyrus hopeiensis and P. ussuriensis and fruit morphological characteristics of P. ussuriensis populations in North China. (A) Holotype of P. hopeiensis, (B) lectotype of P. ussuriensis, (C–E) fruit color, spots, and calyx, (F,G) fruit size, length of pedicel and calyx, (H,I) carpel number (4 vs. 5).
Figure 3.
Type specimens of Pyrus hopeiensis and P. ussuriensis and fruit morphological characteristics of P. ussuriensis populations in North China. (A) Holotype of P. hopeiensis, (B) lectotype of P. ussuriensis, (C–E) fruit color, spots, and calyx, (F,G) fruit size, length of pedicel and calyx, (H,I) carpel number (4 vs. 5).

Figure 4.
Fruits and leaves of Pyrus phaeocarpa and P. betulifolia from the type locality of P. hopeiensis, the Jieshi Mountain, Changli County, Hebei Province. (A,B) Fruits and leaves of P. phaeocarpa and P. betulifolia, (C,D) cross-cut of fruits of P. phaeocarpa and P. betulifolia, respectively, indicating 3–4 carpels per fruit. (A(a),B(b),C) were collected from the same tree of P. phaeocarpa, (B(c),D) were collected from the same tree of P. betulifolia.
Figure 4.
Fruits and leaves of Pyrus phaeocarpa and P. betulifolia from the type locality of P. hopeiensis, the Jieshi Mountain, Changli County, Hebei Province. (A,B) Fruits and leaves of P. phaeocarpa and P. betulifolia, (C,D) cross-cut of fruits of P. phaeocarpa and P. betulifolia, respectively, indicating 3–4 carpels per fruit. (A(a),B(b),C) were collected from the same tree of P. phaeocarpa, (B(c),D) were collected from the same tree of P. betulifolia.

Figure 5.
Integrative evidence indicates that the locally endemic Pyrus hopeiensis is a synonym of P. ussuriensis and should be excluded from the list of key protected wild plants.
Figure 5.
Integrative evidence indicates that the locally endemic Pyrus hopeiensis is a synonym of P. ussuriensis and should be excluded from the list of key protected wild plants.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Morphological characteristics used in the principal coordinate analysis and their coding.
| Specimens of the Flowering Period | Specimens of the Fruiting Period | ||
|---|---|---|---|
| 1 | Leaf blade length [cm] | 1 | Leaf blade length [cm] |
| 2 | Leaf blade width [cm] | 2 | Leaf blade width [cm] |
| 3 | Ratio of leaf blade width to length | 3 | Ratio of leaf blade width to length |
| 4 | Leaf serration: (1) long; (0) short | 4 | Leaf serration: (1) long; (0) short |
| 5 | Length of flower stalk [cm] | 5 | Length of fruit [cm] |
| 6 | Indumentum density on stalk: (0) sparse; (1) dense | 6 | Persisting calyx: (0) no; (1) yes |
| 7 | Length of pedicel [cm] | ||
| 8 | Ratio of fruit to pedicel Length | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mu, X.-Y.; Wu, J.; Wu, J. Taxonomic Uncertainty and Its Conservation Implications in Management, a Case from Pyrus hopeiensis (Rosaceae). Diversity 2022, 14, 417. https://doi.org/10.3390/d14060417
AMA Style
Mu X-Y, Wu J, Wu J. Taxonomic Uncertainty and Its Conservation Implications in Management, a Case from Pyrus hopeiensis (Rosaceae). Diversity. 2022; 14(6):417. https://doi.org/10.3390/d14060417
Chicago/Turabian StyleMu, Xian-Yun, Jiang Wu, and Jun Wu. 2022. "Taxonomic Uncertainty and Its Conservation Implications in Management, a Case from Pyrus hopeiensis (Rosaceae)" Diversity 14, no. 6: 417. https://doi.org/10.3390/d14060417
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.