
Latitudinal Diversity Gradient in the Changing World: Retrospectives and Perspectives


 and
and
Abstract
:1. Introduction
2. Status of LDG from Previous Studies
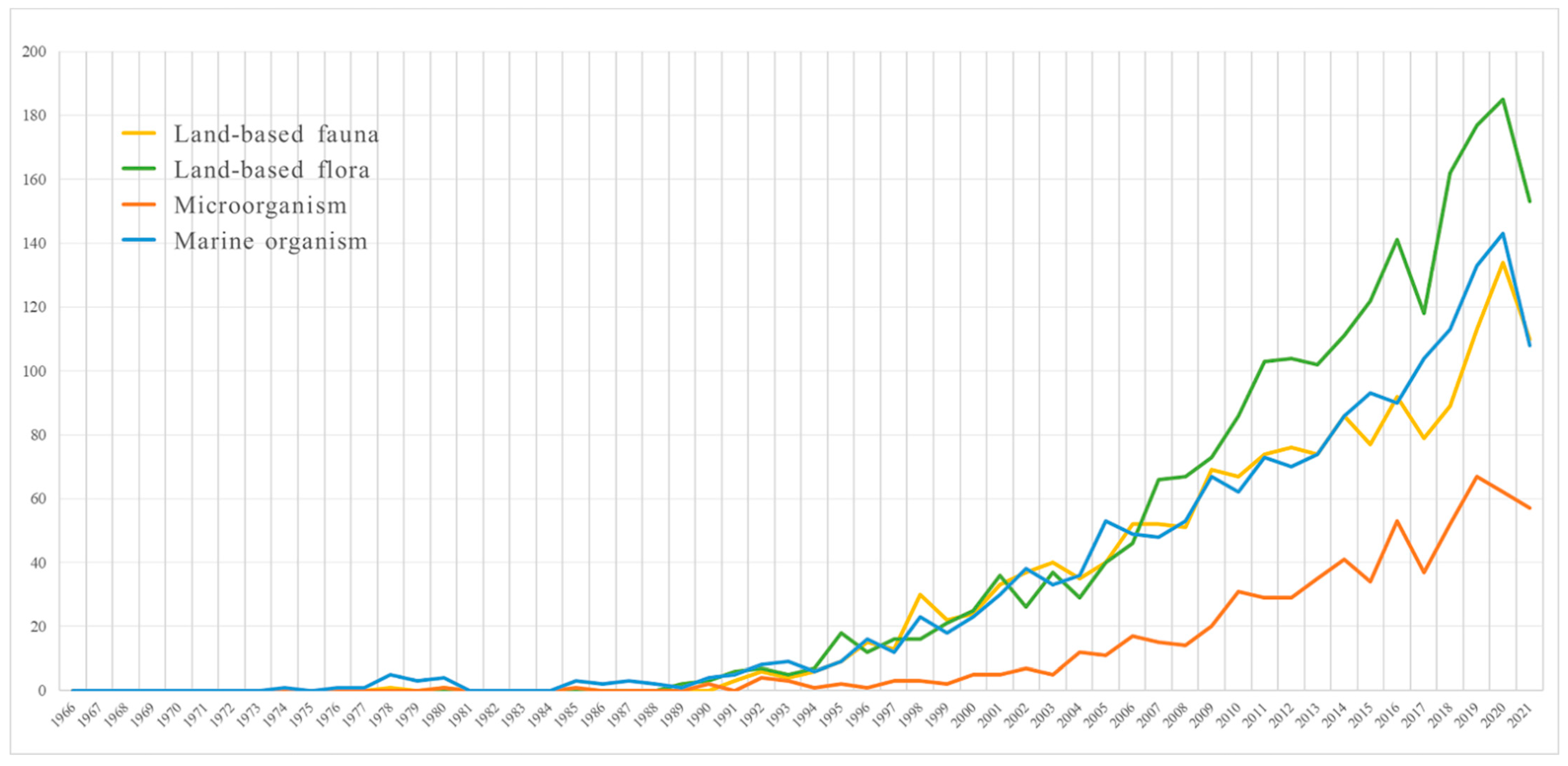
2.1. Land-Based Flora
2.2. Land-Based Fauna
2.3. Microorganisms
2.4. Marine Organisms
2.5. LDG and Biodiversity Conservation
3. Formation Mechanisms of the LDG
3.1. LDG Hypotheses
3.2. Climate Change, Temperature, and Precipitation
4. Evolutionary Responses for LDG
4.1. Speciation Rate
4.2. Extinction Rate
4.3. Net Diversification Rate
4.4. Dispersal Rate
5. Future Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- von Humboldt, A. Views of Nature: Or Contemplations on the Sublime Phenomena of Creation with Scientific Illustrations; (Transl. Otté, E.C., Bohn, H.G., 1st edition in 1850); Higher Education Press: Beijing, China, 2016. [Google Scholar]
- Hawkins, B.A. Ecology’s oldest pattern? Trends Ecol. Evol. 2001, 16, 470. [Google Scholar] [CrossRef]
- Hillebrand, H. On the generality of the latitudinal diversity gradient. Am. Nat. 2004, 163, 192–211. [Google Scholar] [CrossRef] [Green Version]
- Kinlock, N.L.; Prowant, L.; Herstoff, E.M.; Foley, C.M.; Akin-Fajiye, M.; Bender, N.; Umarani, M.; Ryu, H.Y.; Sen, B.; Gurevitch, J. Explaining global variation in the latitudinal diversity gradient: Meta-analysis confirms known patterns and uncovers new ones. Glob. Ecol. Biogeogr. 2017, 27, 125–141. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Schemske, D.W.; Cornell, H.V.; Allen, A.P.; Brown, J.M.; Bush, M.B.; Harrison, S.P.; Hurlbert, A.H.; Knowlton, N.; Lessios, H.; et al. Evolution and the latitudinal diversity gradient: Speciation, extinction and biogeography. Ecol. Lett. 2007, 10, 315–331. [Google Scholar] [CrossRef]
- Pianka, E.R. Latitudinal Gradients in Species Diversity: A Review of Concepts. Am. Nat. 1966, 100, 33–46. [Google Scholar] [CrossRef]
- Pulido–Santacruz, P.; Weir, J.T. Extinction as a driver of avian latitudinal diversity gradients. Evolution 2016, 70, 860–872. [Google Scholar] [CrossRef]
- Rohde, K. Latitudinal gradients in species diversity: The search for the primary cause. Oikos 1992, 65, 514–527. [Google Scholar] [CrossRef] [Green Version]
- Saupe, E.E.; Myers, C.E.; Townsend Peterson, A.; Soberon, J.; Singarayer, J.; Valdes, P.; Qiao, H. Spatio–temporal climate change contributes to latitudinal diversity gradients. Nat. Ecol. Evol. 2019, 3, 1419–1429. [Google Scholar] [CrossRef] [Green Version]
- Schemske, D.W.; Mittelbach, G.G. “Latitudinal gradients in species diversity”: Reflections on Pianka’s 1966 article and a look forward. Am. Nat. 2017, 189, 599–603. [Google Scholar] [CrossRef]
- Stevens, G.C. The latitudinal gradient in geographical range: How so many species coexist in the tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Tomašových, A.; Jablonski, D.; Sandel, B. Decoupling of latitudinal gradients in species and genus geographic range size: A signature of clade range expansion. Glob. Ecol. Biogeogr. 2016, 26, 288–303. [Google Scholar] [CrossRef]
- Jablonski, D.; Belanger, C.L.; Berke, S.K.; Huang, S.; Krug, A.Z.; Roy, K.; Tomašových, A.; Valentine, J.W. Out of the tropics, but how? Fossils, bridge species, and thermal ranges in the dynamics of the marine latitudinal diversity gradient. Proc. Natl. Acad. Sci. USA 2013, 110, 10487–10494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jablonski, D.; Roy, K.; Valentine, J.W. Out of the tropics: Evolutionary dynamics of the latitudinal diversity gradien. Science 2006, 314, 102–106. [Google Scholar] [CrossRef] [Green Version]
- Fine, P.V.A. Ecological and evolutionary drivers of geographic variation in species diversity. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 369–392. [Google Scholar] [CrossRef] [Green Version]
- Fuhrman, J.A.; Steele, J.A.; Hewson, I.; Schwalbach, M.S.; Brown, M.V.; Green, J.L.; Brown, J.H. A latitudinal diversity gradient in planktonic marine bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 7774–7778. [Google Scholar] [CrossRef] [Green Version]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal gradients of biodiversity: Pattern, process, scale, and synthesis. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Jablonski, D.; Huang, S.; Roy, K.; Valentine, J.W. Shaping the latitudinal diversity gradient: New perspectives from a synthesis of paleobiology and biogeography. Am. Nat. 2017, 189, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lomolino, M.V.; Riddle, B.R.; Whittaker, R.J.; Brown, J.H. Biogeography, 4th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2010. [Google Scholar]
- Hanly, P.J.; Mittelbach, G.G.; Schemske, D.W. Speciation and the latitudinal diversity gradient: Insights from the global distribution of endemic fish. Am. Nat. 2017, 189, 604–615. [Google Scholar] [CrossRef] [Green Version]
- Jansson, R.; Rodriguez–Castaneda, G.; Harding, L.E. What can multiple phylogenies say about the latitudinal diversity gradient? A new look at the tropical conservatism, out of the tropics, and diversification rate hypotheses. Evolution 2013, 67, 1741–1755. [Google Scholar] [CrossRef]
- Chen, S.B.; Ferry Slik, J.W.; Gao, J.; Mao, L.F.; Bi, M.J.; Shen, M.W.; Zhou, K.X. Latitudinal diversity gradients in bryophytes and woody plants: Roles of temperature and water availability. J. Syst. Evol. 2015, 53, 535–545. [Google Scholar] [CrossRef]
- Theodoridis, S.; Fordham, D.A.; Brown, S.C.; Li, S.; Rahbek, C.; Nogues-Bravo, D. Evolutionary history and past climate change shape the distribution of genetic diversity in terrestrial mammals. Nat. Commun. 2020, 11, 2557. [Google Scholar] [CrossRef] [PubMed]
- Chown, S.L.; Gaston, K.J. Areas, cradles and museums: The latitudinal gradient in species richness. Trends Ecol. Evol. 2000, 15, 311–315. [Google Scholar] [CrossRef]
- Huang, J.; Ma, K.; Huang, J. Phytogeographical patterns of genera of endemic flora in relation to latitudinal and climatic gradients in China. Plant Syst. Evol. 2017, 303, 689–698. [Google Scholar] [CrossRef]
- Qian, H.; Song, J.S.; Krestov, P.; Guo, Q.; Wu, Z.; Shen, X.; Guo, X. Large–scale phytogeographical patterns in East Asia in relation to latitudinal and climatic gradients. J. Biogeogr. 2003, 30, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.; Brauer, C.J.; Sasaki, M.; Unmack, P.J.; Guillot, G.; Laporte, M.; Bernatchez, L.; Beheregaray, L.B. Latitudinal variation in climate–associated genes imperils range edge populations. Mol. Ecol. 2020, 29, 4337–4349. [Google Scholar] [CrossRef] [PubMed]
- von Konrat, M.; Renner, M.; Soederstroem, L.; Hagborg, A.; Mutke, J. Chapter nine: Early land plants today: Liverwort species diversity and the relationship with higher taxonomy and higher plants. Fieldiana. Bot. 2008, 47, 91–104. [Google Scholar] [CrossRef]
- Liu, H.; Yu, R.; Huang, J.; Liu, Y.; Zang, R.; Guo, Z.; Ding, Y.; Lu, X.; Li, Q.; Chen, H.Y.H. Latitudinal diversity gradients and Rapoport effects in Chineseendemic woody seed plants. Forests 2020, 11, 1029. [Google Scholar] [CrossRef]
- Niskanen, A.K.J.; Niittynen, P.; Aalto, J.; Väre, H.; Luoto, M.; Diez, J. Lost at high latitudes: Arctic and endemic plants under threat as climate warms. Divers. Distrib. 2019, 25, 809–821. [Google Scholar] [CrossRef] [Green Version]
- Pyron, R.A. Temperate extinction in squamate reptiles and the roots of latitudinal diversity gradients. Glob. Ecol. Biogeogr. 2014, 23, 1126–1134. [Google Scholar] [CrossRef]
- Economo, E.P.; Narula, N.; Friedman, N.R.; Weiser, M.D.; Guenard, B. Macroecology and macroevolution of the latitudinal diversity gradient in ants. Nat. Commun. 2018, 9, 1778. [Google Scholar] [CrossRef] [Green Version]
- Saupe, E.E.; Farnsworth, A.; Lunt, D.J.; Sagoo, N.; Pham, K.V.; Field, D.J. Climatic shifts drove major contractions in avian latitudinal distributions throughout the Cenozoic. Proc. Natl. Acad. Sci. USA 2019, 116, 12895–12900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolland, J.; Condamine, F.L.; Jiguet, F.; Morlon, H. Faster speciation and reduced extinction in the tropics contribute to the mammalian latitudinal diversity gradient. PLoS Biol. 2014, 12, e1001775. [Google Scholar] [CrossRef] [PubMed]
- Hillebrand, H.; Azovsky, A.I. Body size determines the strength of the latitudinal diversity gradient. Ecography 2001, 24, 251–256. [Google Scholar] [CrossRef]
- Wilhelm, F. Biogeography and dispersal of micro-organisms: A review emphasizing protists. Acta Protozool. 2006, 45, 111–136. [Google Scholar]
- Andam, C.P.; Doroghazi, J.R.; Campbell, A.N.; Kelly, P.J.; Choudoir, M.J.; Buckley, D.H. A latitudinal diversity gradient in Terrestrial Bacteria of the genus Streptomyces. mBio 2016, 7, e02200-15. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.T.J.; Haas, S.E. Why do parasites exhibit reverse latitudinal diversity gradients? Testing the roles of host diversity, habitat and climate. Glob. Ecol. Biogeogr. 2021, 30, 1810–1821. [Google Scholar] [CrossRef]
- McDonald, C.P.; Rover, J.A.; Stets, E.G.; Striegl, R.G. The regional abundance and size distribution of lakes and reservoirs in the United States and implications for estimates of global lake extent. Limnol. Oceanogr. 2012, 57, 597–606. [Google Scholar] [CrossRef]
- Roy, K.; Jablonski, D.; Valentine, J.W. Dissecting latitudinal diversity gradients: Functional groups and clades of marine bivalves. Proc. R. Soc. Lond. B-Biol. Sci. 2000, 267, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Lambshead, P.J.D.; Tietjen, J.; Timothy, F.; Jensen, P. Latitudinal diversity gradients in the deep sea with special reference to North Atlantic nematodes. Mar. Ecol. Prog. Ser. 2000, 194, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Roy, K.; Jablonski, D.; Valentine, J.W.; Rosenberg, G. Marine latitudinal diversity gradients: Tests of causal hypotheses. Proc. Natl. Acad. Sci. USA 1998, 95, 3699–3702. [Google Scholar] [CrossRef] [Green Version]
- He, Z.W.; Li, X.N.; Yang, M.; Wang, X.F.; Zhong, C.R.; Duke, N.C.; Wu, C.I.; Shi, S.H. Speciation with gene flow via cycles of isolation and migration: Insights from multiple mangrove taxa. Natl. Sci. Rev. 2019, 6, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.H.; Zhou, S.S.; Li, L.; Tan, Y.H.; Li, J.W.; Li, J. Conflict between biodiversity conservation and economic growth: Insight into rare plants in tropical China. Biodivers. Conserv. 2019, 28, 523–537. [Google Scholar] [CrossRef]
- Johnson, C.N.; Balmford, A.; Brook, B.W.; Buettel, J.C.; Galetti, M.; Guangchun, L.; Wilmshurst, J.M. Biodiversity losses and conservation responses in the Anthropocene. Science 2017, 356, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Isbel, F.; Gonzalez, A.; Loreau, M.; Cowles, J.; Diaz, S.; Hector, A.; Mace, M.M.; Wardle, D.A.; O’Connor, M.I.; Duffy, J.M.; et al. Linking the influence and dependence of people on biodiversity across scales. Nature 2017, 546, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Meng, H.H.; Zhou, S.S.; Jiang, X.L.; Gugger, P.F.; Li, L.; Tan, Y.H.; Li, J. Are mountaintops climate refugia for plants under global warming? A lesson from high–mountain oaks in tropical rainforest. Alp. Bot. 2019, 129, 175–183. [Google Scholar] [CrossRef]
- Meng, H.H.; Gao, X.Y.; Song, Y.G.; Cao, G.L.; Li, J. Biodiversity arks in the Anthropocene. Reg. Sustain. 2021, 2, 109–115. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef]
- Li, H.P.; XiangYu, J.G.; Dai, G.Y.; Gu, Z.L.; Ming, C.; Yang, Z.F.; Ryder, O.A.; Li, W.H.; Fu, Y.X.; Zhang, Y.P. Large numbers of vertebrates began rapid population decline in the late 19th century. Proc. Natl. Acad. Sci. USA 2016, 113, 14079–14084. [Google Scholar] [CrossRef] [Green Version]
- Tucker, C.M.; Aze, T.; Cadotte, M.W.; Cantalapiedra, J.L.; Chisholm, C.; Diaz, S.; Grenyer, R.; Huang, D.W.; Mazel, F.; Pearse, W.D.; et al. Assessing the utility of conserving evolutionary history. Biol. Rev. 2019, 94, 1740–1760. [Google Scholar] [CrossRef] [Green Version]
- Barlow, J.; Franca, F.; Gardner, T.A.; Hicks, C.C.; Lennox, G.D.; Berenguer, E.; Castello, L.; Economo, E.P.; Ferreira, J.; Guenard, B.; et al. The future of hyperdiverse tropical ecosystems. Nature 2018, 559, 517–526. [Google Scholar] [CrossRef]
- Bonebrake, T.C.; Brown, C.J.; Bell, J.D.; Blanchard, J.L.; Chauvenet, A.; Champion, C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; et al. Managing consequences of climate–driven species redistribution requires integration of ecology, conservation and social science. Biol. Rev. 2018, 93, 284–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, T.J.; Barraclough, T.G.; Savolainen, V.; Chase, M.W. Environmental causes for plant biodiversity gradients. Philos. Trans. R. Soc. Lond. B-Biol. Sci. 2004, 359, 1645–1656. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, J.; Svenning, J.C. Climate–related range shifts a global multidimensional synthesis and new research directions. Ecography 2015, 38, 15–28. [Google Scholar] [CrossRef]
- Margules, C.R.; Pressey, R.L. Systematic conservation planning. Nature 2000, 405, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Scholl, J.P.; Wiens, J.J. Diversification rates and species richness across the Tree of Life. Proc. R. Soc. Lond. B-Biol. Sci. 2016, 283. [Google Scholar] [CrossRef]
- Chiu, W.T.R.; Yasuhara, M.; Cronin, T.M.; Hunt, G.; Gemery, L.; Wei, C.L. Marine latitudinal diversity gradients, niche conservatism and out of the tropics and Arctic: Climatic sensitivity of small organisms. J. Biogeogr. 2020, 47, 817–828. [Google Scholar] [CrossRef]
- Raja, N.B.; Kiessling, W. Out of the extratropics: The evolution of the latitudinal diversity gradient of Cenozoic marine plankton. Proc. R. Soc. B-Biol. Sci. 2021, 288, 20210545. [Google Scholar] [CrossRef]
- Laenen, B.; Patiño, J.; Hagborg, A.; Désamoré, A.; Wang, J.; Shaw, A.J.; Goffinet, B.; Vanderpoorten, A. Evolutionary origin of the latitudinal diversity gradient in liverworts. Mol. Phylogenet. Evol. 2018, 127, 606–612. [Google Scholar] [CrossRef]
- Qian, H.; Jin, Y.; Ricklefs, R.E. Patterns of phylogenetic relatedness of angiosperm woody plants across biomes and life-history stages. J. Biogeogr. 2017, 44, 1383–1392. [Google Scholar] [CrossRef]
- Wiens, J.J.; Donoghue, M.J. Historical biogeography, ecology and species richness. Trends Ecol. Evol. 2004, 19, 639–644. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The Importance of Keystone Structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.; Eastwood, R. Island radiation on a continental scale: Exceptional rates of plant diversification after uplift of the Andes. Proc. Natl. Acad. Sci. USA 2006, 103, 10334–10339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallimanis, A.S.; Bergmeier, E.; Panitsa, M.; Georghiou, K.; Delipetrou, P.; Dimopoulos, P. Biogeographical determinants for total and endemic species richness in a continental archipelago. Biol. Conserv. 2009, 19, 1225–1235. [Google Scholar] [CrossRef] [Green Version]
- Harms, K.E.; Wright, S.J.; Calderón, O.; Hernández, A.; Herre, E.A. Pervasive density–dependent recruitment enhances seedling diversity in a tropical forest. Nature 2000, 404, 493–495. [Google Scholar] [CrossRef]
- Janzen, D.H. Herbivores and the Number of Tree Species in Tropical Forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Philos. Trans. R. Soc. Lond. B-Biol. Sci. 2004, 359, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Dynesius, M.; Jansson, R. Evolutionary consequences of changes in species’ geographical distributions driven by Milankovitch climate oscillations. Proc. Natl. Acad. Sci. USA 2000, 97, 9115–9120. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.G. Latitudinal variations in organic diversity. Evolution 1960, 14, 64–81. [Google Scholar] [CrossRef]
- Gao, X.Y.; Meng, H.H.; Zhang, M.L. Diversification and vicariance of desert plants: Evidence inferred from chloroplast DNA sequence variation of Lagochilus ilicifolius (Lamiaceae). Biochem. Syst. Ecol. 2014, 55, 93–100. [Google Scholar] [CrossRef]
- Meng, H.H.; Gao, X.Y.; Huang, J.F.; Zhang, M.L. Plant phylogeography in arid Northwest China: Retrospectives and perspectives. J. Syst. Evol. 2015, 53, 33–46. [Google Scholar] [CrossRef]
- Brown, J.H.; Stevens, G.C.; Kaufman, D.M. The geographic range: Size, shape, boundaries, and internal structure. Annu. Rev. Ecol. Evol. Syst. 1996, 27, 597–623. [Google Scholar] [CrossRef] [Green Version]
- Rohde, K. Robust Phylogenies and adaptive radiations: A critical examination of methods used to identify key Innovations. Am. Nat. 1996, 148, 481–500. [Google Scholar] [CrossRef]
- Rohde, K. Latitudinal gradients in species diversity and Rapoport’s rule revisited: A review of recent work and what can parasites teach us about the causes of the gradients? Ecography 1999, 22, 593–613. [Google Scholar] [CrossRef]
- Wright, D.H. Speciesenergy theory: An extension of speciesarea theory. Oikos 1983, 41, 496–506. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broadscale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef] [Green Version]
- Rabosky, D.L. Ecological limits and diversification rate: Alternative paradigms to explain the variation in species richness among clades and regions. Ecol. Lett. 2009, 12, 735–743. [Google Scholar] [CrossRef]
- Rabosky, D.L.; Hurlbert, A.H. Species richness at continental scales is dominated by ecological limits. Am. Nat. 2015, 185, 572–583. [Google Scholar] [CrossRef]
- Theodoridis, S.; Randin, C.; Szovenyi, P.; Boucher, F.C.; Patsiou, T.S.; Conti, E. How do coldadapted plants respond to climatic cycles? Interglacial expansion explains current distribution and genomic diversity in Primula farinosa L. Syst. Biol. 2017, 66, 715–736. [Google Scholar] [CrossRef]
- Tiffney, B.H. erspectives on the origin of the floristic similarity between Eastern Asia and Eastern North America. J. Arn. Arbor. 1985, 66, 73–94. [Google Scholar]
- Meng, H.H.; Su, T.; Gao, X.Y.; Li, J.; Jiang, X.L.; Sun, H.; Zhou, Z.K. Warm-cold colonization: Response of oaks to uplift of the Himalaya-Hengduan Mountains. Mol. Ecol. 2017, 26, 3276–3294. [Google Scholar] [CrossRef] [PubMed]
- Siepielski, A.M.; Morrissey, M.B.; Buoro, M.; Carlson, S.M.; Caruso, C.M.; Clegg, S.M.; Coulson, T.; DiBattista, J.; Gotanda, K.M.; Francis, C.D.; et al. Precipitation drives global variation in natural selection. Science 2017, 355, 959–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldvogel, A.M.; Feldmeyer, B.; Rolshausen, G.; Exposito–Alonso, M.; Rellstab, C.; Kofler, R.; Mock, T.; Schmid, K.; Schmitt, I.; Bataillon, T.; et al. Evolutionary genomics can improve prediction of species’ responses to climate change. Evol. Lett. 2020, 4, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Haffer, J. Speciation in Amazonian Forest Birds. Science 1969, 165, 131–137. [Google Scholar] [CrossRef]
- Jon, F.; Jon, C.L. Geographical patterns of old and young species in African forest biota: The significance of specific montane areas as evolutionary centres. Biol. Conserv. 1997, 6, 325–346. [Google Scholar]
- Parker, L.D.; Hawkins, M.T.R.; Camacho–Sanchez, M.; Campana, M.G.; West-Roberts, J.A.; Wilbert, T.R.; Lim, H.C.; Rockwood, L.L.; Leonard, J.A.; Maldonado, J.E. Little genetic structure in a Bornean endemic small mammal across a steep ecological gradient. Mol. Ecol. 2020, 29, 4074–4090. [Google Scholar] [CrossRef]
- Oliver, M.J.; Tuba, Z.; Mishler, B.D. The Evolution of Vegetative Desiccation Tolerance in Land Plants. Plant Ecol. 2000, 151, 85–100. [Google Scholar] [CrossRef]
- Scheffers, B.R.; De Meester, L.; Bridge, T.C.; Hoffmann, A.A.; Pandolfi, J.M.; Corlett, R.T.; Butchart, S.H.; Pearce-Kelly, P.; Kovacs, K.M.; Dudgeon, D.; et al. The broad footprint of climate change from genes to biomes to people. Science 2016, 354, aaf7671. [Google Scholar] [CrossRef]
- Wiens, J.J.; Ackerly, D.D.; Allen, A.P.; Anacker, B.L.; Buckley, L.B.; Cornell, H.V.; Damschen, E.I.; Jonathan Davies, T.; Grytnes, J.A.; Harrison, S.P.; et al. Niche conservatism as an emerging principle in ecology and conservation biology. Ecol. Lett. 2010, 13, 1310–1324. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T. Extinction, ecological opportunity, and the origins of global snake diversity. Evolution 2012, 66, 163–178. [Google Scholar] [CrossRef]
- Sun, M.; Folk, R.A.; Gitzendanner, M.A.; Soltis, P.S.; Chen, Z.D.; Soltis, D.E.; Guralnick, R.P. Recent accelerated diversification in rosids occurred outside the tropics. Nat. Commun. 2020, 11, 3333. [Google Scholar] [PubMed]
- Gillman, L.N.; Wright, S.D. Species richness and evolutionary speed: The influence of temperature, water and area. J. Biogeogr. 2014, 41, 39–51. [Google Scholar]
- Claramunt, S.; Cracraft, J. A new time tree reveals Earth history’s imprint on the evolution of modern birds. Sci. Adv. 2015, 1, e1501005. [Google Scholar] [PubMed] [Green Version]
- Gaston, K.J. Species–range–size distributions: Patterns, mechanisms and implications. Trends Ecol. Evol. 1996, 11, 197–201. [Google Scholar] [CrossRef]
- Pyron, R.A.; Wiens, J.J. Large–scale phylogenetic analyses reveal the causes of high tropical amphibian diversity. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131622. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.P.; Brown, J.H.; Gillooly, J.F. Global biodiversity, biochemical kinetics, and the energetic–equivalence rule. Science 2002, 297, 1545–1548. [Google Scholar] [CrossRef]
- Cardillo, M. Latitude and rates of diversification in birds and butterflies. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1999, 266, 1221–1225. [Google Scholar]
- Meng, H.H.; Zhang, C.Y.; Song, Y.G.; Yu, X.Q.; Cao, G.L.; Li, L.; Cai, C.N.; Xiao, J.H.; Zhou, S.S.; Tan, Y.H.; et al. Opening a door to the spatiotemporal history of plants from the tropical Indochina Peninsula to subtropical China. Mol. Phylogenet. Evol. 2022, 171, 107458. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araujo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengard, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar]
- Braje, T.J.; Erlandson, J.M. Human acceleration of animal and plant extinctions: A Late Pleistocene, Holocene, and Anthropocene continuum. Anthropocene 2013, 4, 14–23. [Google Scholar]
- Newbold, T.; Hudson, L.N.; Hill, S.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Borger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locke, H.; Ellis, E.C.; Venter, O.; Schuster, R.; Ma, K.; Shen, X.; Woodley, S.; Kingston, N.; Bhola, N.; Strassburg, B.B.N.; et al. Three global conditions for biodiversity conservation and sustainable use: An implementation framework. Natl. Sci. Rev. 2019, 6, 1080–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.Y.; Low, S.L.; Song, Y.G.; Nurainas Kozlowski, G.; Do, T.V.; Li, L.; Zhou, S.S.; Tan, Y.H.; Cao, G.L.; Zhou, Z.; et al. Shining a light on species delimitation in the tree genus Engelhardia Leschenault ex Blume (Juglandaceae). Mol. Phylogenet. Evol. 2020, 152, 106918. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Hypothesis | Primary Focus | References |
|---|---|---|
| P1. The time theory | Ecology and evolution | [6] |
| P2. The theory of spatial heterogeneity | Ecology | [6] |
| P3. The competition hypothesis | Ecology | [6] |
| P4. The predation hypothesis | Ecology | [6] |
| P5. The theory of climatic stability | Ecology and evolution | [6] |
| P6. The productivity hypothesis | Ecology | [6] |
| F1. Time-integrated area, energy, and tropical niche conservatism | Evolution | [15] |
| F2. Climate stability | Evolution | [15] |
| F3. Temperature and evolutionary speed | Evolution | [15] |
| F4. Biotic interactions and speciation rate | Evolution | [15] |
| F5. Biotic interactions and finer niches | Ecology | [15] |
| O1.The ecological regulation hypothesis | Ecology | [32] |
| O2.The “diversification rate hypothesis | Evolution | [57] |
| O3.The out of the tropics hypothesis | Ecology and evolution | [58] |
| O4.The out-of-the-extratropics hypothesis | Ecology and evolution | [59] |
| O5.The evolutionary time hypothesis | Evolution | [32] |
| O6.The time-for-speciation hypothesis | Evolution | [60] |
| O7. The tropical niche conservatism hypothesis | Ecology | [61] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Song, Y.-G.; Zhang, C.-Y.; Wang, T.-R.; Su, T.-H.; Huang, P.-H.; Meng, H.-H.; Li, J. Latitudinal Diversity Gradient in the Changing World: Retrospectives and Perspectives. Diversity 2022, 14, 334. https://doi.org/10.3390/d14050334
Zhang Y, Song Y-G, Zhang C-Y, Wang T-R, Su T-H, Huang P-H, Meng H-H, Li J. Latitudinal Diversity Gradient in the Changing World: Retrospectives and Perspectives. Diversity. 2022; 14(5):334. https://doi.org/10.3390/d14050334
Chicago/Turabian StyleZhang, Yu, Yi-Gang Song, Can-Yu Zhang, Tian-Rui Wang, Tian-Hao Su, Pei-Han Huang, Hong-Hu Meng, and Jie Li. 2022. "Latitudinal Diversity Gradient in the Changing World: Retrospectives and Perspectives" Diversity 14, no. 5: 334. https://doi.org/10.3390/d14050334