
Holocene Genetic Evolution of Pig (Sus scrofa) on Romanian Territory in a European Time and Space Frame


1
Faculty of Biology, Alexandru Ioan Cuza University of Iasi, 700505 Iasi, Romania
2
“Vasile Pârvan” Institute of Archaeology, Romanian Academy, 010667 Bucharest, Romania
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(4), 288; https://doi.org/10.3390/d14040288
Submission received: 24 February 2022
/
Revised: 7 April 2022
/
Accepted: 9 April 2022
/
Published: 11 April 2022
(This article belongs to the Special Issue Revealing the Puzzle of the Past through Ancient Biomolecules: From Wild to Tame Faunal Diversity)
Abstract
:Romanian territory represents a key point in the dispersal of domestic pigs into Europe, due to its geographical position. Our study gathers a high number of samples from different archaeological sites on Romanian territory in order to establish a more accurate chronological view of the spread of domestic pig into Europe and to investigate the possibility of a local domestication process. Approximately 200 samples from 45 archaeological sites on Romanian territory, covering a large period of time, from the Neolithic to the Middle Ages, were subjected to DNA analysis. The sequencing of a short fragment from the D-loop region of the mitochondrial DNA identified a different prevalence of domestic pig genetic signature between two periods of time: the Neolithic period and the Early Bronze Age–Middle Ages period. While the Neolithic period is characterized by the presence of domestic pigs with a Near-Eastern signature, during the Early Bronze Age–Middle Ages period this genetic signature is replaced with a European one. Two European and two Near-Eastern signatures were described for all the analysed samples, each of them prevailing within the wild, respectively domestic Sus scrofa. The data also revealed the introgression process as a form of domestication in Romanian territory.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
The Mesolithic (period between 8500 and 6500 BC) marked some extremely important changes for both humans and their environment, particularly regarding the animals and plants they directly interacted with. Among the many species involved in this transition process, the wild boar proved to be a key element representing an important prey for the human hunters, in vast regions of Eurasia. The domestication of the wild boar was an element of cultural development of human society in the whole world and played an important role in the human migrations, leading to morphological changes in the phenotype of these animals.
The emergence of domestic pigs in Europe has been tried to be explained through different hypotheses: (1) The migration of farmers from the Near East; (2) The cultural diffusion of ideas through exchange networks; (3) The independent development of agriculture with the appearance of different domestication centres for animals such as pigs or cattle [1]. Up to the middle of the XXth century, due to the discovery of the earliest proofs of agriculture in the Near East, the widest spread hypothesis was that the wild boar was domesticated in the Near East and brought to Europe by the migrant farmers. Later evidence established centres of domestication in South-Eastern Asia [2] or discovered swine with an intermediary or semi-domestic phenotype, spread over a much wider region than the domestication centres previously mentioned [3].
The research carried out up to the middle of the XXth century, based on a basic biometric approach, has a very restrictive character, encountering obstacles that will be eliminated later, with the help of genetics and an advanced geometric morphometric technique, to discover details about diachronic and synchronic specific phenotypic changes related to size, shape and teeth morphology [4]. The picture of the domestication process is a very complex one, and it gets clearer only by corroborating various parameters, including the gene flow between wild and domestic individuals.
The first genetic analysis was carried out on mitochondrial DNA from modern domestic pigs and wild boar in the entire world, describing a common genetic signature for both wild and domestic pigs and supporting the hypothesis of the presence of different domestication centres in South-Eastern Asia and Europe [5,6,7].
Later, with the design of special approaches for the molecular analysis of ancient samples, a certain pattern of synchronic and diachronic haplotypes variation was revealed, and this supported the hypothesis according to which the wild boar was domesticated in the Near East and introduced in Europe during the Neolithic period [8,9].
The analysis of domestic pig presence in Europe leads to a wide collection of samples from the entire continent. Romanian territory represented a centre of interest due to its strategic geographic position which favoured the spread of domestic pigs from the Near East to Europe. On the other hand, the cultural context of that period, proved by the archaeologic research, also could have favoured the introduction of domestic pigs into Europe. Between 5000 and 3500 BC in the North-East of Romanian territory, two flourishing cultures are mentioned: Precucuteni and Cucuteni. Their presence motivated the scientists to analyse the emergence and development of farming in this specific area.
The matter of pig domestication on Romanian territory is mentioned for the first time by Alexandra Bolomey, in 1973 [10], in her study called “The present stage of knowledge of mammal exploitation during the Epipalaeolithic and the earliest Neolithic on the territory of Romania”. The corroboration of both morphometric and genetic data obtained [4] showed that only the pig samples from Iron Gates had the same genetic signature as the European wild boars, while the samples from the other locations possessed a genetic signature that was also found in the Neolithic domestic pigs from Near East.
Further details about the domestic pig emergence on Romanian territory from the Near East are presented in detailed archaeogenetic studies, in 2007 and 2019 [8,11]. These studies presented two routes of domestic pig introduction: a northern one, on the Danube and Rhine course up to northern Europe, and a southern one, that crosses the northern Mediterranean shore. Furthermore, considering the idea of the development of local domestication centres in Europe, the study, like the ones which follow, shows that even if a domestication process existed in Europe, too, as on Romanian territory, this process was not an independent phenomenon, but more likely a consequence of the previous introduction of domestic animals from the Near East.
This idea is also debated in another study based on samples from Romanian territory, published in 2015 [4]. This study presents a comprehensive and detailed image of the timing of the arrival of the first domestic pigs in Europe, and the complexity of the domestication process in South-Eastern Europe, more likely as the admixture process between the wild boar and domestic pigs, which took place after the introduction of domestic pigs from the west and central Anatolia.
In support of the results presented in 2015, the present study aims to dig a little bit more into the synchronic and diachronic evolution of domestic pig genetic signature, gathering data from more samples, more archaeologic sites over the entire Romanian territory also covering a large period of time: from the Neolithic, through the Chalcolithic, the Bronze Age, the Iron Age, and continuing with the Roman period and the Middle Ages.
2. Materials and Methods
A vast inventory of 206 swine bone and teeth samples were collected from a total of 45 archaeological sites on Romanian territory, ranging from the Neolithic period until the Middle Ages (20 Neolithic sites and 25 sites dated between Iron Age and Middle Ages) the samples from the Neolithic, were also selected according to the cultural context. The samples were first subjected to biometrical analysis and diagnosticated as belonging to wild or domestic individuals (Table S1).
From the South-East of the country the samples, gathered from archaeological sites dated between 6000 and 3500 BC, belong to different cultures: the oldest ones belong to the Starčevo-Criș culture (6000–5700 BC)—samples from Coțatcu and Bolintineanu culture (5000–4500 BC)—samples from Gălățui; then, on the time scale, follow the Boian-Giulești culture (4700–4500 BC)—samples from Isaccea Suhat and Siliștea Conac; Gumelnița culture (4500–3900 BC)—samples from Bucșani La Pod, Chitila, Șeinoiu, Sultana Malu Roșu, Panduru and Năvodari Tașaul. Some of the oldest samples included in this study were also collected from the Western and Eastern Romanian territory; the western archaeological sites belonged to the Starčevo-Criș culture (6000–5700 BC): the samples from Ocna Sibiului and Șoimuș or Zau culture (5000–4500 BC): the samples from Cluj Napoca site. The samples collected from the Eastern Romanian territory belonged generally to the late Eneolithic Cucuteni culture, except the ones from the Ghigoești site, which belonged to the Precucuteni culture (5000–4700 BC). These are the samples from six more Eneolithic sites included in this study: Cucuteni-Cetățuie, Târgu-Frumos, Poduri, Trușești Tăcuta and Traian.
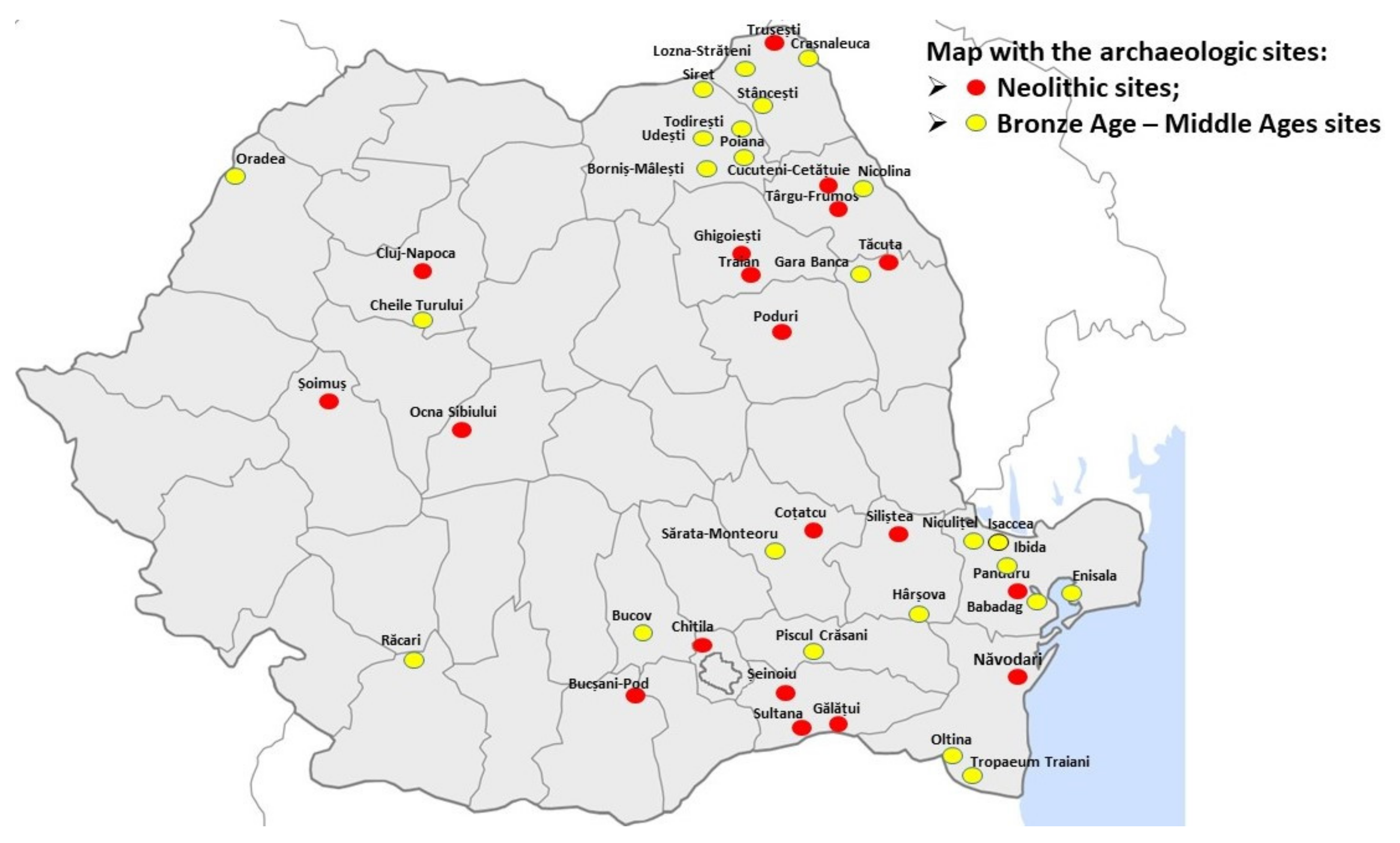
The second wide range of time analyzed in this study spans between the Bronze Age and the Middle Ages and gathers samples from archaeological sites covering again almost the entire Romanian territory, with a higher frequency in the North-East and South-East Romania. The Early Bronze Age (3500–2500 BC) is represented here by samples from Cheile Turului, Western Romania and Sărata-Monteoru, South-Eastern Romania. A later period of the Bronze Age (XIV-XII BC) continues with the Noua culture, represented here by samples from Crasnaleuca in North-East Romania. The Iron Age (XI BC-I AD) is represented by four archaeological sites in South-Eastern Romania: Babadag, Piscul Crăsani, Enisala and Niculițel-Cornet and one in the North-East: Stâncești. The time scale continues with the Roman period (Early and Late Roman period: II-VI AD), represented by samples from Ibida and Niculițel. Other samples from the approximately same period of time were collected from the North-Eastern part of Romanian territory: Lozna Străteni, Udești, Todirești, Gara-Banca, Nicolina, Poiana (Figure 1).
The latest ancient samples included in this study date from the Medieval Ages and their archaeological sites cover the entire Romanian territory: Oradea (North-West), Răcari (South-West), Bucov (South), Oltina, Tropaeum Traiani, Hârșova (South-East), Siret and Borniș-Mâlești (North-East) (Figure 1).
For the Neolithic period, a total of 119 samples were collected, out of which 51% represented bone samples and 49% represented teeth samples.
For the period following the Neolithic, between the Early Bronze Age and Middle Ages, DNA analyses were carried out on a total of 87 samples, out of which the majority (84%) were teeth samples. Bone and teeth samples are chosen for the ancient DNA analysis because they represent a material without any liquids and fewer enzymes, therefore the autolysis is less damaging. They are also less affected by natural contamination with microorganisms and fungi, and the modern DNA contamination can be removed before the DNA extraction process [12]. The teeth samples were mandibles, maxillas or just isolated teeth and the bone samples were fragments coming from the frontal bone, parietal bone, scapula, humerus, cubitus, metacarpal bone, phalange, coxae, tibia, calcaneus or femur.
All procedures starting with the sampling of the bone powder used for DNA extraction until the PCR setup was carried out in laboratories (Archaeo-DNA lab Durham University, UK, and Archaeogenetics Laboratory from Alexandru Ioan Cuza University of Iasi, Romania) dedicated exclusively to working with ancient DNA, to avoid the contamination with other PCR products. The working space was first UV irradiated, then washed with chloride, as well as the equipment. Suitable clothes and shoes were used during all procedures.
Unlike the modern DNA, where the tissue is handled immediately after its sampling, the ancient DNA samples needed a preceding phase of decontamination. All samples were submitted to drilling, to remove the superficial layer which interacted with different contamination agents. A small bone fragment was cut, transformed into powder, incubated in a 15 mL Eppendorf tube with 20 μL proteinase K and 2 mL of extraction buffer and agitated overnight.
On the second day, the actual DNA extraction was carried out using the Amicon Ultra 30K MWCO tubes and the protocol offered by the QIAquick PCR purification kit, from Qiagen. Thus, 2 mL of the sedimented samples were transferred into the Amicon Ultra tubes and centrifuged. After being processed with the help of the Qiaquick kit the extracts reached a final volume of 100 μL of DNA. Then, the extracted DNA concentration and the blank purity were verified spectrophotometrically.
Once the DNA was obtained the next step was its amplification through the PCR reaction. The ancient DNA is always very fragmented because of its high degradation. This includes base modifications or their loss, but also breaks within each of the two DNA strands. Besides the molecular modifications, there are also external factors that can interfere with the PCR reaction: the environmental conditions of the sites where the samples were collected, or the catabolism products of the samples [13]. The DNA extract also contains PCR inhibitors: products of the thymine or cytosine oxidation or collagen catabolic products. To increase the quality of the PCR product, certain concentrations of magnesium chloride and bovine albumin were used. Furthermore, a blank (PCR negative control) and a PCR positive control were included in every batch of samples in order to verify their contamination. The purpose was to amplify an approximately 120 base pairs fragment from the D-loop region of the mitochondrial DNA, firstly designed and tested by Larson and collaborators [8], using the ANC1 pair of primers: ANC1-F: CTTTAAAACAAAAAAACCCATAAAAA and ANC1-R: TTAATGCACGACGTACATAGG.
Due to the ancient DNA’s high fragmentation, the PCR protocol was designed in order to have an amplification capacity inversely proportional to the length of the amplified DNA fragment: a small fragment of proximately 120 base pairs from the D-loop region of the mitochondrial DNA.
Based on phylogeographical analysis carried out with both modern and museum samples, Larson and collaborators [8] designed and tested this fragment for its capacity for discrimination through certain SNPs between several haplotypes described for the European and Near-Eastern Sus scrofa. For all of the analyzed samples, the PCR amplicons were further processed for sequencing. The samples worked in Durham, UK, were sequenced at the Department of Biosciences, by a Sanger sequencing protocol, using the Applied Biosystems 3730 capillary system. The samples worked in Romania, at the Alexandru Ioan Cuza University from Iasi, were also sequenced by the Sanger method, using the Beckman Coulter CEQ8000 capillary system. All resulted sequences were visualized and edited using the Geneious software.
The data obtained from the samples genetically analysed within this study was corroborated with genetic data obtained from other previous studies [4,8,11] which included or were based on samples also collected from Romanian territory. According to these studies the Neolithic Romanian samples had already been identified as carrying three different genetic signatures from the ones described in this study too: the ANC-Aside, ANC-Cside and ANC-Y1-6A haplotypes. Further morphometric and biometric analysis described the analyzed samples as wild boar or domestic pig [4,8,11].
A resulting total dataset of approximately 450 DNA sequences was further subjected to statistical analysis in order to calculate the frequency of each identified haplotype in wild boar or domestic pig.
3. Results and Discussion
3.1. Characterization of Genetic Signature for the Neolithic Samples Collected from Archaeologic Sites in Romanian Territory
The genetic analysis for the Neolithic samples revealed four different haplotypes belonging to the two previously described haplogroups, based on the analysis of the ANC1 diagnosis fragment: the European haplogroup and the Near-Eastern haplogroup.
These four haplotypes, named ANC-Aside and ANC-Cside and ANC-Y1-6A and ANC-Y2-5A differ through 5 mutations: three transitions between the ANC-Aside haplotype and the ANC-Y1-6A and ANC-Y2-5A haplotypes, and two other particular mutations: one transversion for the European ANC-Aside haplotype and one deletion for the Near-Eastern ANC-Y2-5A haplotype (Figure 2). When first described, the first two haplotypes, ANC-Aside and ANC-Cside were assigned to the European haplogroup, while the latter two, the ANC-Y1-6A and ANC-Y2-5A were assigned to the Near-Eastern haplogroup [8]. Later studies [11] showed that the ANC-Y2-5A haplotype actually has a European origin. This questioned haplotype has a very low frequency both on Romanian territory and the rest of Europe and the meaning of its spatial–temporal presence in both domestic and wild individuals is yet to be debated.
Reported to the entire number of Neolithic samples (with positive, diagnosticated results) the haplotype with the highest frequency (56.43%—57 samples) was the Near-Eastern ANC-Y1-6A, which prevailed in domestic pigs, and it was followed by the European ANC-Cside haplotype (32.67%—33 samples), found generally in wild boar. Only a few individuals (11), both domestic and wild, presented the European ANC-Aside haplotype (6) and the Near-Eastern ANC-Y2-5A haplotype (5).
3.2. Timing of the Near-Eastern ANC-Y1-6A Haplotype Spread on Romanian Territory during the Neolithic
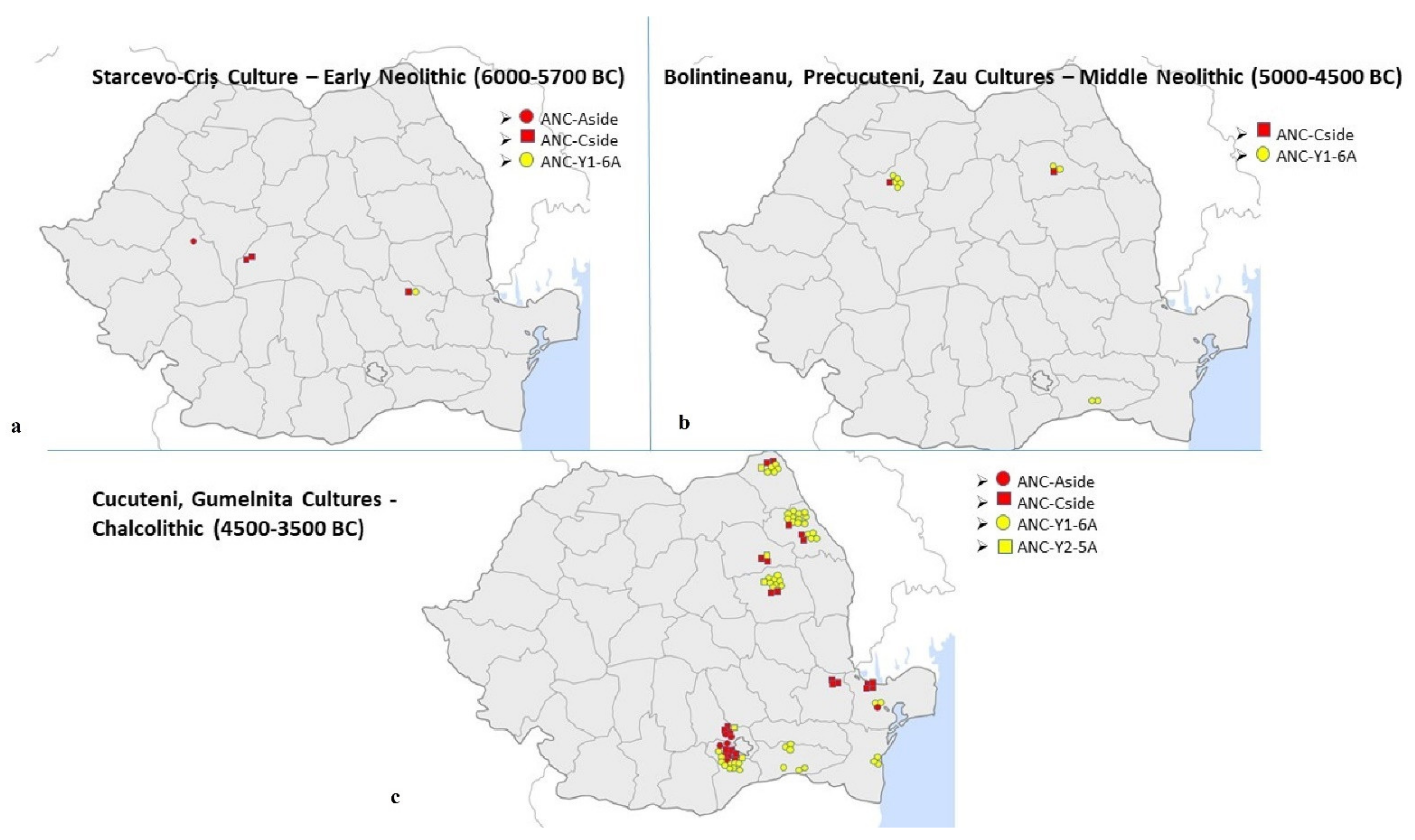
A total of 119 Neolithic samples were analysed, out of which 101 held positive results (84.9%). The earliest samples subjected to DNA analysis within this study date from the Starčevo-Criș culture and were collected from Ocna Sibiului, Șoimuș and Coțatcu, from the Western and Eastern regions of Romanian territory. The genetic signature obtained for these samples shows that at this time (6000–5700 BC) in the Western Romanian territory the European haplotype was still prevailing since only the European ANC-Cside and ANC-Aside haplotypes were described for the two wild and one domestic Sus scrofa, respectively. Only in the Eastern site (Coțatcu) one Near-Eastern ANC-Y1-6A haplotype was described, which supports previous data, that by 6000–5700 BC the spread of the Near-Eastern farming into Europe had already begun [4] (Figure 3a).
During the next analysed period, 5000–4500 BC, with the Bolintineanu culture represented by samples from Gălățui, Boian Culture with samples from Siliștea Conac and Isaccea Suhat (both cultures from South-Eastern Romania), Precucuteni culture in North-Eastern Romania represented by samples from Ghigoești and Zau culture in Western Romania represented by samples from Cluj Napoca, the frequency of the ANC-Y1-6A haplotype had already increased considerably (Figure 3b), although the European ANC-Cside haplotype is still present in a high degree. Biometric analysis assigned the ANC-Y1-6A haplotype for the Romanian samples of this period of time only to domestic pigs.
During the latest Neolithic period included in this study, the Chalcolithic, with the Cucuteni, and Gumelnița cultures (4500–3500 BC) the Near-Eastern ANC-Y1-6A haplotype’s frequency reaches the highest level in the majority of the archaeological sites dating from this period (in the Eastern Romanian territory) (Figure 3c). Corroboration of biometric and genetic data reveals that, at this time, this Near-Eastern genetic signature starts to appear not only in the domestic pig, but also in the wild boar. This supports the previously presented data, according to which the admixture process and gene flow had an important role in the emergence of domestic pigs on Romanian territory [4].
3.3. Timing of the Near-Eastern ANC-Y1-6A Haplotype Spread on Romanian Territory during the Period between Early Bronze Age and Middle Ages
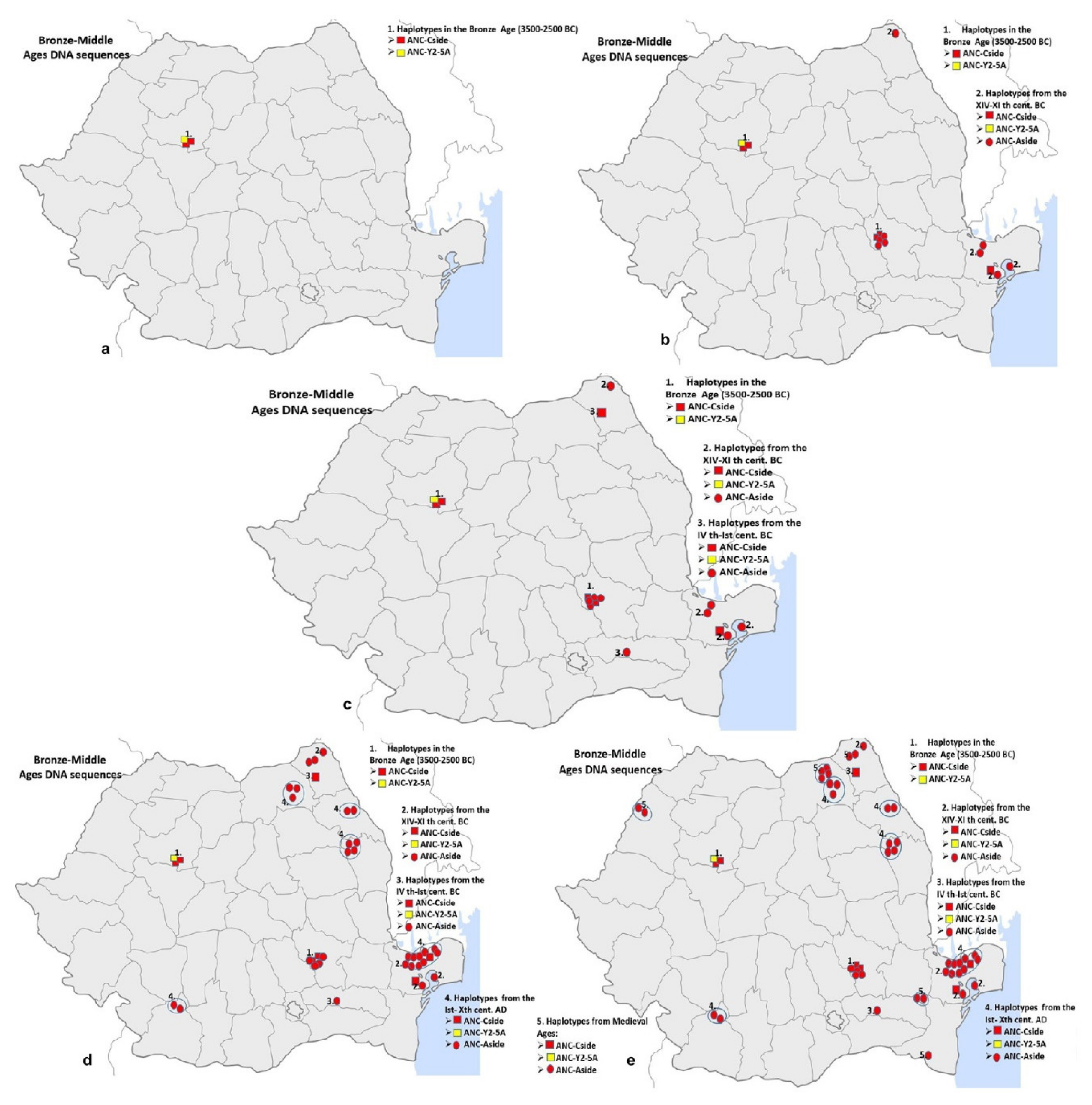
From the 87 samples ranging between the Early Bronze Age and the Middle Ages only 46 held positive results (52.9%) after the PCR reaction. One single Near-Eastern haplotype was identified for one of the earliest samples from this period, coming from Western Romania, Cheile Turului (Early Bronze Age: 3500–2500 BC): the ANC-Y2-5A haplotype (Figure 4a). Six samples, one from Moldova (Stâncești), three from the East and South-East Romania (Sărata Monteoru and Babadag) and two others from Transilvania (Cheile Turului), were morphologically identified as belonging to the wild boar, presented the European ANC-Cside haplotype (Figure 4c). The Near-Eastern ANC-Y1-6A haplotype seemed to disappear in this second period analysed (Early Bronze Age–Middle Ages). The rest of the samples which were successfully PCR amplified encoded only the European ANC-Aside haplotype, all of them belonging to the domestic pig.
A better image of the relevance of this haplotype variation is given by the correlation between the dating of the samples and their genetic signature. Thus, starting with the Early Bronze Age the overall picture of pig’s genetic signature on Romanian territory changed, acquiring a prevalent European genetic signature: the Near-Eastern ANC-Y1-6a haplotype is now replaced by the European ANC-Aside haplotype and together with this one, but to a lesser extent, the presence of the European ANC-Cside haplotype is maintained (Figure 4).
A previous study [11] has already demonstrated that the introduction of domestic pigs in Europe was followed by their genomic turnover and the timing of this process has also been analysed. Yet, the samples from Romanian sites analysed so far [4,8,11] did not exceed the time limit of the Iron Age. The present study extends the time boundary for the samples analysed from Romanian territory up to the Middle Ages. Thus, corroborating the data obtained within this study with the previous genetic data, we obtained a more comprehensive picture of the haplotype’s succession on Romanian territory, a strategic point for the spread of domestic pig into Europe.
More than that, to investigate how the genetic changes at this gate of European Neolithization correlated with the ones registered for the entire European continent, a larger set of samples, including samples from other European countries, was analysed. The genomic turnover strongly correlates with two big issues in the pig domestication process: the analysis of the European ANC-Aside haplotype’s frequency as well as the synchronic and diachronic analysis of the Near-Eastern ANC-Y1-6A haplotype’s spread.
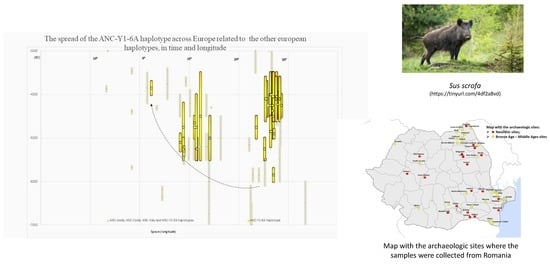
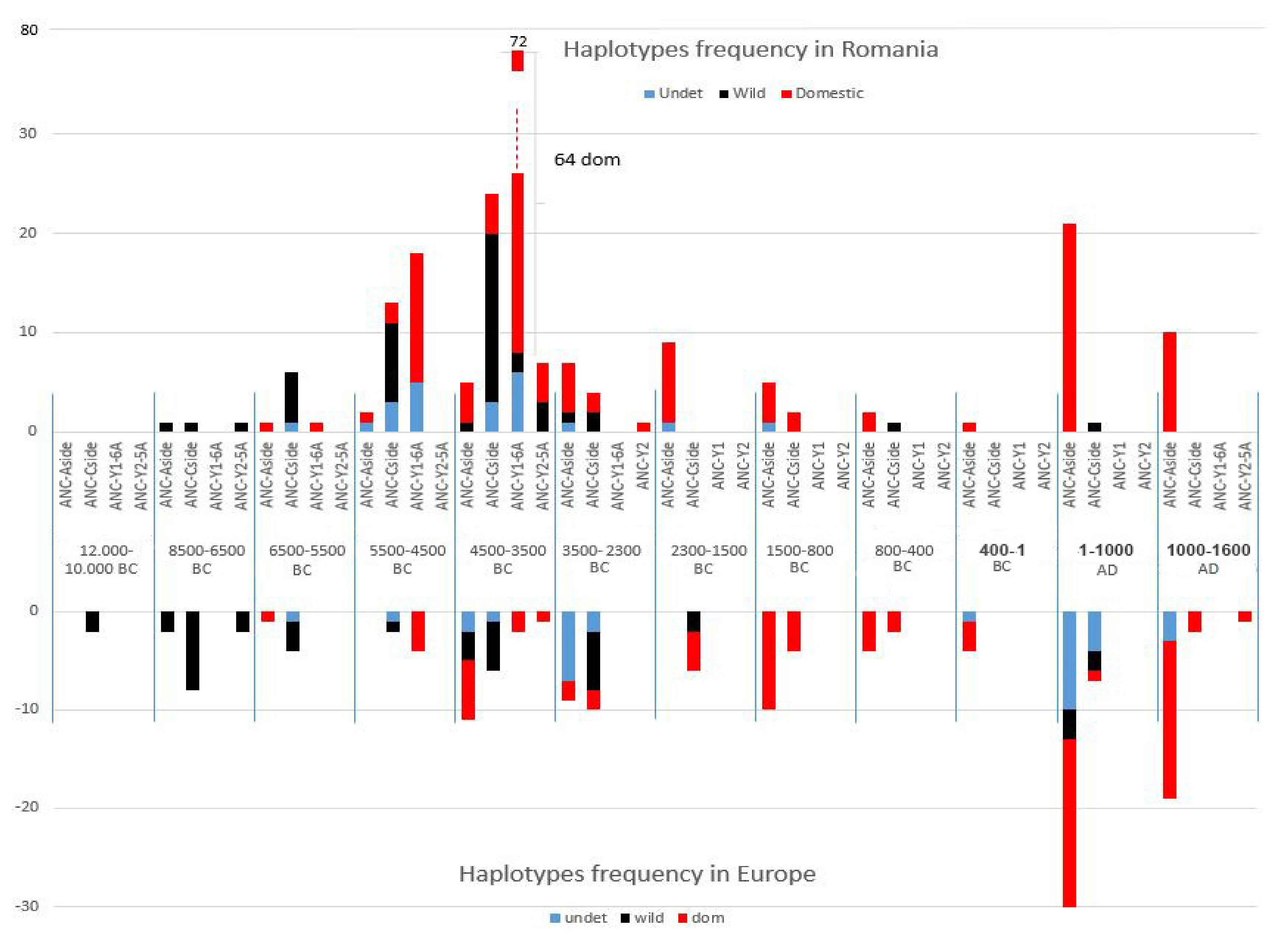
The comparison between the genetic signature turnover on Romanian territory and on the entire European territory [4,8,14] revealed that the pattern of the Near-Eastern haplotype ANC-Y1-6A spread on Romanian territory is maintained for the entire continent, with the same timing of its first appearance, prevalence, and extinction (Figure 5).
The domestic pigs ANC-Aside haplotype appeared both on Romanian territory and the rest of Europe in the Early Neolithic (6500–5500 BC) and will be replaced later by the Near-Eastern haplotype. It maintained a very low frequency during the next 1000 years (5500–4500) on Romanian territory (while missing in the rest of Europe), and its frequency increased again in domestic pigs during the Chalcolithic period (4500–3300) both on Romanian and the rest of the European territory. Still, in the same period of time, this European haplotype’s frequency in domestic pigs is much lower than the Near-Eastern one. The lack of the European ANC-Aside haplotype’s presence in Europe between 5500 and 4500 and its different frequency varies, of course, with the different number of samples collected and successfully amplified from site to site.
Yet, not the exact number, but the prevalence of a certain domestic signature in wild or domestic, as well as its emergence and disappearance on a larger temporal scale described a certain pattern of evolution for the Near-Eastern domestic pigs spread in Europe.
On Romanian territory, for domestic pigs, the earliest European ANC-Aside haplotype identification synchronizes with the earliest ANC-Y1 Aside identification. The same European genetic signature appeared even earlier (8500–6500 BC) both on Romanian territory and the rest of Europe, but only in the wild boar. For the next 1000 years, the corresponding phenotype of the European ANC-Aside haplotype changed from the wild to domestic, at a very low frequency between 6500 and 5500 BC, when the frequency of the introduced Near-Eastern haplotype was also very low. This happened both on Romanian territory and the rest of Europe and could be the first proof of the wild boar introgression into the introduced domestic stocks, a hypothesis supported by previous data [11], yet questioned by the lack of the wild boar with the ANC-Aside genetic signature within the same spatial–temporal boundary. Between 4500 and 3500 BC, the ANC-Aside’s frequency increases even more in the period when the Near-Eastern genetic signature reaches its peak, but follows an inverse correlation within the next thousands of years, until 1600 AD.
On the other hand, there is a presence of the European ANC-Cside haplotype only in the wild boar of the Early Neolithic both on Romanian territory and Europe. Later on (5500–4500), after the emergence of the Near-Eastern haplotype in the same geographical boundaries, the European ANC-Cside haplotype is identified both for the wild boar and domestic pig with low frequencies for domestics and high frequencies for the wild individuals, revealing a clear introgression of the wild individuals into the domestic stocks.
Still, considering the previous morphometric results corroborated with genetic data, neither the geographic timing of the European and Near-Eastern haplotypes in domestic pigs nor the introgression process of the wild boars into the domestic stocks do not particularly support the existence of independent domestication centres in Romanian or European territory [4].
The complexity of the domestication process is far deeper and certain traits of it are underlined for example with the gene flow, which is more pronounced within the later periods, covering the Neolithic, Chalcolithic and less the Bronze Age, when interchanges in genotypes characteristic for certain phenotypes can be noted.
Instead, as the time boundaries of the analysed samples widen, they reveal a clearer image of the genomic turnover for the domestic pig both in Romanian territory and Europe: it took approximately 2000 years for the introduced Near-Eastern domestic pig to reach its frequency peak and another 1000 years for its genome to be replaced by the European one, and this European signature maintained later for further thousands of years.
4. Conclusions
With the big number of archaeological sites (41 with morpho-genetic diagnosticated samples out of 45 sites) and samples (147 morpho-genetic diagnosticated samples out of the 206) and the wide period of time covered (6000 BC–1600 AD), the present study reveals a relevant diachronic and synchronic image that supports the previously presented results regarding the first introduction of the domestic pig on Romanian territory and Europe.
The total replacement of the Near-Eastern genetic signature with the European one after the Bronze Age is another feature of the domestication complexity which is yet to be explained.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d14040288/s1, Table S1: The samples from Romanian territory used in the study.
Author Contributions
Conceptualization, M.L., A.B. and M.S.S.; methodology, M.L. and A.B.; software, M.L.; validation, M.L., A.B. and M.S.S.; formal analysis, M.L.; investigation, M.L.; resources, M.L.; data curation, M.L.; writing—original draft preparation, M.L.; writing—review and editing, A.B. and M.S.S.; visualization, M.S.S. and L.B.; supervision, A.B. and L.B.; project administration, M.S.S.; funding acquisition, M.S.S. All authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank R. Andreescu, C. Bem, C. Lazăr, C. Nicolae, V. Voinea, S. Pandrea, C. Micu, and D. Bindea for providing material and support. We thank G. Larson for the support in the faunal material analysis at Archaeo-DNA lab, Durham University, UK.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Larson, G.; Liu, R.; Zhao, X.; Yuan, J.; Fuller, D.; Barton, L.; Dobney, K.; Fan, Q.; Gu, Z.; Liu, X.H.; et al. Patterns of East Asian pig domestication, migration, and turnover revealed by modern and ancient DNA. Proc. Natl. Acad. Sci. USA 2010, 107, 7686–7691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albarella, U.; Dobney, K.; Ervynck, A.; Rowley-Conwy, P. Pigs and Humans: 10000 Years of Interaction; Oxford University Press: Oxford, UK, 2007; pp. 1–13. [Google Scholar]
- Ervynck, A.; Dobney, K.; Hongo, H.; Meadow, R. New evidence for the status of Sus scrofa at Neolithic Çayönü Tepesi (Southeastern Anatolia, Turkey). Paléorient 2001, 27, 47–73. [Google Scholar] [CrossRef]
- Evin, A.; Girdland Flink, L.; Bălășescu, A.; Popovici, D.; Andreescu, R.; Bailey, D.; Mirea, P.; Lazăr, C.; Boroneanț, A.; Bonsall, C.; et al. Unravelling the complexity of domestication: A case study using morphometrics and ancient DNA analyses of archaeological pigs from Romania. Phil. Trans. R. Soc. B 2015, 370, 20130616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuffra, J.; Kijas, J.M.H.; Amarger, V.; Carlborg, O.; Jeon, J.T.; Andersson, L. The Origin of the Domestic Pig: Independent Domestication and Subsequent Introgression. Genet. Soc. Am. 2000, 154, 1785–1791. [Google Scholar] [CrossRef] [PubMed]
- Larson, G.; Dobney, K.; Albarella, U.; Fang, M.; Matisoo-Smith, E.; Robins, J.; Lowden, S.; Finlayson, H.; Brand, T.; Willerslev, E.; et al. Worldwide phylogeography of wild boar reveals multiple centers of pig domestication. Science 2005, 307, 1618–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, M.; Andersson, L. Mitochondrial diversity in European and Chinese pigs is consistent with population expansions that occurred prior to domestication. Proc. R. Soc. B 2006, 273, 1803–1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, G.; Albarella, U.; Dobney, K.; Rowley-Conwey, P.; Schibler, J.; Tresset, A.; Vigne, J.-D.; Edwardsh, C.; Schlumbaum, A.; Dinu, A.; et al. Ancient DNA, pig domestication, and the spread of the Neolithic into Europe. Proc. Natl. Acad. Sci. USA 2007, 104, 15276–15281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottoni, C.; Flink, L.G.; Evin, A.; Georg, C.; De Cupere, B.; Van Neer, W.; Bartosiewicz, L.; Linderholm, L.; Barnett, R.; Peters, J.; et al. Pig Domestication and Human-Mediated Dispersal in Western Eurasia Revealed through Ancient DNA and Geometric Morphometrics. Mol. Biol. Evol. 2013, 30, 824–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolomey, A. The present stage of knowledge of mammal exploitation during the Epipalaeolithic and the earliest Neolithic on the territory of Romania. In Domestikationforschung und Geschichte der Haustiere; Matolcsi, J., Ed.; Akademiai Kiado: Budapest, Hungary, 1973; pp. 197–203. [Google Scholar]
- Frantz, L.A.F.; Haile, J.; Lin, A.T.; Scheu, A.; Geörg, C.; Benecke, N.; Alexander, M.; Linderholm, A.; Mullin, V.E.; Daly, K.G.; et al. Ancient pigs reveal a near-complete genomic turnover following their introduction to Europe. Proc. Natl. Acad. Sci. USA 2019, 116, 17231–17238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baubliene, J.; Daugnora, L.; Miceikiene, I. Evaluation of the DNA extraction method from ancient animal bones. Ekologija 2003, 1, 8–11. [Google Scholar]
- Chelomina, G.N. Ancient DNA. Russ. J. Genet. 2006, 42, 219–233. [Google Scholar] [CrossRef]
- Krause-Kyora, B.; Makarewicz, C.; Evin, A.; Flink, L.G.; Dobney, K.; Larson, G.; Hartz, S.; Schreiber, S.; von Carnap-Bornheim, C.; von Wurmb-Schwark, N.; et al. Use of domesticated pigs by Mesolithichunter-gatherers in northwestern Europe. Nat. Commun. 2013, 4, 2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Map with the archaeologic sites where the samples were collected from Romania.

Figure 2.
Differences between the four haplotypes described for Romanian samples.

Figure 3.
The spread of the Near-Eastern Sus scrofa genetic signature on Romanian territory during the Neolithic: (a). Haplotypes’ spread in the Early Neolithic; (b). Haplotypes’spread in the Middle Neolithic; (c). Haplotypes’ spread in the Chalcolithic.
Figure 3.
The spread of the Near-Eastern Sus scrofa genetic signature on Romanian territory during the Neolithic: (a). Haplotypes’ spread in the Early Neolithic; (b). Haplotypes’spread in the Middle Neolithic; (c). Haplotypes’ spread in the Chalcolithic.

Figure 4.
The prevalence of the European ANC-Aside haplotype on Romanian territory for the pig samples dating between the Early Bronze Age and Middle Ages: (a) Haplotypes’ spread between 3500–2500 BC; (b) Haplotypes’ spread between 3500-XIth century BC; (c) Haplotypes’ spread between 3500-Ist century BC; (d) Haplotypes’ spread between 3500 BC-Xth century AD; (e) Haplotypes’ spread between 3500 BC-Medieval Ages.
Figure 4.
The prevalence of the European ANC-Aside haplotype on Romanian territory for the pig samples dating between the Early Bronze Age and Middle Ages: (a) Haplotypes’ spread between 3500–2500 BC; (b) Haplotypes’ spread between 3500-XIth century BC; (c) Haplotypes’ spread between 3500-Ist century BC; (d) Haplotypes’ spread between 3500 BC-Xth century AD; (e) Haplotypes’ spread between 3500 BC-Medieval Ages.

Figure 5.
Timing of haplotypes frequency for Sus scrofa on Romanian territory versus Europe.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Stanc, M.S.; Luca, M.; Bălășescu, A.; Bejenaru, L. Holocene Genetic Evolution of Pig (Sus scrofa) on Romanian Territory in a European Time and Space Frame. Diversity 2022, 14, 288. https://doi.org/10.3390/d14040288
AMA Style
Stanc MS, Luca M, Bălășescu A, Bejenaru L. Holocene Genetic Evolution of Pig (Sus scrofa) on Romanian Territory in a European Time and Space Frame. Diversity. 2022; 14(4):288. https://doi.org/10.3390/d14040288
Chicago/Turabian StyleStanc, Margareta Simina, Monica Luca, Adrian Bălășescu, and Luminița Bejenaru. 2022. "Holocene Genetic Evolution of Pig (Sus scrofa) on Romanian Territory in a European Time and Space Frame" Diversity 14, no. 4: 288. https://doi.org/10.3390/d14040288
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.